Deep-Sequence Identification and Role
in Virus Replication of a JC Virus
Quasispecies in Patients with Progressive
Multifocal Leukoencephalopathy
Kenta Takahashi,
aTsuyoshi Sekizuka,
bHitomi Fukumoto,
aKazuo Nakamichi,
cTadaki Suzuki,
aYuko Sato,
aHideki Hasegawa,
aMakoto Kuroda,
bHarutaka Katano
aDepartment of Pathology,aPathogen Genomics Center,band Department of Virology 1,cNational Institute of
Infectious Diseases, Tokyo, Japan
ABSTRACT
JC virus (JCV) is a DNA virus causing progressive multifocal
leukoencepha-lopathy (PML) in immunodeficient patients. In the present study, 22 genetic quasispecies
with more than 1.5% variant frequency were detected in JCV genomes from six clinical
samples of PML by next-generation sequencing. A mutation from A to C at nucleotide
(nt) 3495 in JCV Mad1 resulting in a V-to-G amino acid substitution at amino acid (aa)
position 392 of the large T antigen (TAg) was identified in all six cases of PML at 3% to
19% variant frequencies. Transfection of JCV Mad1 DNA possessing the V392G
substitu-tion in TAg into IMR-32 and human embryonic kidney 293 (HEK293) cells resulted in
dramatically decreased production of JCV-encoded proteins. The virus DNA copy
num-ber was also reduced in supernatants of the mutant virus-transfected cells. Transfection
of the IMR-32 and HEK293 cells with a virus genome containing a revertant mutation
re-covered viral production and protein expression. Cotransfection with equal amounts of
wild-type genome and mutated JCV genome did not reduce the expression of viral
pro-teins or viral replication, suggesting that the mutation did not have any
dominant-negative function. Finally, immunohistochemistry demonstrated that TAg was expressed
in all six pathological samples in which the quasispecies were detected. In conclusion,
the V392G amino acid substitution in TAg identified frequently in PML lesions has a
function in suppressing JCV replication, but the frequency of the mutation was restricted
and its role in PML lesions was limited.
IMPORTANCE
DNA viruses generally have lower mutation frequency than RNA viruses,
and the detection of quasispecies in JCV has rarely been reported. In the present study,
a next-generation sequencer identified a JCV quasispecies with an amino acid
substitu-tion in the T antigen in patients with PML.
In vitro
studies showed that the mutation
strongly repressed the expression of JC viral proteins and reduced the viral replication.
However, because the frequency of the mutation was low in each case, the total
expres-sion of virus proteins was sustained
in vivo
. Thus, JC virus replicates in PML lesions in
the presence of a mutant virus which is able to repress virus replication.
KEYWORDS
JCV, PML, large T antigen, quasispecies, next-generation sequencer,
virus replication, progressive multifocal leukoencephalopathy
T
he JC virus (JCV) is a DNA virus of the human polyomavirus family (1–3). Serum
antibodies to JCV are found in 51% to 76% of a healthy general population,
indicating that JCV is ubiquitous in humans (4–7). The primary infection of JCV occurs
asymptomatically during infancy, with latent infections occurring in the kidney,
lym-phocyte, spleen, and bone marrow. JCV spreads hematogenously to the central nervous
system, or remains latent in the brain during primary viremia, and reactivates locally,
Received6 July 2016Accepted3 October 2016
Accepted manuscript posted online19 October 2016
CitationTakahashi K, Sekizuka T, Fukumoto H, Nakamichi K, Suzuki T, Sato Y, Hasegawa H, Kuroda M, Katano H. 2017. Deep-sequence identification and role in virus replication of a JC virus quasispecies in patients with progressive multifocal leukoencephalopathy. J Virol 91:e01335-16. https://doi.org/10.1128/ JVI.01335-16.
EditorLawrence Banks, International Centre for Genetic Engineering and Biotechnology
Copyright© 2016 American Society for Microbiology. All Rights Reserved. Address correspondence to Harutaka Katano, [email protected].
crossm
on November 7, 2019 by guest
http://jvi.asm.org/
causing progressive multifocal leukoencephalopathy (PML), a fatal demyelinating
dis-ease, in immunocompromised patients such as AIDS sufferers or patients receiving
immunomodulatory therapies (8–10). In the PML lesion, remarkable demyelination is
observed histologically in the white matter, accompanying enlarged oligodendroglial
nuclei and bizarre astrocytes with nuclear atypia (11, 12). JCV is detected in the
enlarged nucleus of oligodendroglia by immunohistochemistry (13, 14).
JCV encodes a large T antigen (TAg), a small t antigen, three virus capsid proteins
(VP1 to -3), and an agnoprotein in its genome (15). JCV-encoded TAg plays an
important role in virus replication that is similar to the role of the TAg encoded by other
polyomaviruses (16, 17). TAg binds to the viral origin of replication through a DNA
binding domain, and the nuclear localization signal in TAg recruits TAg to the nucleus
(17). The helicase domain in TAg unwinds the DNA double helix and promotes viral
DNA replication (18).
DNA viruses, including JCV, generally have fewer mutations than RNA viruses. JCV
isolates from PML patients have unique mutations, deletions, and/or rearrangements
(so called PML-type rearrangements) in the regulatory region; in addition,
⬎
90% of JCV
isolates from PML patients carry point mutations in VP1 (19–23). Rearrangement of the
regulatory region in polyomavirus has been detected frequently in patients with
polyomavirus infection, including PML (10). The JCV genome with the archetype
regulatory region undergoes insertion and deletion errors during replication,
generat-ing rearranged regulatory region genomes. Polyomaviruses with a rearranged
regula-tory region have increased levels of early gene expression and higher replication
capacity, resulting in elimination of polyomaviruses with the archetype regulatory
region from the PML lesion in immunodeficient hosts (21, 24). Mutations in the
agnoprotein have been identified also in PML-variant JCV encephalopathy (25). Thus,
mutation, deletion, and rearrangement are sometimes observed in JCV genomes
isolated from patients with PML; however, the details concerning the frequency of
quasispecies and mutations in the JCV genome in individuals have not been reported
to date. In the present study, next-generation sequencing (NGS) was performed on
pathological samples from PML lesions, which identified common quasispecies in the
JCV genome among PML cases. In addition, the role of TAg in the replication of the
common quasispecies was investigated.
RESULTS
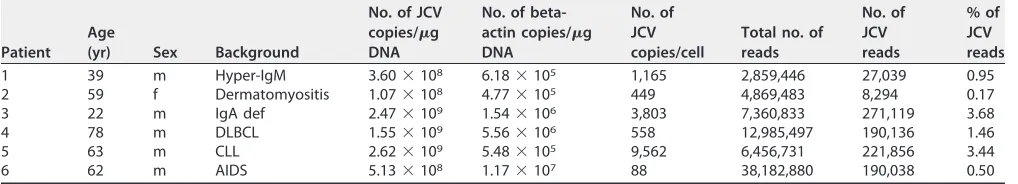
[image:2.585.41.550.84.176.2]JCV detection and full sequence determination in each PML case.
JCV DNA was
quantified in the six PML samples. High numbers of JCV DNA ranging from 88 to 9,562
copies per cell were detected (Table 1). PCR and direct-sequence analysis revealed that
the JCV genomes in all six PML cases had PML-type regulatory regions (Fig. 1A). NGS
was performed on DNA extracted from pathological samples from the six PML cases.
The full-length JCV genomes were constructed from the sequences of the regulatory
region and the NGS contigs in each case. Phylogenetic tree analysis based on
whole-genome sequences demonstrated that all six cases belonged to the B CY or the B MY
cluster of JCV, each of which is a major cluster in Japan (Fig. 1A). NGS reads were
mapped on each full-length genome. Finally, a minimum depth of 100 reads was
TABLE 1List of patients and samplesaPatient Age
(yr) Sex Background
No. of JCV copies/g DNA
No. of beta-actin copies/g DNA
No. of JCV copies/cell
Total no. of reads
No. of JCV reads
% of JCV reads
1 39 m Hyper-IgM 3.60⫻108 6.18⫻105 1,165 2,859,446 27,039 0.95
2 59 f Dermatomyositis 1.07⫻108 4.77⫻105 449 4,869,483 8,294 0.17
3 22 m IgA def 2.47⫻109 1.54⫻106 3,803 7,360,833 271,119 3.68
4 78 m DLBCL 1.55⫻109 5.56⫻106 558 12,985,497 190,136 1.46
5 63 m CLL 2.62⫻109 5.48⫻105 9,562 6,456,731 221,856 3.44
6 62 m AIDS 5.13⫻108 1.17⫻107 88 38,182,880 190,038 0.50
aCLL, chronic lymphocytic leukemia; DLBCL, diffuse large B cell lymphoma; IgA def, IgA deficiency; f, female; m, male.
on November 7, 2019 by guest
http://jvi.asm.org/
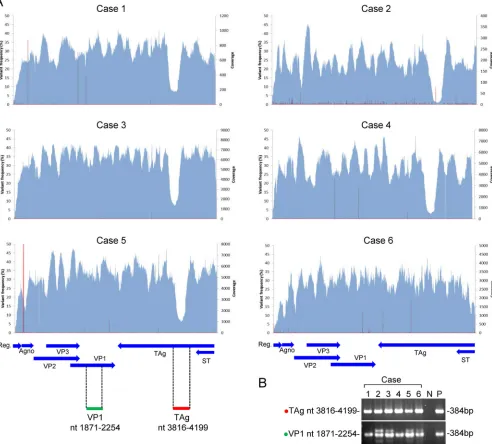
completed in more than 95% of the JCV genome in all samples (Fig. 2A). Since low
levels of coverage were observed at nucleotide (nt) 3830 to 4100 of the JCV genome
in 5 cases, a fragment covering the region was amplified with a standard PCR and
subjected to direct sequencing. The fragment was amplified at an expected size in all
6 cases (Fig. 2B), and no mutation or sequence variation was detected by direct
sequencing.
Identification of JCV quasispecies from PML patients.
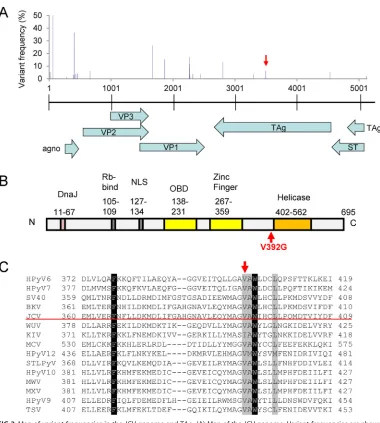
Mapping of the NGS reads
to the full-length JCV genome identified 22 quasispecies which showed more than 1.5%
variant frequency with respect to the JCV genome (Table 2 and Fig. 3A). Among the 22
quasispecies, A
3495to C
3495mutations were found in all six cases. G
1664to C
1664mutations were found in two cases, whereas the other 20 quasispecies were observed
in a single case. The mutations of G
2261to A
2261and C
2274to T
2274result in amino acid
substitutions from asparagine
264to aspartic acid
264and serine
268to cysteine
268in the
JCV oligosaccharide-binding sites of VP1, respectively (23, 26). The variant frequency of
A
3495to C
3495ranged from 3.29% to 19.68% in the six cases. The mutation of A to C at
this position resulted in a single amino acid substitution from valine
392to glycine
392in
TAg immediately upstream of the helicase domain (Fig. 3B). The valine at position 392
FIG 1Typing of the JCV genome in PML patients. (A) Recombination of the regulatory region of JCV in cases 1 to 6. (B) Phylogenetic tree of the whole genome of JCV. The samples of cases 1 to 6 are indicated in red. The genotypes of the JCV samples and the GenBank accession numbers of the reference strains are as follows: AB077870 (B SC), AB362351 (B 2E), AB077855 (B SC), AB048577 (B MY), AB092584 (B MX), AB118654 (B CY), AB113145 (B B3a), AB048557 (B B2), AB372037 (B B1-d), AB372038 (B B1-c), AB262402 (B B1-b), AB113144 (B B1-a), AB127013 (B Af2), AB038253 (C Af1), AB074575 (A EU-c), AB048563 (A EU-b), AB074580 (A EU-a), AB127349 (A EU-a), AB127348 (A EU-a), AB038254 (Tky-1), U61771 (Taiwan-3), and J02226 (Mad1).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.54.541.69.460.2]in the JCV TAg is highly conserved among the other 13 human polyomaviruses except
KI polyomavirus (Fig. 3C).
Repression of JCV replication by the V392G mutation in TAg and its limited
role.
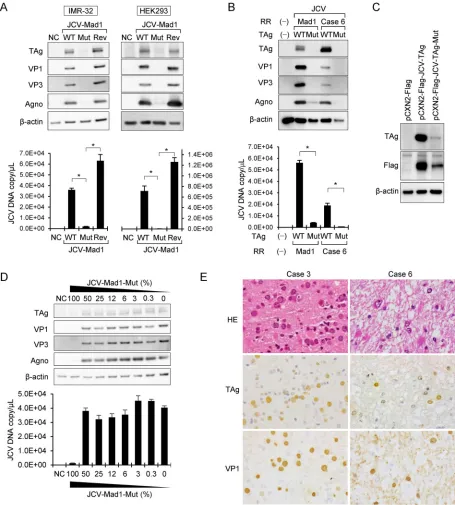
To investigate the biological function of the V392G amino acid substitution in TAg,
the wild-type JCV genome and the JCV genome with the V392G mutation and its
revertant genome were each transfected into human neuroblastoma IMR-32 and
human embryonic kidney 293 (HEK293) cells, and cell lysates were collected 4 days
later. Immunoblot analysis revealed that expression of both early and late JCV proteins
was strongly repressed in the cells transfected with the mutant JCV genome. The
expression of these proteins completely recovered in cells transfected with the
rever-tant JCV genome (Fig. 4A). Real-time PCR analysis revealed that the viral copy number
FIG 2Coverage and variant frequencies in the JCV genome determined by NGS. (A) Coverage and variant frequencies. Coverages are shown in light blue, and variant frequencies are indicated by red. The transcripts encoded by JCV are shown by blue arrows at the bottom of the panels. (B) PCR amplification of the region with low coverage. A DNA fragment at JCV nt 3816 to 4199 of TAg was amplified in all six cases and electrophoresed. JCV region nt 1871 to 2254 of VP1 was also amplified as a positive control for JCV DNA. The amplified regions are indicated by green and red bars at bottom left of panel A. Predicted sizes of amplicons are indicated at the right. Agno, agnoprotein gene; Reg., regulatory region; P, positive control (JCV-Mad1 strain plasmid); N, negative control (no DNA).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.48.540.77.521.2]was significantly decreased in the supernatant of IMR-32 and HEK293 cells transfected
with the mutated JCV genome and that the copy numbers recovered in the cells
transfected with revertant genome (Fig. 4A). It has been demonstrated that sequences
of regulatory regions affect expression of early proteins encoded by polyomavirus (21,
24). Therefore, the JCV genome with the PML-type regulatory region from a patient was
also transfected to IMR-32 cells in the presence or absence of the V392G amino acid
substitution. Western blot analysis revealed that expression of both early and late JCV
proteins was strongly repressed in the cells transfected with the V392G-mutated JCV
genome with a regulatory region from case 6 (Fig. 4B). JCV copy numbers decreased in
the culture supernatant of IMR-32 cells transfected with the V392G-mutated JCV
genome with a regulatory region from case 6. These data suggest that the repression
of TAg expression resulting from the V392G amino acid substitution was
indepen-dent of sequences in the regulatory region. To evaluate the effect on TAg
expres-sion of the V392G amino acid substitution, TAg expresexpres-sion plasmids with or without
the V392G mutation were transfected to IMR-32 cells. Western blot analysis using
anti-TAg and anti-Flag antibodies demonstrated that expression of mutant TAg was
much lower than that of wild-type TAg in transfected IMR-32 cells (Fig. 4C),
suggesting that the V392G amino acid substitution directly caused low expression
of TAg. Since NGS detected the mutant in 3% to 19% of reads in each clinical sample
(Table 2), we investigated the effect of cotransfection with mutated and wild-type
JCV genomes in various ratios. Western blot analysis revealed that transfection with
a combination containing up to 50% of the mutated genome did not reduce
expression of any JCV proteins whereas the mutant genome at a level of 100%
repressed their expression (Fig. 4D). In addition, analysis of the viral copy numbers
in the supernatant showed results comparable to those seen with the protein
expression, indicating that contamination by the mutant genome at a level of less
than 50% did not affect total JCV replication (Fig. 4D). Finally, expression of TAg was
confirmed by immunohistochemistry in all the clinical samples of PML in which the
quasispecies was detected (Fig. 4E), suggesting that the mutation of A to C at nt
3495 did not reduce expression of TAg and that the role of the mutation in the
clinical samples was limited.
TABLE 2Quasispecies in JCV genomes isolated from PML lesionsa
Coding
Position in J02226
Reference sequence Variant
Variant frequency (%)
Case 1 Case 2 Case 3 Case 4 Case 5 Case 6
No coding 14 G t 5.06
No coding 61 T acg 52.70
agno 281 T g 1.62
agno 390 C t 3.47
agno 403 G t 2.76
agno 406 S c 36.46
agno 429 A g 3.65
agno 458 G c 4.05
VP2 661 C g 6.19
VP1 1664 G c 29.58 22.79
VP1 1865 G a 15.31
VP1 2261 G a 7.86
VP1 2262 A c 17.31
VP1 2268 A c 11.81
VP1 2274 C t 2.68
VP1 2317 A c 3.73
VP1 2446 T g 3.83
TAg 2804 G c 13.06
TAg 3299 T c 3.73
TAg 3495 A c 3.39 3.76 3.29 3.73 3.46 19.68
TAg/ST 4534 C g 5.06
No coding 5119 C a 2.01
aBold data indicate a common quasispecies among the PML cases. agno, agnoprotein.
on November 7, 2019 by guest
http://jvi.asm.org/
DISCUSSION
In the present study, NGS identified a JCV quasispecies with the amino acid
substitution V392G in TAg in all 6 PML patients examined. Although it was difficult to
detect a small population of variants in the host genome using a traditional approach
such as PCR, NGS, owing to its depth, enabled us to detect a novel genomic variation
(27). NGS has strongly supported the studies of viral genetic diversity, especially in RNA
viruses (28, 29), whereas the detection by NGS of quasispecies in DNA virus has been
reported less frequently (30–33). Using PCR analysis, the presence of VP1 quasispecies
has been reported in polyomavirus BK (BKV) (34). In addition, the presence of
quasi-species in the regulatory region of BKV has also been reported, with some of the
quasispecies being associated with virus replication (35). JCV quasispecies have been
reported in the regulatory region and in VP1 from urine samples using deep
sequenc-FIG 3Map of variant frequencies in the JCV genome and TAg. (A) Map of the JCV genome. Variant frequencies are shown by blue rods in the upper graph. A quasispecies with a mutation of A to C at nt 3495 in JCV Mad1 is shown by the red arrow. Transcripts encoded by JCV are shown by light blue arrows at the bottom of the panel. (B) Map of TAg. The V392G mutation is upstream of the helicase domain. (C) Alignment of large T antigens encoded by human polyomaviruses. The red arrow indicates the JCV valine at position 392. The accession codes for the amino acid sequences used in the alignment, with the corresponding polyomavirus names in parentheses, are as follows: P03070 (SV40), P03071 (BKV), A3R4N4 (KI polyomavirus [KIV]), P03072 (JCV), A5HBG1 (WU virus [WUV]), AER35104 (Merkel cell polyomavirus [MCV]), BAO09095 (Trichodysplasia spinulosa polyomavirus [TSV]), ADE45440 (human polyomavirus 6 [HPyV6]), ADE45455 (HPyV7), ADV15633 (HPyV9), AFN43007 (HPyV10), AGL07668.1 (MW polyomavirus [MWV]), AFS65330 (MX polyomavirus [MXV]), AGH58117 (HPyV12), and AGC03170.1 (STL polyomavirus [STLPyV]).
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.44.424.67.490.2]FIG 4The effects of the V392G mutation in the TAg of JCV on the expression of JC viral proteins and virus replication. (A) Transfection of JCV-Mad1 genome with and without a V392G mutation. Results of immunoblot analysis of the expression of the viral proteins in the JCV genome-transfected cells are shown (upper panels). JCV wild-type (WT) or V392G TAg mutant (Mut) or revertant (Rev) genome vectors were transfected into IMR-32 (left) and HEK293 (right) cells. The cell lysates were evaluated by immunoblotting using the indicated antibodies. The lower panels show results of real-time PCR assays for the detection of the JCV genome in the cultured supernatant. DNA was extracted from each cultured supernatant, and real-time PCR was performed.*,P⫽ ⬍0.001. (B) Transfection of JCV-Mad1 or JCV-case 6 regulatory region (RR) genome with and without V392G mutation in TAg. A JCV genome with a regulatory region from case 6 was transfected to IMR-32 cells in the presence or absence of the V392G amino acid substitution. Results of immunoblot analysis of the expression of the viral proteins in the JCV genome-transfected cells are shown (upper panels). The lower panel shows results of a real-time PCR assay for the detection of the JCV genome in the cultured supernatant.*,P⫽
⬍0.001. (C) Transfection of TAg-expressing plasmid to IMR-32 cells. Equal amounts (200 ng per well) of pCXN2-Flag vector expressing wild-type TAg (pCXN2-Flag-JCV-TAg) or V392G mutant TAg (pCXN2-Flag-JCV-TAg-mut) were transfected into IMR-32 cells. TAg, Flag, and beta-actin were detected by immunoblotting. Duplicate experiments showed similar results. (D) Cotransfection with wild-type and mutated JCV vectors. JCV wild-type and mutated vectors were cotransfected into IMR-32 cells in various ratios. Cell lysates were collected and analyzed by immunoblotting (upper panels). DNA was extracted from each supernatant, and JC viral copy numbers were determined by real-time PCR (lower panels). (E) Histology of PML clinical samples from cases 3 (left) and 6 (right). Hematoxylin and eosin (HE) staining of PML lesions shows enlarged nuclei of the oligodendrocytes and atypical astrocytes in the demyelinated lesion. Positive signals for TAg and VP1 in JCV-infected cells are indicated by immunohistochemistry.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.42.497.71.576.2]ing (36, 37). NGS analysis revealed that the JC viral population is often a complex
mixture composed of multiple viral variants that contribute to the quasispecies in the
cerebrospinal fluid (CSF) of PML patients (37). As far as we are aware, finding JCV
quasispecies with a mutated TAg in the brain of patients with PML has not been
reported previously.
The V392G mutation in JCV TAg was observed in all cases of PML examined in this
study. This indicates that JCV replicates in PML lesions in the presence of a mutant virus
which can repress virus replication. Results of a cotransfection experiment performed
with wild-type and mutant JCV genomes demonstrated that the mutation does not
function as a dominant negative. Intact TAg could fully recover the dysfunction induced
by a small population of mutant TAg (Fig. 4D). Indeed, immunohistochemistry
dem-onstrated that TAg was expressed in all six pathological samples in which the
quasi-species was detected (Fig. 4E), implying that the role of the mutation in PML lesions was
limited. Although the reason for the occurrence of such a common mutation which
reduces virus replication is unknown, the frequency of the mutation is controlled by
unknown factors, and the total expression of virus proteins is sustained in PML lesions.
Generally, replication of polyomavirus is associated with the immune status of patients
(7). Recently, immunoepitopes targeted by HLA-restricted T cells were identified in the
early viral gene region of BK polyomavirus (38). Interestingly, a prediction of
immu-noepitopes (http://www.syfpeithi.de/bin/MHCServer.dll/EpitopePrediction.htm)
identi-fied the V392 in TAg of JCV as an immunodominant epitope with a top 20 score for
HLA-A02 and -A24 nonamers, which are major HLA types in members of the Japanese
population (39). It is possible that targeting by T cells might be associated with the
occurrence of the common mutation in the JCV genome among patients with PML.
The mutation that we found causes an amino acid substitution of V392G in TAg of
the quasispecies. After the entry of JCV into the nucleus of the host cell, TAg is
transcribed and translated immediately as an early protein. TAg has DNA binding and
helicase activities and stimulates transcription of late genes and suppresses that of the
early genes (40). The amino acid substitution of V392G was located just upstream of the
helicase domain in TAg (Fig. 3B). In our experiment performed
in vitro
, the V392G amino
acid substitution dramatically decreased expression of both JCV-encoded early proteins
and JCV-encoded late proteins (Fig. 4A and B), including TAg itself (Fig. 4C). The strong
repressive function of the V392G substitution in TAg might be associated with the low
expression of TAg, the ensuing production of other viral proteins, and the inhibition of
viral replication. A recent study demonstrated that a single amino acid change in the
C-terminal pocket of the TAg origin-binding domain impaired the role of TAg in DNA
replication, suggesting that the conformational structure of TAg is crucial for its
function (41). Because V392 in TAg is highly conserved among human polyomaviruses
(Fig. 3C), the V392G substitution might cause a conformational change and low
expression in TAg. Further studies on the structure of TAg will be required to clarify the
role of the mutation in the structure, expression, and function of this protein. There are
no drugs that can reduce polyomavirus replication efficiently
in vivo
(42–44). Since the
inhibition of TAg function results in impairing virus replication (45–47), the single amino
acid V392 might be a target for the inhibition of JCV replication.
MATERIALS AND METHODS
Samples.Studies using human tissue were performed with the approval of the Institutional Review Board of the National Institute of Infectious Diseases (Approval No. 272). We examined frozen tissue from the brains of patients with PML. For NGS analysis, we examined brain samples from the six patients listed in Table 1. All patients were confirmed histologically to have PML and to be positive for JCV infection by PCR and using immunohistochemistry directed against JCV VP1. Since all specimens were obtained for pathological diagnosis, the patients did not receive any antiviral drug before the diagnosis of PML except for patient 6, who received antiretroviral therapy.
DNA extraction and real-time PCR.DNA was extracted from unfixed frozen brain tissue using a DNeasy blood and tissue kit (Qiagen, Hilden, Germany). Real-time PCR for detecting JCV and human beta-actin DNA was performed using a standard TaqMan kit protocol (Applied Biosystems, Foster City, CA) on a Mx3005P system (Agilent Technologies, Santa Clara, CA) with previously reported probe and primer sets (48, 49). Amplicon sizes of JCV and beta-actin real-time PCR were 89 bp and 60 bp, respectively. For real-time PCR, the 20-l PCR mixture included 1⫻QuantiTect probe PCR master mix
on November 7, 2019 by guest
http://jvi.asm.org/
(Qiagen), 0.3M TaqMan probe, 0.3M concentrations of forward and reverse primers, and approxi-mately 100 ng of template DNA. The reaction mixtures were incubated at 95°C for 5 min, followed by 40 cycles of 95°C for 15 s and 60°C for 1 min. The JCV DNA copy numbers per cell were calculated by dividing the JCV copy numbers by half of the beta-actin copy numbers, because each cell had two copies of the gene in two alleles (50).
PCR and sequencing.The regulatory region of JCV was amplified by PCR as described previously (51). The fragments of nt 3830 to 4100 and nt 1871 to 2254 in the JCV genome were also amplified using JCV-Mad1-3816f (5=-TCCTCACACTTGGTTTCCAAG-3=)/JCV-Mad1-4199r (5= -TGCTTCTTTTGCTGTGTATACC-3=) and JCV-Mad1-1871f (5=-AAGCCAGTGCAGGGCACCAGCT-3=)/JCV-Mad1-2254r (5=-CATGCCACAGACAT CAACAGCT-3=) primers. PCR amplification was carried out at 98°C for 30 s (one cycle), 98°C for 10 s, 55°C for 30 s, and 72°C for 30 s (35 cycles), and 72°C for 7 min (one cycle) with Phusion High-Fidelity PCR master mix (New England BioLabs, Ipswich, MA) using GeneAmp PCR System 9700 (Applied Biosystems). PCR products were purified using a QIAquick PCR purification kit (Qiagen), followed by direct sequencing with an ABI 3130 sequencer (Applied Biosystems) using a Big-Dye terminator ready reaction kit (Applied Biosystems) according to the manufacturer’s instructions.
Deep sequencing by NGS.The DNA library was prepared using a Genomic DNA Sample Prep kit (Illumina, San Diego, CA). DNA clusters were generated on a slide using a Cluster Generation kit (ver. 4) on an Illumina Cluster Station (Illumina) according to the manufacturer’s instructions. The sequencing run was performed using Illumina Genome Analyzer IIx (GA IIx) with TruSeq SBS kit v5. Fluorescent images were analyzed using the Illumina RTA1.8/SCS2.8 bacalling pipeline to obtain FASTQ-formatted se-quence data. Sese-quence reads were trimmed and assembled using VirusTAP pipeline software (https:// gph.niid.go.jp/cgi-bin/virustap/index.cgi) (52). To identify the potential pathogens, sequencing reads were analyzed by MePIC2 (53). Sequence reads were analyzed using CLC Genomics Workbench (Qiagen).
Multiple alignment and phylogenetic tree analysis.Nucleotide sequences of whole JCV genomes or TAg of 15 polyomaviruses were multiply aligned using Clustal W version 1.83 (54), and a phylogenetic tree was constructed using the neighbor-joining plot method with Genetyx software (Genetyx, Tokyo, Japan). In addition to our samples, previously reported JCV gene sequences were obtained from the GenBank database and used as reference sequences for comparisons.
Cell culture.The IMR-32 human neuroblastoma cell line was purchased from the Health Science Research Resource Bank (Osaka, Japan). IMR-32 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Thermo Fisher Scientific, Rockford, IL) with 10% fetal bovine serum (FBS), 0.1 mM MEM nonessential amino acids (Thermo Fisher Scientific, Waltham, MA), and penicillin and streptomycin (Thermo Fisher Scientific). Human embryonic kidney 293 (HEK293) cells were maintained in DMEM with 5% FBS, penicillin, and streptomycin. Both cell lines were incubated in an atmosphere of 5% CO2at 37°C
(55).
Antibodies.A mouse monoclonal antibody to simian virus 40 (SV40) TAg (PAb416; Calbiochem, San Diego, CA) was previously confirmed to cross-react with JCV TAg (56). Rabbit polyclonal antibodies to JCV VP1, VP2/3, and agnoprotein were prepared as described previously (56, 57). A mouse monoclonal antibody to beta-actin and anti-Flag rabbit polyclonal antibody were obtained from Sigma-Aldrich (St. Louis, MO).
JCV genome recombination experiments.The complete genome of JCV Mad1 strain (GenBank J02226), with an insertion of GGTC between nt positions 109 and 110, was subcloned into pUC19. Mutagenesis to construct the JCV Mad1 genome with a single A3495to C3495alteration, resulting in the
TAg Val392being altered to Gly392, was performed with a PrimeSTAR Mutagenesis Basal kit (TaKaRa
Biotechnology, Tokyo, Japan) following the manufacturer’s protocol. The sequences of the primers were as follows: forward (5=to 3=), ATCCAGGCCCCCCCAGCCATATATTGCT; reverse (5=to 3=), TGGGGGGGCCT GGATTCATTGCTT (the mutagenesis sites are underlined). After amplification, the plasmid was digested and self-ligated to construct a complete circular JCV Mad1 genome. A revertant JCV genome was also constructed from the mutated vector in the same way, and the sequences of the primers were as follows: forward (5=to 3=), ATCCAGGCCACCCCAGCCATATATTGCT; reverse (5=to 3=), TGGGGTGGCCTGGATTCATT GCTT. Both the mutated and revertant circular JCV Mad1 genomes were fully sequenced to confirm that all the sequences were identical to the wild-type JCV genome sequences except for the altered nucleotide. The JCV genome with a regulatory region from case 6 was generated by two-step recom-bination of the JCV Mad1 genome by the use of a KOD-mutagenesis kit (Toyobo, Osaka, Japan) using following primers: JCV-Mad1-36r (5=to 3=, CCTTCCCTTTTTTTTATATATACAG) and JCV-niid11-53-i37-120f (5=to 3=, TAGGGAGGAGCTGGCTAAAACTGGATGGCTGCCAGCCAAGCATGAGCTCATAATCACAAGTAAACAA AGCACAAGGGGAAGTGG) for the first step of deletion mutagenesis and JCV-niid-12-31-58r (5=to 3=, AGTTTTAGCCAGCTCCTCCCTACCT) and JCV-niid-12-31-i59-193f (5=to 3=, GGATGGCTGCCAGCCAAGCATG AGCTCATACCGTAAACAAAGCACAAGGGATGAGCTCATACCGTAAACAAAGCACAAGGGATGGCTGCCAGCCA AGCATGAGCTCATACCGTAAACAAAGCACAAGGGGAAGTG) for the second step of insertion mutagenesis. Recombination was confirmed by sequencing the plasmid DNA. pCXN2-Flag-JCV-TAg was used as a plasmid expressing Flag-tagged JCV TAg (58). Mutagenesis on pCXN2-Flag-JCV-TAg was performed in a manner similar to that described for the JCV Mad1 genome.
Transfection with viral genomes or plasmids. Cells were seeded onto type I collagen-coated 24-well plates and transfected with 200 ng of the viral genomes or pCXN2-Flag-JCV-TAg using Attractene transfection reagent (Qiagen) according to the manufacturer’s instructions. One day after transfection with viral genomes, transfected cells were transferred to type I collagen-coated 6-well plates and cultured for an additional 3 days. Cells transfected with pCXN2-Flag-JCV-TAg were collected 4 days after transfection.
on November 7, 2019 by guest
http://jvi.asm.org/
Immunoblotting.Cell lysates were prepared using M-PER mammalian protein extraction reagent (Thermo Fisher Scientific). After sonication and centrifugation, lysate proteins (total, 5l [originating from 12,500 cells in each sample]) were separated by SDS-polyacrylamide gel electrophoresis and examined by immunoblotting using anti-SV40 TAg (1:5,000), anti-VP1 (1:5,000), anti-VP2/3 (1:5,000), anti-agnoprotein (1:5,000), anti-Flag (1:5,000), and anti-beta-actin (1:5,000) antibodies. The immunopo-sitive signals were visualized with horseradish peroxidase-conjugated secondary antibodies (Promega, Madison, WI) (1:10,000), Stable Peroxidase Solution and Luminol/Enhancer Solution (Thermo Fisher Scientific), and LAS-3000 (Fujifilm, Tokyo, Japan).
Detection of JCV DNA in culture supernatants. Four days after transfection, the cell culture supernatants were filtered through a 0.22-m-pore-size filter (Merck Millipore, Darmstadt, Germany). The filtered supernatants were treated with DNase (Thermo Fisher Scientific), which was then inactivated at 95°C for 5 min. DNA was extracted from the DNase-treated culture supernatants using a DNeasy blood and tissue kit (Qiagen).
Immunohistochemistry.Immunohistochemistry was performed as described previously (58).
Statistics.All the experiments performedin vitrowere replicated at least three times. Statistical differences between experimental groups were analyzed using Student’sttest.
Accession number(s).GenBank accession numbers of JCV sequences from the clinical samples are LC164349 to LC164354. The JCV DNA sequence data analyzed by NGS in this study were deposited in the DNA Data Bank of Japan (DDBJ) (accession number DRA004832) (BioProject PRJDB4940).
ACKNOWLEDGMENTS
We are grateful to Yasuko Orba and Hirofumi Sawa of Hokkaido University for their
kind gift of the JCV genome coding vector, the plasmid expressing Flag-tagged JCV
TAg, and the antibodies to JCV proteins.
This work was supported financially by Research Program on HIV/AIDS (grant
number 15fk0108011h0503 to H. Katano) and Research Program on Emerging and
Re-emerging Infectious Diseases (grant number 16fk0108119j0001 to M. Kuroda) from
the Agency for Medical Research and Development, the Research Committee of Prion
Disease and Slow Virus Infection, Research on Policy Planning and Evaluation of Rare
and Intractable Diseases from the Ministry of Health, Labor and Welfare of Japan (to T.
Suzuki), and Grants-in-Aid for Young Scientists (B) from the Japan Society for the
Promotion of Science (grant number JP26860270 to K. Takahashi). The funders had no
role in study design, data collection and interpretation, or the decision to submit the
work for publication.
REFERENCES
1. Dalianis T, Hirsch HH. 2013. Human polyomaviruses in disease and cancer. Virology 437:63–72. https://doi.org/10.1016/j.virol.2012.12.015. 2. Jiang M, Abend JR, Johnson SF, Imperiale MJ. 2009. The role of
polyo-maviruses in human disease. Virology 384:266 –273. https://doi.org/ 10.1016/j.virol.2008.09.027.
3. Imperiale MJ. 2000. The human polyomaviruses, BKV and JCV: molecular pathogenesis of acute disease and potential role in cancer. Virology 267:1–7. https://doi.org/10.1006/viro.1999.0092.
4. Antonsson A, Pawlita M, Feltkamp MC, Bouwes Bavinck JN, Euvrard S, Harwood CA, Naldi L, Nindl I, Proby CM, Neale RE, Waterboer T. 2013. Longitudinal study of seroprevalence and serostability of the human polyomaviruses JCV and BKV in organ transplant recipients. J Med Virol 85:327–335. https://doi.org/10.1002/jmv.23472.
5. Viscidi RP, Rollison DE, Sondak VK, Silver B, Messina JL, Giuliano AR, Fulp W, Ajidahun A, Rivanera D. 2011. Age-specific seroprevalence of Merkel cell polyomavirus, BK virus, and JC virus. Clin Vaccine Immunol 18: 1737–1743. https://doi.org/10.1128/CVI.05175-11.
6. Li TC, Iwasaki K, Katano H, Kataoka M, Nagata N, Kobayashi K, Mizutani T, Takeda N, Wakita T, Suzuki T. 2015. Characterization of self-assembled virus-like particles of Merkel cell polyomavirus. PLoS One 10:e0115646. https://doi.org/10.1371/journal.pone.0115646.
7. Egli A, Infanti L, Dumoulin A, Buser A, Samaridis J, Stebler C, Gosert R, Hirsch HH. 2009. Prevalence of polyomavirus BK and JC infection and replication in 400 healthy blood donors. J Infect Dis 199:837– 846. https://doi.org/10.1086/597126.
8. Kothary N, Diak IL, Brinker A, Bezabeh S, Avigan M, Dal Pan G. 2011. Progressive multifocal leukoencephalopathy associated with efalizumab use in psoriasis patients. J Am Acad Dermatol 65:546 –551. https:// doi.org/10.1016/j.jaad.2010.05.033.
9. Padgett BL, Walker DL, ZuRhein GM, Eckroade RJ, Dessel BH. 1971.
Cultivation of papova-like virus from human brain with progressive multifocal leucoencephalopathy. Lancet i:1257–1260.
10. Hirsch HH, Kardas P, Kranz D, Leboeuf C. 2013. The human JC polyoma-virus (JCPyV): virological background and clinical implications. APMIS 121:685–727. https://doi.org/10.1111/apm.12128.
11. Richardson EP, Jr, Webster HD. 1983. Progressive multifocal leukoencephalopathy: its pathological features. Prog Clin Biol Res 105: 191–203.
12. Aksamit AJ, Jr. 1995. Progressive multifocal leukoencephalopathy: a review of the pathology and pathogenesis. Microsc Res Tech 32: 302–311. https://doi.org/10.1002/jemt.1070320405.
13. Aksamit AJ, Sever JL, Major EO. 1986. Progressive multifocal leukoencephalopathy: JC virus detection by in situ hybridization com-pared with immunohistochemistry. Neurology 36:499 –504. https:// doi.org/10.1212/WNL.36.4.499.
14. Wharton KA, Jr, Quigley C, Themeles M, Dunstan RW, Doyle K, Cahir-McFarland E, Wei J, Buko A, Reid CE, Sun C, Carmillo P, Sur G, Carulli JP, Mansfield KG, Westmoreland SV, Staugaitis SM, Fox RJ, Meier W, Goelz SE. 2016. JC polyomavirus abundance and distribution in progressive multifocal leukoencephalopathy (PML) brain tissue implicates myelin sheath in intracerebral dissemination of infection. PLoS One 11: e0155897. https://doi.org/10.1371/journal.pone.0155897.
15. Frisque RJ, Bream GL, Cannella MT. 1984. Human polyomavirus JC virus genome. J Virol 51:458 – 469.
16. Simmons DT. 2000. SV40 large T antigen functions in DNA replication and transformation. Adv Virus Res 55:75–134. https://doi.org/10.1016/ S0065-3527(00)55002-7.
17. An P, Saenz Robles MT, Pipas JM. 2012. Large T antigens of polyomaviruses: amazing molecular machines. Annu Rev Microbiol 66: 213–236. https://doi.org/10.1146/annurev-micro-092611-150154.
on November 7, 2019 by guest
http://jvi.asm.org/
18. Borowiec JA, Dean FB, Bullock PA, Hurwitz J. 1990. Binding and unwind-ing— how T antigen engages the SV40 origin of DNA replication. Cell 60:181–184. https://doi.org/10.1016/0092-8674(90)90730-3.
19. Ault GS, Stoner GL. 1993. Human polyomavirus JC promoter/enhancer rearrangement patterns from progressive multifocal leukoencephalop-athy brain are unique derivatives of a single archetypal structure. J Gen Virol 74(Pt 8):1499 –1507. https://doi.org/10.1099/0022-1317-74-8-1499. 20. Gorelik L, Reid C, Testa M, Brickelmaier M, Bossolasco S, Pazzi A, Bestetti A, Carmillo P, Wilson E, McAuliffe M, Tonkin C, Carulli JP, Lugovskoy A, Lazzarin A, Sunyaev S, Simon K, Cinque P. 2011. Progressive multifocal leukoencephalopathy (PML) development is associated with mutations in JC virus capsid protein VP1 that change its receptor specificity. J Infect Dis 204:103–114. https://doi.org/10.1093/infdis/jir198.
21. Gosert R, Kardas P, Major EO, Hirsch HH. 2010. Rearranged JC virus noncoding control regions found in progressive multifocal leukoen-cephalopathy patient samples increase virus early gene expression and replication rate. J Virol 84:10448 –10456. https://doi.org/10.1128/ JVI.00614-10.
22. Reid CE, Li H, Sur G, Carmillo P, Bushnell S, Tizard R, McAuliffe M, Tonkin C, Simon K, Goelz S, Cinque P, Gorelik L, Carulli JP. 2011. Sequencing and analysis of JC virus DNA from natalizumab-treated PML patients. J Infect Dis 204:237–244. https://doi.org/10.1093/infdis/jir256.
23. Sunyaev SR, Lugovskoy A, Simon K, Gorelik L. 2009. Adaptive mutations in the JC virus protein capsid are associated with progressive multifocal leukoencephalopathy (PML). PLoS Genet 5:e1000368. https://doi.org/ 10.1371/journal.pgen.1000368.
24. Gosert R, Rinaldo CH, Funk GA, Egli A, Ramos E, Drachenberg CB, Hirsch HH. 2008. Polyomavirus BK with rearranged noncoding control region emerge in vivo in renal transplant patients and increase viral replication and cytopathology. J Exp Med 205:841– 852. https://doi.org/10.1084/ jem.20072097.
25. Ellis LC, Norton E, Dang X, Koralnik IJ. 2013. Agnogene deletion in a novel pathogenic JC virus isolate impairs VP1 expression and virion production. PLoS One 8:e80840. https://doi.org/10.1371/journal.pone.0080840. 26. Neu U, Maginnis MS, Palma AS, Stroh LJ, Nelson CD, Feizi T, Atwood
WJ, Stehle T. 2010. Structure-function analysis of the human JC polyomavirus establishes the LSTc pentasaccharide as a functional receptor motif. Cell Host Microbe 8:309 –319. https://doi.org/10.1016/ j.chom.2010.09.004.
27. Capobianchi MR, Giombini E, Rozera G. 2013. Next-generation sequenc-ing technology in clinical virology. Clin Microbiol Infect 19:15–22. https://doi.org/10.1111/1469-0691.12056.
28. Bartolini B, Chillemi G, Abbate I, Bruselles A, Rozera G, Castrignano T, Paoletti D, Picardi E, Desideri A, Pesole G, Capobianchi MR. 2011. As-sembly and characterization of pandemic influenza A H1N1 genome in nasopharyngeal swabs using high-throughput pyrosequencing. New Microbiol 34:391–397.
29. Kuroda M, Katano H, Nakajima N, Tobiume M, Ainai A, Sekizuka T, Hasegawa H, Tashiro M, Sasaki Y, Arakawa Y, Hata S, Watanabe M, Sata T. 2010. Characterization of quasispecies of pandemic 2009 influenza A virus (A/H1N1/2009) by de novo sequencing using a next-generation DNA sequencer. PLoS One 5:e10256. https://doi.org/ 10.1371/journal.pone.0010256.
30. Solmone M, Vincenti D, Prosperi MC, Bruselles A, Ippolito G, Capobianchi MR. 2009. Use of massively parallel ultradeep pyrosequencing to char-acterize the genetic diversity of hepatitis B virus in drug-resistant and drug-naive patients and to detect minor variants in reverse transcriptase and hepatitis B S antigen. J Virol 83:1718 –1726. https://doi.org/10.1128/ JVI.02011-08.
31. Kukimoto I, Maehama T, Sekizuka T, Ogasawara Y, Kondo K, Kusumoto-Matsuo R, Mori S, Ishii Y, Takeuchi T, Yamaji T, Takeuchi F, Hanada K, Kuroda M. 2013. Genetic variation of human papillomavirus type 16 in individual clinical specimens revealed by deep sequencing. PLoS One 8:e80583. https://doi.org/10.1371/journal.pone.0080583.
32. Ruiz-Carrascoso G, Romero-Gomez MP, Plaza D, Mingorance J. 2013. Rapid detection and quantitation of ganciclovir resistance in cytomeg-alovirus quasispecies. J Med Virol 85:1250 –1257. https://doi.org/ 10.1002/jmv.23570.
33. Nishijima N, Marusawa H, Ueda Y, Takahashi K, Nasu A, Osaki Y, Kou T, Yazumi S, Fujiwara T, Tsuchiya S, Shimizu K, Uemoto S, Chiba T. 2012. Dynamics of hepatitis B virus quasispecies in association with nucleos-(t)ide analogue treatment determined by ultra-deep sequencing. PLoS One 7:e35052. https://doi.org/10.1371/journal.pone.0035052.
34. Luo C, Hirsch HH, Kant J, Randhawa P. 2012. VP-1 quasispecies in human
infection with polyomavirus BK. J Med Virol 84:152–161. https://doi.org/ 10.1002/jmv.22147.
35. Olsen GH, Hirsch HH, Rinaldo CH. 2009. Functional analysis of polyo-mavirus BK non-coding control region quasispecies from kidney transplant recipients. J Med Virol 81:1959 –1967. https://doi.org/ 10.1002/jmv.21605.
36. Van Loy T, Thys K, Tritsmans L, Stuyver LJ. 2013. Quasispecies analysis of JC virus DNA present in urine of healthy subjects. PLoS One 8:e70950. https://doi.org/10.1371/journal.pone.0070950.
37. Van Loy T, Thys K, Ryschkewitsch C, Lagatie O, Monaco MC, Major EO, Tritsmans L, Stuyver LJ. 2015. JC virus quasispecies analysis reveals a complex viral population underlying progressive multifocal leukoen-cephalopathy and supports viral dissemination via the hematogenous route. J Virol 89:1340 –1347. https://doi.org/10.1128/JVI.02565-14. 38. Cioni M, Leboeuf C, Comoli P, Ginevri F, Hirsch HH. 2016.
Characteriza-tion of immunodominant BK polyomavirus 9mer epitope T cell re-sponses. Am J Transplant 16:1193–1206. https://doi.org/10.1111/ ajt.13598.
39. Ikeda N, Kojima H, Nishikawa M, Hayashi K, Futagami T, Tsujino T, Kusunoki Y, Fujii N, Suegami S, Miyazaki Y, Middleton D, Tanaka H, Saji H. 2015. Determination of HLA-A, -C, -B, -DRB1 allele and haplotype frequency in Japanese population based on family study. Tissue Anti-gens 85:252–259. https://doi.org/10.1111/tan.12536.
40. Chen NN, Khalili K. 1995. Transcriptional regulation of human JC polyo-mavirus promoters by cellular proteins YB-1 and Pur alpha in glial cells. J Virol 69:5843–5848.
41. Meinke G, Phelan PJ, Shin J, Gagnon D, Archambault J, Bohm A, Bullock PA. 2016. Structural based analyses of the JC virus T-antigen F258L mutant provides evidence for DNA dependent conformational changes in the C-termini of polyomavirus origin binding domains. PLoS Pathog 12:e1005362. https://doi.org/10.1371/journal.ppat.1005362.
42. Knoll GA, Humar A, Fergusson D, Johnston O, House AA, Kim SJ, Ramsay T, Chasse M, Pang X, Zaltzman J, Cockfield S, Cantarovich M, Karpinski M, Lebel L, Gill JS. 2014. Levofloxacin for BK virus prophylaxis following kidney transplantation: a randomized clinical trial. JAMA 312:2106 –2114. https://doi.org/10.1001/jama.2014.14721.
43. Clifford DB, Nath A, Cinque P, Brew BJ, Zivadinov R, Gorelik L, Zhao Z, Duda P. 2013. A study of mefloquine treatment for progressive multi-focal leukoencephalopathy: results and exploration of predictors of PML outcomes. J Neurovirol 19:351–358. https://doi.org/10.1007/s13365-013 -0173-y.
44. Kalisch A, Wilhelm M, Erbguth F, Birkmann J. 2014. Progressive multifocal leukoencephalopathy in patients with a hematological malignancy: re-view of therapeutic options. Chemotherapy 60:47–53. https://doi.org/ 10.1159/000368072.
45. Tuesuwan B, Kern JT, Thomas PW, Rodriguez M, Li J, David WM, Kerwin SM. 2008. Simian virus 40 large T-antigen G-quadruplex DNA helicase inhibition by G-quadruplex DNA-interactive agents. Biochemistry 47: 1896 –1909. https://doi.org/10.1021/bi701747d.
46. Ali SH, Chandraker A, DeCaprio JA. 2007. Inhibition of simian virus 40 large T antigen helicase activity by fluoroquinolones. Antivir Ther 12: 1– 6.
47. Sharma BN, Li R, Bernhoff E, Gutteberg TJ, Rinaldo CH. 2011. Fluoro-quinolones inhibit human polyomavirus BK (BKV) replication in primary human kidney cells. Antiviral Res 92:115–123. https://doi.org/10.1016/ j.antiviral.2011.07.012.
48. Pal A, Sirota L, Maudru T, Peden K, Lewis AM, Jr. 2006. Real-time, quantitative PCR assays for the detection of virus-specific DNA in sam-ples with mixed populations of polyomaviruses. J Virol Methods 135: 32– 42. https://doi.org/10.1016/j.jviromet.2006.01.018.
49. Kuramochi H, Hayashi K, Uchida K, Miyakura S, Shimizu D, Vallbohmer D, Park S, Danenberg KD, Takasaki K, Danenberg PV. 2006. Vascular endothelial growth factor messenger RNA expression level is preserved in liver metas-tases compared with corresponding primary colorectal cancer. Clin Cancer Res 12:29 –33. https://doi.org/10.1158/1078-0432.CCR-05-1275.
50. Asahi-Ozaki Y, Sato Y, Kanno T, Sata T, Katano H. 2006. Quantitative analysis of Kaposi sarcoma-associated herpesvirus (KSHV) in KSHV-associated dis-eases. J Infect Dis 193:773–782. https://doi.org/10.1086/500560.
51. Sugimoto C, Ito D, Tanaka K, Matsuda H, Saito H, Sakai H, Fujihara K, Itoyama Y, Yamada T, Kira J, Matsumoto R, Mori M, Nagashima K, Yogo Y. 1998. Amplification of JC virus regulatory DNA sequences from cere-brospinal fluid: diagnostic value for progressive multifocal leukoenceph-alopathy. Arch Virol 143:249 –262. https://doi.org/10.1007/ s007050050284.
on November 7, 2019 by guest
http://jvi.asm.org/
52. Yamashita A, Sekizuka T, Kuroda M. 2016. VirusTAP: viral genome-targeted assembly pipeline. Front Microbiol 7:32. https://doi.org/ 10.3389/fmicb.2016.00032.
53. Takeuchi F, Sekizuka T, Yamashita A, Ogasawara Y, Mizuta K, Kuroda M. 2014. MePIC, metagenomic pathogen identification for clinical speci-mens. Jpn J Infect Dis 67:62– 65. https://doi.org/10.7883/yoken.67.62. 54. Jeanmougin F, Thompson JD, Gouy M, Higgins DG, Gibson TJ. 1998.
Multiple sequence alignment with Clustal X. Trends Biochem Sci 23: 403– 405. https://doi.org/10.1016/S0968-0004(98)01285-7.
55. Takahashi K, Orba Y, Kimura T, Wang L, Kohsaka S, Tsuda M, Tanino M, Nishihara H, Nagashima K, Sawa H, Tanaka S. 2013. Relationship between methyl CpG binding protein 2 and JC viral proteins. Jpn J Infect Dis 66:126 –132. https://doi.org/10.7883/yoken.66.126.
56. Okada Y, Sawa H, Endo S, Orba Y, Umemura T, Nishihara H, Stan AC,
Tanaka S, Takahashi H, Nagashima K. 2002. Expression of JC virus agnoprotein in progressive multifocal leukoencephalopathy brain. Acta Neuropathol 104:130 –136. https://doi.org/10.1007/s00401-002 -0526-8.
57. Suzuki S, Sawa H, Komagome R, Orba Y, Yamada M, Okada Y, Ishida Y, Nishihara H, Tanaka S, Nagashima K. 2001. Broad distribution of the JC virus receptor contrasts with a marked cellular restriction of virus replication. Virology 286:100 –112. https://doi.org/10.1006/ viro.2001.0972.
58. Sunden Y, Suzuki T, Orba Y, Umemura T, Asamoto M, Nagashima K, Tanaka S, Sawa H. 2006. Characterization and application of poly-clonal antibodies that specifically recognize JC virus large T antigen. Acta Neuropathol 111:379 –387. https://doi.org/10.1007/s00401-005 -0025-9.
on November 7, 2019 by guest
http://jvi.asm.org/