Developing and Evaluating Inhibitors against the RNase H
Active Site of HIV-1 Reverse Transcriptase
Paul L. Boyer,
aSteven J. Smith,
aXue Zhi Zhao,
bKalyan Das,
cKevin Gruber,
d,eEddy Arnold,
d,eTerrence R. Burke, Jr.,
bStephen H. Hughes
aaHIV Dynamics and Replication Program, National Cancer Institute, National Institutes of Health, Frederick, Maryland, USA
bChemical Biology Laboratory, Center for Cancer Research, National Cancer Institute, National Institutes of Health, Frederick, Maryland, USA
cRega Institute for Medical Research and Department of Microbiology and Immunology, Leuven, Belgium
dCenter for Advanced Biotechnology and Medicine, Rutgers University, Piscataway, New Jersey, USA
eDepartment of Chemistry and Chemical Biology, Rutgers University, Piscataway, New Jersey, USA
ABSTRACT
We tested three compounds for their ability to inhibit the RNase H
(RH) and polymerase activities of HIV-1 reverse transcriptase (RT). A
high-resolu-tion crystal structure (2.2 Å) of one of the compounds showed that it chelates
the two magnesium ions at the RH active site; this prevents the RH active site
from interacting with, and cleaving, the RNA strand of an RNA-DNA
heterodu-plex. The compounds were tested using a variety of substrates: all three
com-pounds inhibited the polymerase-independent RH activity of HIV-1 RT.
Time-of-addition experiments showed that the compounds were more potent if they
were bound to RT before the nucleic acid substrate was added. The compounds
significantly inhibited the site-specific cleavage required to generate the
polypu-rine tract (PPT) RNA primer that initiates the second strand of viral DNA
synthe-sis. The compounds also reduced the polymerase activity of RT; this ability was a
result of the compounds binding to the RH active site. These compounds appear
to be relatively specific; they do not inhibit either
Escherichia coli
RNase HI or
human RNase H2. The compounds inhibit the replication of an HIV-1-based
vec-tor in a one-round assay, and their potencies were only modestly decreased by
mutations that confer resistance to integrase strand transfer inhibitors (INSTIs),
nucleoside analogs, or nonnucleoside RT inhibitors (NNRTIs), suggesting that
their ability to block HIV replication is related to their ability to block RH
cleav-age. These compounds appear to be useful leads that can be used to develop
more potent and specific compounds.
IMPORTANCE
Despite advances in HIV-1 treatment, drug resistance is still a
prob-lem. Of the four enzymatic activities found in HIV-1 proteins (protease, RT
polymer-ase, RT RNase H, and integrase), only RNase H has no approved therapeutics
di-rected against it. This new target could be used to design and develop new classes
of inhibitors that would suppress the replication of the drug-resistant variants that
have been selected by the current therapeutics.
KEYWORDS
HIV-1, RNase H, active site inhibitors, magnesium chelating, structure
W
hile significant progress has been made in the treatment of HIV-1 infections, drug
resistance continues to be a problem, and new drugs will be a needed for the
foreseeable future. Generally speaking, the most effective anti-HIV drugs have targeted
three of the four enzymatic activities of HIV. There are FDA-approved drugs that target
protease (PR), integrase (IN), and the polymerase activity of reverse transcriptase (RT).
Received18 December 2017Accepted31 March 2018
Accepted manuscript posted online11 April 2018
CitationBoyer PL, Smith SJ, Zhao XZ, Das K, Gruber K, Arnold E, Burke TR, Jr, Hughes SH. 2018. Developing and evaluating inhibitors against the RNase H active site of HIV-1 reverse transcriptase. J Virol 92:e02203-17.https://doi .org/10.1128/JVI.02203-17.
EditorWesley I. Sundquist, University of Utah
Copyright© 2018 American Society for Microbiology.All Rights Reserved.
Address correspondence to Stephen H. Hughes, [email protected].
VACCINES AND ANTIVIRAL AGENTS
crossm
on November 6, 2019 by guest
http://jvi.asm.org/
However, despite serious efforts, there are no approved drugs that target the RNase H
(RH) activity of RT (1–7).
After infection of a cell by HIV-1, the single-strand RNA genome (there are two
copies of the viral RNA in HIV virions) must be converted into a linear double-stranded
DNA. The ends of this linear viral DNA are the substrates for integration into the host
cell DNA; therefore, the ends of the viral DNA must be appropriate substrates for the
viral enzyme IN. HIV-1 RT carries out the conversion of the single-stranded RNA into
double-stranded DNA. RT is a heterodimer, comprising a larger p66 subunit and a
smaller p51 subunit. The p66 and p51 proteins are initially synthesized as part of the
Gag-Pol polyprotein and are produced, in the maturing virion, by cleavage of Gag-Pol
by the viral enzyme PR. The p66 subunit of RT contains the DNA polymerase active site;
the polymerase can use either RNA or DNA as the template. Like most DNA
poly-merases, RT needs a primer to initiate DNA synthesis. The p66 subunit also contains an
RH activity, which degrades the RNA strand only if the RNA strand is in an RNA-DNA
heteroduplex, leaving a 5
=
phosphate group after cleavage. The p51 subunit does not
have any enzymatic activity and appears to play a structural role. Structural analyses of
RT with bound nucleic acid template/primers show that the 3
=
end of the primer strand
is preferentially bound at the polymerase active site. The double-stranded portion of
the nucleic acid spans the region between the two active sites, which are
approxi-mately 17 nucleotides (nt) apart. The template strand passes near the RH active site. If
the template strand is RNA, RH can interact with it and cleave the phosphate backbone
(for a current review, see reference 8).
Viral RNA is sense (plus) strand. Once the polymerase has begun copying the RNA
genome, generating minus-strand viral DNA and creating an RNA-DNA duplex, RH is
able to cleave the RNA template. Although the RH preferentially cleaves at some
positions in an RNA-DNA duplex, most of the cleavages of the duplex are relatively
nonspecific. However, there are specific RH cleavages that play an essential role in the
synthesis of the linear viral DNA. The polymerase activity of RT requires a primer, and
RH generates the RNA primers that are used to initiate synthesis of the second viral
DNA strand (plus strand). One of these plus-strand primers is particularly important, the
polypurine tract (PPT) primer. There is a second related sequence called the central PPT
(cPPT), which is in the IN-coding region; this second PPT presumably undergoes similar
processing. The RH cleavage at the 3
=
end of the PPT sequence in the RNA-DNA hybrid
determines the site where plus-strand DNA synthesis is initiated. The remaining PPT
sequence appears to be resistant to RH cleavage (possibly due to its structure), and it
is not certain whether the cleavage at the 5
=
end of the PPT is really necessary. Once
the cleavage at the 3
=
end of the PPT occurs, the resulting RNA primer can be used to
initiate second-strand DNA synthesis. RH is also responsible for the site-specific
cleav-age of the junction between the PPT RNA primer and the newly created DNA strand;
this cleavage determines the upstream end of the viral DNA. Lastly, RH cleaves near the
junction of the tRNA Lys
3primer (which serves as the primer for minus-strand DNA
synthesis) and the viral DNA. This cleavage takes place 1 nucleotide from the RNA-DNA
junction, leaving a riboA attached to the 3
=
end of the minus-strand DNA (8).
Compounds referred to as “integrase strand transfer inhibitors,” or INSTIs, that
selectively interfere with the second, or strand transfer, step of integration have been
developed. INSTIs chelate the two magnesium ions that are present at the IN active site,
and other parts of the compounds interact with both IN and the 3
=
ends of the viral
DNA. Both HIV-1 RH and HIV-1 IN belong to the functionally diverse superfamily of
DDE(D) nucleotidyltransferases. These enzymes all have two magnesium ions bound at
their active sites, which interact with the acidic amino acids of the DDE(D) motif. We
have previously reported 4-amino-1-hydroxy-2-oxo-1,8-naphthyridine-containing
com-pounds that were designed to inhibit IN and showed that some of the comcom-pounds
could also inhibit the RH and polymerase activities of RT (9). Based on these promising
results, we developed a series of compounds that were specifically intended to bind to
the RH active site. These compounds are based on the aforementioned INSTI scaffold,
which consists of a 1,8-naphthyridine ring with a chelating motif formed from a
on November 6, 2019 by guest
http://jvi.asm.org/
1-N-hydroxyl group, the 2-oxo group, and the 8-naphthyridine nitrogen. However, the
new compounds lack the 2
=
,4
=
-difluorobenzyl moiety, which was, in the parental
compounds, linked via a carboxamide to the 3-position of the heterobicyclic core. The
function of the modified benzyl ring is to stack with the penultimate dC at the 3
=
end
of the incoming viral DNA when the compounds are bound to HIV-1 IN. Here, we report
that three compounds, XZ456, XZ460, and XZ462, interfere with the RH and polymerase
activities of HIV-1 RT in enzymatic assays and block HIV-1 replication in cultured cells.
We also present a high-resolution crystal structure which demonstrates that one of the
compounds, XZ462, is bound at the RH active site.
RESULTS
The compounds inhibit the replication of an HIV-1 vector in a one-round assay.
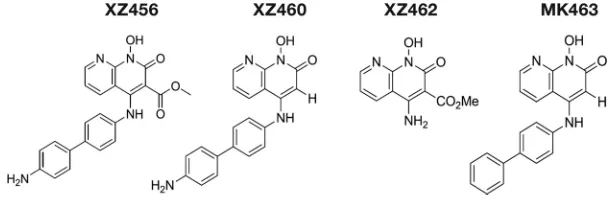
The compounds XZ456, XZ460, XZ462, and MK463 (Fig. 1), which is a reference Merck
compound (patent number WO2008010964), were tested for their abilities to inhibit
the replication of HIV-based vectors in one-round viral replication assays. XZ456 and
XZ462 weakly inhibited wild-type (WT) HIV-1, with 50% effective concentrations (EC
50s)
of 1.22
M and 1.5
M, respectively, while XZ460 and MK463 were more potent at
inhibiting WT HIV-1 (EC
50s of 94 nM and 139 nM, respectively) (Fig. 2A and B). In parallel,
these compounds were also tested for toxicity by using a sensitive assay that measures
the level of ATP in treated cells. Many RH inhibitors are toxic to cells, presumably
because the compounds can bind to host cellular enzymes that belong to the
nucle-otidyltransferase superfamily. XZ456 had no detectable toxicity, with a 50% cytotoxic
concentration (CC
50) of
⬎
250
M, while XZ462 had minor toxicity, with a CC
50of 22.4
M, which is a toxicity level similar to that seen in several FDA-approved nonnucleoside
RT inhibitors (NNRTIs). XZ460 and MK463 showed some toxicity, with CC
50s of 2.44
M
and 1.54
M, respectively (Fig. 3A and B). Thus, XZ456 had a therapeutic index of
⬎
200,
while the therapeutic indexes for XZ460 and XZ462 were 26 and 15, respectively, which
are better than that of MK463, which is 11 (6).
If this series of compounds selectively targets the RH active site of HIV-1 RT, then
HIV-1-resistant mutants that affect the potency of INSTIs, NNRTIs, and NRTIs should not
cause a significant change in the potency of the compounds relative to WT HIV-1. We
first tested the ability of these compounds to inhibit the INSTI-resistant G140S/Q148H
mutant. XZ456, XZ460, and XZ462 inhibited the G140S/Q148H double mutant with
efficacies similar to that of WT HIV-1, and the G140S/Q148H double mutant displayed
a 2-fold drop in susceptibility to MK463 compared to WT HIV-1 (Fig. 2A and B). This
initial screen suggests that these compounds do not target HIV-1 IN. Previous assays
done with the FDA-approved INSTIs raltegravir and elvitegravir showed 475-fold and
891-fold losses in potency, respectively, with this particular resistant double mutant. We
also tested these compounds against the well-known NNRTI-resistant V106A, Y181C,
Y188L, and L100I/K103N mutants. XZ456, XZ460, and XZ462 inhibited these
NNRTI-resistant mutants with antiviral activities equivalent to that against WT HIV-1; however,
there was, in some cases, an apparent increase in potency. We have no good
expla-nation for this result. MK463 inhibited all of the NNRTI-resistant mutants with efficacies
similar to that against WT HIV-1, except for the Y181C mutant, which caused a minor
FIG 1Structures of the compounds. Depiction of the chemical structures of XZ456, XZ460, XZ462, and MK463.Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:3.585.52.360.71.170.2]drop in susceptibility. Lastly, we tested the compounds against the NRTI-resistant
M184V and K70R mutants to see if there was any evidence that the compounds could
bind at the polymerase active site. XZ456 and XZ460 both inhibited these NRTI mutants
with efficacies similar to their antiviral activities against WT HIV-1. Conversely, the
NRTI-resistant mutants caused a 3- to 4-fold reduction in susceptibility to XZ462 and
MK463 (Fig. 2A and B). Although the data obtained with the viral mutants suggest that
XZ462 could be interacting with the polymerase active site, both the biochemical data
FIG 2Antiviral activities of XZ456, XZ460, XZ462, and MK463 against HIV-1 vectors that carry well-known INSTI, NNRTI, and NRTI resistance mutations. (A) Graphical representation of the EC50s (M) of XZ456, XZ460, XZ462, and MK463 against WT HIV-1
and several INSTI-, NNRTI-, and NRTI-resistant mutants, represented in different colors. The EC50s were measured using a
single-round infection assay. Error bars represent standard deviations of results of independent experiments,n⫽4. (B) Table showing the numerical values of the EC50s⫾standard deviations.
FIG 3Cytotoxicities of XZ456, XZ460, XZ462, and MK463. (A) Graphical representation of the cellular cytotoxicities of XZ456, XZ460, XZ462, and MK463. The CC50 values (M) of the compounds were
measured by monitoring the ATP levels in HOS cells. Error bars represent standard deviations of results of independent experiments,n⫽4. (B) Table showing the numerical values of cellular cytotoxicities of the compounds⫾standard deviations.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:4.585.41.431.73.389.2] [image:4.585.87.326.572.687.2]and the crystallographic data (presented below) show that XZ462 binds to and inhibits
RH. Even if XZ462 binds at both the polymerase active site and the RH active site, the
most potent compound, XZ460, was not significantly affected by the mutations we
tested.
Crystal structure of XZ462 in a complex with HIV-1 RT.
To better understand the
interaction between the RH inhibitors and the RH active site of HIV-1 RT, crystals of
HIV-1 RT were grown in a complex with the NNRTI drug rilpivirine (RPV) (10). RPV, which
binds in a hydrophobic pocket about 10 Å from the polymerase active site, helps to
prepare crystals that diffract to high resolution (11). Because the RPV was bound
⬃
50
Å from the RH active site, binding did not affect the structure of the RH domain, nor
should it affect the binding of the RH inhibitors. Preformed RT-RPV crystals were soaked
in solutions that contained each of the three RNase H inhibitors. When the structures
of the RT complexes in the three sets of soaked crystals were solved, only the RT that
was soaked with XZ462 showed the electron density for a bound inhibitor. The XZ462
complex structure was solved at a resolution of 2.2 Å, making it possible to
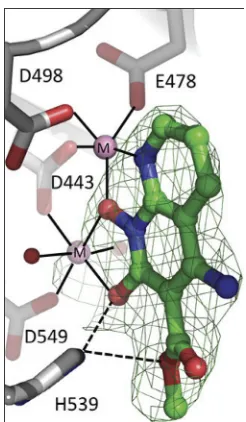
unambig-uously position the inhibitor at the RH active site (Fig. 4; Table 1). As expected, the
inhibitor chelates the two magnesium ions at the active site, which are coordinated by
the RH active site residues D443, E478, D498, and D549. The binding of XZ462 is similar
to the binding of MK1 and MK2 seen in previously solved structures (12). The inhibitor
is bound in close proximity (
⬃
4.5 Å) to residues G444, S499, Q500, A538, and H539.
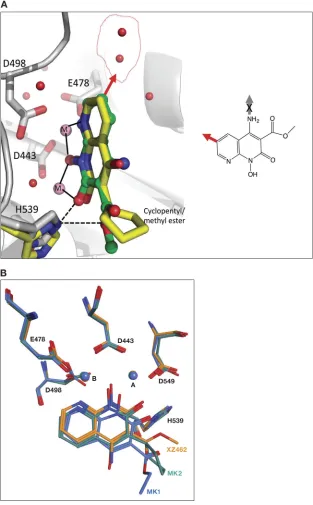
Superposition of the RT-XZ462 and RT-MK2 crystal structures shows structural
differ-ences in the contacts made by different constituents that protrude from the 3
=
position
of the naphthyridine pharmacophore. The methyl ester modification of XZ462 and the
cyclopentyl group of MK2 occupy a similar spatial orientation; however, the methyl
ester group of XZ462 has a hydrogen-bonding interaction with conserved residue H539
of RH (Fig. 5A). In addition, superposing the RT-MK1 structure with the corresponding
RT-XZ462 and RT-MK2 structures suggests that the longer ethyl ester of MK1 likely
prevents it from interacting with H539 of RH. In the crystal structure, this functionality
extends away from the RH active site (Fig. 5B). There were, in the RT-XZ462 structure,
several ordered water molecules near the RH active site. These ordered water molecules
were not seen in the previously solved RH inhibitor structures. The methyl ester
FIG 4Relative position of XZ462 in the RNase H active site of HIV-1 RT, showing contacts with residue H539. The crystal structure of XZ462 bound to the RNase H active site of HIV-1 RT shows the contacts made between XZ462 (green) and the RNase H active site residues (gray). The chelating motif of the hydroxylnaphthyridine scaffold binds the magnesium ions (shown in pink), while the methyl ester group of XZ462 forms a hydrogen bond with H539 of HIV-1 RT. Several ordered H2Omolecules are shown in red.
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:5.585.144.267.69.280.2]modification of XZ462 interacts with two water molecules within
⬃
3.15 Å, while three
water molecules are located within
⬃
4 Å of the 6
=
position on the naphthyridine core
(Fig. 5A, red arrow), and one water molecule is 3.8 Å away from the 7
=
position of the
naphthyridine ring (data not shown). The close proximities of these water molecules to
the inhibitor suggest that there are chemically feasible substitutions of the
naphthyri-dine ring that could mimic the bound water molecules and enhance the interactions of
the compounds with the RH domain (Fig. 5A, red arrow). Lastly, the RT-XZ462 structure
shows that there are two water molecules that interact with the magnesium A ion,
giving it 6 coordinating ligands; the other magnesium ion (B) has 5 coordinating
ligands (Fig. 5A), which was not observed in the previously solved RH inhibitor
structures (12).
[image:6.585.41.370.83.346.2]Polymerase-independent RH assays.
Polymerase-independent RH assays do not
contain deoxynucleoside triphosphates (dNTPs), and there is, in these assays, no primer
extension by the polymerase. However, RT is free to move along the template/primer
(T/P) substrate. The first substrate we tested falls into a class of substrates that are
designated to be DNA 3
=
end-directed substrates (13–16). In our version of this assay,
the 5
=
end of the RNA was labeled and then annealed to a shorter DNA oligonucleotide.
This creates a recessed end with the 5
=
end of the RNA extending past the 3
=
end of the
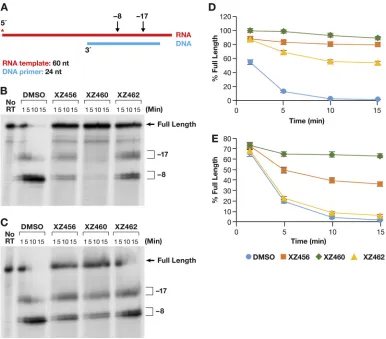
DNA primer (Fig. 6A). This substrate has been previously described (17). The results of
typical assays are shown in Fig. 6B and C. Figure 6B shows a time course in which the
RT and the compounds were allowed to interact before the radioactive T/P was added
to the reaction. Because the compounds were dissolved in dimethyl sulfoxide (DMSO),
the reaction mixture in the control lane had the same amount of DMSO added, but with
no compound. As can be seen in Fig. 6B, the RH of HIV-1 RT generated, from this
substrate, two families of RNA fragments designated
⫺
17 and
⫺
8. These are described
in more detail in Discussion. As seen in Fig. 6B, the compounds interfered with the
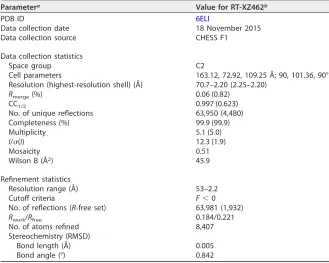
TABLE 1Crystallographic data and refinement statistics for HIV-1 RT-XZ462 complex
Parametera Value for RT-XZ462b
PDB ID 6ELI
Data collection date 18 November 2015
Data collection source CHESS F1
Data collection statistics
Space group C2
Cell parameters 163.12, 72.92, 109.25 Å; 90, 101.36, 90° Resolution (highest-resolution shell) (Å) 70.7–2.20 (2.25–2.20)
Rmerge(%) 0.06 (0.82)
CC1/2 0.997 (0.623)
No. of unique reflections 63,950 (4,480)
Completeness (%) 99.9 (99.9)
Multiplicity 5.1 (5.0)
I/(I) 12.3 (1.9)
Mosaicity 0.51
Wilson B (Å2) 45.9
Refinement statistics
Resolution range (Å) 53–2.2
Cutoff criteria F⬍0
No. of reflections (R-free set) 63,981 (1,932)
Rwork/Rfree 0.184/0.221
No. of atoms refined 8,407
Stereochemistry (RMSD)
Bond length (Å) 0.005
Bond angle (°) 0.842
aCC
1⁄2⫽ ⌺共ai⫺a兲共bi⫺b兲⁄关⌺共ai⫺a兲2⌺共bi⫺b兲2兴1⁄2, whereaiandbiare the intensities of unique reflections
merged across the observations, andaandbare their averages;Rmerge⫽ ⌺ ⌺
n
hkli⫽1
|li共hkl兲⫺I共hkl兲| ⁄⌺ ⌺ n
hkli⫽1 |li共hkl兲,
whereli共hkl兲andI共hkl兲are intensity of reflectioniand average intensity, respectively; RMSD, root mean
square deviation;I, intensity;, standard deviation.
bValues in parentheses are for the highest-resolution shell, 2.25–2.2 Å.
on November 6, 2019 by guest
http://jvi.asm.org/
FIG 5Superposing the crystal structures of XZ462 and MK2 in the RNase H active site of HIV-1 RT. (A) The crystal structures of XZ462 (green) and MK2 (yellow) in the RNase H active site of HIV-1 RT were superposed to show the different contacts between the inhibitor and residues that comprise HIV-1 RT RNase H active site and how the chelating motif interacts with the two magnesium ions (pink). Superposing both structures revealed that XZ462 (green) forms a hydrogen bond interaction with HIV-1 RT H539 (gray); MK2 (yellow) does not interact with its respective H539 (yellow). The chemical structure of XZ462 is shown in the inset. It is flipped relative to the structure shown in Fig. 1. The red arrow pointing from the 6=position of the XZ462 pharmacophore indicates sites where modifications could be made. The red arrow in the larger figure represents the same 6=position and indicates where modifi-cations could be made based on the contacts with water (depicted as red spheres inside the red circle). (B) Comparison of the binding contacts of XZ462, MK1, and MK2 in the HIV-1 RT RH active site. The crystal structures of XZ462 (orange), MK1 (blue), and MK2 (green) in their respective HIV-RT RH active sites were superposed. HIV-1 RT RH active site residues D443, E478, D498, and D549, as well as H539 and the magnesium ions A and B, are also shown.
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.49.362.73.578.2]cleavages that give rise to both the
⫺
17 and
⫺
8 products. Phosphoimager analysis
makes it possible to quantitate the results; the graph is shown in Fig. 6D. “Percent
full-length” (shown on the
y
axis) indicates the amount of the starting full-length RNA
that remained at a given time. The DMSO control lane indicates that in the absence of
an inhibitor, RH cleaved the substrate efficiently. The compounds prevented this cleavage
(to various degrees), and significantly more full-length RNA remained (Fig. 6D). XZ460 is the
most potent of the tested inhibitors, followed by XZ456 and then XZ462.
We then asked how the compounds behave when they are required to compete
with the T/P for binding to RT. The reaction was initiated by the addition of RT to the
reaction, which already contained the compound being tested (if any) and the labeled
RNA-DNA. The pattern of RNA fragments (
⫺
17 and
⫺
8) produced by the RH was not
altered in this reaction in comparison to reactions in which RT and the compounds
FIG 6Effects of the compounds on RH cleavage (3=-end-directed cleavage). (A) Overall structure of the template/primer (T/P) used for the DNA 3=-end-directed RH assay. Red, RNA; blue, DNA. The asterisk indicates the32P end label. The locations
of the RH cleavages shown in the figure are indicated above the nucleic acid. (B) Cleavages when RT and compounds were allowed to interact before the T/P was added. The control lane (no RT) indicates the size of the intact, full-length RNA. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction. The sizes of the cleavage fragments are based on their distance from the 3=end of the DNA oligonucleotide located at the polymerase active site.⫺17 is approximately the distance (in bp) between the polymerase active site and the RH active site. (C) Cleavages when compounds and T/P were mixed together first and the addition of RT was used to initiate the reaction. (D) Graphic representation of the Phosphoimager data of the RH cleavage fragments in panel B. The amount of the full-length RNA template present at a given time was measured as a percentage of the total amount of cleaved and uncleaved RNA in the reaction. For all of the polymerase-independent RH assays, the same amount of T/P and the same amount of RT was used, allowing direct comparisons to be made among the various assays. Differences in the extent of cleavage depends only on how well the RT can bind to the T/P and how well RH is able to cleave the RNA. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3. (E) Graphic representation of the Phosphoimager data of the RH cleavage fragments in panel C. The assay is similar to the assay described for panel D, except that the compounds and T/P are mixed together first. The reaction was initiated by the addition of the RT, so that the compounds and T/P competed for binding to RT. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.41.427.70.409.2]were allowed to interact before the substrate was added (Fig. 6C). However, there were
differences in the abilities of the compounds to inhibit RH. The order of the
com-pounds’ abilities to inhibit was still XZ460
⬎
XZ456
⬎
XZ462. However, in the
competition assay, XZ462 had little impact on the extent of cleavage. XZ460 was
slightly less effective in the competition assay than it was when it was able bind to RT
without T/P being present. XZ456 was significantly less active in blocking RH cleavages
in the competition assay than in the initial assay (Fig. 6E). This suggests that if T/P is
already bound, it can compete with the compounds for binding to RT. We will revisit
this question below in the section that discusses the effects of the compounds on
polymerization by RT.
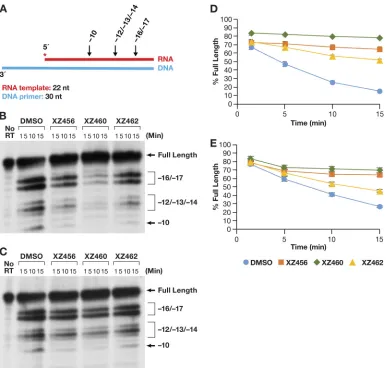
Another mode of RH activity has been designated RNA 5
=
end directed (13–16, 18).
In this case, a relatively short 5
=
-end-labeled RNA was annealed to a longer DNA
fragment (Fig. 7A). The 5
=
end of the RNA was recessed. It is unclear how the 5
=
end of
the RNA directs the positioning of the T/P, because if the RNA strand is cleaved, it must
FIG 7Effects of the compounds on RH cleavage (5=-end-directed cleavage). (A) Overall structure of the template/primer (T/P) used for the RNA 5=-end-directed RH assay. Red, RNA; blue, DNA. The asterisk indicates the32P end label. The locations
of the RH cleavages shown in the figure are indicated above the nucleic acid. (B) Cleavages when RT and compounds were allowed to interact before the T/P was added. The control lane (no RT) indicates the size of the intact, full-length RNA. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction. The sizes of the cleavage fragments are based on their distance from the 5=end of the RNA oligonucleotide located at the polymerase active site.⫺17 is approximately the distance (in bp) between the polymerase active site and the RH active site. (C) Cleavages when compounds and T/P were mixed together first and the addition of RT was used to initiate the reaction. (D) Graphic representation of the Phosphoimager results from panel B. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3. (E) Graphic representation of the Phosphoimager data from panel C. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3.
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.42.428.69.437.2]contact the RH active site. This means that the RNA strand must still be acting as the
template. When the compounds and the RT were allowed to interact before the T/P was
added, the amount of RH cleavage was much less than what was seen for the DNA
3
=
-end-directed T/P substrate (Fig. 7B), even in the DMSO control lane (see Discussion).
However, the order of effectiveness of the compounds was the same (XZ460
⬎
XZ456
⬎
XZ462) (Fig. 7D). When the T/P and a compound were allowed to interact with
RT at the same time (the reaction was initiated by the addition of RT), the pattern of
cleavages remained the same (Fig. 7C) but the amount of cleavage was reduced (Fig.
7E). The inhibition curves were more tightly clustered than they were in the assays done
with the DNA 3
=
-end-directed template.
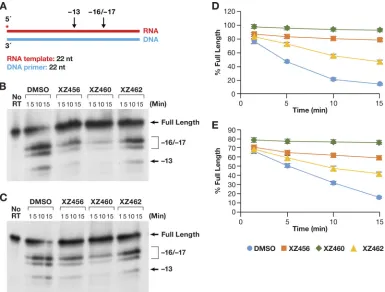
One of the last modes of polymerase-independent RH reactions has been
desig-nated internal cleavages (13, 19). The T/P is flush on both ends; there are no recessed
ends (Fig. 8A). The pattern of cleavages seen in this assay (Fig. 8B and C) was similar to
that seen for the RNA 5
=
-end-directed reaction (Fig. 7B and C). This indicates that the
RT tends to bind with the 3
=
OH group of the DNA strand located at the polymerase
active site and with the RNA acting as the template strand. RT seems to be able to bind
to this substrate more efficiently than the 5
=
-end-directed substrate. Again, the effect
of the compounds on the amount of RH cleavage was greater when the compounds
were allowed to bind to RT before the nucleic acid substrate was added (Fig. 8D) than
when the compounds competed against the T/P (Fig. 8E). The pattern of cleavages was
FIG 8Effects of the compounds of RH cleavage (internal cleavage substrate). (A) Overall structure of the template/primer (T/P) used for the internal RH assay. Red, RNA; blue, DNA. The asterisk indicates the32P end label. The location of the RH
cleavages shown in the figure are indicated above the nucleic acid. (B) Cleavages when RT and compounds were allowed to interact before the T/P was added. The control lane (no RT) indicates the size of the intact, full-length RNA. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction. The sizes of the cleavage fragments are based on their distance from the 3=end of the DNA oligonucleotide located at the polymerase active site.⫺17 is approximately the distance (in bp) between the polymerase active site and the RH active site. (C) Cleavages when compounds and T/P were mixed together first and the addition of RT was used to initiate the reaction. (D) Graphic representation of the Phosphoimager data from panel B. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3. (E) Graphic representation of the Phospho-imager data from panel C. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:10.585.42.427.71.363.2]not altered (Fig. 8B and C), and the hierarchy of the compounds was the same (XZ460
⬎
XZ456
⬎
XZ462), whether the compounds were added before the T/P or at the same
time.
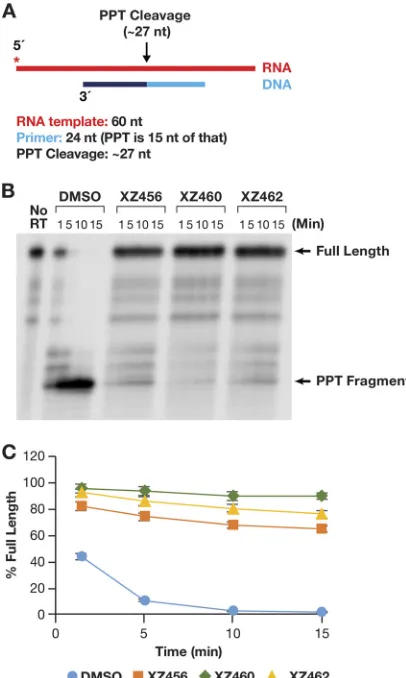
PPT cleavage.
As described in the introduction, there are a few cases in which
specific RNA primers must be generated and/or removed by the activity of RH. The
cleavage at the 3
=
end of the RNA PPT is one of two RH cleavages that generate the RNA
primer that is used to initiate second-strand viral DNA synthesis (8). As shown in Fig. 9A,
an RNA-DNA heteroduplex was used to test cleavage at the PPT site. In the absence of
any compound, the initial RNA product was a bit longer than the expected size
(probably due to the 17-nt spacing between the polymerase and RH active sites). This
fragment was quickly processed to the correct size (Fig. 9B). There was no evidence of
further processing of the secondary RNA product to produce smaller-sized fragments.
This supports the consensus that the PPT sequence is generally resistant to RH
degradation. The compounds were all able to potently inhibit this specific RNA
cleav-age (Fig. 9B and C). In this assay, it appeared that XZ462 was somewhat more potent
than XZ456 (Fig. 9C). However, the results were similar enough that it is difficult to be
certain whether this apparent difference is significant.
FIG 9Effects of the compounds on RH cleavage (PPT substrate). (A) Overall structure of the template/ primer (T/P) used for the PPT assay. Red, RNA; dark and light blue, DNA (the PPT region is dark blue). The asterisk indicates the32P end label. The locations of the RH cleavages shown in the figure are indicated
above the nucleic acid. (B) Cleavage products produced in the presence and absence of the compounds. The control lane (no RT) indicates the size of the intact, full-length RNA. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction. The location of the PPT fragment is shown. (C) Graphic representation of the Phosphoimager data for the PPT substrate (see the text). The amount of full-length product was calculated as a percentage of the total amount of RNA in the assay. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3.
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
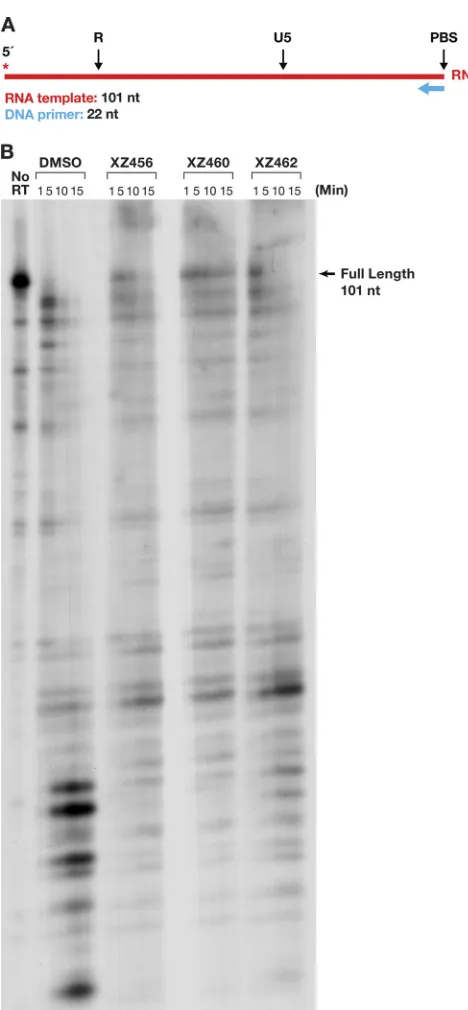
[image:11.585.103.306.72.411.2]Polymerase-dependent RNase H activity and polymerase assays.
We prepared
an RNA-DNA substrate in which the 5
=
-end-labeled RNA template strand was 101 nt
long, and a small DNA oligonucleotide was annealed to this RNA (Fig. 10A). The RT was
allowed to polymerize in the presence of a 20.0
M concentration of each dNTP. As
shown in the DMSO control lane (Fig. 10B), most of the prominent cleavages were
made near the 5
=
terminus of the RNA, where RT slowed down as it neared the end of
the template. Because the cleavage pattern was complicated, we used the amount of
FIG 10Effects of the compounds and polymerization on RH cleavages. (A) Template/primer (T/P) used for the polymerase-dependent RH assay. Red, RNA; blue, DNA. The asterisk indicates the32P end label.(B) RH cleavages made when the RT was polymerizing (polymerase-dependent RH activity). The control lane (no RT) indicates the size of the intact, full-length RNA. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:12.585.85.319.65.570.2]residual full-length RNA to monitor the ability of the compounds to inhibit the RH
activity and showed that the effectiveness of the compounds was XZ460
⬎
XZ456
⬎
XZ462 (Fig. 10B). The data also suggest that the polymerase activity of RT may have
been affected by the compounds, a possibility that is addressed below.
To test the effects of the compounds on the polymerase activity directly, the same
RNA-DNA substrate was used, but in this case, the 5
=
end of the primer DNA was
labeled (Fig. 11A). This makes it possible to monitor the sizes of the DNA extension
products that were produced by RT by using RNA as the template (RNA-dependent
DNA polymerase [RDDP] assay). An RT that is deficient in RH activity (RH
⫺) was included
in this assay. This RH-defective RT has two of the active site aspartic acid residues
changed to alanines (D443A/D549A). There are two magnesium ions at the RH active
site of RT (designated magnesium A and magnesium B), and the aspartic acid residues
FIG 11Effects of the compounds on polymerization on RNA and DNA templates. (A) T/Ps used for the polymerization assays. The long DNA and RNA templates have identical sequences and are identical in sequence to the template in Fig. 10A. The asterisk indicates the
32P end label. (B) RDDP assays done in the presence or absence of the compounds. The WT RT is shown on the left, while the RH⫺ RT is on the right. “Full-length” indicates the extension of the primer to the end of the RNA template. The control lane (no RT) indicates the size of the DNA primer. The DMSO lanes had no inhibitor present, while the other lanes had a 10.0M concentration of the indicated compound in the reaction. (C) The assays are similar to those in panel B, except that the template is DNA (DDDP assay). (D) Graphic representation of the Phosphoimager data for the RNA-dependent DNA polymerase (RDDP) assay using the WT RT (panel B). The error bars represent standard deviations of results of individual experiments as calculated by Excel. n ⫽ 3. (E) Graphic representation of the Phosphoimager data for the RDDP assay using the RH⫺RT (D443A D549A) (panel B). The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3. (F) Graphic representation of the Phosphoimager data for the DNA-dependent DNA polymerase (DDDP) assay using the WT RT (panel C). The DNA template has the same sequence as the RNA template. The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3. (G) Graphic representation of the Phosphoimager data for the DNA-dependent DNA polymerase (DDDP) assay using the RH⫺RT (panel C). The error bars represent standard deviations of results of individual experiments as calculated by Excel.n⫽3.
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:13.585.41.459.74.439.2]D443 and D549 interact with the same magnesium ion at the RH active site
(magne-sium A). With both of the aspartic acids changed to alanine, RT should not be able to
effectively bind magnesium A (13). The reason an RH
⫺RT was included was to address
the possibility that the compounds could interact with other parts of RT, including the
polymerase active site (which also has two bound magnesium ions). The loss or
FIG 11(Continued)
on November 6, 2019 by guest
http://jvi.asm.org/
[image:14.585.40.429.68.645.2]distortion of the binding of one of the magnesium ions at the RH active site should
decrease the ability of the compounds to bind at this location, which helped us
determine whether the effect of the compounds on the polymerase activity was direct
(through the polymerase active site) or indirect (through the RH active site). Figures 11B
and D show the RDDP activity of the WT RT. The presence of the compounds
significantly decreased the ability of the WT RT to extend the DNA primer. However, the
polymerase activity of the RH
⫺RT was similar in the presence or the absence of the
compounds, indicating that the polymerase inhibition was due to the compounds
interacting with the RH active site (Fig. 11B and E). Another substrate, a DNA-DNA
nucleic acid, was also tested. The sequence of the DNA template was the same as that
of the RNA template (Fig. 11A). This allowed us to determine the effects of the
compounds on the DNA-dependent DNA polymerase (DDDP) activity of RT. The results
were similar to those obtained in the RDDP assay. The compounds were able to inhibit
the DDDP activity; in this case, their ability to inhibit the polymerase activity was the
same as their ability to inhibit RH activity (XZ460
⬎
XZ456
⬎
XZ462) (Fig. 11C and 11F).
As was seen with an RNA template, the polymerase activity of the RH
⫺RT was not
affected by the addition of the compounds (Fig. 11C and G).
To ask whether the compounds could bind to RT if it was already bound to a T/P,
we did an experiment in which RT was allowed to bind to the labeled T/P before the
compounds were added. The reaction was initiated by the addition of the dNTPs and
the inhibitors, in either the presence or absence of an unlabeled nucleic acid trap. This
is basically a variation on a processivity assay. When a cold trap is present, whenever
the prebound RT falls off the labeled T/P, it will likely bind to the unlabeled nucleic acid
trap (which is present in excess) rather than rebinding to a labeled T/P, and, as a
consequence, all of the polymerization of the labeled primer that occurs involves the
prebound RT. Thus, if there is any measurable inhibition of the polymerase reaction, the
inhibitor must have been able to bind to the complex of RT and the T/P. As shown in
Fig. 12, none of the compounds had any measurable effect in the reactions that
included the cold trap. Thus, the compounds were not able to inhibit the WT RT
polymerization by a prebound RT to any significant extent, suggesting that if the T/P
is already bound to the RT, the compounds are not able to bind to the RH active site
(see Discussion).
General RH inhibition.
Because the compounds target the catalytic magnesium
ions in the HIV-1 RH, there is a concern that the active sites of other RHs could be
targeted as well. It has been reported that many RH enzymes share similar structures,
even if the amino acid homologies are low. In human cells, two RH proteins (RNase H1
and RNase H2) are known, and loss of either one causes significant cell morbidity (20).
The
Escherichia coli
RNase HI enzyme and the human RNase H2 enzyme were tested
against the compounds. The substrate was the same as that used in Fig. 6A. Neither
enzyme has an appended polymerase domain that would dictate how the RH would
bind to the nucleic acid substrate, and both enzymes should cleave at many sites in the
RNA strand in the heteroduplex. As seen in Fig. 13A and B, the compounds did not have
a significant effect on the activity of either of these enzymes. This result supports the
idea that the RH active site of HIV-1 RT is sufficiently different from the active sites of
these two cellular enzymes that compounds can be developed as selective inhibitors of
viral RH.
DISCUSSION
Considering the multiple roles the RH of HIV-1 RT has in the generation of
double-stranded DNA from the single-double-stranded RNA genome, it is an attractive target for
anti-HIV therapy. The two magnesium ions bound at the RH active site are essential for
the cleavage of the RNA phosphate backbone; compounds that interact with these ions
can block the activity of the enzyme. However, there are large number of host enzymes
for which catalysis depends on two bound two magnesium ions. To be useful as
antiviral drugs, compounds should have chemical substituents that make them specific
for the RH of HIV-1 RT. We examined the abilities of three compounds to interfere with
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
FIG 12Effects of the compounds on polymerization in the presence of a cold trap. For each reaction, RT was allowed to bind to the T/P for 5 min in the reaction mixture. The reaction was initiated by the addition of a mixture containing dNTPs, the compound to be tested (10.0M in the final reaction buffer, or an equal volume of DMSO for the control lane), and either poly(rC)-oligo(dG) for the “⫹trap” reactions or an equal volume of water for the “⫺trap” reactions. In the “Trap Test,” allowing the cold trap to bind to the RT before addition of the labeled T/P (⫹lane) caused a marked reduction in the extension of the labeled primer compared to when no trap (⫺) is added. This indicates that the cold trap is able to sequester the RT from the labeled T/P.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:16.585.122.290.71.668.2]the RH activity of HIV-1 RT and inhibit viral replication. The compounds are able to
inhibit the replication of a one-round HIV vector in a cell-based assay. The hierarchy of
potency is the same in the vector-based experiments and the
in vitro
assays. This, taken
together with the fact that mutations that reduce the susceptibility of the vectors to
several NNRTIs and IN inhibitors did not reduce the susceptibility of the vectors to the
new compounds, supports the idea that the inhibitors inhibit viral replication by
binding to the RH active site. It appears that the potency of one of the compounds,
XZ462, was reduced by mutations in the polymerase active site of RT (K70R and M184V)
that affect the susceptibility of viral replication to nucleoside analogs. It is possible that
this compound may bind at both the polymerase and the RH active sites; however, this
is the least potent of our compounds. Moreover, the crystal structure of XZ462 in a
complex with HIV-1 RT shows that this compound can bind to the RH active site, and
there is no structural evidence that it binds at the polymerase site. While all three
compounds interfered with RH activity, they differed in their potencies, which probably
reflects their abilities to bind at the RH active site.
From the known structures of RT bound to nucleic acid, it appears that for RH to
cleave the RNA strand, the DNA strand must act as the primer and the RNA strand must
act as the template. This configuration allows the RNA to be near the RH active site,
where it could interact with the magnesium ions, and be cleaved. The 3
=
OH group of
the DNA primer is known to preferentially bind at the P site of the polymerase active
site, in a configuration that allows an incoming dNTP to bind at the N site. In the
absence of an inhibitor, RNA templates are cleaved, first at
⫺
17 (relative to the end of
the DNA primer) and then at
⫺
8. The polymerase and RH active sites are approximately
17 bp apart, which explains why
⫺
17 is a favored site for cleavage. To carry out the
⫺
8
cleavages, RT must move relative to that T/P so that the 3
=
end of the primer is closer
to the RH site. The addition of the compounds to the RH assay did not alter the pattern
of cleavage fragments that were produced by the RH of HIV RT. This would match the
proposed model in which the compounds bind at the RH active site. If binding of the
compounds interfered indirectly with the binding of the nucleic acid substrate and/or
altered the ways in which RT bound the nucleic acid substrate, the compounds might
FIG 13The compounds do not affect the cleavage byE. coliRNase HI or human RNase H2. (A) RNA cleavage fragments generated byE. coliRNase HI; (B) fragments generated by human RNase H2.Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
[image:17.585.104.308.74.326.2]have altered the cleavage pattern. Although the compounds did not alter the pattern
of cleavage, they reduced the amount of cleavage; the potency of inhibition was
XZ460
⬎
XZ456
⬎
XZ462.
The assays were repeated, but the reactions were initiated by the addition of RT, so
that the compounds and T/P competed for binding to the RT. The pattern of RH
cleavage products was not affected, but the ability of the compounds to inhibit the
activity of RH was reduced. XZ462 is only modestly effective against HIV-1 RH in this
assay. This indicates that XZ462 weakly competes with the T/P for binding to the RH
active site. The T/P, which was present in the assay in excess, relative to the
com-pounds, is known to bind to RT at nanomolar concentrations. HIV-1 RT, both in the
virion and in these assays, is free to associate and disassociate from the T/P, providing
an opportunity for the compounds to bind to the free RT. Although XZ460 can still
inhibit RH in the competition assay, it is more effective if it is allowed to associate with
RH before the T/P is added. The results obtained with XZ456 suggest that it is a better
than XZ462 in terms of its ability to compete with the T/P but is less effective than
XZ460.
The RNA 5
=
-end-directed substrate does not seem to be a favored one for RH
cleavage. It is probable that the single-stranded portion of the DNA oligonucleotide
affects the ability of RT to bind to the T/P in a way that facilitates RH cleavage.
Inappropriate or ineffective binding would cause a reduction in the amount of RH
cleavage. However, the ability of the compounds to inhibit RH cleavage was unaffected:
XZ460
⬎
XZ456
⬎
XZ462.
It has been suggested that a substrate with blunt ends would not have a recessed
end to influence how RT binds to the T/P (13, 14, 19). It has also been suggested that
this would allow the RH to cleave at multiple places within the nucleic acid substrate
(13, 14, 19). It is not clear, given the structure of RT around the polymerase active RT,
just how it could bind to the internal portion of a long RNA-DNA duplex. We found,
when using this type of substrate, that the pattern of cleavages was similar to that seen
for the RNA 5
=
-end-directed assay. The cleavages were apparently not directed by RT
binding internally on the substrate but rather by binding at the 3
=
end of the DNA
strand. The same pattern was seen in the RT-initiated reactions. The compounds, as
before, inhibit in the order XZ460
⬎
XZ456
⬎
XZ462. This order was seen both in the
experiments in which the compounds were allowed to bind to RT first and in the
competition assays. However, the amount of RH cleavage is reduced for all samples
relative to the first substrate (DNA 3
=
end directed) we tested.
We also tested the impact of the compounds on one of the sites where specific
cleavage is essential for HIV replication, the cleavage that gives rise to the PPT primer.
The compounds were all able to inhibit this RH cleavage. Interestingly, the order of
effectiveness was changed to XZ460
⬎
XZ462
⬎
XZ456. It has been suggested that the
PPT region in an RNA-DNA heteroduplex has a somewhat distorted structure compared
to a “normal” nucleic acid heteroduplex. The data suggest that the compounds might
be interacting with this structure differently than they do with the other RT-substrate
complexes.
All of the assays described above are polymerase independent, i.e., the RT is not
polymerizing and the assay measures only RH activity. For RH cleavage by HIV-1 RT, the
polymerase domain helps bind the T/P and orient it so that the RNA strand is in the
proper location for cleavage to occur. In the polymerase-dependent RH activity assay,
we used a long RNA template (101 nt) to examine the pattern of RH cleavages. There
is evidence that there is little, if any, RNase H cleavage during active polymerization (13,
21, 22), although there have been suggestions that the nucleic acid can interact with
both active sites simultaneously (1, 23). In agreement with the idea that there is little
RH cleavage during polymerization, we found relatively few cleavages in the body of
the long RNA template. The inhibition order was the same as in the 3
=
-end-directed
assay: XZ460
⬎
XZ456
⬎
XZ462.
Because we know how well the compounds inhibit RH activity in assays using these
substrates, the effects of the compounds on RNA-dependent RNA polymerase (RDDP)
on November 6, 2019 by guest
http://jvi.asm.org/
and DNA-dependent DNA polymerase (DDDP) activities can be examined. One possible
problem is that if the polymerase activity is inhibited, the compounds could be bound
not only at the RH active site but also at some other site, possibly the polymerase active
site, which has two bound magnesium ions. To address this issue, we prepared an RT
variant with two of the RH active site acidic residues mutated (D443A/D539A). These
two residues interact with the magnesium ion A in the RH active site, and their removal
should affect the binding of this magnesium ion, which should reduce the ability of the
compounds to bind at the RH active site. When tested with WT RT, all of the
com-pounds inhibited polymerization; however, none of the comcom-pounds inhibited the
ability of the RH mutant to polymerize. When a DNA template was used, the
com-pounds still inhibited polymerization with the same hierarchy as they did in the RH
inhibition assays (XZ460
⬎
XZ456
⬎
XZ462), suggesting that the polymerase inhibition
is a result of the compounds binding at the RH active site. These data provide a strong
support for the idea that the inhibition of the polymerase reaction that was seen with
the WT RT was due to the binding of the compounds to the RH active site. We think it
is likely that the bound inhibitor interferes with polymerization by inhibiting
translo-cation of RT along the T/P.
To ask whether a bound T/P could prevent the compounds from binding at the RH
active site, we used a nucleic acid cold trap. In this type of assay, once RT falls off the
labeled T/P, the trap prevents the RT from rebinding to the labeled T/P. In the presence
of the cold trap, the compounds had no measurable effect on polymerization; thus, the
compounds are unable to inhibit polymerization with a prebound template primer.
The simplest explanation is that the presence of the bound template primer prevented
the compounds from binding to the RH active site. This explanation fits the observation
that the compounds are more effective if they are allowed to bind to RT before the T/P
is added to the reaction. Fortunately, both in
in vitro
assays and in infected cells, RT falls
off the template frequently, which gives the compounds a chance to bind to the RH
active site.
One last challenge for any inhibitor of the RH of HIV-1 RT is its specificity. Human
cells contain two RH enzymes, RH1 and RH2 (16). The RH domain of HIV-1 RT and the
human RH1 and RH2 enzymes share structural similarities. Inhibition of either of these
human RH enzymes has catastrophic results for the cells. We did not have human RH1
and tested the related RHI from
E. coli
. None of the compounds were able to inhibit the
E. coli
RHI nor did the compounds inhibit human RH2. The fact that the compounds did
not inhibit these cellular RHs helps to explain the observation that the best compound
(XZ460) has a respectable therapeutic index (
⬃
26) in assays done with cultured cells.
More importantly, these data show that it is possible to develop compounds that are
able to inhibit the RH of HIV-1 without also inhibiting related host enzymes.
The compounds we describe here are not the first that have been developed to bind
to the RH active site. The magnesium ions provide a site where inhibitors can bind; the
RH active site is relatively flat. Chelation of the crucial magnesium ions would prevent
the RH from cleaving the RNA strand of the RNA-DNA heteroduplex (for a review, see
reference 3). Given the need for a close association between the magnesium ions and
the RNA template for cleavage, it is not surprising that neither of our compounds, nor
those described by others, can bind to the RH active site if the nucleic acid substrate
is bound first. This conclusion is based on biochemical data and is supported by crystal
structures of the compounds bound to the RH active site (Fig. 4 and 5A and B), as well
as by other structures of the RH domain bound with RH inhibitors (for examples, see
references 2, 12, 24, and 25). However, there are differences in the behavior of our
compounds and those that have been described by others. First, some of the
com-pounds that have been reported to inhibit the RH of purified HIV RT do not inhibit viral
replication (e.g., 2). There are several possible explanations (low cell permeability,
binding to proteins in the medium, etc.); however, in terms of being a useful lead for
drug development, a compound should be able to inhibit viral replication with low
cytotoxicity. Second, our compounds inhibit both the initial primary and secondary
RNase H cleavages. A number of previously described compounds mainly inhibit the
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
secondary cleavages but have little or no effect on the primary cleavages (3, 4). Third,
our compounds inhibit the polymerase activity of HIV-1 RT, with either an RNA and/or
a DNA template, apparently by binding to the RH active site. The compounds described
by others have not been reported to show this effect. We suggest that the ability of the
compounds to inhibit the polymerase activity (as well as the RNase H activity) of HIV-1
RT by binding to the RNase H active site is an advantage. By interfering with both
activities, the compounds will further decrease the replication efficiency of the virus.
Whether this might make it more difficult for RT to develop resistance is unclear. Finally,
having a high-resolution crystal structure of XZ462 bound at the HIV-1 RH active site
which shows the surrounding ordered water molecules should help us develop
deriv-atives of these compounds with increased binding affinity and greater potency.
MATERIALS AND METHODS
Preparation of XZ456, XZ460, XZ463, and XZ462. (i) General procedures.Proton (1H) and carbon
(13C) NMR spectra were recorded on a Varian 400 MHz spectrometer or a Varian 500 MHz spectrometer
and are reported in parts per million (ppm) relative to tetramethylsilane (TMS) and referenced to the solvent in which the spectra were collected. Solvent was removed by rotary evaporation under reduced pressure, and anhydrous solvents were obtained commercially and used without further drying. Purifi-cation by silica gel chromatography was performed using Combiflash with an ethyl acetate (EtOAc)-hexane solvent system. Preparative high-pressure liquid chromatography (HPLC) was conducted using a Waters Prep LC4000 system having photodiode array detection and Phenomenex C18columns (catalog
no. 00G-4436-P0-AX; 250 mm by 21.2 mm, 10-m particle size, 110-Å pore size) at a flow rate of 10 ml/min. Binary solvent systems consisting of 0.1% aqueous trifluoroacetic acid (TFA) (A) and 0.1% TFA in acetonitrile (B) were employed with gradients as indicated. Products were obtained as amorphous solids following lyophilization. Electrospray ionization-mass spectrometric (ESI-MS) data were acquired with an Agilent liquid chromatography-mass selective detector (LC-MSD) system equipped with a multimode ion source. The purities of samples subjected to biological testing were assessed using this system and shown to beⱖ95%.
(ii) Scheme 1 (Fig. 14). (a) Methyl 4-((4=-amino-[1,1= -biphenyl]-4-yl)amino)-1-(benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridine-3-carboxylate (compound 3).
Methyl 1-(benzyloxy)-2-oxo-4-(((trifluoromethyl)sulfonyl)oxy)-1,2-dihydro-1,8-naphthyridine-3-carboxylate (compound 2, 277 mg, 0.61 mmol) was dissolved in dimethylformamide (DMF) (1.0 ml) (21). Benzidine (134 mg, 0.73 mmol) andN-ethyl-N-isopropylpropan-2-amine (0.25 ml, 1.46 mmol) were added to the clear solution. A brown solution was formed. The reaction is exothermic. The reaction mixture was stirred (room temperature, 30 min). The mixture was purified by silica gel column chromatography. Methyl 4-((4= -amino-[1,1=-biphenyl]-4-yl)amino)-1-(benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridine-3-carboxylate (compound 3) was provided as a yellow oil in an 81% yield.1H NMR (400 MHz, CDCl
3)␦10.52 (s, 1H), 8.54-8.53 (m, 1H), 7.82
(dd,J⫽8.2, 1.2 Hz, 1H), 7.67 (d,J⫽6.4 Hz, 2H), 7.41 (d,J⫽8.4 Hz, 2H), 7.33-7.28 (m, 5H), 6.98 (d,J⫽8.3 Hz, 2H), 6.80 (dd,J⫽8.2, 4.6 Hz, 1H), 6.68 (d,J⫽8.4 Hz, 2H), 5.25 (s, 2H), 3.89 (s, 3H), 3.73 (bs, 2H).13C NMR FIG 14 Scheme 1. Synthesis of compounds XZ456, XZ460, and XZ463. Reagents and conditions were as follows: (i) trifluoromethanesulfonic anhydride, triethylamine (TEA), CH2Cl2, 0°C; (ii) benzidine or 4-aminobiphenyl, N,N
-diisopropylethylamine (DIPEA), N,N-dimethylformamide (DMF); (iii) H2, Pd/C (10%), H; (iv) sodium methoxide
(NaOMe), MeOH; (v) HBr/HOAc (33%), H2O, 80°C.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:20.585.41.402.71.275.2](101 MHz, CDCl3)␦169.35, 157.22, 153.25, 152.79, 149.59, 146.21, 139.90, 138.37, 137.09, 134.44, 130.07 (2C),
129.96, 128.89, 128.36 (2C), 127.68 (2C), 127.38 (2C), 123.34 (2C), 117.13, 115.42 (2C), 108.92, 102.69, 78.00, 52.61. ESI-MSm/z: 493.1 (MH⫹).
(b) Methyl 4-((4=-amino-[1,1= -biphenyl]-4-yl)amino)-1-hydroxy-2-oxo-1,2-dihydro-1,8-naphthyri-dine-3-carboxylate (compound XZ456).Methyl 4-((4=-amino-[1,1= -biphenyl]-4-yl)amino)-1-(benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridine-3-carboxylate (compound 3, 800 mg, 0.81 mmol) was suspended in methanol (MeOH) (12 ml). Palladium on carbon (Pd/C) (10%, 80 mg) was added. After degassing, the reaction mixture was stirred at room temperature under hydrogen for 1 h. The mixture was filtered and concentrated. The residue was purified by preparative HPLC (with a linear gradient of 0 to 40% B over 30 min; retention time, 24.4 min) and provided methyl 4-((4=-amino-[1,1= -biphenyl]-4-yl)amino)-1-hydroxy-2-oxo-1,2-dihydro-1,8-naphthyridine-3-carboxylate (XZ456, 56 mg) as a yellow solid in a 17% yield.1H NMR (400 MHz, DMSO-d
6)␦9.03 (s, 1H), 8.72 (dd,J⫽4.5, 1.3 Hz, 1H), 8.56-8.54 (m, 1H), 7.58 (dd,
J⫽12.5, 8.5 Hz, 4H), 7.37 (dd,J⫽8.1, 4.6 Hz, 1H), 7.09 (dd,J⫽8.3, 6.8 Hz, 4H), 3.20 (s, 3H). ESI-MSm/z: 403.1 (MH⫹).
(c) 1-(Benzyloxy)-4-hydroxy-1,8-naphthyridin-2(1H)-one (compound 4).A suspension of methyl 1-(benzyloxy)-4-hydroxy-2-oxo-1,2-dihydro-1,8-naphthyridine-3-carboxylate (compound 1, 1.34 g, 4.13 mmol) in MeOH (30 ml) was mixed with NaOH (1.0 N, 20 ml) (26). The white suspension was heated to boiling. After 1 h, the reaction mixture turned to a clear solution. The solvent was evaporated, and the residue was refluxed (16 h). The reaction mixture was cooled with ice and adjusted to pH 2 with concentrated HCl (aqueous). The formed suspension was filtered, and the white solid was collected and dried. Compound 1-(benzyloxy)-4-hydroxy-1,8-naphthyridin-2(1H)-one (compound 4, 1.11 g) was pro-vided as a white solid in a 98% yield.1H NMR (400 MHz, DMSO-d
6)␦12.08 (brs, 1H), 8.72 (dd,J⫽4.7,
1.8 Hz, 1H), 8.27 (dd,J⫽7.9, 1.8 Hz, 1H), 7.66-7.63 (m, 2H), 7.45-7.39 (m, 3H), 7.35 (dd,J⫽7.8, 4.7 Hz, 1H), 6.05 (s, 1H), 5.14 (s, 2H), 4.05 (s, 4H).13C NMR (101 MHz, DMSO-d
6)␦160.40, 159.81, 151.79, 148.67,
135.27, 133.14, 129.83 (2C), 129.16, 128.73 (2C), 118.92, 111.71, 99.36, 77.58. ESI-MSm/z: 269.1 (MH⫹) (26, 27).
(d) 1-(Benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridin-4-yl trifluoromethanesulfonate (com-pound 5).1-(Benzyloxy)-4-hydroxy-1,8-naphthyridin-2(1H)-one (compound 4, 0.88 mg, 3.30 mmol) was dissolved in CH2Cl2(15 ml). Triethylamine (1.1 ml, 7.91 mmol) and trifluoromethanesulfonic anhydride
(3.95 ml, 3.95 mmol, 1.0 M in CH2Cl2) were added dropwise at 0°C. The solution was stirred at 0°C for 30
min. The resultant reaction mixture was concentrated and purified by silica gel column chromatography. The title compound (compound 5, 0.99 g) was provided as a white solid in a 75% yield.1H NMR (400 MHz,
CDCl3)␦8.80 (dd,J⫽4.8, 1.7 Hz, 1H), 8.09 (dd,J⫽8.0, 1.7 Hz, 1H), 7.67-7.65 (m, 2H), 7.40-7.34 (m, 4H),
6.86 (s, 1H), 5.32 (s, 2H). ESI-MSm/z: 400.9 (MH⫹).
(e) 4-((4=-Amino-[1,1=-biphenyl]-4-yl)amino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6a).1-(Benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridin-4-yl trifluoromethanesulfonate (compound 5, 167 mg, 0.41 mmol) and benzidine (92 mg, 0.50 mmol) were dissolved in DMF (1.0 ml) in a sealed vial. The solution was microwave heated (140°C, 1 h). The resultant mixture was purified by silica gel column and provided 4-((4=-amino-[1,1=-biphenyl]-4-yl)amino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6a, 73 mg) as a yellow solid in a 40% yield.1H NMR (400 MHz, DMSO-d
6)␦8.94 (s, 1H), 8.75 (dd,J⫽4.6,
1.5 Hz, 1H), 8.69 (dd,J⫽8.1, 1.6 Hz, 1H), 7.67-7.63 (m, 4H), 7.44-7.41 (m, 6H), 7.35 (d,J⫽8.4 Hz, 2H), 6.70 (d,J⫽8.5 Hz, 2H), 5.85 (s, 1H), 5.15 (s, 2H).13C NMR (101 MHz, DMSO-d
6)␦159.49, 151.55, 148.72, 148.15,
148.05, 137.71, 137.62, 135.40, 132.80, 129.82 (2C), 129.13, 128.74 (2C), 127.74, 127.48 (2C), 126.70 (2C), 124.70 (2C), 118.47, 115.11 (2C), 110.74, 94.52, 77.52. ESI-MSm/z: 435.1 (MH⫹).
(f) 4-((4=-Amino-[1,1=-biphenyl]-4-yl)amino)-1-hydroxy-1,8-naphthyridin-2(1H)-one (XZ460). The mixture of 4-((4=-amino-[1,1=-biphenyl]-4-yl)amino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6a, 73 mg, 0.17 mmol) in HBr in acetic acid (33%, 2.0 ml) and H2O (1.0 ml) was heated and stirred (80°C, 1 h). The
solvent was removed by vacuum, and the residue was triturated with acetonitrile and filtered. The solid was purified by preparative HPLC (with a linear gradient of 0 to 40% B over 30 min; retention time, 24.5 min) and provided 4-((4=-amino-[1,1=-biphenyl]-4-yl)amino)-1-hydroxy-1,8-naphthyridin-2(1H)-one (compound XZ460, 24 mg) as yellow flurry solid in 80% yield.1H NMR (400 MHz, DMSO-d
6)␦9.05 (s, 1H), 8.77 (dd,J⫽8.1, 1.5
Hz, 1H), 8.70 (dd,J⫽4.9, 1.5 Hz, 1H), 7.85-7.81 (m, 2H), 7.79-7.75 (m, 2H), 7.49-7.42 (m, 5H), 6.03 (s, 1H).13C
NMR (101 MHz, DMSO-d6)␦159.67, 149.82, 148.07, 146.82, 140.12, 139.84, 135.00, 133.99, 131.21, 128.18 (2C),
128.12 (2C), 124.18 (2C), 123.81 (2C), 117.86, 110.95, 95.66. ESI-MSm/z: 345.1 (MH⫹).
(g) 4-([1,1=-Biphenyl]-4-ylamino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6b). Com-pounds 1-(benzyloxy)-2-oxo-1,2-dihydro-1,8-naphthyridin-4-yl trifluoromethanesulfonate (compound 5, 143 mg, 0.36 mmol) and [1,1=-biphenyl]-4-amine (121 mg, 0.72 mmol) were dissolved in DMF (1 ml). The solution was microwave heated (140°C, 2 h). The resultant brown mixture was purified by silica gel column chroma-tography. 4-([1,1=-Biphenyl]-4-ylamino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6b, 106 mg) was produced as a yellow solid in a 70% yield.1H NMR (400 MHz, CDCl
3)␦8.70 (dd,J⫽4.6, 1.5 Hz, 1H), 8.18
(d,J⫽8.0 Hz, 1H), 7.69-7.67 (m, 2H), 7.60-7.55 (m, 4H), 7.46-7.42 (m, 2H), 7.38-7.31 (m, 7H), 6.76 (bs, 1H), 6.27 (s, 1H), 5.29 (s, 2H). ESI-MSm/z: 420.1 (MH⫹), 442.0 (MNa⫹), 861.0 (M
2Na⫹).
(h) 4-([1,1=-Biphenyl]-4-ylamino)-1-hydroxy-1,8-naphthyridin-2(1H)-one (compound XZ463).
The mixture of 4-([1,1=-biphenyl]-4-ylamino)-1-(benzyloxy)-1,8-naphthyridin-2(1H)-one (compound 6b, 62 mg, 0.15 mmol) in HBr-HOAc (33%, 2.0 ml) and H2O (1.0 ml) was heated and stirred (80°C, 1 h). The
solvent was removed by vacuum, and the residue was mixed with acetonitrile and filtered. The yellow residue was purified by preparative HPLC (with a linear gradient of 20% B to 80% B over 30 min; retention time, 21.9 min) and provided 4-([1,1=-biphenyl]-4-ylamino)-1-hydroxy-1,8-naphthyridin-2(1H)-one (com-pound XZ463, 37 mg) as a yellow fluffy solid in a 75% yield.1H NMR (500 MHz, DMSO-d
6)␦8.91 (s, 1H),
8.70 (dd,J⫽4.6, 1.5 Hz, 1H), 8.65 (d,J⫽8.0 Hz, 1H), 7.76 (d,J⫽8.4 Hz, 2H), 7.70 (s, 2H), 7.49 (t,J⫽
Inhibitors of HIV-1 RNase H Journal of Virology
on November 6, 2019 by guest
http://jvi.asm.org/
7.7 Hz, 2H), 7.43 (d,J⫽8.4 Hz, 2H), 7.40-7.36 (m, 2H), 6.01 (s, 1H).13C NMR (126 MHz, DMSO-d
6)␦159.89,
151.18, 148.86, 146.88, 140.02, 139.77, 136.36, 132.65, 129.43 (2C), 128.07 (2C), 127.70, 126.86 (2C), 123.84 (2C), 117.81, 110.39, 95.53. ESI-MSm/z: 330.1 (MH⫹), 681.0 (M
2Na⫹) (27, 28). (iii) Scheme 2 (Fig. 15). (a) 2-((Benzyloxy)amino)nicotinonitrile (compound 8).
2-Fluoronicotinonitrile (compound 7, 1.59 g, 13.0 mmol) andO-benzylhydroxylamine (4.55 ml, 39.0 mmol) were dissolved in DMSO (2.0 ml). The reaction mixture was microwave heated and stirred (120°C, 10 h). The reaction mixture was extracted with EtOAc. The organic phase was washed by brine and dried by sodium sulfate. After filtration and concentration, the residue was purified by silica gel column chromatography. Compound 2-((benzyloxy)amino)nicotinonitrile (compound 8, 2.78 g) was provided as a yellow oil in a 95% yield.1H NMR (400 MHz, CDCl
3)␦8.22 (dd,J⫽1.8, 5.0 Hz, 1H), 7.90 (brs, 1H), 7.84
(dd,J⫽1.6, 8.0 Hz, 1H), 7.46-7.44 (m, 2H), 7.39-7.33 (m, 3H), 6.87 (dd,J⫽5.2, 8.0 Hz, 1H), 5.04 (s, 2H).
13C NMR (100 MHz, CDCl
3)␦160.90, 151.83, 143.21, 135.71, 129.24 (2C), 128.61 (3C), 116.80, 116.17, 94.32,
78.73. ESI-MSm/z: 226.1 (MH⫹) (21, 22).
(b) Methyl 3-((benzyloxy)(3-cyanopyridin-2-yl)amino)-3-oxopropanoate (compound 9). To a solution of 2-((benzyloxy)amino)nicotinonitrile (compound 8, 3.03 g, 13.4 mmol) and triethylamine (5.64 ml, 40.3 mmol) in CH2Cl2(50 ml), methyl 3-chloro-3-oxopropanoate (4.45 ml, 40.3 mmol) was added
dropwise. The mixture was stirred (room temperature, 2 h) and then filtered and concentrated. The residue was purified by silica gel column chromatography. Compound methyl 3-((benzyloxy)(3-cyanopyridin-2-yl)amino)-3-oxopropanoate (compound 9, 3.85 g) was provided as a brown oil in an 88% yield.1H NMR (400 MHz, CDCl
3)␦8.62 (dd,J⫽2.0, 4.8 Hz, 1H), 7.91 (dd,J⫽2.0, 8.0 Hz, 1H), 7.33-7.31
(m, 2H), 7.28-7.23 (m, 4H), 4.96 (s, 2H), 3.70 (s, 3H), 3.59 (s, 2H).13C