Late Endosomal Trafficking of Alternative Serotype Adenovirus
Vaccine Vectors Augments Antiviral Innate Immunity
Jeffrey E. Teigler,aJonathan C. Kagan,bDan H. Baroucha,c
Center for Virology and Vaccine Research, Beth Israel Deaconess Medical Center, Boston, Massachusetts, USAa; Department of Pediatrics, Harvard Medical School, Boston, Massachusetts, USAb; Ragon Institute of MGH, MIT, and Harvard, Boston, Massachusetts, USAc
ABSTRACT
Adenovirus (Ad) vaccine vectors have found widespread use as vaccine platforms against multiple infections and cancers, and multiple serotypes have been shown to differ significantly in their biological properties and immune phenotypes. Our laboratory and others have previously described differential innate immune stimulation elicited by various Ad serotypes. Here, we show that Ad serotype 5 (Ad5) traffics rapidly to the nucleus following infection, whereas Ad35 and Ad26 accumulate in late endo-somes 2 to 8 h postinfection. Innate immune cytokine elicitation by all Ad serotypes was abrogated by blockade of endosomal acidification, cathepsin B, and caspase 1, suggesting that virus interactions with acid-dependent sensors, such as Toll-like recep-tor- and cathepsin-dependent inflammasome activation in late endosomes, may trigger innate immunity. These data suggest a mechanism by which Ad vectors from various serotypes differentially trigger innate antiviral pathways via distinct intracellular trafficking to late endosomes.
IMPORTANCE
Adenoviruses (Ad) are widely used for vaccination and gene therapy applications. Importantly, Ad vectors have been shown to
differ significantly in their innate immune profiles bothin vivoandin vitro. The molecular mechanism that underlies these
ob-served differences has important implications for the development of improved vaccines. In this study, we propose a mechanism in which the degree of late endosomal trafficking of Ad vectors results in differential stimulation of late endosomal pattern rec-ognition receptors.
G
iven the limitations that have become evident with adenovi-rus serotype 5 (Ad5) vectors, alternative serotype Ad vectors have been developed from serotypes against which high-level hu-moral responses are less common in the global population, including Ad35 and Ad26 (1–3). Interestingly, in addition to cir-cumventing high levels of Ad5 neutralizing antibodies, alternative serotype Ad vectors differ substantially from Ad5 in the pheno-types of innate and adaptive immune responses elicited by vacci-nation (1,4,5). Moreover, several alternative serotype Ad vector-based vaccines confer increased protection relative to Ad5 against pathogenic simian immunodeficiency virus (SIVmac251) chal-lenges in rhesus monkeys (4,6,7).Innate immune stimulation is a critical determinant for estab-lishing the magnitude and phenotype of adaptive immune re-sponses (8–11). Previously, our group and others have shown that Ad vectors differ markedly in their innate immune-stimulatory properties (12,13). In particular, Ad35 and Ad26 induce signifi-cantly higher levels of antiviral and proinflammatory cytokines and chemokines in sera from vaccinated rhesus monkeys and in fresh human peripheral blood mononuclear cells (PBMC) than Ad5 (12). However, the mechanism underlying these observed differences has not previously been determined.
Results from several groups have suggested that Toll-like re-ceptor 9 (TLR9), late endosomal lysis, and/or differences in tro-pism may contribute to innate stimulation by Ad vectors (14–17). Previous reports utilizing a variety of cell systems have also shown that Ad vectors can utilize different endosomal trafficking path-ways, such as clathrin-mediated endocytosis or macropinocytosis, and they may exit in either early or late endosomes (18–24). We therefore hypothesized that differential Ad vector trafficking and
access to late endosomal pattern recognition receptors (PRR) might account for the differential innate stimulation elicited by these vectors.
In this study, we show that Ad35 and Ad26 accumulate in the late endosomal compartment more extensively than Ad5 at 2 to 8 h following infection. Innate immune stimulation by all Ad vec-tors was sensitive to inhibivec-tors of endosomal acidification, cathep-sin B, and caspase 1. Thus, whereas Ad vectors stimulate common innate immune-sensing pathways in the late endosome, marked differences in intracellular trafficking and access to this late endo-somal compartment may account for the different innate immune phenotypes elicited by these Ad vectors.
MATERIALS AND METHODS
Viruses.Replication-incompetent Ad5, Ad35, Ad26, Ad35k5, and Ad5f35 vectors with E1/E3 deleted expressing no transgene and enhanced green fluorescent protein (EGFP) were generated as previously described (1,
25). Briefly, viruses were produced in E1-complementing PER.55K cells and purified by CsCl gradient ultracentrifugation.
Cells.A549 cells (ATCC) were serially passaged in Dulbecco’s modi-fied Eagle’s medium (DMEM) supplemented with 10% fetal calf serum
Received2 April 2014Accepted24 June 2014
Published ahead of print2 July 2014
Editor:B. H. Hahn
Address correspondence to Dan H. Barouch, dbarouch@bidmc.harvard.edu.
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.00936-14
The authors have paid a fee to allow immediate free access to this article.
on November 7, 2019 by guest
http://jvi.asm.org/
(FCS) and grown at 37°C and 5% CO2. Normal human blood was col-lected in the presence of sodium heparin, and PBMC were isolated by the Ficoll-Hypaque density gradient method (26). The cells were resuspended at a concentration of 1⫻106cells/ml in R10 medium (RPMI, 10% FCS, 50 U/ml penicillin, 50g/ml streptomycin) and further utilized inin vitro assays.
Chemicals.The endosomal inhibitor bafilomycin A1 (BafA) (Enzo Life Sciences, Farmingdale, NY); the cathepsin inhibitors Ca074-Me, CAA0225, cathepsin G inhibitor (Millipore, Billerica, MA), and Z-FA (R&D Systems, Minneapolis, MN); and the caspase inhibitors Z-LEHD, Z-WEHD, Z-YVAD, and Z-VAD (R&D Systems, Minneapolis, MN) were dissolved in sterile dimethyl sulfoxide (DMSO). The endosomal acidifi-cation inhibitors chloroquine (Sigma-Aldrich, St. Louis, MO) and ammo-nium chloride (Sigma-Aldrich, St. Louis, MO) were reconstituted in ster-ile deionized water. The concentrations of the inhibitors utilized were determined from the literature or by titration for function and cellular viability.
In vitrostimulation assay.For all reactions, 1⫻106PBMC were stimulated with 1,000 viral particles (vp)/cell of the indicated Ad vectors. The cells were treated with various chemical inhibitors prior to or follow-ing stimulation with Ad vectors. The cells were cultured at 37°C, 10% CO2, and culture supernatants were harvested 24 h following Ad vector stimulation by centrifugation at 1,400 rpm for 5 min. The supernatants were analyzed using the Millipore Milliplex MAP Magnetic Human Cy-tokine/Chemokine Panel (Millipore, Billerica, MA) according to the manufacturer’s protocol. Luminex data were acquired on a BioPlex 200 instrument running BioPlex Manager v4.1 (Bio-Rad, Hercules, CA) with an 80% to 120% standard acceptance range. The data were analyzed using GraphPad Prism v5.0. Means were compared between groups using Kruskal-Wallis tests and plotted as means and standard errors of the mean (SEM).
Fluorescent labeling of adenoviral vectors.Adenoviral vectors were fluorescently labeled with the N-hydroxysuccinimide (NHS) ester of Atto633 (Atto-tec GmbH, Siegen, Germany) as previously described (20). Briefly, the Ad vectors were incubated for 10 to 20 min at room temper-ature (RT) with Atto633 and washed 3 times at 4°C with ice-cold phos-phate-buffered saline (PBS) containing 5% (wt/vol) sucrose by centrifu-gation at 3,000 rpm for 30 min in Amicon 10,000-kDa-molecular-mass centrifuge concentration vials (Millipore, Billerica, MA). The Atto633/ capsomere ratio was determined using the extinction coefficient of Atto633 and theA260. All preparations had ratios between 0.5 and 1.8 and displayed no defect in cellular binding or trafficking.
Infection of A549 cells with Atto633-labeled Ad vectors.A549 cells (15,000) were plated onto no. 1 German glass 15-mm coverslips (BD Biosciences, San Jose, CA) and incubated overnight (O/N) at 37°C, 5% CO2. The cells were placed on ice, washed 2 times with ice-cold PBS, and overlaid with 5,000 to 50,000 vp/cell of the indicated Atto633-labeled Ad vectors diluted in unsupplemented DMEM. The cells were incubated on ice with Ad vector for 90 min with gentle rocking every 15 min. The cells were washed 2 times with ice-cold PBS, and viral infection was induced with an overlay of 37°C prewarmed DMEM supplemented with 10% FCS. The cells were cultured at 37°C, 10% CO2. At the indicated time points, the cells were fixed by immersion into ice-cold 4% paraformaldehyde diluted in PBS for 10 min. The cells were washed 2 times with ice-cold PBS and permeabilized by immersion in 0.01% saponin diluted in PBS sup-plemented with 5% FCS for 15 min. The cells were washed 3 times in ice-cold PBS and stored for further use.
Immunofluorescence histochemistry.Cells were stained by being overlaid onto a drop of the indicated antibodies at the indicated concen-trations diluted in PBS supplemented with 5% FCS in a humidified cham-ber protected from light. Primary antibodies were incubated for 1 h, and secondary antibodies were incubated for 30 min. All staining steps were FIG 1Ad35 and Ad26 accumulate more extensively than Ad5 in the late endosomal compartment. A549 cells were cold synchronized for endocytosis and infected with 5,000 to 50,000 vp/cell of Atto633-labeled Ad5, Ad35, or Ad26. The cells were fixed in 4% paraformaldehyde at various time points postinfection. Nuclei were stained for with DAPI, and EEA1, LAMP1, and M6P were stained by immunofluorescence histochemistry. The cells were imaged at⫻630 magnification by confocal microscopy with a z-stack slice depth of 0.5m and a slice interval of 0.3m. (A) Percentages of Ad5, Ad35, and Ad26 virions colocalized with nuclei, EEA1, LAMP1, and M6P per cell following synchronized infection. Z-stack images (n⫽2 to 7 replicate experiments) were analyzed for colocalization utilizing Volocity image analysis software. Compartments were identified as⬎3 times the image standard deviation, and virions were identified as particles of⬎0.078m3with intensities⬎3 times the image standard deviation. Virion colocalization with compartments was measured using Manders’ overlap
coefficients of⬎0.5. The symbols indicate time points where both Ad35 and Ad26 colocalization values were significantly different than those of Ad5, with the less significant value indicated (*,P⬍0.01;⫹,P⬍0.001; Student’sttest). The error bars indicate SEM. (B) Representative images of Ad5, Ad35, and Ad26 subcellular localization 2 h postinfection, with the boxed areas magnified in the insets. The indicated virus is shown in green, and the indicated endosomal compartment or nucleus is shown in red, with areas of overlap shown in yellow. The images were processed using Fiji. Scale bars⫽20m.
Adenovirus Vector Trafficking and Innate Immunity
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.124.462.67.285.2]followed by 3 washes with 1 ml ice-cold PBS supplemented with 5% FCS. The antibodies and concentrations used were as follows: rabbit anti-lyso-some-associated membrane protein 1 (LAMP1) (1:1,000; Abcam, Cam-bridge, MA), mouse anti-early endosome antigen 1 (EEA1) (1:500; BD Biosciences, San Jose, CA), mouse anti-mannose-6-phosphate receptor (M6P) (1:100; Abcam, Cambridge, MA), and goat anti-mouse IgG (2
g/ml) and donkey anti-rabbit IgG (2g/ml; Life Technologies, Grand Island, NY). Following staining, coverslips were inverted onto a drop of ProLong Gold containing DAPI (4=,6-diamidino-2-phenylindole) mounting medium (Life Technologies, Grand Island, NY) on microscope slides, allowed to cure overnight at room temperature, and stored at
⫺20°C for future use.
Confocal microscopy.The slides were analyzed at the Beth Israel Dea-coness Medical Center Imaging Core on a Zeiss LSM Meta with⫻10 optical magnification and a Zeiss 63⫻oil immersion lens. Images were acquired using a Zeiss LSM 510 (Carl Zeiss GmbH, Jena, Germany) in the z-stack setting with optical slices taken at a width of 0.5m and an interval of 0.3m. For each virus, time point, and endosomal marker, at least 2 to 7 independent experiments were performed with triplicate image field coverage.
Image processing and analysis.Images were processed utilizing Vo-locity Software (Perkin-Elmer, Waltham, MA). Virions were detected as particles with an intensity⬎3 times the standard deviation of the image with a volume of at least 0.078m3. Endosomal compartments were measured as positive by a signal intensity⬎3 times the standard deviation of the image. Cells were drawn manually as regions of interest (ROIs), and virion colocalization within ROIs was measured by a Manders’ overlap coefficient (42) of 0.5 or greater. Virion colocalization with a given com-partment was calculated on a per ROI basis as a percentage of the total
individual ROI virions. Images were generated using the Fiji package for ImageJ. All images for a given endosomal marker or virus were adjusted identically for brightness and contrast for the purpose of image printing, and images utilized for Volocity analysis were unaltered prior to analysis. Colocalization of viruses with a given compartment was analyzed with GraphPad Prism 5.0 (GraphPad Software, Inc., La Jolla, CA), and differ-ences in colocalization between viruses were assessed by Student’sttests.
Data analysis and statistics.Cytokine and chemokine levels were as-sessed from Luminex assays as determined using a 5-parameter logistic model. Levels of cytokines and chemokines for inhibitor-treated and un-treated cells were assessed by Kruskal-Wallis tests. The number of colo-calized virions per cell was calculated for each endosomal marker and virus analyzed, and colocalization amounts were compared between vi-ruses by Student’sttests.
RESULTS
Ad35 and Ad26 accumulate to higher levels than Ad5 in the late
endosomal compartment at 2 to 8 h following infection.To
as-sess the intracellular-trafficking pathways for Ad5, Ad35, and Ad26, vectors were covalently labeled with the NHS ester of Atto633 and tested for viability as previously described (20). Briefly, A549 cells were infected at an MOI of 5,000 to 50,000 vp/cell of fluorescently labeled Ad vector, and colocalization was measured over time by con-focal microscopy for the early endosome marker EEA1 or the late endosomal marker LAMP1 or M6P by immunofluorescence histo-chemistry, and nuclei were labeled with DAPI (27–29). Viruses were measured for colocalization in a given endosomal/nuclear compart-ment by calculation of the Manders’ overlap coefficients, and
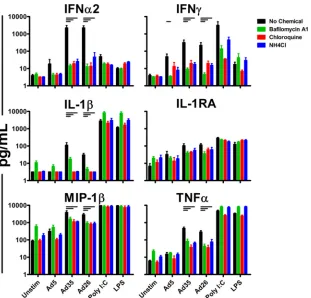
differ-FIG 2Ad innate immune stimulation is blocked by inhibitors of late endosomal acidification. Fresh human PBMC (n⫽4/group) were preincubated for 1 h with BafA (1M), chloroquine (200M), or ammonium chloride (1 mM) prior to infection with 1,000 vp/cell of Ad5, Ad35, or Ad26. Induced cytokines and chemokines were measured 24 h postinfection by Luminex assays. The data are shown as mean responses of individual donors and SEM. The horizontal bars indicatePvalues of 0.0286; Kruskal-Wallis test. High-molecular-weight (HMW) poly(I·C) (10g/ml) and LPS (1 ng/ml) were included as positive controls.
Teigler et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.140.451.64.362.2]ences between vector localizations at a given time point were deter-mined by Student’sttests (42).
Ad5 virions rapidly accumulated in the perinuclear region, with 72.6% of virions colocalizing with nuclear staining by 2 h postinfection (Fig. 1AandB). Colocalization of Ad5 with the early endosomal marker EEA1 occurred only minimally and never ex-ceeded 7% at any time point measured (Fig. 1A). Ad5 virions also localized to minor levels with late endosomes, as indicated by low levels of colocalization with LAMP1 or M6P at 2 h postinfection (4.0% and 2.7% colocalization, respectively) (Fig. 1A and B). These results indicate that Ad5 rapidly trafficked to the nucleus and only minimally accumulated in early and late endosomal compartments.
Ad35 and Ad26 exhibited marked differences in intracellu-lar-trafficking pathways compared with Ad5. Ad35 and Ad26 displayed substantial but transient colocalization with the early endosomal marker EEA1 at 30 min (18.5% and 45.5% colocal-ization, respectively) and at 1 h (11.9% and 19.9% colocaliza-tion, respectively) postinfection (Fig. 1A). The colocalization of Ad35 and Ad26 with early endosomes was greater than that observed for Ad5 at both 30 min (3.2- and 7.8-fold greater colocalization, respectively;P ⫽0.0001 andP⬍ 0.0001, re-spectively; Student’sttests) and at 60 min (2.8- and 4.6-fold
greater colocalization, respectively;P⫽0.0082 andP⫽0.0048, respectively). At later time points, both Ad35 and Ad26 dem-onstrated substantial colocalization with the late endosomal marker LAMP1, with 12.6% and 25.7% of virions colocalizing with late endosomes at 2 h postinfection, respectively (Fig. 1A
andB). Ad35 and Ad26 also colocalized with the late endo-somal marker M6P at 2 h postinfection (15.4% and 26.2% of virions, respectively) (Fig. 1A and B). Analysis of adherent PBMC, which include multiple cellular subsets (30), showed that Ad35 and Ad26 similarly colocalized with late endosomes by 2 h postinfection (data not shown). Both Ad35 and Ad26 accumulated in the late endosomal compartment to signifi-cantly higher levels than Ad5 at 2 h postinfection as measured by both LAMP1 (3.1- and 6.4-fold greater colocalization, re-spectively;P⫽0.0088 andP⫽0.0001, respectively) and M6P (5.8- and 9.8-fold greater colocalization, respectively; P ⬍
0.0001 for both) colocalization. This association of Ad35 and Ad26 in the late endosomes persisted for all time points ob-served between 2 and 8 h postinfection. Taken together, these results indicate that Ad35 and Ad26 display a markedly differ-ent intracellular trafficking pattern than Ad5, characterized by substantially greater association with late endosomes 2 to 8 h postinfection.
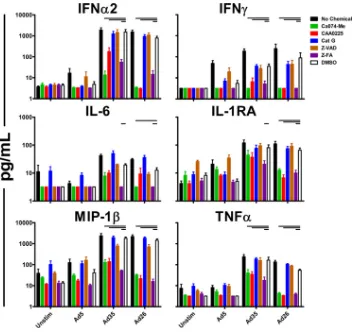
FIG 3Ad innate immune stimulation is sensitive to cathepsin B, cathepsin L, and pancathepsin inhibition. Fresh human PBMC (n⫽4/group) were preincu-bated for 1 h with the cathepsin B inhibitor Ca074-Me (10M), the cathepsin L inhibitor CAA0225 (10M), a cathepsin G inhibitor (10M), the pancaspase inhibitor Z-VAD (100M), the pancathepsin inhibitor Z-FA (100M), or a DMSO vehicle control. The cells were stimulated with 1,000 vp/cell of Ad5, Ad35, or Ad26, and cytokines were measured 24 h postinfection by Luminex assays. The data are shown as mean responses of individual donors and SEM. The horizontal bars indicatePvalues of 0.0286; Kruskal-Wallis test.
Adenovirus Vector Trafficking and Innate Immunity
on November 7, 2019 by guest
http://jvi.asm.org/
[image:4.585.113.473.64.396.2]Ad innate immune stimulation is blocked by inhibitors of
late endosomal acidification.Ad35 and Ad26 induce markedly
higher levels of antiviral and proinflammatory cytokines and chemokines, including alpha interferon (IFN-␣), IFN-␥, IP-10, interleukin 6 (IL-6), and IL-1RA, than Ad5 in sera from vacci-nated rhesus monkeys and in human PBMC (12). Interestingly, these differences in innate immune cytokines and chemokines corresponded to the observed levels of Ad vector trafficking to the late endosome. To probe the possible late endosomal innate sens-ing pathways the different Ad vectors might activate, we assessed the dependence of innate triggering by these vectors on late endo-somal acidification. Fresh human PBMC (n⫽4/group) were iso-lated and preincubated with the endosomal acidification inhibitor BafA, chloroquine, or ammonium chloride for 1 h prior to infec-tion with 1,000 vp/cell of Ad5, Ad35, or Ad26 (31,32). Cytokine induction was measured 24 h following infection by Luminex as-says, as previously described (12).
Preincubation of human PBMC with BafA, chloroquine, or ammonium chloride markedly decreased induction of innate cy-tokines and chemokines by all the Ad vectors studied (Fig. 2). This inhibition was most pronounced for IFN-␣2 with preincubation of PBMC with BafA, chloroquine, or ammonium chloride, lead-ing to reduced levels of IFN-␣2 upon stimulation with Ad5, Ad35, or Ad26 (Fig. 2). Activities of inhibitors on intracellular innate
immune signaling were confirmed by the inhibition of intracellu-lar poly(I·C) sensing, but not extracelluintracellu-lar lipopolysaccharide (LPS) sensing, by all three endosomal acidification inhibitors. To-gether, these data suggest that innate stimulation by all three vec-tors was sensitive to inhibivec-tors of late endosomal acidification.
Innate immune stimulation by Ad5, Ad35, and Ad26 is sen-sitive to cathepsin B, cathepsin L, and pancathepsin inhibition.
We next evaluated the roles of specific cathepsins in Ad innate sensing. Cathepsins are a group of late endosomal papain-like proteases that play several important roles in late endosomal cargo processing, as well as autophagy and apoptosis (33). Fresh human PBMC (n⫽4/group) were preincubated for 1 h with the cathepsin B inhibitor Ca074-Me, the cathepsin L inhibitor CAA0225, a ca-thepsin G inhibitor, the pancaspase inhibitor Z-VAD, the pan-cathepsin inhibitor Z-FA, or a DMSO vehicle control. The cells were then stimulated with 1,000 vp/cell of Ad5, Ad35, or Ad26. Elicited cytokine and chemokine levels were analyzed 24 h postin-fection by Luminex assays as described above.
Human PBMC preincubated with Ca074-Me, CAA0225, or Z-FA exhibited reduced levels of multiple cytokines and chemo-kines produced in response to Ad vectors. In particular, levels of IFN-␣2, IFN-␥, MIP-1, and tumor necrosis factor alpha (TNF-␣) were significantly reduced following preincubation of human PBMC with these cathepsin inhibitors in response to Ad35
FIG 4Ad innate immune stimulation is reduced by inhibition of caspase 1. Fresh human PBMC (n⫽3 or 4/group) were preincubated for 1 h with the caspase 9 inhibitor Z-LEHD (100M), the caspase 1 inhibitor Z-WEHD (100M), the caspase 4 inhibitor Z-YVAD (100M), or DMSO vehicle control. The PBMC were then stimulated with 1,000 vp/cell of Ad5, Ad35, or Ad26, and the elicited cytokine and chemokine levels were measured 24 h following infection by Luminex assays. The data are shown as mean responses of individual donors and SEM. The horizontal bars indicatePvalues of 0.0286; Kruskal-Wallis test.
Teigler et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:5.585.113.472.64.394.2]and Ad26 relative to the DMSO vehicle control, and similar trends were observed for Ad5 (Fig. 3). Preincubation with a cathepsin G inhibitor, however, had minor effects on the cytokines and chemokines elicited by Ad35 and Ad26 (Fig. 3). These results sug-gest that Ad35 and Ad26, and to a lesser extent Ad5, are inhibited by cathepsin B and pancathepsin inhibition, suggesting the func-tional relevance of late endosomal localization to triggering innate immunity.
Inhibition of caspase 1 reduces innate immune stimulation
elicited by Ad5, Ad35, and Ad26.Caspases are proteases that serve
as major components of proapoptotic and proinflammatory path-ways (34). To assess the role of caspases in Ad innate stimulation, fresh human PBMC (n⫽3 or 4/group) were stimulated with Ad5, Ad35, or Ad26 following preincubation for 1 h with the caspase 9 inhibitor Z-LEHD, the caspase 1 inhibitor Z-WEHD, the caspase 4 inhibitor Z-YVAD, or DMSO vehicle control. Preincubation of human PBMC with Z-WEHD led to marked reduction of IFN-␣2 and IFN-␥elicited by Ad35 and Ad26 (Fig. 4). In contrast, prein-cubation of human PBMC with the caspase 9 inhibitor Z-LEHD or the caspase 4 inhibitor Z-YVAD had little to no effect on the innate cytokine stimulation by Ad5, Ad35, and Ad26. Similar to experiments performed with cathepsin inhibitors, the overall lev-els of cytokines and chemokines elicited by Ad5 were lower than those by Ad35 and Ad26, yet they remained sensitive to Z-WEHD inhibition. These data suggest a role for caspase 1 in the induction of immune responses by these Ad vectors.
Innate stimulation by Ad5, Ad35, and Ad26 is initiated
be-tween 0 and 6 h postinfection.To determine the timing of the
initial triggering of the innate sensing pathways by the Ad vectors, fresh human PBMC were stimulated with 1,000 vp/cell of Ad5, Ad35, or Ad26. At 0, 4, 6, 8, or 12 h postinfection, the cells were
treated with chloroquine, ammonium chloride, Ca074-Me, CAA0225, or Z-FA at the concentrations described above. Cyto-kine and chemoCyto-kine levels were assessed 24 h following infection by Luminex assays and compared to those of infected cells with mock inhibition.
Addition of inhibitors at the time of infection (0 h) effectively inhibited induction of IFN-␣2, IFN-␥, and TNF-␣(Fig. 5), in agreement with the previous experiments (Fig. 2to4). For all three vectors, the majority of inhibition of innate sensing by chlo-roquine and Z-FA occurred earlier than 6 h postinfection, with the most prominent inhibition caused by Z-FA occurring up to 12 h postinfection (Fig. 5). Interestingly, inhibition of cytokine re-sponses elicited by Ad26, and to a lesser extent by Ad5, was sensi-tive to inhibition up to 6 h postinfection, while those elicited in response to Ad35 were able to be significantly inhibited up to 4 h postinfection. Taken together, these data indicate that innate im-mune sensing of Ad5, Ad35, and Ad26 was initiated between 0 and 6 h postinfection. These findings are consistent with the possibility that localization of Ad vectors to the late endosome is important for Ad innate sensing, as inhibition of innate immune sensors reduced Ad innate immune stimulation during times at which Ad vectors were observed to be present in late endosomes.
Chemical inhibition of innate immune stimulation does not
perturb transduction efficiency.We next investigated whether
chemical inhibition of Ad innate immune stimulation was simply due to blockade of Ad infectivity. To test whether chemical inhib-itors had an effect on Ad35 or Ad26 transduction efficiency, hu-man PBMC (n⫽4/group) were treated at the time of infection with chloroquine, ammonium chloride, Ca074-Me, CAA0225, Z-FA, or DMSO vehicle control as described above. The cells were then stimulated with 1,000 vp/cell of Ad35 or Ad26 expressing
FIG 5Ad innate immune stimulation occurs with similar kinetics and is sensitive to chemical inhibitors from 0 to 6 h postinfection. Fresh human PBMC (n⫽ 4/group) were stimulated with 1,000 vp/cell of Ad5, Ad35, or Ad26. The chemical inhibitor chloroquine, ammonium chloride, Ca074-Me, CAA0225, or Z-FA was added at the indicated times following infection. The levels of elicited cytokines and chemokines were measured 24 h following infection by Luminex assays and compared to an infected, mock-inhibited control (Media). The error bars indicate SEM.
Adenovirus Vector Trafficking and Innate Immunity
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.122.462.64.317.2]EGFP. The levels of elicited cytokines and chemokines were mea-sured 24 h postinfection by Luminex assays, and the levels of transduced cells were measured by flow cytometry.
Similar to our results described above, chloroquine, ammo-nium chloride, Ca074-Me, CAA0225, and Z-FA reduced cytokine and chemokine induction in human PBMC elicited by Ad35 or Ad26 expressing EGFP at 24 h postinfection (data not shown). Flow cytometry on stimulated cells showed no inhibition of trans-duction of PBMC by either Ad35 or Ad26 for any of the chemical inhibitors analyzed, as measured by the percentage of EGFP-pos-itive cells 24 h postinfection (Fig. 6). These results show that chemical inhibition of Ad35 and Ad26 innate sensing did not sub-stantially reduce vector infectivity.
Both Ad fiber and capsid components influence
intracellu-lar-trafficking pathways.Both fiber and capsid components of
Ad5 and Ad35 can influence innate stimulatory properties of Ad vectors (12). In particular, both Ad35 fiber and capsid compo-nents were required to confer the increased innate stimulatory properties of Ad35 relative to Ad5. We sought to determine the influence of Ad fiber and capsid components on access to the late endosomal compartment and the associated Ad innate sensing mechanisms. To explore the influence of the Ad fiber and capsid on Ad intracellular trafficking, we utilized Ad35k5, a chimeric Ad35 with its fiber knob domain replaced with that of Ad5, and Ad5f35, a chimeric Ad5 with its fiber protein replaced with that of Ad35. A549 cells were infected with Ad35k5 and Ad5f35 vectors and subsequently processed for immunofluorescence histochem-istry and confocal microscopy as described in the legend toFig. 1. Ad5f35 and Ad35k5 both displayed initial colocalization with the EEA1 compartment at 30 min (32.9% and 20.2%, respectively) and 60 min (21.4% and 22.4%, respectively) similar to that ob-served with Ad35. Furthermore, both Ad5f35 and Ad35k5 dis-played increased late endosomal trafficking beginning at 2 h postinfection, as indicated by higher colocalization than Ad5 at 2 h for both LAMP1 (2.8- and 5.9-fold greater colocalization than Ad5, respectively;P⫽0.0215 andP⬍0.0001, respectively; Stu-dent’sttests) and M6P (5.6- and 8.8-fold higher colocalization, respectively;P⫽0.0003 andP⬍0.0001, respectively) (Fig. 7). These data indicate that both fiber and capsid components of Ad vectors influence intracellular trafficking patterns and suggest in-creased late endosomal association as a mechanism for the
previ-ously observed increase in innate cytokine and chemokine stimu-lation by both Ad5f35 and Ad35k5 relative to Ad5 (12).
DISCUSSION
In this study, we show that Ad5, Ad35, and Ad26 differ substan-tially in their intracellular trafficking patterns, with Ad35 and Ad26 accumulating to a greater degree than Ad5 in late endo-somes and Ad5 rapidly trafficking to the nucleus. All three vectors were sensitive to chemical inhibitors of late endosomal innate im-mune sensing, in particular endosomal acidification, cathepsin B, and caspase 1. These studies suggest a mechanism that could ac-count for the differential innate immune-stimulatory phenotypes of Ad vectors in which intracellular trafficking to the late endo-some contributes to these observed differences in innate immu-nity (12,13) (Fig. 8). In the late endosome, virions stimulate ste-reotypical innate immune pathways that require endosomal acidification, cathepsins, and caspase 1 (Fig. 8). Taken together, these data suggest that intracellular Ad vector trafficking critically influences innate immune stimulation by various Ad serotypes.
Innate immune stimulation elicited in response to all three Ad vectors was sensitive to inhibition of acidification, cathepsins, and caspase 1, suggesting that accumulation in late endosomes, rather than differential PRR stimulationper se, may be responsible for
FIG 6Chemical inhibition of Ad5, Ad35, or Ad26 innate immune signaling does not block Ad transduction. Fresh human PBMC (n⫽4/group) were synchronously inhibited with chloroquine, ammonium chloride, Ca074-Me, CAA0225, Z-FA, or DMSO vehicle control at the same concentrations as for
Fig. 2and3and stimulated with 1,000 vp/cell of Ad35 or Ad26 expressing EGFP. Twenty-four hours following infection, the percent EGFP cellular transduction was measured by flow cytometry. Unstim, unstimulated. The error bars indicate SEM.
FIG 7Both fiber and capsid components influence Ad vector intracellular-trafficking patterns. A549 cells were incubated with 50,000 vp/cell of Atto633-labeled Ad35k5 or Ad5f35 and fixed at the indicated times postinfection by immersion in 4% paraformaldehyde. The cells were stained for the indicated markers by immunohistochemistry, and virion colocalization was assessed as described forFig. 1. The graphs indicate the percentages of virions per cell colocalized with the indicated compartment at the indicated time points, with Ad5 and Ad35 data fromFig. 1shown as dashed lines for reference. The sym-bols indicate the time points where both Ad35k5 and Ad5f35 colocalization values were significantly different than those of Ad5, with the less significant value indicated (*,P⬍0.01;⫹,P⬍0.001; Student’sttest). The error bars indicate SEM.
Teigler et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.331.515.64.331.2] [image:7.585.70.256.64.164.2]the observed differences in innate immune phenotypes elicited by these vectors. The requirement for endosomal acidification in in-nate sensing suggests that late endosomal TLRs may be the endo-somal acidification-dependent portion of the sensing pathway (Fig. 2and8). For example, TLR9 has been previously implicated in Ad vector double-stranded DNA (dsDNA) genome sensing, and TLR9 requires endosomal acidification for its function (14,
32, 35–37). Additionally, late endosomal lysis and cathepsin B release into the cytosol activate the NALP3 inflammasome, and cathepsin B activity is required for this process (38). The sensitiv-ity of Ad vector innate stimulation to both the cathepsin B inhib-itor Ca074-Me and the pancathepsin inhibinhib-itor Z-FA for an ex-tended period following Ad endocytosis is temporally consistent with our observation that Ad35 and Ad26 vectors potentially be-gin exiting the late endosome at 2 to 6 h postinfection (Fig. 1A). Additionally, the fact that Ad vector sensing was sensitive to ca-thepsin inhibitors for up to 6 h following infection indicates that a defect in endosomal TLR processing is unlikely to account for the reduced innate immune stimulation observed. Of note, recent studies have suggested the importance of the cytosolic DNA sen-sor cGAS for innate sensing of Ad5 in murine cells (39). The po-tential influence of cGAS on Ad innate immune sensing, as well as putative differences between Ad5, Ad35, and Ad26 on cGAS stim-ulation, will therefore be of interest for further understanding of Ad innate immune sensing. Importantly, our data confirm prior observations of the importance of acid-dependent innate immune sensors and inflammasome activation and extend these findings by suggesting that differential trafficking of several Ad serotypes currently in preclinical and clinical development to late
endo-somal compartments contributes to their innate immune-stimu-latory phenotypes. Importantly, these data also suggest, in agree-ment with previous studies showing Ad vector innate sensing is redundantin vivo, that several pattern recognition receptors are likely important for Ad vector sensing (13,16,40).
Our studies utilized A549 cells and human PBMC for the anal-ysis of Ad vector trafficking and human PBMC for the assessment of innate immune phenotypes (12,18,22,23,41). A limitation of studying vector trafficking in A549 cells is that they are more ho-mogeneous than the diverse cellular subsets in PBMC. We previ-ously showed that the innate immune cytokines and chemokines elicited by Ad vectors are dependent on several PBMC subsets (12). Due to the bulk nature of the measurements of innate stim-ulation performed on PBMC, it is possible that inhibition of par-ticular cellular subsets may be responsible for the results observed. As such, the chemical inhibitor data shown above cannot formally differentiate between induction of common sensing pathways in all PBMC subsets and triggering of different innate sensing path-ways in different PBMC subsets. Nevertheless, our results strongly suggest that important innate signaling pathways likely involve late endosomes.
Both the fiber and other capsid components appear to contrib-ute to the innate stimulatory properties of Ad vectors, suggesting roles for receptor binding proteins, as well as acid-dependent cap-sid protease components, in determining Ad innate immune-stimulatory phenotypes (12). Interestingly, CD46 binding by the fiber protein appears to be sufficient to confer late endosomal trafficking on Ad vectors (22). These data suggest that Ad innate immune stimulation phenotypes observed in previous studies
FIG 8Proposed model of differential innate immune stimulation elicited by Ad5, Ad35, and Ad26. Ad5, Ad35, and Ad26 virions enter the cell through an EEA1-positive early endosomal compartment. Upon entry into this location, the majority of Ad5 rapidly exits the endocytic pathway into the cytosol, where it subsequently traffics to the nucleus by 60 min postinfection. Ad35 and Ad26, as well as a small subset of Ad5 virions, accumulate in the late endosomal compartment 2 to 6 h following infection. At approximately 2 to 6 h postinfection, late endosomal Ad virions exit the late endosome and traffic to the nucleus. Accumulation of Ad virions in late endosomes and their release into the cytosol stimulate both an endosomal acidification-dependent and a cathepsin-dependent sensor.
Adenovirus Vector Trafficking and Innate Immunity
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.123.462.66.320.2]may be reflective of increased late endosomal trafficking conferred by the fiber rather than primary receptor usage (15,17,37). Fur-thermore, the increased innate stimulatory capacity of Ad5f35 rel-ative to Ad35k5, despite their roughly equivalent abilities to access late endosomal compartments, suggests an active role of CD46 in influencing Ad vector innate immune stimulation, a possibility that warrants further investigation.
In summary, our data suggest a potential mechanism by which Ad5, Ad35, and Ad26 vectors differentially stimulate innate im-munity. These vectors trigger similar innate immune pathways, but the extent of stimulation is driven by differential intracellular trafficking to late endosomes, resulting in profound differences in innate immunity. These results have important implications for the development of vaccine and gene therapy vectors and could lead to strategies for rational vector design that specifically tune desired innate immune phenotypes.
ACKNOWLEDGMENTS
We thank S. Blackmore, E. Borducci, A. McNally, L. Parenteau, J. Smith, and K. Stanley for technical assistance. We thank D. Cureton, L. Ang, Y. Zheng, and the Imaging Core at BIDMC for confocal microscopy exper-tise and advice.
We acknowledge support from the National Institutes of Health (AI078526 and AI096040), the Bill and Melinda Gates Foundation (OPP1033091 and OPP1040741), and the Ragon Institute of MGH, MIT, and Harvard.
REFERENCES
1.Abbink P, Lemckert AAC, Ewald BA, Lynch DM, Denholtz M, Smits S, Holterman L, Damen I, Vogels R, Thorner AR, Brien KLO, Carville A, Mansfield KG, Goudsmit J, Havenga MJE, Barouch DH.2007. Com-parative seroprevalence and immunogenicity of six rare serotype recom-binant adenovirus vaccine vectors from subgroups B and D. J. Virol.81: 4654 – 4663.http://dx.doi.org/10.1128/JVI.02696-06.
2.Barouch DH, Kik SV, Weverling GJ, Dilan R, King SL, Maxfield LF, Clark S, Ng’ang’a D, Brandariz KL, Abbink P, Sinangil F, de Bruyn G, Gray GE, Roux S, Bekker L-G, Dilraj A, Kibuuka H, Robb ML, Michael NL, Anzala O, Amornkul PN, Gilmour J, Hural J, Buchbinder SP, Seaman MS, Dolin R, Baden LR, Carville A, Mansfield KG, Pau MG, Goudsmit J.2011. International seroepidemiology of adenovirus sero-types 5, 26, 35, and 48 in pediatric and adult populations. Vaccine29: 5203–5209.http://dx.doi.org/10.1016/j.vaccine.2011.05.025.
3.Santra S, Sun Y, Korioth-Schmitz B, Fitzgerald J, Charbonneau C, Santos G, Seaman MS, Ratcliffe SJ, Montefiori DC, Nabel GJ, Ertl HCJ, Letvin NL. 2009. Heterologous prime/boost immunizations of rhesus monkeys using chimpanzee adenovirus vectors. Vaccine27:5837–5845.
http://dx.doi.org/10.1016/j.vaccine.2009.07.050.
4.Liu J, O’Brien KL, Lynch DM, Simmons NL, La Porte A, Riggs AM, Abbink P, Coffey RT, Grandpre LE, Seaman MS, Landucci G, Forthal DN, Montefiori DC, Carville A, Mansfield KG, Havenga MJ, Pau MG, Goudsmit J, Barouch DH.2009. Immune control of an SIV challenge by a T-cell-based vaccine in rhesus monkeys. Nature457:87–91.http://dx.doi .org/10.1038/nature07469.
5.Penaloza-MacMaster P, Provine NM, Ra J, Borducchi EN, McNally A, Simmons NL, Iampietro MJ, Barouch DH.2013. Alternative serotype adenovirus vaccine vectors elicit memory T cells with enhanced anamnes-tic capacity compared to Ad5 vectors. J. Virol.87:1373–1384.http://dx.doi .org/10.1128/JVI.02058-12.
6.Barouch DH, Liu J, Li H, Maxfield LF, Abbink P, Lynch DM, Iampietro MJ, SanMiguel A, Seaman MS, Ferrari G, Forthal DN, Ourmanov I, Hirsch VM, Carville A, Mansfield KG, Stablein D, Pau MG, Schuite-maker H, Sadoff JC, Billings EM, Rao M, Robb ML, Kim JH, Marovich MA, Goudsmit J, Michael NL.2012. Vaccine protection against acquisi-tion of neutralizaacquisi-tion-resistant SIV challenges in rhesus monkeys. Nature 482:89 –93.http://dx.doi.org/10.1038/nature10766.
7.Letvin NL, Rao SS, Montefiori DC, Seaman MS, Sun Y, Lim S-Y, Yeh WW, Asmal M, Gelman RS, Shen L, Whitney JB, Seoighe C, Lacerda M, Keating S, Norris PJ, Hudgens MG, Gilbert PB, Buzby AP, Mach LV,
Zhang J, Balachandran H, Shaw GM, Schmidt SD, Todd J-P, Dodson A, Mascola JR, Nabel GJ.2011. Immune and genetic correlates of vaccine protection against mucosal infection by SIV in monkeys. Sci. Transl. Med. 3:81ra36.http://dx.doi.org/10.1126/scitranslmed.3002351.
8.Kasturi SP, Skountzou I, Albrecht RA, Koutsonanos D, Hua T, Nakaya HI, Ravindran R, Stewart S, Alam M, Kwissa M, Villinger F, Murthy N, Steel J, Jacob J, Hogan RJ, García-Sastre A, Compans R, Pulendran B. 2011. Programming the magnitude and persistence of antibody responses with innate immunity. Nature470:543–547.http://dx.doi.org/10.1038 /nature09737.
9.Manicassamy S, Pulendran B.2009. Modulation of adaptive immunity with Toll-like receptors. Semin. Immunol.21:185–193.http://dx.doi.org /10.1016/j.smim.2009.05.005.
10. Pulendran B.2004. Modulating TH1/TH2 responses with microbes, den-dritic cells, and pathogen recognition receptors. Immunol. Res.29:187– 196.http://dx.doi.org/10.1385/IR:29:1-3:187.
11. Querec TD, Akondy RS, Lee EK, Cao W, Nakaya HI, Teuwen D, Pirani A, Gernert K, Deng J, Marzolf B, Kennedy K, Wu H, Bennouna S, Oluoch H, Miller J, Vencio RZ, Mulligan M, Aderem A, Ahmed R, Pulendran B.2009. Systems biology approach predicts immunogenicity of the yellow fever vaccine in humans. Nat. Immunol.10:116 –125.http: //dx.doi.org/10.1038/ni.1688.
12. Teigler JE, Iampietro MJ, Barouch DH.2012. Vaccination with adeno-virus serotypes 35, 26, and 48 elicits higher levels of innate cytokine re-sponses than adenovirus serotype 5 in rhesus monkeys. J. Virol.86:9590 – 9598.http://dx.doi.org/10.1128/JVI.00740-12.
13. Johnson MJ, Petrovas C, Yamamoto T, Lindsay RWB, Loré K, Gall JGD, Gostick E, Lefebvre F, Cameron MJ, Price D, Haddad E, Sekaly R-P, Seder R, Koup R.2012. Type I IFN induced by adenovirus serotypes 28 and 35 has multiple effects on T cell immunogenicity. J. Imunol.188: 6109 – 6118.http://dx.doi.org/10.4049/jimmunol.1103717.
14. Iacobelli-Martinez M, Nemerow GR. 2007. Preferential activation of Toll-like receptor nine by CD46-utilizing adenoviruses. J. Virol.81:1305– 1312.http://dx.doi.org/10.1128/JVI.01926-06.
15. Barlan AU, Danthi P, Wiethoff CM.2011. Lysosomal localization and mechanism of membrane penetration influence nonenveloped virus acti-vation of the NLRP3 inflammasome. Virology412:306 –314.http://dx.doi .org/10.1016/j.virol.2011.01.019.
16. Barlan U, Griffin TM, McGuire K, Wiethoff CM. 2011. Adenovirus membrane penetration activates the NLRP3 inflammasome. J. Virol.85: 146 –155.http://dx.doi.org/10.1128/JVI.01265-10.
17. Alba R, Bradshaw AC, Coughlan L, Denby L, McDonald R, Wadding-ton SN, Buckley SMK, Greig JA, Parker AL, Miller AM, Wang H, Lieber A, van Rooijen N, McVey JH, Nicklin SA, Baker AH.2010. Biodistri-bution and retargeting of FX-binding ablated adenovirus serotype 5 vec-tors. Blood 116:2656 –2664. http://dx.doi.org/10.1182/blood-2009-12 -260026.
18. Gastaldelli M, Imelli N, Boucke K, Amstutz B, Meier O, Greber UF. 2008. Infectious adenovirus type 2 transport through early but not late endosomes. Traffic9:2265–2278.http://dx.doi.org/10.1111/j.1600-0854 .2008.00835.x.
19. Kälin S, Amstutz B, Gastaldelli M, Wolfrum N, Boucke K, Havenga M, DiGennaro F, Liska N, Hemmi S, Greber UF.2010. Macropinocytotic uptake and infection of human epithelial cells with species B2 adenovirus type 35. J. Virol.84:5336 –5350.http://dx.doi.org/10.1128/JVI.02494-09. 20. Leopold PL, Ferris B, Grinberg I, Worgall S, Hackett NR, Crystal RG.
1998. Fluorescent virions: dynamic tracking of the pathway of adenoviral gene transfer vectors in living cells. Hum. Gene Ther.9:367–378.http://dx .doi.org/10.1089/hum.1998.9.3-367.
21. Meier O, Boucke K, Hammer SV, Keller S, Stidwill RP, Hemmi S, Greber UF.2002. Adenovirus triggers macropinocytosis and endosomal leakage together with its clathrin-mediated uptake. J. Cell Biol.158:1119 – 1131.http://dx.doi.org/10.1083/jcb.200112067.
22. Miyazawa N, Leopold PL, Hackett NR, Ferris B, Worgall S, Falckped-ersen E, Crystal RG.1999. Fiber swap between adenovirus subgroups B and C alters intracellular trafficking of adenovirus gene transfer vectors. J. Virol.73:6056 – 6065.
23. Miyazawa N, Crystal RG, Leopold PL.2001. Adenovirus serotype 7 retention in a late endosomal compartment prior to cytosol escape is mod-ulated by fiber protein. J. Virol.75:1387–1400.http://dx.doi.org/10.1128 /JVI.75.3.1387-1400.2001.
24. Shayakhmetov DM, Li Z, Ternovoi V, Gaggar A, Gharwan H, Lieber A. 2003. The interaction between the fiber knob domain and the cellular
Teigler et al.
on November 7, 2019 by guest
http://jvi.asm.org/
attachment receptor determines the intracellular trafficking route of ad-enoviruses. J. Virol. 77:3712–3723. http://dx.doi.org/10.1128/JVI.77.6 .3712-3723.2003.
25. Nanda A, Lynch DM, Goudsmit J, Angelique A, Lemckert C, Ewald BA, Sumida SM, Diana M, Abbink P, Kishko MG, Gorgone DA, Lifton MA, Shen L, Carville A, Keith G, Havenga MJE, Barouch DH, Lemckert AAC, Truitt DM, Mansfield KG.2005. Immunogenicity of recombinant fiber-Chimeric adenovirus serotype 35 vector-based vaccines in mice and rhesus monkeys. J. Virol.79:14161–14168.http://dx.doi.org/10.1128/JVI .79.22.14161-14168.2005.
26. Bennett S, Breit SN.1994. Variables in the isolation and culture of human monocytes that are of particular relevance to studies of HIV. J. Leukoc. Biol.56:236 –240.
27. Chen JW, Murphy TL, Willingham MC, Pastan I, August JT.1985. Identification of two lysosomal membrane glycoproteins. J. Cell Biol.101: 85–95.http://dx.doi.org/10.1083/jcb.101.1.85.
28. Griffiths G, Hoflack B, Simons K, Mellman I, Kornfeld S.1988. The mannose 6-phosphate receptor and the biogenesis of lysosomes. Cell52: 329 –341.http://dx.doi.org/10.1016/S0092-8674(88)80026-6.
29. Mu F-T, Callaghan JM, Steele-Mortimer O, Stenmark H, Parton RG, Campbell PL, McCluskey J, Yeo J-P, Tock EP, Toh B-H.1995. EEA1, an early endosome-associated protein. J. Biol. Chem.270:13503–13511.http: //dx.doi.org/10.1074/jbc.270.22.13503.
30. Chandra J, McCormick TS, Imamura Y, Mukherjee PK, Ghannoum MA.2007. Interaction of Candida albicans with adherent human periph-eral blood mononuclear cells increases C. albicans biofilm formation and results in differential expression of pro- and anti-inflammatory cytokines. Infect. Immun.75:2612–2620.http://dx.doi.org/10.1128/IAI.01841-06. 31. Lund JM, Alexopoulou L, Sato A, Karow M, Adams NC, Gale NW,
Iwasaki A, Flavell RA.2004. Recognition of single-stranded RNA viruses by Toll-like receptor 7. Proc. Natl. Acad. Sci. U. S. A.101:5598 –5603.
http://dx.doi.org/10.1073/pnas.0400937101.
32. Lund J, Sato A, Akira S, Medzhitov R, Iwasaki A. 2003. Toll-like receptor 9-mediated recognition of Herpes simplex virus-2 by plasmacy-toid dendritic cells. J. Exp. Med.198:513–520.http://dx.doi.org/10.1084 /jem.20030162.
33. Turk V, Stoka V, Vasiljeva O, Renko M, Sun T, Turk B, Turk D.2012. Cysteine cathepsins: from structure, function and regulation to new
fron-tiers. Biochim. Biophys. Acta 1824:68 – 88.http://dx.doi.org/10.1016/j .bbapap.2011.10.002.
34. McIlwain, DR, Berger T, Mak TW.2013. Caspase functions in cell death and disease. Cold Spring Harb. Perspect. Biol.5:a008656.http://dx.doi .org/10.1101/cshperspect.a008656.
35. Pahl JHW, Verhoeven DHJ, Kwappenberg KMC, Vellinga J, Lankester AC, van Tol MJD, Schilham MW.2012. Adenovirus type 35, but not type 5, stimulates NK cell activation via plasmacytoid dendritic cells and TLR9 signaling. Mol. Immunol.51:91–100.http://dx.doi.org/10.1016/j .molimm.2012.02.119.
36. Zhu J, Huang X, Yang Y.2007. Innate immune response to adenoviral vectors is mediated by both Toll-like receptor-dependent and -indepen-dent pathways. J. Virol. 81:3170 –3180. http://dx.doi.org/10.1128/JVI .02192-06.
37. Perreau M, Welles HC, Pellaton C, Gjoksi B, Potin L, Martin R, Harari A, Bett A, Casimiro D, Gall J, Barouch DH, Kremer EJ, Pantaleo G. 2012. The number of Toll-like receptor 9-agonist motifs in the adenovirus genome correlates with induction of dendritic cell maturation by adeno-virus immune complexes. J. Virol.86:6279 – 6285.http://dx.doi.org/10 .1128/JVI.00123-12.
38. Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL, Fitzgerald KA, Latz E.2008. Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat. Im-munol.9:847– 856.http://dx.doi.org/10.1038/ni.1631.
39. Lam E, Stein S, Falck-Pedersen E.2013. Adenovirus detection by the cGAS/STING/TBK1 DNA sensing cascade. J. Virol.88:974 –981.http://dx .doi.org/10.1128/JVI.02702-13.
40. Rhee EG, Blattman JN, Kasturi SP, Kelley RP, Kaufman DR, Lynch DM, La Porte A, Simmons NL, Clark SL, Pulendran B, Greenberg PD, Barouch DH.2011. Multiple innate immune pathways contribute to the immunogenicity of recombinant adenovirus vaccine vectors. J. Virol.85: 315–323.http://dx.doi.org/10.1128/JVI.01597-10.
41. Greber UF, Willetts M, Webster P, Helenius A.1993. Stepwise disman-tling of adenovirus 2 during entry into cells. Cell75:477– 486.http://dx .doi.org/10.1016/0092-8674(93)90382-Z.
42. Manders EMM, Verbeek FJ, Aten JA.1993. Measurement of colocaliza-tion of objects in dual-colour confocal images. J. Microsc.169:375–382.
http://dx.doi.org/10.1111/j.1365-2818.1993.tb03313.x.
Adenovirus Vector Trafficking and Innate Immunity