Analysis of Select Herpes Simplex Virus 1 (HSV-1) Proteins for
Restriction of Human Immunodeficiency Virus Type 1 (HIV-1):
HSV-1 gM Protein Potently Restricts HIV-1 by Preventing
Intracellular Transport and Processing of Env gp160
Sachith Polpitiya Arachchige,
aWyatt Henke,
aAnkita Pramanik,
aMaria Kalamvoki,
aEdward B. Stephens
aaDepartment of Microbiology, Molecular Genetics and Immunology, University of Kansas Medical Center, Kansas City, Kansas, USA
ABSTRACT
Virus-encoded proteins that impair or shut down specific host cell
func-tions during replication can be used as probes to identify potential
proteins/path-ways used in the replication of viruses from other families. We screened nine
pro-teins from herpes simplex virus 1 (HSV-1) for the ability to enhance or restrict
human immunodeficiency virus type 1 (HIV-1) replication. We show that several
HSV-1 proteins (glycoprotein M [gM], US3, and UL24) potently restricted the
replica-tion of HIV-1. Unlike UL24 and US3, which reduced viral protein synthesis, we
ob-served that gM restriction of HIV-1 occurred through interference with the
process-ing and transport of gp160, resultprocess-ing in a significantly reduced level of mature
gp120/gp41 released from cells. Finally, we show that an HSV-1 gM mutant lacking
the majority of the C-terminal domain (HA-gM[Δ345-473]) restricted neither gp160
processing nor the release of infectious virus. These studies identify proteins from
heterologous viruses that can restrict viruses through novel pathways.
IMPORTANCE
HIV-1 infection of humans results in AIDS, characterized by the loss of
CD4
⫹T cells and increased susceptibility to opportunistic infections. Both HIV-1 and
HSV-1 can infect astrocytes and microglia of the central nervous system (CNS). Thus,
the identification of HSV-1 proteins that directly restrict HIV-1 or interfere with
path-ways required for HIV-1 replication could lead to novel antiretroviral strategies. The
results of this study show that select viral proteins from HSV-1 can potently restrict
HIV-1. Further, our results indicate that the gM protein of HSV-1 restricts HIV-1
through a novel pathway by interfering with the processing of gp160 and its
incor-poration into virus maturing from the cell.
KEYWORDS
UL24, US3, gM, glycoprotein transport, herpes simplex virus, human
immunodeficiency virus, restriction
D
ominant-negative (DN) mutants have been used extensively to study the
impor-tance of viral proteins in replication and to decipher virus-host interactions. The
viral proteins from heterologous viruses that act in a dominant-negative fashion to
decrease the replication or release of infectious virus of another viral family may
provide alternative antiviral strategies. Herpes simplex virus 1 (HSV-1) is a major
opportunistic pathogen in human immunodeficiency virus type 1 (HIV-1)-infected
patients that may be a cofactor by interacting with HIV-1 at the cellular and/or
molecular level. HSV-1 is a highly cytopathic virus that is maintained in a latent state in
humans (1–3). The HSV-1 genome has more than 80 open reading frames (ORFs) that
encode proteins, including transactivators of transcription, DNA replication proteins,
RNA binding proteins, proteases, kinases, and structural components (viral
glycopro-teins and capsid proglycopro-teins). Due to the complex replication scheme of HSV-1, which
Received28 August 2017Accepted16
October 2017
Accepted manuscript posted online1
November 2017
CitationPolpitiya Arachchige S, Henke W,
Pramanik A, Kalamvoki M, Stephens EB. 2018. Analysis of select herpes simplex virus 1 (HSV-1) proteins for restriction of human immunodeficiency virus type 1 (HIV-1): HSV-1 gM protein potently restricts HIV-1 by preventing intracellular transport and processing of Env gp160. J Virol 92:e01476-17.
https://doi.org/10.1128/JVI.01476-17.
EditorRozanne M. Sandri-Goldin, University of
California, Irvine
Copyright© 2018 American Society for
Microbiology.All Rights Reserved. Address correspondence to Edward B. Stephens, [email protected].
crossm
on November 6, 2019 by guest
http://jvi.asm.org/
involves the cytoplasm, nucleus, and membrane organelles, HSV-1 encodes a unique
set of proteins that could act as dominant-negative proteins to block the replication or
release of a heterologous virus.
HIV-1 encodes proteins that facilitate replication and integration of the viral genome
into the host cell chromosome (i.e., reverse transcriptase and integrase) or viral
tran-scription and RNA export (i.e., Tat and Rev) (4) and that interact with host cell factors
that would normally restrict viral replication or promote immune responses against the
virus (i.e., Vpu, Nef, Vif, and Vpr) (5, 6). While many molecular aspects of HIV-1
replication have been deciphered, the identification of novel cellular proteins/pathways
that affect HIV-1 replication continues to be explored.
While HIV-1 and HSV-1 generally do not infect the same target cells (HIV-1, CD4
⫹T
cells and macrophages; HSV-1, epithelial cells and neurons), one exception is the central
nervous system (CNS). Both viruses are neurotropic, with HSV-1 the most common
cause of sporadic encephalitis in the United States and HIV-1 capable of causing overt
encephalitis and HIV-1-associated dementia (HAD) (7, 61). Despite current treatments
for HIV-1 (combined antiretroviral therapy [cART]), approximately 50% of infected
patients continue to develop HIV-1-associated neurocognitive disorders (HAND) (8–10).
Both HIV-1 and HSV-1 infect astrocytes and microglia of the CNS (11–18). Astrocytes are
the most abundant cells of the CNS and have several functions, including biochemical
support of brain endothelial cells essential to maintaining the blood-brain barrier,
production of neurotrophic immune-modulatory factors, neuronal homeostasis, and
repair and scarring following CNS trauma (19). Microglia account for 1 to 15% of all cells
found within the brain. As the resident macrophages, they function as the first and
main form of active immune defense in the CNS (20). Thus, disruption of the function
of these types of cells and release of immune modulators toxic to neurons by viral
infection can have detrimental consequences for the CNS.
Here, we screened nine HSV-1 proteins for the ability to restrict the replication of
HIV-1. Our data show differential effects of these proteins on HIV-1 replication, with
three HSV-1 proteins, US3, UL24, and gM, potently restricting HIV-1 replication. We have
further characterized the novel mechanism through which HSV-1 gM restricts HIV-1
replication.
RESULTS
Identification of HSV-1 proteins that potently restrict HIV-1.
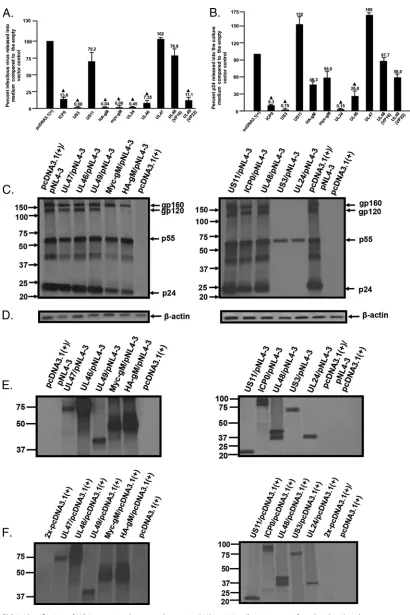
We analyzed nine
HSV-1 proteins for the ability to restrict HIV-1. These proteins were ICP0, VP16, VP22,
US3, US11, gM, UL24, UL46, and UL47. The proteins have different functions in the
HSV-1 life cycle, which are listed in Table 1. We first analyzed the expression of these
proteins to ensure that they were expressed well in 293 cells. Transfection of 293 cells
with each plasmid construct and analysis of expression at 48 h posttransfection using
anti-myc, anti-Flag, or protein-specific antibodies and Western blotting showed that all
nine proteins were expressed (Fig. 1). We then constructed a pcDNA3.1(
⫹
) vector
expressing a human codon-optimized glycoprotein M (gM) with a hemagglutinin (HA)
tag at the N terminus that was expressed well in 293 cells for further experiments (Fig.
1E). We next cotransfected 293 cells with each of the above-mentioned vectors and a
vector with the complete genome of HIV-1 strain NL4-3 (pNL4-3). At 48 h
posttrans-fection, the levels of infectious virus and p24 released into the culture medium were
determined. The results indicated that three proteins, VP16, UL47, and US11, had no
significant effect on HIV-1 replication and release of infectious virus (Fig. 2A). Three
other HSV-1 proteins, ICP0, UL46, and VP22, resulted in an intermediate level of
restriction (
⬃
7 to 14% that of the control) (Fig. 2A). Finally, three HSV-1 proteins, US3,
UL24, and gM, restricted the release of infectious virus with averages of 0.08, 0.45, and
0.04% of the pcDNA3.1(
⫹
) control restriction, respectively (Fig. 2A). HA-gM and myc-gM
restricted HIV-1 at equivalent levels. Viability assays using the Zombie Aqua fixable
assay kit (BioLegend) revealed that over 98% of the cells transfected with HA-gM,
myc-UL24, and myc-US3 were viable at 48 h (data not shown). The levels of p24
released from cultures generally correlated with levels of infectious virus released. US3
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 2
on November 6, 2019 by guest
http://jvi.asm.org/
and UL24 released p24 at levels that were 0.75 and 0.15%, respectively, of the pcDNA3.1
control level (Fig. 2B). In contrast, the levels of p24 released from cells transfected with
the vectors expressing HA-gM and pNL4-3 were 46.3% of the control level (Fig. 2B).
These discordant results suggested that HSV-1 gM restriction of HIV-1 might be due to
interaction with a specific HIV-1 protein. We next analyzed the expression of HIV-1
proteins in the presence of the individual HSV-1 proteins. 293 cells were cotransfected
as described above and at 24 h starved for methionine/cysteine and radiolabeled for 6
h. Cell lysates were prepared, and aliquots were used to immunoprecipitate HIV-1
proteins,

-actin, and individual HSV-1 proteins (Fig. 2C to E). Companion cultures were
cotransfected with pcDNA3.1(
⫹
) and with vectors expressing each HSV-1 protein for
comparative purposes (Fig. 2F). Our results indicated that, with the exceptions of gM,
US3, and UL24, the synthesis of HIV-1 proteins was similar to that of the pNL4-3/
pcDNA3.1(
⫹
) control. Additionally, expression of the HSV-1 proteins in the presence of
pNL4-3 was not significantly different from that of companion cultures expressing only
the HSV-1 protein, indicating that the HIV-1 proteins did not result in degradation of
TABLE 1HSV-1 gene products, classes of transcripts, functions, and locations in HSV-1-infected cells
Gene Protein Transcript class Function Location HIV-1 restrictiona
RL2 ICP0 Alpha (␣) Promiscuous transactivator; disrupts DNA repressor complexes; E3 ligase; blocks innate immunity
Nucleus, cytoplasm ⫹
UL10 gM Gamma (␥2) Efficient viral entry; regulates capsid envelopment; redirects surface proteins totrans-Golgi
Nucleolus,trans-Golgi ⫹⫹⫹
UL24 UL24 Gamma (␥1) Nucleolar dispersal of host proteins; nuclear egress; polykaryocyte formation in the absence of UL24
Nucleus; membrane association
⫹⫹⫹
UL46 VP11/12 Gamma (␥1) Tegument; modulates VP16 transactivation function; PI3Kb/AKT
activation; blocks innate immunity through STING
Nucleus, cytoplasm ⫹⫹
UL47 VP13/14 Gamma (␥1) Tegument; RNA binding protein; modulates the activity of Vhs; modulates VP16 transactivation function
Shuttles between nucleus and cytoplasm
⫺
UL48 VP16 Gamma (␥) Tegument; induces␣genes; virion assembly; together with VP22 blocks Vhs; interacts with gH
Nucleus, cytoplasm ⫺
UL49 VP22 Gamma (␥) Tegument; with VP16 neutralizes Vhs; binds RNA; chromatin remodeling
Nucleus, cytoplasm ⫹
US3 US3 Beta () Tegument; serine/threonine protein kinase (PKA-like) Nucleus, cytoplasm ⫹⫹⫹ US11 US11 Gamma (␥2) Tegument; RNA binding protein; antagonizes PKR; inhibits
RIG-I and MDA-5; capsid egress
Nucleolus, cytoplasm ⫺
aLevel of HIV-1 restriction:⫺, 0 to 30% restriction;⫹, 90 to 31% restriction;⫹⫹, 91 to 99% restriction;⫹⫹⫹,⬎99% restriction. bPI3K, phosphatidylinositol 3-kinase.
FIG 1Expression of HSV proteins in 293 cells. 293 cells were transfected with vectors expressing different HSV-1 proteins or the empty vector. At 48 h, cell lysates were prepared and processed for Western blotting. Proteins were detected using antibodies to either myc- or Flag-tagged HSV-1 proteins. (A) Western blot showing expression of myc-tagged HSV-1 proteins. (B) Western blot showing expression of Flag-tagged HSV-1 proteins. (C) Western blot showing expression of ICP0. (D) Western blot showing expression of VP16. (E) Western blot showing expression of myc-gM and HA-gM. The gels at the bottom show the loading controls (-actin) for the Western blots.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:3.585.42.438.511.681.2]FIG 2Identification of HSV-1 proteins that potently restricted HIV-1. 293 cells were cotransfected with either the empty-vector plasmid or plasmids expressing each HSV-1 protein (ICP0, US3, US11, myc-gM, HA-gM, UL24, UL46, UL47, VP16, or VP22) and a plasmid with the HIV-1 genome (pNL4-3). At 48 h, the culture supernatants were collected. (A and B) The levels of infectious virus released into the culture supernatants were determined using TZM.bl cell assays, and p24 levels in the culture supernatants were determined using p24 antigen capture assays. (A) Levels of infectious virus released into the culture medium from cells cotransfected with the pNL4-3 genome and plasmids expressing the different HSV-1 proteins. (B) Levels of p24 protein produced
(Continued on next page)
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 4
on November 6, 2019 by guest
http://jvi.asm.org/
[image:4.585.39.449.66.681.2]the HSV-1 proteins tested. Taken together, these results indicate that ectopic
expres-sion of select HSV-1 proteins potently restricted the production of infectious HIV-1.
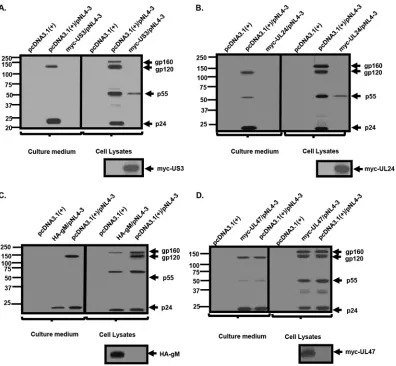
Processing and release of viral proteins in the presence of US3, UL24, and gM.
As US3, UL24, and gM restricted the production of infectious HIV-1 to the greatest
extent, we examined the processing and release of the HIV-1 proteins in cells
express-ing these HSV-1 proteins. 293 cells were cotransfected with either the empty
pcDNA3.1(
⫹
) vector or one expressing HA-US3, HA-UL24, HA-gM, or UL47 and with
pNL4-3. The transfected cells were starved for 2 h and radiolabeled with [
35S]methio-nine/cysteine. The viral proteins immunoprecipitated from the culture medium and
from cell lysates at 16 h postlabeling. Cells cotransfected with pcDNA3.1(
⫹
) and pNL4-3
expressed gp160, gp120, p55 Gag, and p24 in the cell lysates, while gp120 and p24
were readily detected in the supernatants from culture medium (Fig. 3A to D). In cells
transfected with HA-US3 or HA-UL24 and pNL4-3, the detection of viral proteins by
immunoprecipitation was significantly decreased, with only minor amounts of p55
detected in the cell lysates and no viral proteins detected in the supernatants from
culture medium (Fig. 3A and B). However, in cells cotransfected with HA-gM and
pNL4-3, p24 was readily detectable in cell lysates and supernatants from culture
medium, although at reduced levels compared to the pcDNA3.1(
⫹
)/pNL4-3 control
(Fig. 3C). Additionally, in HA-gM/pNL4-3 cultures, while the gp160 precursor was
detected in cell lysates, no gp120 was observed in cell lysates, which was also reflected
in the levels of gp120 released from cells (Fig. 3C). Finally, in myc-UL47/NL4-3 cultures,
p24 and gp120 were released into the culture medium at levels similar to those of the
pcDNA3.1(
⫹
)/pNL4-3 control (Fig. 3D). These results correlated well with the p24
assays.
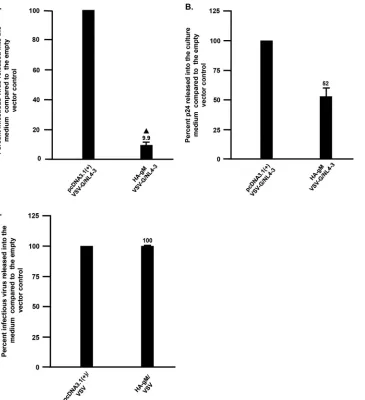
Inoculation of HA-gM-expressing cultures with infectious virus restricts HIV-1,
but not VSV.
To determine if gM would restrict infectious HIV-1, we generated
vesicular stomatitis virus G protein (VSV-G)-pseudotyped virus. 293 cells were
trans-fected with either the empty pcDNA3.1(
⫹
) vector or one expressing HA-gM. The cells
were then inoculated with VSV-G/NL4-3 (multiplicity of infection [MOI]
⫽
0.01), and the
levels of infectious virus released were determined using TZM.bl cells (Fig. 4A) while
those of p24 were determined by antigen capture assays. The results indicated that
the levels of infectious virus released were significantly decreased (9.9% those of the
empty-vector control). We also examined whether VSV replication was restricted in the
presence of HA-gM. 293 cells were transfected with either the empty vector or one
expressing HA-gM, and at 24 h posttransfection, the cells were inoculated with VSV. At
24 h postinoculation, the virus-containing culture medium was collected and titrated
on 293 cells. Our results showed that the presence of HA-gM did not restrict VSV
replication, indicating the specificity of the HA-gM-mediated HIV-1 restriction (Fig. 4C).
gM from HSV-2 also restricts HIV-1.
We also analyzed the ability of the gM protein
from HSV-2 to restrict HIV-1. We chose HSV-2 gM for sequence similarity to HSV-1
(
⬃
80% at the amino acid level). Both proteins were expressed well in 293, as
deter-mined by Western blotting (Fig. 5A). The slight decrease in relative molecular weight
(
M
r) of myc-tagged gM of HSV-2 (myc-gM[HSV-2]) was due to a 7-amino-acid deletion
FIG 2Legend (Continued)
from cells cotransfected with the pNL4-3 genome and plasmids expressing the different HSV-1 proteins. The experiments were performed at least four times, and statistical differences with the pcDNA3.1(⫹)/HIV-1 control were evaluated using a two-tailed Studentttest, with aPvalue of⬍0.05 (Œ) considered significant. The error bars indicate standard deviations and the numbers the mean levels of infectious virus released compared to the pcDNA3.1(⫹) control. (C to E) Expression of HIV-1 proteins in the presence of HSV-1 proteins. 293 cells were transfected with pcDNA3.1(⫹) or a vector expressing each HSV-1 protein and a plasmid expressing the pNL4-3 viral genome. At 24 h, the cells were starved and radiolabeled with [35S]methionine/cysteine for 6 h, at which time the culture medium was removed and cell lysates were prepared. Equal aliquots of the cell lysates were immuno-precipitated with antibodies against the HIV-1 proteins (C),-actin (D), or HSV-1 proteins (E). (F) Companion cultures from the experiments in panel C to E were cotransfected with the empty vector pcDNA3.1(⫹) and vectors expressing each of the HSV-1 proteins. 293 cells were cotransfected with pcDNA3.1(⫹) and vectors expressing each HSV-1 protein listed above. At 24 h, the cells were starved and radiolabeled with [35S]methionine/cysteine for 6 h. The culture medium was removed, and cell lysates were prepared. Equal aliquots of the cell lysates were immunoprecipitated with appropriate antibodies against the HSV-1 proteins. The numbers to the left indicate the position of the molecular weight markers (in thousands).
on November 6, 2019 by guest
http://jvi.asm.org/
(amino acids 384 to 390) in the C-terminal domain. While the levels of p24 were similar
in cells transfected with HA-gM[HSV-1], myc-gM[HSV-2], and pNL4-3, we observed that
myc-gM[HSV-2] was also capable of significant restriction of HIV-1 replication at levels
similar to those of HA-gM[HSV-1] (Fig. 5B and C).
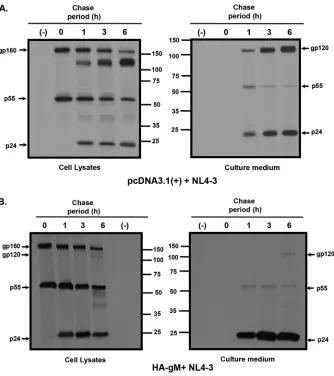
Pulse-chase analyses reveal that HIV-1 gp160 is inefficiently processed and
degraded in the presence of HSV-1 gM.
As the above-described results suggested
that gp160 was inefficiently processed, we determined if expression of HA-gM would
prevent the processing of gp160 in the presence or absence of other viral proteins. In
the first set of experiments, 293 cells were cotransfected with the empty pcDNA3.1(
⫹
)
vector or the vector expressing HA-gM and plasmid pNL4-3. At 48 h posttransfection,
the cells were starved for methionine/cysteine and radiolabeled, and pulse-chase
experiments were performed with chase periods of 0, 1, 3, and 6 h (Fig. 6A and B). The
culture medium was harvested, and cell lysates were prepared as described in Materials
and Methods. HIV-1 proteins were immunoprecipitated using an anti-simian-human
immunodeficiency virus (SHIV) serum and an antibody directed against p24. As shown
FIG 3Processing and release of HIV-1 proteins in the presence of US3, UL24, and gM. 293 cells were cotransfected with the empty pcDNA3.1(⫹) vector or one expressing myc-UL24, myc-US3, HA-gM, or myc-UL47 and a plasmid containing the HIV-1 NL4-3 genome (pNL4-3). At 24 h, the cells were incubated in medium lacking methionine/cysteine for 2 h, followed by radiolabeling the cultures with [35S]methionine/cysteine for 16 h. The culture medium was harvested, cell lysates were prepared as described in Materials and Methods, and HIV-1 Env and Gag proteins were immunoprecipitated with appropriate antibodies. In addition, HSV-1 proteins (Us3, myc-UL24, and UL47) immunoprecipitated with anti-myc and gM immunopre-cipitated with anti-HA are shown below panels A to D. The immunoprecipitates were collected on protein A-Sepharose, washed, and boiled in sample reducing buffer. The proteins were separated on 10% SDS gels and visualized using standard radiographic techniques. (A) myc-US3. (B) Myc-UL24. (C) HA-gM. (D) myc-UL47. Numbers to the left indicate the position of molecular weight markers (in thousands).
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 6
on November 6, 2019 by guest
http://jvi.asm.org/
[image:6.585.43.439.69.435.2]in Fig. 6A, in the presence of the empty vector, gp160 and p55 were detected in the 0-h
chase period, with gp120 and p24 detected at the 1-, 3-, and 6-h chase periods in the
cell lysates. Both gp120 and p24 were detected in the cell culture medium at 1-, 3-, and
6-h chase periods. Pulse-chase analysis of cells transfected with HA-gM and pNL4-3
resulted in gp160 and p55 being detected in the 0-h chase period and little to no gp120
being immunoprecipitated from cell lysates in the 3- and 6-h chase periods (Fig. 6B).
This was also reflected in the HIV-1 proteins immunoprecipitated from the culture
medium. Almost no gp120 was detected in the culture, although p24 was clearly
released into the culture medium (Fig. 6B).
In a second pulse-chase experiment, 293 cells were cotransfected with a vector
expressing HA-gM and the pBal.26 vector. Pulse-chase analyses were performed
simi-FIG 4HSV-1 gM restricts the production of infectious HIV-1 but not VSV. (A) Levels of infectious HIV-1 produced from cells transfected with the empty vector pcDNA3.1(⫹) or one expressing HA-gM and inoculated with VSV-G/NL4-3. 293 cells were transfected with the empty vector pcDNA3.1(⫹) or one expressing HA-gM. At 24 h posttransfection, the cells were inoculated with VSV-G/NL4-3 at an MOI of 0.01. At 72 h posttransfection, the cell culture medium was collected, clarified by low-speed centrifugation, and analyzed for infectious virus using TZM.bl cell assays and for p24 levels using p24 antigen capture assays. (B) Levels of p24 released from transfected/infected cells from panel A. (C) Levels of infectious VSV released into culture supernatants from cells transfected with the empty vector pcDNA3.1(⫹) or one expressing HA-gM and inoculated with VSV. 293 cells were transfected as for panel A, and at 24 h posttransfection were inoculated with VSV (MOI⫽0.01). The virus was collected 24 h after virus inoculation, clarified, and titrated for levels of infectious virus. The results shown represent the means from two experiments. The error bars indicate standard deviations.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:7.585.48.413.70.470.2]larly to those with the virus-transfected cells discussed above, except with chase
periods of 0, 1, 3, and 6 h (Fig. 7). HIV-1 Env was immunoprecipitated using anti-Env
antibodies. Our results indicated that in cells cotransfected with the empty vector
pcDNA3.1(
⫹
) and the vector expressing HIV-1 strain Bal Env, gp160 was synthesized
and processed into gp120 by the 1-h chase period in cell lysates and was barely
detected in the culture medium at the 1-h chase period but was detected at later time
points (Fig. 7A). In contrast, the processing of gp160 into gp120 was not detected in
cells cotransfected with vectors expressing HA-gM and HIV-1 Bal Env glycoprotein until
the 6-h chase period, and the levels of gp120 were reduced in the culture medium in
the 6-h chase period (Fig. 7B).
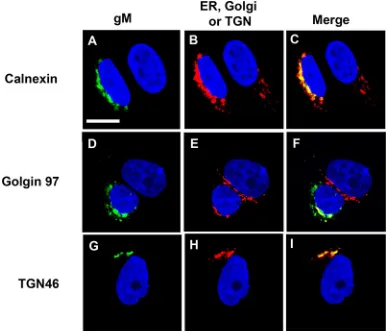
Intracellular localization of Env in the presence of HSV-1 HA-gM protein.
The
intracellular expression of HIV-1 Env was analyzed in the absence or presence of gM. We
first examined the expression of gM in the absence of HIV-1 proteins. COS-7 cells were
transfected with the empty pcDNA3.1(
⫹
) vector or the vector expressing HA-gM and
examined by immunofluorescence for its intracellular localization. At 24 h, the cells
were fixed, permeabilized, and stained with an antibody against the HA tag and
antibodies against calnexin (endoplasmic reticulum [ER]), Golgin 97 (Golgi apparatus),
or TGN46 (
trans
-Golgi network [TGN]). Our results indicated that gM partially
colocal-ized with calnexin, Golgin 97, and TGN46, as was previously reported (Fig. 8A to I). We
did not observe significant amounts of HA-gM on the cell plasma membrane. We next
analyzed the cells cotransfected with either the pcDNA3.1(
⫹
) vector or one expressing
HA-gM and a vector expressing the Bal Env (pBal.26). Our results indicated that after
cotransfection of the vector expressing HA-gM with pBal.26, the level of Env expression
was significantly diminished, with most of the gp160 staining occurring in intracellular
compartments (Fig. 9A to C). In contrast, cotransfection of cells with the empty
pcDNA3.1(
⫹
) vector and the pBal.26 vector resulted in Env that was easily detected
in intracellular compartments and on the cell plasma membrane (Fig. 9G to I). These
results agree with the pulse-chase analysis showing decreased expression with
time.
FIG 5The gM from HSV-2 also restricts HIV-1 replication. 293 cells were cotransfected with the pcDNA3.1(⫹)vector or one expressing HA-gM[HSV-1], myc-gM[HSV-2], or HSV-1 myc-UL47 and a plasmid containing the NL4-3 genome. At 24 h posttransfection with pNL4-3, the culture medium was collected and assayed for release of p24 and for infectious virus using TZM.bl assays. The experiments were performed at least three times, and statistical differences with the pcDNA3.1(⫹)/HIV-1 control were evaluated using a two-tailed Studentttest, with aPvalue of⬍0.05 considered significant. (A) Expression of HA-gM[HSV-1] and myc-gM[HSV-2] proteins in 293 cells. (B) Levels of p24 protein produced from cells cotransfected with pcDNA3.1(⫹), HA-gM[HSV-1], or myc-gM[HSV-2] and a plasmid with the pNL4-3 genome. (C) Levels of infectious virus in the culture medium from cells cotransfected with pcDNA3.1(⫹), HA-gM[HSV-1], or myc-gM[HSV-2] and a plasmid with the pNL4-3 genome. The error bars indicate standard deviations.
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 8
on November 6, 2019 by guest
http://jvi.asm.org/
[image:8.585.42.444.69.277.2]Truncation of gM abrogates restriction of HIV-1.
A current model of the structure
of HSV-1 gM suggests that the protein is a type III membrane protein that spans the
membrane eight times, with both the N and C termini located on the cytoplasmic side
of the membrane (11, 21–24). We determined if removal of the C-terminal domain
would abrogate gM restriction of HIV-1. A mutant was constructed in which the
C-terminal 126 amino acids were deleted from gM (just after the last transmembrane
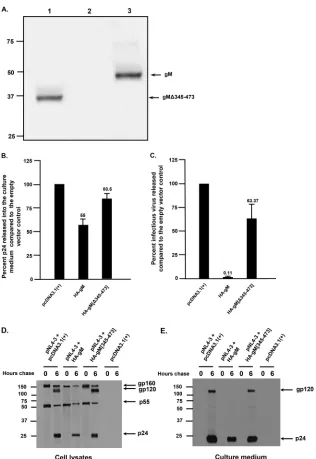
domain). This mutant, HA-gM[Δ345-473], was expressed well in 293 cells (Fig. 10A).
When the culture medium was analyzed for p24, we observed that the levels of p24 in
the medium of cells transfected with HA-gM[Δ345-473] were similar to those with the
empty-vector control (Fig. 10B). When used in restriction assays with pNL4-3, our results
indicated that, unlike the full-length protein, HA-gM[Δ345-473] was unable to restrict
HIV-1 (Fig. 10C). We also determined if the processing of Env was normal in the absence
FIG 6HSV-1 gM inhibits the processing of HIV-1 gp160 and release of gp120. 293 cells were cotransfected with the empty pcDNA3.1(⫹) vector or one expressing HA-gM and pNL4-3. At 48 h posttransfection, the cells were starved for methionine/cysteine and radiolabeled with [35S]methionine/cysteine for 30 min. The radiolabel was removed, and the cells were washed twice and chased in medium containing 100⫻ methionine/cysteine for either 0, 1, 3, or 6 h. The culture medium was removed, and cell lysates were prepared as described in Materials and Methods. HIV-1 Env and Gag proteins were immunoprecipitated with appropriate antibodies, and the immunoprecipitates were collected on protein A-Sepharose. Samples were washed and then boiled in sample reducing buffer. The proteins were separated on 10% SDS gels and visualized using standard radiographic techniques. (A) HIV-1 proteins immunoprecipitated from cell lysates and culture medium from 293 cells cotransfected with pcDNA3.1(⫹) and pNL4-3. (B) HIV-1 proteins immunoprecipitated from cell lysates and culture medium from 293 cells cotransfected with vectors expressing HA-gM and pNL4-3. Numbers in the middle of the panels indicate the position of the molecular weight markers (in thousands).
on November 6, 2019 by guest
http://jvi.asm.org/
[image:9.585.43.377.69.445.2]of other viral proteins and HA-gM[Δ345-473]. 293 cells were cotransfected with either
the empty pcDNA3.1(
⫹
) vector or one expressing HA-gM or HA-gM[Δ345-473] and with
vector pNL4-3. At 48 h, the cells were starved and radiolabeled with [
35S]methionine/
cysteine for 30 min, and the radiolabel was chased in cold excess methionine/cysteine
for 6 h. HIV-1 proteins were immunoprecipitated with anti-SHIV and anti-p24
antibod-FIG 7HSV-1 gM inhibits the processing of HIV-1 gp160 and release of gp120 in the absence of other HIV-1 proteins. 293 cells were cotransfected with the empty pcDNA3.1(⫹) vector or one expressing HA-gM and pBalEnv26, which expresses the Bal strain Env gp160 protein. At 48 h posttransfection, the cells were incubated in medium lacking methionine/cysteine for 2 h, followed by radiolabeling the cultures with [35S]methionine/cysteine for 30 min. The radiolabel was removed, and the cells were washed twice and chased in medium containing 100⫻methionine/cysteine for either 0, 1, 3, or 6 h. The culture medium was removed, and cell lysates were prepared as described in Materials and Methods. HIV-1 Env proteins were immunoprecipitated with an anti-gp160 antibody, and the immunoprecipitates were collected on protein A-Sepharose. Samples were washed and boiled in sample reducing buffer, and proteins were separated on 10% SDS gels and visualized using standard radiographic techniques. (A) HIV-1 Env proteins immunoprecipitated from cell lysates and culture medium from 293 cells cotrans-fected with pcDNA3.1(⫹) and pBalEnv. (B) HIV-1 Env proteins immunoprecipitated from cell lysates and culture medium from 293 cells cotransfected with vectors expressing HA-gM and pBalEnv.
FIG 8Intracellular localization of HA-gM. COS-7 cells on coverslips were transfected with the vector expressing HA-gM. At 24 h posttransfection, the cells were fixed, permeabilized, and blocked as described in Materials and Methods. The cells were reacted with antibodies against calnexin (ER), Golgin 97 (Golgi complex), or TGN46 (trans-Golgi network); washed; and reacted with secondary antibodies as described in Materials and Methods. The cells were then reacted with a rabbit anti-HA-FITC antibody to identify the gM proteins. The cells were washed and counterstained with DAPI for 5 min. All the coverslips were mounted and examined with a Leica TCS SPE confocal microscope with a 63⫻objective and a 2⫻digital zoom using the LAS X software package. A 488-nm filter was used to visualize FITC, a 647-nm filter to visualize Alexa Fluor 647, and a 405-nm filter to visualize DAPI staining. (A,D, and G) Cells stained for HA-gM. (B, E, and H) Cells stained for calnexin (B), Golgin 97 (E), and TGN46 (H). (C, F, and I) Merge of panels A and B (C), of panels D and E (F), and of panels G and H (I). The bar in panel A represents 10m.
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 10
on November 6, 2019 by guest
http://jvi.asm.org/
[image:10.585.72.341.70.234.2] [image:10.585.109.303.459.626.2]ies. The results of the Env immunoprecipitation from cell lysates revealed that gp120
was released from cells cotransfected with pcDNA3.1(
⫹
) and pNL4-3 (Fig. 10D and E).
In contrast, cells cotransfected with vectors expressing HA-gM and pNL4-3 showed very
little gp120 released into the culture medium. Finally, cotransfection of cells with
HA-gM[Δ345-473] and pNL4-3 revealed release of gp120 into the culture medium at
levels similar to those with the empty vector (Fig. 10D and E). We also examined cells
cotransfected with the vector expressing HA-gM[Δ345-473] and the pBal.26 vector by
immunofluorescence (Fig. 9D to F). Our results indicated that expression of Env was
very similar to that observed with cells cotransfected with the empty vector,
pcDNA3.1(
⫹
), and pBal.26 (Fig. 9G to I). Taken together, these results indicate that the
C-terminal domain of gM is required to restrict Env transport and for release of
infectious HIV-1 from cells.
DISCUSSION
Previous studies have shown that herpesviruses can reactivate HIV-1, although this
phenomenon depended on the herpesvirus, experimental model, and cell line used (2,
25–31). Several HSV-1 proteins encoded by immediate-early (IE) genes, such as ICP0
(infected cell polypeptide 0), ICP4, and ICP27, have been shown to transactivate the
HIV-1 long terminal repeat (LTR) (26, 29, 30, 32, 33). More recently, it was shown that
CEM cells chronically infected with either HIV-1 (CEM
HIV) or HSV-1 (CEM
HSV) had
different effects on the replication of HIV-1 or HSV-1 (34). The investigators showed that
superinfection of CEM
HIVcells with HSV-1 resulted in an increase in the release of virus
from the cells as measured by reverse transcriptase activity. This effect was inversely
proportional to the multiplicity of infection used to inoculate cultures. In contrast, HIV-1
superinfection of CEM
HSVcells resulted in a delay in the kinetics of HIV-1 replication,
suggesting a negative influence of HSV-1 on HIV-1 replication (34). To date, no studies
FIG 9Intracellular expression of HIV-1 gp160 in the presence of HA-gM or HA-gM[Δ345-473]. COS-7 cells were cotransfected with either the pcDNA3.1(⫹) vector, one expressing HA-gM, or one expressing HA-gM[Δ345-473] and a plasmid expressing the gp160 (pBal.26). At 24 h posttransfection, the cells were fixed, permeabilized, and blocked as described in Materials and Methods. The cells were then reacted with a goat anti-gp160 antibody to identify the gp160 proteins and with anti-HA to identify gM, washed, and reacted with secondary antibody as described in the text. The coverslips were counterstained with DAPI for 5 min. All the coverslips were mounted and examined with a Leica TCS SPE confocal microscope with a 63⫻objective and a 2⫻digital zoom using the LAS X software package. A 488-nm filter was used to visualize FITC, a 647-nm filter to visualize Alexa Fluor 647, and a 405-nm filter to visualize DAPI staining. (A to C) Cells cotransfected with vectors expressing HA-gM and Bal Env. (D to F) Cells cotransfected with vectors expressing HA-gM[Δ345-473] and Bal Env. (G to I) Cells cotransfected with vector pcDNA3.1(⫹) and Bal Env. (A, D, and G) Cells stained for HA-containing proteins. (B, E, and H) Cells stained for gp160. (C, F, and I) Merge of panels A and B (C), of panels D and E (F), and of panels G and
H (I). The bar in panel A represents 10m.
on November 6, 2019 by guest
http://jvi.asm.org/
[image:11.585.71.338.71.271.2]FIG 10Truncation of gM abrogates restriction of HIV-1 replication. 293 cells were cotransfected with either the empty pcDNA3.1(⫹) vector or one expressing full-length HA-gM or HA-gM[Δ345-473] and with pNL4-3. At 48 h, the culture supernatants were collected and cell lysates were prepared. The levels of p24 determined in the culture supernatants were quantified using p24 antigen capture assays, and equivalent levels of p24 were used to determine virus infectivity using TZM.bl cell assays. (A) Immunoprecipitation analysis showing the expression of HSV-1 HA-gM and HA-gM[Δ345-473]. 293 cells were transfected with pcDNA3.1(⫹) expressing HA-gM[Δ345-473] (lane 1), empty pcDNA3.1(⫹) (lane 2), or HA-gM (lane 3). At 24 h posttransfection, cells were starved for methionine/ cysteine and radiolabeled with [35S]methionine/cysteine for 6 h. Cell lysates were prepared, and immunoprecipitations were performed with an anti-HA polyclonal serum. (B) Levels of p24 protein produced from cells cotransfected with pcDNA3.1(⫹), HA-gM, or HA-gM[Δ345-473] and the pNL4-3 genome. (C) Levels of infectious virus released into the culture medium from cells cotransfected with pcDNA3.1(⫹), HA-gM, or HA-gM[Δ345-473] and pNL4-3. (D and E) Pulse-chase analysis of 293 cells cotransfected with the empty pcDNA3.1(⫹) vector and pNL4-3, a vector expressing HA-gM and pNL4-3, a vector expressing HA-gM[Δ345-473] and pNL4-3. At 48 h posttransfection, the cells were starved for methionine/cysteine and radiolabeled with [35S]methionine/cysteine. The radiolabel was chased for 0 or 6 h, and viral proteins immunoprecipitated from the cell lysates (D) or cell culture medium (E) as described in Materials and Methods. The error bars indicate standard deviations. Numbers to the left indicate the positions of the molecular weight markers (in thousands).
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 12
on November 6, 2019 by guest
http://jvi.asm.org/
[image:12.585.48.364.67.526.2]have critically examined whether HSV-1 proteins could specifically restrict the
replica-tion of HIV-1. We explored the hypothesis that proteins expressed by HSV-1 could
impair the replication of HIV-1. We screened nine HSV-1 proteins by transfection for the
ability to restrict the replication of HIV-1. Of these proteins, US3 and UL24 profoundly
restricted HIV-1 replication, as determined by infectivity, viral protein biosynthesis, and
p24 release from cells.
In addition, our results indicated that HSV-1 and HSV-2 gM proteins restricted HIV-1
replication. In this study, we further analyzed the mechanism by which the HSV-1 gM
protein restricted HIV-1. Thirteen envelope glycoproteins that are involved in different
aspects of the viral replication cycle, including viral entry, egress from the nucleus, and
acquisition of the final envelope, are known to be incorporated into HSV-1 virions.
HSV-1 gM is a 473-amino-acid type III transmembrane protein that synthesizes as a
47-kDa precursor that matures into a family of 53- to 63-kDa glycosylated proteins (35,
36). Ectopic expression of HSV-1 gM results in the protein being targeted to the TGN
(37). While a detailed topological map of gM is lacking, the available evidence predicts
that it has eight membrane-spanning domains, with the N- and C-terminal ends
oriented toward the cytosol (22, 37, 38). In a recent study, the roles of several gM
domains were analyzed with respect to the subcellular distribution (23). The
investi-gators found that expression of a gM protein consisting of amino acids 19 to 343
resulted in localization to the TGN, while the expression of other truncated proteins,
such as gM 1 to 342, lacking the final residue of the last transmembrane domain, and
gM 133 to 473, lacking the first two transmembrane domains, resulted in the retention
of gM in the ER (23). These data suggested that all the transmembrane helices and the
C-terminal domain were required for transport to the TGN (23). While gM is not
essential for HSV-1 replication in cell culture, its deletion reduces viral yields by
approximately 10-fold (35, 39). HSV gM is known to interact with multiple viral proteins,
including gN (24, 40, 41). HSV-1 gN is a 91-amino-acid ER-resident protein when
expressed in the absence of other HSV proteins (24). However, in the presence of gM,
gN is transported to the TGN and requires the presence of the hydrophobic core (i.e.,
the region with eight transmembrane helices) (24). Site-directed mutagenesis showed
that gM and gN interact through the formation of a disulfide bond between C-46 of gN
and C-59 of gM, although the disulfide bond is not necessary for transport to the TGN
(24). The gM glycoprotein also interacts with other HSV-1 glycoproteins, such as gB, gD,
and gH/gL. The interaction of these glycoproteins results in redirection to the TGN, the
site of final envelopment (37). Additionally, gM can interact with and cause moderate
antagonism of BST-2 (tetherin), leading to enhanced HSV-1 release from cells (42, 43).
BST-2 is a protein that is antagonized by HIV-1 Vpu to facilitate virus release from cells
(44, 45). In another study, it was shown that BST-2 restricted the release of infectious
HSV-2 particles into the extracellular medium and that the HSV-2 glycoproteins gB, gD,
gH, and gL downregulated the endogenous expression of BST-2. Interestingly, HSV-2
gM did not downregulate the expression of BST-2 (46).
A salient question we have attempted to address in this study is what the
mecha-nism by which gM restricts HIV-1 is. Our results indicate that gp160 was not efficiently
cleaved into gp120/gp41, which occurs in the Golgi complex (47, 48). gp160 appeared
to be degraded with time, and very low levels of gp120 were observed in the culture
medium (and presumably the virus). One potential explanation for these observations
is that gM may perturb the transport of membrane proteins from the ER to the Golgi
complex. While we cannot totally rule out this scenario, other HSV-1 proteins are
synthesized and transported to the cell surface in the presence of gM. Also, the
production of infectious VSV from cells transfected with the empty pcDNA3.1(
⫹
) vector
or one expressing HA-gM was not affected, suggesting that the presence of gM had no
impact on the well-studied VSV G protein. Another potential explanation is that gM
may interact with gp160 within the rough endoplasmic reticulum (RER) to prevent
transport of gp160 to the Golgi complex and subsequent processing into gp120/gp41.
The HSV-1 gM contains four tyrosine-based motifs with the consensus sequence YXX
⌽
(where X is any amino acid and
⌽
is a bulky hydrophobic amino acid) and an acidic
on November 6, 2019 by guest
http://jvi.asm.org/
cluster (DEXXDDXXD). YXX
⌽
motifs, either alone or in combination with acidic clusters,
play crucial roles in efficient transport of the well-studied VSV-G protein and of the gB
proteins of different herpesviruses from the ER to the Golgi complex (21, 49–52).
Consistent with this hypothesis, the expression and processing of gp160 in the
pres-ence of the HA-gM[Δ345-473] mutant, lacking the C-terminal 129 amino acids, was
similar to transfection with the empty vector, indicating that the C-terminal domain of
gM is important for the inhibition of Env transport and processing. Alternatively, if gM
interacts with gp160, it could prevent proper folding of gp160. This in turn could signal
an endoplasmic reticulum-associated protein degradation response, leading to the
degradation of gp160 (53, 54). A working model is that the cytoplasmic domain of
HA-gM may interact with the cytoplasmic domain of gp160 to mask the tyrosine-based
motifs and or acidic clusters of gM that are essential for transport to the TGN. A
consequence of this interaction would be decreased transport of gp160 to the Golgi
complex and subsequent processing. This would decrease the levels of gp120/gp41
transported to the cell plasma membrane and incorporated into virions, resulting in the
restriction of HIV-1 spread through a novel mechanism. These studies also indicate that
the proteins from heterologous viruses, such as the herpesviruses, may provide
infor-mation on unique mechanisms of restriction that could possibly be used in the
development of novel antiviral approaches.
MATERIALS AND METHODS
Cells, plasmids, and viruses.293 cells were used for transfection of vectors expressing the HSV-1 proteins. The TZM-bl cell line, which was used as an indicator to measure virus infectivity, was obtained from the NIH AIDS Reagent Branch (55–58). A plasmid expressing Rev and gp160 from HIV-1 Env (HIV-1 strain Bal; pBal.26) and a plasmid with the entire HIV-1 NL4-3 genome (pNL4-3) were obtained from the NIH AIDS Reagent Branch. Both cell lines were maintained in Dulbecco’s minimal essential medium (DMEM) with 10% fetal bovine serum (R10FBS), 10 mM HEPES buffer, pH 7.3, 100 U/ml penicillin, 100 g/ml streptomycin, and 5g gentamicin. The HSV-1 proteins analyzed in this study were expressed as either untagged proteins (ICP0 and VP16), with a myc tag (US3, gM, UL24, UL46, UL47, and VP22), or with a Flag tag (US11) fused to the N termini of the proteins. All HSV-1 ORFs were PCR amplified from the HSV-1 (F) genome, and the sequences were verified by sequence analysis. A vector [pcDNA3.1(⫹)] expressing human codon-optimized HA-tagged HSV-1 gM was synthesized by GenScript, and myc-tagged HSV-2 gM was obtained from Origene (catalog number VC100874). An HSV-1 gM mutant in which the C-terminal 128 amino acids were deleted (HA-gM[Δ345-473]) was constructed by GenScript and expressed in pcDNA3.1(⫹). The sequence was confirmed by sequence analysis. Expression of HSV-1 proteins was confirmed by transfection with the TurboFect transfection reagent (ThermoFisher) and Western blot analysis using a mouse monoclonal antibody directed against the Flag tag (Sigma; F3165), a mouse monoclonal antibody against the c-myc tag (Santa Cruz; 9E10; sc-40), or rabbit antibodies against the individual proteins (ICP0 and VP16).
Analysis of virus production in the presence of HSV-1 proteins.To analyze the virus restriction properties of the HSV-1 proteins, 293 cells were transfected with either empty pcDNA3.1(⫹) vector or a vector expressing one of the HSV-1 proteins and with pNL4-3. At 48 h posttransfection, the culture medium was collected and clarified by low-speed centrifugation, and the supernatant was analyzed for p24 using antigen capture assays. The levels of infectious virus in the culture supernatants were titrated on TZM.bl cells (59, 60). TZM.bl cells have an integrated copy of a retrovirus vector expressing -galactosidase under the control of the HIV-1 LTR. If TZM.bl cells are infected with HIV-1, Tat protein expression will result in the expression of-galactosidase, and infected cells can be visualized by fixing and staining with X-Gal (5-bromo-4-chloro-3-indolyl--D-galactopyranoside). All assays were performed at least four times and analyzed for statistical significance using a two-tiered Studentttest with cells cotransfected with the empty vector pcDNA3.1(⫹) and pNL4-3 set at 100% infectivity.
Biosynthesis and processing of viral proteins in the presence of HSV-1 proteins.The biosyn-thesis and processing of viral proteins were examined in the presence of US3, UL24, and gM. 293 cells were cotransfected with empty pcDNA3.1(⫹) or pcDNA3.1(⫹) expressing HA-US3, HA-UL24, or HA-gM and with pNL4-3. At 24 h, the cells were washed and incubated in DMEM without methionine/cysteine for 2 h. The cells were washed and radiolabeled in DMEM containing 500Ci [35S]Translabel (methionine and cysteine; MP Biomedical) for 16 h. The culture medium was collected, subjected to low-speed centrifugation, and made 1⫻with respect to radioimmunoprecipitation assay (RIPA) buffer (1⫻RIPA buffer is 50 mM Tris-HCl, pH 7.5, 50 mM NaCl, 0.5% deoxycholate, 0.2% SDS, 10 mM EDTA). Cell lysates were prepared by lysis in RIPA buffer for 10 min on ice. The lysates were centrifuged in a microcentrifuge at 14,000 rpm for 10 min, and the supernatants were transferred to fresh tubes. HIV-1 proteins were immunoprecipitated using a monkey anti-SHIV antibody (which recognizes the gp160/gp120 and gp41 proteins of HIV-1) and a monoclonal antibody against p24 (number 24-4 from the NIH AIDS Reagent Program). The immunoprecipitates were collected by incubation with protein A-Sepharose beads at 4°C. The beads were washed three times with RIPA buffer, and the samples were resuspended in sample
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 14
on November 6, 2019 by guest
http://jvi.asm.org/
reducing buffer. The samples were boiled, and the proteins were separated by SDS-PAGE (10% gel) and visualized using standard radiographic techniques.
Restriction of infectious-virus production by HA-gM.Following the initial screen, we examined HSV-1 gM for the ability to restrict the replication of infectious virus. We generated infectious HIV-1 NL4-3 pseudotyped with the VSV-G protein by cotransfecting 293 cells with a vector expressing VSV-G protein and one containing the full-length HIV-1 NL4-3 genome. At 48 h, the culture medium was collected, clarified by low-speed centrifugation (1,200 rpm; 10 min), and frozen at⫺86°C. The number of infectious units was determined using 293 cells, serial 10-fold dilutions of virus, and p24 assays. To determine if HA-gM would restrict the production of infectious NL4-3, 293 cells were transfected with the vector expressing HA-gM. At 24 h posttransfection, the cultures were inoculated with VSV-G/NL4-3 at a multiplicity of infection of 0.01. At 72 h posttransfection, the levels of infectious virus in the culture supernatants were titrated on TZM.bl cells as described above (59, 60). The 50% tissue culture infectious doses (TCID50) of virus in TZM.bl cells were determined, and the numbers of TCID50in the empty vector and the vector expressing HA-gM were compared. All assays were performed at least four times and analyzed for statistical significance using two-tiered Studentttests.
To examine the specificity of HA-gM restriction, we also analyzed the ability of HA-gM to restrict VSV. 293 cells were transfected with the empty vector or one expressing HA-gM. At 24 h posttransfection, the cells were inoculated with VSV (MOI⫽0.01) for 1 h, at which time the inoculum was removed and the cells were washed three times. The infected cells were incubated in medium for 24 h, at which time the culture medium was removed and centrifuged at 1,200 rpm for 10 min. The levels of infectious virus in the culture supernatants were titrated using 10-fold dilutions and 293 cells in 96-well plates. The TCID50of virus in the 293 cells was determined, and the numbers of TCID50in the empty vector and the vector expressing HA-gM were compared.
Pulse-chase analysis of biosynthesis and processing of viral proteins in the presence of HSV-1 gM.293 cells were transfected with the empty pcDNA3.1(⫹) vector or pcDNA3.1(⫹) expressing HA-gM and pNL4-3. At 48 h, the cells were washed and incubated in medium without methionine/cysteine for 2 h, followed by radiolabeling with 500Ci [35S]methionine/cysteine (Translabel) for 30 min. The medium was removed, and the cells were washed twice in medium containing 100⫻excess cold methionine and cysteine and then incubated in the same medium for 0, 1, 3, and 6 h. At various times, the culture medium was collected, subjected to low-speed centrifugation, and made 1⫻with respect to RIPA. The cell lysates were prepared as described above. HIV-1 proteins were immunoprecipitated and processed, and analysis was performed as described above. In addition, the synthesis, processing, and transport of HIV-1 Env were also analyzed in the absence of other HIV-1 proteins. 293 cells were cotransfected with plasmid pBal.26, which expresses HIV-1 Env (strain Bal), and either empty pcDNA3.1(⫹) vector or one expressing HA-gM. The pulse-chase analysis was performed as described above, except that goat anti-gp160 antibodies were used in the immunoprecipitation assays (NIH AIDS Reagent Program; catalog numbers 189 and 51).
Intracellular localization of HIV-1 Env in the presence of HSV-1 gM. We determined if the intracellular localization of HIV-1 Env was altered in the presence of HSV-1 gM. We first determined the intracellular localization of gM protein in the absence of HIV-1. COS-7 cells were plated on 13-mm coverslips in 6-well plates and transiently transfected with the empty pcDNA3.1(⫹) vector or one expressing HA-gM protein using TurboFect transfection reagent (ThermoFisher) according to the man-ufacturer’s instructions. The cultures were maintained for 24 h, and the medium was removed; the cultures were washed three times in PBS (pH 7.4) and fixed in 4% paraformaldehyde (prepared in PBS) for 30 min. The coverslips were washed in PBS, permeabilized in 0.1% Triton X-100, washed, and incubated in PBS containing 5% BSA for 1 h. For colocalization of gM and the endoplasmic reticulum, the coverslips were reacted with a mouse monoclonal antibody against calnexin (mouse anti-calnexin; Novus Biologicals; 3H4A7) overnight at 4°C. The coverslips were washed and reacted with a secondary rabbit anti-mouse IgG Alexa Fluor 594 (Abcam ab150128). The coverslips were then reacted with rabbit anti-HA-fluorescein isothiocyanate (FITC) (Abcam; ab1208) and then counterstained with DAPI (4= ,6-diamidino-2-phenylindole) (1g/ml) for 5 min. For colocalization of HA-gM with the Golgi complex or TGN, the coverslips were reacted with rabbit anti-Golgin 97 (Abcam; ab84340) or rabbit anti-TGN46 (Novus Biologicals; NBP1-49643). The coverslips were washed and reacted with goat anti-rabbit IgG Alexa Fluor 647 (Abcam; ab150083), washed, and reacted with rabbit anti-HA-FITC (Abcam; ab1208) for 1 h. The coverslips were counterstained with DAPI (1g/ml) for 5 min. All the coverslips were washed and mounted in glycerol-containing mounting medium (SlowFade antifade solution A; Invitrogen). The coverslips were examined using a Leica TCS SPE confocal microscope with a 63⫻objective and a 2⫻ digital zoom using the Leica Application Suite X (LAS X) software package. A 405-nm filter was used to visualize DAPI staining, a 488-nm filter was used to visualize FITC, and a 647-nm filter was used to visualize Alexa Fluor 647. A minimum of 50 cells were examined for each sample, and the results shown in the figures are representative of each sample.
To analyze the intracellular localization of gp160 in the presence or absence of gM proteins, COS-7 cells were cotransfected with either empty pcDNA3.1(⫹) vector or one expressing HA-gM or HA-gM[Δ345-473] and pBal.26Env. At 24 h, the cells on coverslips were washed, fixed, permeabilized, and blocked as described above. The coverslips were reacted with a mixture of three goat anti-gp160 antibodies (NIH AIDS Reagents Branch; catalog numbers 51, 189, and 191) and with mouse monoclonal anti-HA antibody (SC7392) overnight at 4°C. The coverslips were washed and then reacted with donkey anti-goat IgG Alex Fluor 568 antibody (Abcam; ab175704) and Alexa Fluor 488 rabbit anti-mouse IgG (Thermo Fisher; A11059) for 2 h and then counterstained with DAPI (1g/ml) for 5 min. The coverslips
on November 6, 2019 by guest
http://jvi.asm.org/
were mounted, and a minimum of 50 cells were examined using the appropriate filters as described above.
ACKNOWLEDGMENTS
This work was supported by NIH grant R21 AI108391 to E.B.S.
We thank the Northwestern Sequence facility for sequence analyses and the KUMC
Biotechnology Core for oligonucleotide synthesis. The HIV-1 p24 Gag monoclonal
antibody (number 24-4) from Michael H. Malim was obtained through the NIH AIDS
Reagent Program, Division of AIDS, NIAID, NIH. pNL4-3 was obtained from Malcolm
Martin through the NIH AIDS Reagent Program, Division of AIDS, NIAID, NIH. The HIV-1
BaL.26 clone was obtained from J. R. Mascola through the NIH AIDS Reagent Program,
Division of AIDS, NIAID (catalog number 11446).
REFERENCES
1. Bloom DC. 2016. Alphaherpesvirus latency: a dynamic state of transcrip-tion and reactivatranscrip-tion. Adv Virus Res 94:53– 80.https://doi.org/10.1016/ bs.aivir.2015.10.001.
2. Kennedy PG, Rovnak J, Badani H, Cohrs RJ. 2015. A comparison of herpes simplex virus type 1 and varicella-zoster virus latency and reactivation. J Gen Virol 96:1581–1602.https://doi.org/10.1099/vir.0.000128. 3. Nicoll MP, Proença JT, Efstathiou S. 2012. The molecular basis of herpes
simplex virus latency. FEMS Microbiol Rev 36:684 –705.https://doi.org/ 10.1111/j.1574-6976.2011.00320.x.
4. Karn J, Stoltzfus CM. 2012. Transcriptional and posttranscriptional reg-ulation of HIV-1 gene expression. Cold Spring Harb Perspect Med 2:a006916.https://doi.org/10.1101/cshperspect.a006916.
5. Simon V, Bloch N, Landau NR. 2015. Intrinsic host restrictions to HIV-1 and mechanisms of viral escape. Nat Immunol 16:546 –553.https://doi .org/10.1038/ni.3156.
6. Strebel K. 2013. HIV accessory proteins versus host restriction factors. Curr Opin Virol 3:692– 699.https://doi.org/10.1016/j.coviro.2013.08.004. 7. Granerod J, Ambrose HE, Davies NW, Clewley JP, Walsh AL, Morgan D, Cunningham R, Zuckerman M, Mutton KJ, Solomon T, Ward KN, Lunn MP, Irani SR, Vincent A, Brown DW, Crowcroft NS, UK Health Protection Agency (HPA) Aetiology of Encephalitis Study Group. 2010. Causes of encephalitis and differences in their clinical presentations in England: a multicentre, population-based prospective study. Lancet Infect Dis 10: 835– 844.https://doi.org/10.1016/S1473-3099(10)70222-X.
8. Heaton RK, Franklin DR, Ellis RJ, McCutchan JA, Letendre SL, Leblanc S, Corkran SH, Duarte NA, Clifford DB, Woods SP, Collier AC, Marra CM, Morgello S, Mindt MR, Taylor MJ, Marcotte TD, Atkinson JH, Wolfson T, Gelman BB, McArthur JC, Simpson DM, Abramson I, Gamst A, Fennema-Notestine C, Jernigan TL, Wong J, Grant I, CHARTER Group, HNRC Group. 2011. HIV-associated neurocognitive disorders before and during the era of combination antiretroviral therapy: differences in rates, nature, and predictors. J Neurovirol 17:3–16.https://doi.org/10.1007/s13365-010 -0006-1.
9. Nightingale S, Winston A, Letendre S, Michael BD, McArthur JC, Khoo S, Solomon T. 2014. Controversies in HIV-associated neurocognitive disor-ders. Lancet Neurol 13:1139 –1151.https://doi.org/10.1016/S1474-4422 (14)70137-1.
10. Sacktor N, Skolasky RL, Seaberg E, Munro C, Becker JT, Martin E, Ragin A, Levine A, Miller E. 2016. Prevalence of HIV-associated neurocognitive disorders in the Multicenter AIDS Cohort Study. Neurology 86:334 –340. https://doi.org/10.1212/WNL.0000000000002277.
11. Adams O, Besken K, Oberdörfer C, MacKenzie CR, Takikawa O, Däubener W. 2004. Role of indoleamine-2,3-dioxygenase in alpha/beta and gamma interferon mediated antiviral effects against herpes simplex virus infec-tions. J Virol 78:2632–2636.https://doi.org/10.1128/JVI.78.5.2632-2636 .2004.
12. Chauhan A. 2015. Enigma of HIV-1 latent infection in astrocytes: an in-vitro study using protein kinase C agonist as a latency reversing agent. Microbes Infect 17:651– 659.https://doi.org/10.1016/j.micinf.2015 .05.006.
13. Chauhan A, Mehla R, Vijayakumar TS, Handy I. 2014. Endocytosis-mediated HIV-1 entry and its significance in the elusive behavior of the virus in astrocytes. Virology 456-457:1–19.https://doi.org/10.1016/j.virol .2014.03.002.
14. Chen NC, Partridge AT, Sell C, Torres C, Martín-García J. 2017. Fate of
microglia during HIV-1 infection: from activation to senescence? Glia 65:431– 446.https://doi.org/10.1002/glia.23081.
15. Joseph SB, Arrildt KT, Sturdevant CB, Swanstrom R. 2015. HIV-1 target cells in the CNS. J Neurovirol 21:276 –289.https://doi.org/10.1007/ s13365-014-0287-x.
16. Li J, Hu S, Zhou L, Ye L, Wang X, Ho J, Ho W. 2011. Interferon lambda inhibits herpes simplex virus type I infection of human astrocytes and neurons. Glia 59:58 – 67.https://doi.org/10.1002/glia.21076.
17. Lokensgard JR. 2001. Robust expression of TNF-, IL-1, RANTES, and IP-10 by human microglial cells during non-productive infection with herpes simplex virus. J Neurovirol 7:208 –219.https://doi.org/10 .1080/13550280152403254.
18. Reinert LS, Lopušná K, Winther H, Sun C, Thomsen MK, Nandakumar R, Mogensen TH, Meyer M, Vægter C, Nyengaard JR, Fitzgerald KA, Paludan SR. 2016. Sensing of HSV-1 by the cGAS-STING pathway in microglia orchestrates antiviral defence in the CNS. Nat Commun 7:13348.https:// doi.org/10.1038/ncomms13348.
19. Seifert G, Schilling K, Steinhauser C. 2006. Astrocyte dysfunction in neurological disorders: a molecular perspective. Nat Rev Neurosci 7:194 –206.https://doi.org/10.1038/nrn1870.
20. Lawson LJ, Perry VH, Gordon S. 1992. Turnover of resident microglia in the normal adult mouse brain. Neuroscience 48:405– 415.https://doi .org/10.1016/0306-4522(92)90500-2.
21. Heineman TC, Hall SL. 2001. VZV gB endocytosis and Golgi localization are mediated by YXXphi motifs in its cytoplasmic domain. Virology 285:42– 49.https://doi.org/10.1006/viro.2001.0930.
22. Raschbichler V, Lieber D, Bailer SM. 2012. NEX-TRAP, a novel method for in vivo analysis of nuclear export of proteins. Traffic 13:1326 –1334. https://doi.org/10.1111/j.1600-0854.2012.01389.x.
23. Striebinger H, Zhang J, Ott M, Funk C, Radtke K, Duron J, Ruzsics Z, Haas J, Lippé R, Bailer SM. 2015. Subcellular trafficking and functional impor-tance of herpes simplex virus type 1 glycoprotein M domains. J Gen Virol 96:3313–3325.https://doi.org/10.1099/jgv.0.000262.
24. Striebinger H, Funk C, Raschbichler V, Bailer SM. 2016. Subcellular traf-ficking and functional relationship of the HSV-1 glycoproteins N and M. Viruses 8:83.https://doi.org/10.3390/v8030083.
25. Davis MG, Kenney SC, Kamine J, Pagano JS, Huang ES. 1987. Immediate-early gene region of human cytomegalovirus trans-activates the pro-moter of human immunodeficiency virus. Proc Natl Acad Sci U S A 84:8642– 8646.https://doi.org/10.1073/pnas.84.23.8642.
26. Golden MP, Kim S, Hammer SM, Ladd EA, Schaffer PA, DeLuca N, Albrecht MA. 1992. Activation of human immunodeficiency virus by herpes simplex virus. J Infect Dis 166:494 – 499.https://doi.org/10.1093/ infdis/166.3.494.
27. Kenney S, Kamine J, Markovitz D, Fenrick R, Pagano J. 1988. An Epstein-Barr virus immediate-early gene product trans-activates gene expression from the human immunodeficiency virus long terminal repeat. Proc Natl Acad Sci U S A 85:1652–1656.https://doi.org/10.1073/pnas.85.5.1652. 28. Lusso P, Ensoli B, Markham PD, Ablashi DV, Salahuddin SZ, Tschachler E,
Wong-Staal F, Gallo RC. 1989. Productive dual infection of human CD4⫹
T lymphocytes by HIV-1 and HHV-6. Nature 337:370 –373.https://doi .org/10.1038/337370a0.
29. Mosca JD, Bednarik DP, Raj NB, Rosen CA, Sodroski JG, Haseltine WA, Hayward GS, Pitha PM. 1987. Activation of human immunodeficiency virus by herpesvirus infection: identification of a region within the long
Polpitiya Arachchige et al. Journal of Virology
January 2018 Volume 92 Issue 2 e01476-17 jvi.asm.org 16
on November 6, 2019 by guest
http://jvi.asm.org/
terminal repeat that responds to a trans-acting factor encoded by herpes simplex virus 1. Proc Natl Acad Sci U S A 84:7408 –7412.https:// doi.org/10.1073/pnas.84.21.7408.
30. Mosca JD, Bednarik DP, Raj NB, Rosen CA, Sodroski JG, Haseltine WA, Pitha PM. 1987. Herpes simplex virus type-1 can reactivate transcription of latent human immunodeficiency virus. Nature 325:67–70.https://doi .org/10.1038/325067a0.
31. Schafer SL, Vlach J, Pitha PM. 1996. Cooperation between herpes simplex virus type 1-encoded ICP0 and Tat to support transcription of human immunodeficiency virus type 1 long terminal repeat in vivo can occur in the absence of the TAR binding site. J Virol 70:6937– 6946.
32. Albrecht MA, DeLuca NA, Byrn RA, Schaffer PA, Hammer SM. 1989. The herpes simplex virus immediate-early protein, ICP4 is required to poten-tiate replication of human immunodeficiency virus in CD4⫹
lympho-cytes. J Virol 63:1861–1868.
33. Vlach J, Pitha PM. 1993. Differential contribution of herpes simplex virus type 1 gene products and cellular factors to the activation of human immunodeficiency virus type 1 provirus. J Virol 67:4427– 4431. 34. Calistri A, Parolin C, Palù G. 2003. Herpes simplex virus type 1 can either
suppress or enhance human immunodeficiency virus type 1 replication in CD4-positive T lymphocytes. J Med Virol 70:163–170.https://doi.org/ 10.1002/jmv.10350.
35. Baines JD, Roizman B. 1991. The open reading frames UL3, UL4, UL10, and UL16 are dispensable for the replication of herpes simplex virus 1 in cell culture. J Virol 65:938 –944.
36. Baines JD, Roizman B. 1993. The UL10 gene of herpes simplex virus 1 encodes a novel viral glycoprotein, gM, which is present in the virion and in the plasma membrane of infected cells. J Virol 67:1441–1452. 37. Crump CM, Bruun B, Bel S, Pomeranz LE, Minson T, Browne HM. 2004.
Alphaherpesvirus glycoprotein M causes the relocalization of plasma membrane proteins. J Gen Virol 85:3517–3527.https://doi.org/10.1099/ vir.0.80361-0.
38. Zhang J, Nagel CH, Sodeik B, Lippé R. 2009. Early, active, and specific localization of herpes simplex virus type 1 gM to nuclear membranes. J Virol 83:12984 –12997.https://doi.org/10.1128/JVI.01180-09.
39. MacLean CA, Robertson LM, Jamieson FE. 1993. Characterization of the UL10 gene product of herpes simplex virus type 1 and investigation of its role in vivo. J Gen Virol 74:975–983.https://doi.org/10.1099/0022 -1317-74-6-975.
40. El Kasmi I, Lippé R. 2015. Herpes simplex virus 1 gN partners with gM to modulate the viral fusion machinery. J Virol 89:2313–2323.https://doi .org/10.1128/JVI.03041-14.
41. Koyano S, Mar EC, Stamey FR, Inoue N. 2003. Glycoproteins M and N of human herpesvirus 8 form a complex and inhibit cell fusion. J Gen Virol 84:1485–1491.https://doi.org/10.1099/vir.0.18941-0.
42. Blondeau C, Pelchen-Matthews A, Mlcochova P, Marsh M, Milne RS, Towers GJ. 2013. Tetherin restricts herpes simplex virus 1 and is antag-onized by glycoprotein M. J Virol 87:13124 –13133.https://doi.org/10 .1128/JVI.02250-13.
43. Zenner HL, Mauricio R, Banting G, Crump CM. 2013. Herpes simplex virus 1 counteracts tetherin restriction via its virion host shutoff activity. J Virol 87:13115–13123.https://doi.org/10.1128/JVI.02167-13.
44. Neil SJ, Zang T, Bieniasz PD. 2008. Tetherin inhibits retrovirus release and is antagonized by HIV-1 Vpu. Nature 451:425– 430.https://doi.org/10 .1038/nature06553.
45. Van Damme N, Goff D, Katsura C, Jorgenson RL, Mitchell R, Johnson MC, Stephens EB, Guatelli JC. 2008. The interferon-induced protein BST-2 restricts HIV-1 release and is downregulated from the cell surface by the viral Vpu protein. Cell Host Microbe 3:245–252.https://doi.org/10.1016/ j.chom.2008.03.001.
46. Liu Y, Luo S, He S, Zhang M, Wang P, Li C, Huang W, Hu B, Griffin GE, Shattock RJ, Hu Q. 2015. Tetherin restricts HSV-2 release and is
counter-acted by multiple viral glycoproteins. Virology 475:96 –109.https://doi .org/10.1016/j.virol.2014.11.005.
47. Hallenberger S, Bosch V, Angliker H, Shaw E, Klenk HD, Garten W. 1992. Inhibition of furin-mediated cleavage activation of HIV-1 glycoprotein gp160. Nature 360:358 –361.https://doi.org/10.1038/360358a0. 48. Stein BS, Engleman EG. 1990. Intracellular processing of the gp160 HIV-1
envelope precursor. Endoproteolytic cleavage occurs in a cis or medial compartment of the Golgi complex. J Biol Chem 265:2640 –2649. 49. Favoreel HW. 2006. The why’s of Y-based motifs in alphaherpesvirus
envelope proteins. Virus Res 117:202–208.https://doi.org/10.1016/j .virusres.2005.11.007.
50. Heineman TC, Krudwig N, Hall SL. 2000. Cytoplasmic domain signal sequences that mediate transport of varicella-zoster virus gB from the endoplasmic reticulum to the Golgi. J Virol 74:9421–9430.https://doi .org/10.1128/JVI.74.20.9421-9430.2000.
51. Heineman TC, Connolly P, Hall SL, Assefa D. 2004. Conserved cytoplas-mic domain sequences mediate the ER export of VZV, HSV-1, and HCMV gB. Virology 328:131–141.https://doi.org/10.1016/j.virol.2004.07.011. 52. Sevier CS, Weisz OA, Davis M, Machamer CE. 2000. Efficient export of the
vesicular stomatitis virus G protein from the endoplasmic reticulum requires a signal in the cytoplasmic tail that includes both tyrosine-based and di-acidic motifs. Mol Biol Cell 11:13–22.https://doi.org/10 .1091/mbc.11.1.13.
53. Caramelo JJ, Parodi AJ. 2015. A sweet code for glycoprotein folding. FEBS Lett 589:3379 –3387.https://doi.org/10.1016/j.febslet.2015.07.021. 54. Wang Q, Groenendyk J, Michalak M. 2015. Glycoprotein quality control and endoplasmic reticulum stress. Molecules 20:13689 –13704.https:// doi.org/10.3390/molecules200813689.
55. Derdeyn CA, Decker JM, Sfakianos JN, Wu X, O’Brien WA, Ratner L, Kappes JC, Shaw GM, Hunter E. 2000. Sensitivity of human immunode-ficiency virus type 1 to the fusion inhibitor T-20 is modulated by coreceptor specificity defined by the V3 loop of gp120. J Virol 74: 8358 – 8367.https://doi.org/10.1128/JVI.74.18.8358-8367.2000. 56. Platt EJ, Bilska M, Kozak SL, Kabat D, Montefiori DC. 2009. Evidence that
ecotropic murine leukemia virus contamination in TZM-bl cells does not affect the outcome of neutralizing antibody assays with human immu-nodeficiency virus type 1. J Virol 83:8289 – 8292.https://doi.org/10.1128/ JVI.00709-09.
57. Takeuchi Y, McClure HO, Pizzato M. 2008. Identification of gammaretro-viruses constitutively released from cell lines used for human immuno-deficiency virus research. J Virol 82:12585–12588.https://doi.org/10 .1128/JVI.01726-08.
58. Wei X, Decker JM, Liu H, Zhang Z, Arani RB, Kilby JM, Saag MS, Wu X, Shaw GM, Kappes JC. 2002. Emergence of resistant human immunode-ficiency virus type 1 in patients receiving fusion inhibitor (T-20) mono-therapy. Antimicrob Agents Chemother 46:1896 –1905.https://doi.org/ 10.1128/AAC.46.6.1896-1905.2002.
59. Schmitt K, Katuwal M, Zdong CL, Wang Y, Stephens EB. 2014. Analysis of the N–terminal positively charged residues of the simian immunodefi-ciency virus Vif reveals a critical amino acid required for antagonism of rhesus APOBEC3D, G, and H. Virology 449:140 –149.https://doi.org/10 .1016/j.virol.2013.10.037.
60. Wang Y, Wang Z, Pramanik A, Santiago ML, Qiu J, Stephens EB. 2016. A chimeric human APOBEC3A protein with a three amino acid insertion confers differential HIV-1 and adeno-associated virus restriction. Virology 498:149 –163.https://doi.org/10.1016/j.virol.2016.08.001.
61. Solomon T, Michael BD, Smith PE, Sanderson F, Davies NW, Hart IJ, Holland M, Easton A, Buckley C, Kneen R, Beeching NJ, National Enceph-alitis Guidelines Development and Stakeholder Groups. 2012. Manage-ment of suspected viral encephalitis in adults: Association of British Neurologists and British Infection Association National Guidelines. J Infect 64:347–373.https://doi.org/10.1016/j.jinf.2011.11.014.
on November 6, 2019 by guest
http://jvi.asm.org/

![FIG 5 The gM from HSV-2 also restricts HIV-1 replication. 293 cells were cotransfected with the pcDNA3.1(HA-gM[HSV-1] and myc-gM[HSV-2] proteins in 293 cells](https://thumb-us.123doks.com/thumbv2/123dok_us/140101.17245/8.585.42.444.69.277/restricts-hiv-replication-cells-cotransfected-pcdna-proteins-cells.webp)
![FIG 9 Intracellular expression of HIV-1 gp160 in the presence of HA-gM or HA-gM[Δ345-473]](https://thumb-us.123doks.com/thumbv2/123dok_us/140101.17245/11.585.71.338.71.271/fig-intracellular-expression-hiv-presence-ha-ha-d.webp)