0022-538X/91/010061-10$02.00/0
CopyrightX) 1991, American Society forMicrobiology
The
Target
DNA
Sequence
for Resolution of Poxvirus
Replicative
Intermediates
Is
an
Active
Late
Promoter
DAVID STUART, KATHRYN GRAHAM, MARTHASCHREIBER,
COLINMACAULAY, AND GRANTMcFADDEN*
Department of Biochemistry, Universityof Alberta, Edmonton, Alberta, Canada T6G 2H7
Received 6July1990/Accepted 2 October 1990
The linear double-stranded genomes of poxviruses such as Shope fibroma virus (SFV) replicate
autono-mouslywithinthe cytoplasm of infected cells, and it is believedthat allofthereplication functionsarevirally
encoded. During DNA replicationthe incompletely base-paired terminal hairpin loops of the viral genome
transientlyexistintheform ofinvertedrepeatreplicative intermediates. These invertedrepeat structuresform
thetargetfor telomereresolutioneventsthatincludesequence-specffic cleavage and directedstrand exchange toformthe hairpintermini ofprogenyvirusgenomes.Theterminalsequencedomain which formsthe telomere
resolution target (TRT) shares considerable sequence similarity with viral latepromoters. In this study we
demonstrate that theTRT of SFV is capable of functioning as astrongviral promoter late in infection. A
spectrumofTRTmutationsaffectstelomereresolutionand latetranscription inastrictly concordant fashion,
suggestingthat thetwoactivitiesmaybeinextricably linked. Furthersupportfor thisconceptcomesfrom the
demonstrationthatalate SFVpromotersequencedesignatedcrypticTRT, which differs substantially from the
nativeTRTintermsofsequence,cansupporttelomere resolution when placed in thecorrectspatialcontext.
The proposed model for telomere resolution invokes directed unwinding of the TRT double helix by a
transcription initiation complexandprocessing oftheresulting secondarystructurebyvirallate-geneproducts.
Linear DNAgenomes are a commonfeature of complex
eucaryotic organisms and many viruses (2, 25). The
com-plete and accurate replication of linear DNA molecules
requiressomespecial mechanism to account for the need of
allknownDNApolymerases toutilize a 3' hydroxyl to prime
synthesis (8, 50). Many eucaryotic organisms have
over-cometheproblem ofreplicating chromosome termini bythe
useofatelomereterminaltransferase (telomerase) activity,
which synthesizes terminalsequenceina
template-indepen-dentfashion (reviewed in references 4 and53). An
alterna-tivestrategyfor thereplicationandmaintenance of
chromo-some termini isthe possessionof covalentlylinkedterminal
hairpin loops. This typeofDNA structure has been
identi-fied in avariety ofeucaryotic organelle and plasmid
chro-mosomes (5, 18,42), parvoviruses (2), African swine fever
virus(22),and poxviruses(1,16). TheleporipoxvirusShope
fibroma virus (SFV)and orthopoxvirus vacciniavirus have
double-stranded linearDNA genomes with hairpin termini
which exist in"flip" and"flop"configurations(1,16).During
the viralreplicative cycle,DNAsynthesis throughthe
hair-pin loops transiently generates inverted repeat structures
(38)which fusenascent viral genomestogether in arrays of
linear concatemers (13, 35). Production ofmature progeny
viral DNA requires that the inverted repeat replicative
intermediates be resolvedto yield unit-lengthgenomes with
flip and flop hairpin termini. The cytoplasmic site of viral
replication and the ability of poxviruses to
replicate
inenucleatedcells(37)suggestthatalloftheactivities
required
for telomere replication and resolution are virally encoded
(reviewed inreference15).
Investigations into the resolution process have been
greatly aidedbytheobservationthatcloned versions ofthe
telomere replicative intermediates are replicated and
re-solved into linear minichromosomes with viral hairpin
ter-*Corresponding author.
mini when transfected into virus-infected cells (16, 33). The
minimalviral DNA sequences requiredincisfor the
resolu-tion ofreplicative intermediates have been determined by
bothdeletionandpointmutationanalysis(14, 16, 34)tobea
stretch of about 20 nucleotides located near the hairpin
termini.This target sequenceforthe viralresolution
machin-eryhasbeendesignatedthetelomereresolutiontarget(TRT)
(15). Despitetheoverall lackofsequencehomologynearthe
terminibetweendifferentgeneraof poxviruses for which this
information is available, the deduced TRT among various
poxviruses
shows remarkable conservation(15),suggesting
thatacommonresolution mechanism may beutilizedbyall
poxviruses. This is
underscored
by the fact that SFV andvaccinia virus canresolveeach other's telomeres (16) andis
consistentwith theobservationthat theseviruses
recognize
eachother'spromoters (28)andearly
transcription
termina-tion signals (49, 52).
Inspection of the conserved minimal TRT sequence
re-veals that it is verysimilartotherecentlydefinedconsensus
sequence for poxvirus late promoters (12). This sequence
similarity and the recent report that the telomeres ofthe
orthopoxvirus vacciniavirusaretranscribedlate ininfection
(40) suggest the
possibility
thattranscriptional
events insomewayinfluence DNA
replication
ortelomereresolution.In order to study the possible role of
transcription
intelomere resolution,we haveundertakenan
investigation
oftheabilityof the SFV TRTtofunctionas aviral promoter. In
this communication we report the existence ofan intrinsic
relationship between telomere resolution and
specific
late-promoteractivity. Potential mechanisms by which the viral
transcriptional machinerymightactivate telomere resolution
arediscussed.
MATERIALS ANDMETHODS
Cells and viruses. BGMK cells
(African
greenmonkey)
obtained from the American Type Culture Collection were
61
on November 10, 2019 by guest
http://jvi.asm.org/
grown in monolayer culture in Dulbeco modified Eagle's
medium (DME; GIBCO Laboratories) supplemented with
10% newborn calf serum (GIBCO). Human thymidine
kinase-minus (TK-) H143 cells usedtoselectrecombinant vaccinia
viruses were obtainedfrom D. Panicaliandgrownin
mono-layerculture inDMEsupplementedwith5%fetal calfserum
and 20 ,ug of
5-bromo-2'-deoxyuridine
(Sigma)per ml.Vaccinia virus (strain WR) obtained from the American
Type CultureCollection was propagatedby the infection of
suspensionculturesofHeLaS3cells, whichwere agenerous
gift from V. Paetkau. SFV (Kasza strain) was similarly
propagated by theinfectionofmonolayer culturesof BGMK
cells. Virus stocks were prepared from cells harvested by
centrifugation at 48 h postinfection. The cell pellet was
swelled andDouncehomogenized(11, 51).Virustiters were
determined by infecting monolayers of 1.6 x
106
BGMKcells with 10-fold serial dilutions of the crude virus stocks.
After1 h ofadsorptionina200-,u volume, fresh mediumwas
added and the infections wereallowedtoproceed for 48h, at
whichtime thecells were fixed andstained with 0.1% crystal
violet.
The recombinant vaccinia virus vCST-2a was generated
by the standard mechanism ofhomologous recombination
between aplasmid containingafusion oftheSFV TRTand
a bacterial chloramphenicol acetyltransferase (CAT)
re-portercassette intothevaccinia virusthymidine kinase(TK)
gene (30). Theinsertion vector usedin this study, pVV5.1,
hasbeenpreviously described(20). Thisvectorwascleaved
at
unique
sites with restriction enzymes SalI and BamHI,and the linearized vector, with the vaccinia virus 7.5-kDa
protein (7.5K protein) promoter removed, was purified and
blunt ended with T4 DNA polymerase in the presence of
equalconcentrations of thefour
deoxyribonucleotides.
Thisvectorwasused astherecipientfor aDNAfragment which
had been excised from pCST-1
(described
below) withBamHI and SstI and whose overhanging ends had been
blunted with T4 DNA polymerase. The resulting insertion
vector, pVCST-1, consisted of the CAT gene under the
regulation of theminimal TRT sequence of SFVflankedon
both sides by vaccinia virus TK sequences. Recombinant
TK-viruses were selected on H143 cells in thepresence of
5-bromo-2'-deoxyuridine
and screened for the presence oftheinsertby dotblothybridization withaCATgene-specific
probe. The insertion site of the TRT-CAT sequence was
confirmedby Southern blotting.
Plasmids and strains. All bacterial plasmids which
con-tainedinverted repeats of viralsequence were maintainedin
Escherichia coli DB1256 (16), and all other plasmids were
propagated in E. coli HB101. The construction of all
plas-mids which contained inverted repeats of wild-type SFV
sequence and the deletion derivatives ofthis sequence has
been described previously (14, 16). Plasmids which
con-tained large inverted repeats of SFV telomere sequence
(pSAB-67,
pSXB-102,
andpSCB-la)
were all derived byremoving theappropriateAccI,
XhoI,
orClaIfragment fromthe plasmid pYSF1-30 described previously (16) and
sub-cloning the fragment into the SmaI site of pUC19. The
plasmid pSCX-1 was derived from pSCB-la by cleavage
with XhoI to delete the central-axis 0.7-kb fragment and
self-ligation ofthe vector to generate a palindromic insert
with acentral-axis XhoI site (see Fig. 6a).
Plasmids containingthe SFV TRTsequences upstream of
a promoterless CAT gene were generated by excising one
half of the inverted repeatofpSD-19 (14) ordeletion
deriv-atives of this construct as a
HindIII-AflIl
fragment. Theoverhanging 5' ends ofthese fragments were filled withT4
DNA
polymerase,
and the bluntedfragments
wereligated
intotheSmaIsiteofthevector
pMTL-24 (9).
Thetelomeredeletions of interest were then isolated from the
pMTL
constructsas
KpnI
fragments
andcloned into theKpnI
site ofpUC19-CAT 4,
whichcontains theCATgeneclonedintothe SmaI site of
pUC19
oriented such that CAT can beplaced
under thecontrolof viral sequences introducedinto theKpnI,
SstI,
orEcoRI sites(28).
The prototypemember ofthis series ofvectors,
pCST-1,
contains the entirewild-type
TRTsequence ofSFVobtainedfromthevector
pSAD-2
(14)
and
directing
expression
ofthe CAT gene.Point mutations were
generated
inthe TRTby
blunt-endligating
anEcoRI-AflII fragment
frompSAD-2
intotheSmaIsite of
M13mpl9.
Thisligation destroys
the EcoRI site butregenerates the central-axis
AflIl
site.Oligonucleotide-di-rected
site-specific
mutagenesis
wasperformed
(26),
and theappropriate
mutationswereidentifiedby
dot blothybridiza-tion and then confirmed
by sequencing.
ThemutatedEcoRI-AflIl fragments
were isolated andself-ligated
orligated
to afragment
ofwild-type
sequencetogenerateinvertedrepeats
whichwere subclonedinto the SstI site of
pUC19
and usedforassaysof telomere resolution. Forpromoter assays, the
mutated
fragments
wereligated
into thevectorpUC19-CAT-4 so that the sequences could be tested fortheir
ability
todirect CAT
expression.
Analysis of telomere resolution in transfected cells. The
ability
of thewild-type
or mutated TRT invertedrepeat
sequences toberesolvedinto linearminichromosomeswith
hairpin
termini was assayed aspreviously
described(16).
Plasmid DNA
containing
inverted repeat telomerese-quences was transfected into
previously
infectedmonolay-ers of1.6 x 106 BGMK
cells,
and totalDNA washarvested24 h after transfection. This DNA was treated with
restric-tion enzyme
DpnI
to cleave theinput unreplicated
DNA,
electrophoresed through
0.7% agarose gels, transfered tonitrocellulosepaper,
hybridized
withnick-translatedplasmid
DNAprobes, and visualizedby autoradiography.
Promoter andtranscript analyses. BGMKcells which had
been infected with virus andtransfected with plasmidDNA
wereharvested20 hpostinfection, and thelysateswereused
for CATassaysasmodified forSFV-infected cells(28). RNA
synthesis driven from the TRT promoter in vaccinia virus
recombinantswasanalyzed by primer extension asfollows.
Monolayers
of BGMK cellsin 100-mmdishes were infectedwiththerecombinant vaccinia virus vCST-2a at a
multiplic-ity
of 10 PFU percell;at2and 16 h postinfection, RNAwasharvestedfromtheinfected monolayersby scrapingthecells
into
guanidinium isothiocyanate
(Sigma) and pelleting theRNAthrough cesium chloride (10). Ofthis RNA, 40,ugwas
hybridized with 200 ng ofa 5'-end-labeled 17-mer
oligonu-cleotide
primer
whichwascomplementarytothe CAT gene.This primer was extended with avian myeloblastosis virus
reversetranscriptase (Life Sciences Inc.) in the presence or
absence ofdideoxynucleotides as described previously (3).
The extended products of these reactions were
electro-phoresed through 8% polyacrylamide sequencing gels
con-taining8M ureaand visualizedvia autoradiography. Unless
otherwise specified, all of the restriction and modification
enzymes used in this study were obtained from Bethesda
Research Laboratories and used under the conditions
rec-ommendedbythe manufacturer.
RESULTS
Similarity between consensus TRT and late-promoter
se-quences. During the productive replication cycle of
on November 10, 2019 by guest
http://jvi.asm.org/
I IA
a
replication t resolution
I IA
Hairpin terminus
Transient inverted repeat
IA I
b
Box I Box IASFV TTTTTTTCTAG GGTTA TAAATTA VAC TTTTTTTCTAG ACAC- TAAATAA CPV TTTTTTTCTAG ACAC- TAAATAA RCN TTTTTTTCTAG ACAT- TAAATAA
FIG. 1. (a) The poxviral telomere in thematurehairpin form and in theinvertedrepeatconfigurationwhich exists transiently during
DNA replication. The inverted repeat structure is resolved by
trans-acting viral factors to yield two daughter hairpin telomere structures.Therelative positions of boxesIandIAoftheminimal TRTsequences areindicated, but extrahelicalbases on thehairpin andnonpalindromic sitesonthe invertedrepeat are not shown. (b) One copy of the TRT sequence of SFV, vaccinia virus (VAC), cowpoxvirus(CPV), andracoonpoxvirus (RCN).
ruses, the hairpin termini aretransiently convertedinto the
inverted repeat replicative intermediate which is the
sub-stratefortrans-actingviralfactorsthatcatalyzethe
isomer-izationof these sequences into two daughter hairpin termini
(15). As shown in Fig. 1, the minimal sequence domain
required for resolution consists oftwo blocks of sequence,
designated boxesIand IA, that arehighlyconservedamong
poxviruses and are separatedbya spacerof4 or 5
nucleo-tides that has diverged. Inspection of the conserved
se-quences which constitute boxes I and IA reveals a strong
similarity to the consensus structure ofvaccinia virus late
promoters (12). As can be seen in Fig. 2, the pll late
promoterdiffers by onlyonenucleotide from theconsensus
TRTwithinthealigned boxIand IAdomains,andmany of
the other characterized late promoters have a conserved
TAAAT motif which corresponds exactly to the TRT box
IA.
Transcription directed by the TRT sequences of SFV. To
investigate
thepossibilitythat theTRT sequenceofSFV iscapable
ofdirecting
orientation-specific transcription, weconstructedaseries ofplasmidvectorsinwhichthebacterial
CAT gene was placedunder the regulation of
well-charac-terized poxviral promoters or a single copy of the TRT
sequence and a series of mutated versions of the TRT that
had been created for resolution assays. CAT activity
as-sayedfromcellsharvestedatlatetimes showed that the TRT
was capableof directing gene expression in an
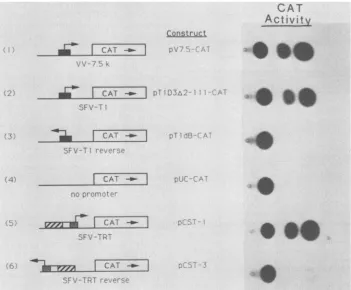
orientation-specificmanner(Fig. 3, lanes5 and6). Theeffectivenessof
the TRT as a promoter can be inferred by comparing the
amount of CAT activity directed from the TRT with the
levels from the SFV Ti early promoter (Fig. 3, lane 2) and
the vaccinia virus 7.5Kearly-late promoter(Fig. 3, lane 1).
A series of deletions of the SFV telomere sequence
indicate thatthe minimal TRT sequence in boxes I and IA
includes the essential sequence elements for promoter
activ-ity. 5' deletions which remove nucleotides from the
se-quence TT TTTT of box I (Fig. 4, lane 7) or 3' deletions
from the TAAAT of box IA(lane 8)eliminate the promoter
activity of the TRT. It has been observed that mutations
within the conserved TAAAT sequence of vaccinia virus
promoters will abrogate promoter function (12). When the
sequence TAAAT in box IA of the TRT is mutated to
TAACT, the ability of the TRT todirectgeneexpressionis
lost (Fig. 4, lane 10).
The mutatedversions of the TRT have been tested in an in
vivo resolution assay for their ability to support the
resolu-tion of inverted repeats into hairpin structures (14, 16).
Deletions from the 5' side which remove sequence from box
I orfromthe 3' side which remove box IA of the TRT onone
or both sides of the inverted repeat fail to support in vivo
hairpin resolution. Infact, thesamedeletionconstructs that
define the 5' and 3' boundriesof the TRT inresolution assays
also define the domain of the promoter function of the SFV
TRT. Thesame result has beenobserved forpointmutations
which alter the highly conserved TAAAT of box IA in terms
ofpromoteractivity (Fig. 4, lane 10) and resolution (seeFig.
7c). Thus a clearcorrelation canbe demonstratedbetween
theability of the TRT to direct transcription and the ability
to support theresolution of inverted repeats in vivo.
SFV TRT functions as a late promoter in a
transient-expression assay. Previouslyithas beenshown thatanearly
SFV promoter, Ti, will functioncorrectlyin a transfection
assay in cells infected with SFV or vacciniavirus (28). CAT
assays on lysates from infected/transfected cells indicate
that the TRTsequence of SFV can also direct the expression
of CATactivity in cells which had beeninfected with either
SFV orvaccinia virus(Fig. 5,lanes 1 and2). Thus the TRT
isrecognized bybothleporipoxvirusesandorthopoxviruses
as apromoter whenfusedas asinglecopy to the CAT gene
Box I -(spacer)- Sos IA
T T T T T T T C s a 0(4 or 5)- T A A A T TRT consensus.
T T T T T T T C T A T-( 4 ) T A A A T pll late promoter.
T T T T T T T A T A A-( 0 ) T A A A T ATI late promoter.
T T T T T T T T T 0 ( 4 ) T A A A T Synthetic MJ480 promoter. T T T T T A ! A 0 T A-( 13 ) T A A A T 7.5 k late promoter.
FIG. 2. The TRTisverysimilartopoxviruslatepromoters. The TRT sequence which is conserved among all of thepoxvirusesforwhich thesequenceis available (TRTconsensus)is shown alignedwith thesequences of the vaccinia virus 11-kDa gene late promoter(plllate promoter), cowpox A-type inclusion body promoter (ATI late promoter), an artificial strong late promoter synthesized to match the determinedconsensussequencefor vaccinia virus latepromoters(12)(SyntheticMJ480promoter),and thevacciniavirus 7.5-kDa genelate promoter(7.5klatepromoter).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.58.296.73.292.2] [image:3.612.150.459.610.680.2]Construct
L:*"
1
CAT-r
I
^C A T _ st'-7t i
CAT
Act
ivity
.4,,
4.
_t,1v r C!
.
S s...
S.,
FIG. 3. Promoterproperties of the SFVTRT sequence. Plasmid constructscontainingtheCATgenedownstream ofthevacciniavirus 7.5-kDa early-late promoter(pV7.5-CAT), the twoorientations of the SFV Ti early promoter(pTlD3A2-111-CAT andpTldB-CAT), no promoter(pUC-CAT),orthetwoorientations of the SFVTRT(pCST-1 andpCST-3)weretested fortheirabilitytoexpressCATactivityin transientassays whentransfected into SFV-infected cells. Theshadings of boxIandIAarethesame asin Fig.1.
and as a resolution signal when in an inverted repeat
configuration. When the DNAreplication inhibitorcytosine
arabinofuranoside (araC)is usedto treatcellsduring
poxvi-rusinfection, late-gene expressionand DNAreplication are
prevented while early-geneexpression continues(37). In the
presence ofaraC, SFV-infected cellscontinue todirect the
expression of CAT activity from theSFVTiearlypromoter;
however, verylittle CATactivity is observedtobedirected
fromthe TRT (Fig. 5, lanes 3 and 4),suggestingthat the TRT
functionsas apromoterprimarilylate in infection.
SFV TRT functions as a late promoter in a recombinant
vaccinia virus. The ability of vaccinia virus trans-acting
factors to recognize and utilize the TRT of SFV as a late
promoterprompted us to test theability of the TRT to act as
apromoterfrom within the context ofthe viral genome. A
recombinant vaccinia virus, vCST-2a, in which the
TRT-directed CAT genewasinserted into the vaccinia virus TK
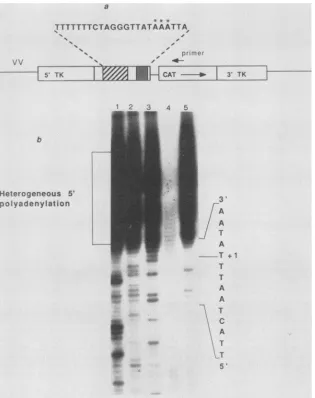
gene, was generated (Fig. 6). The 5' start site of RNA
collected at 16 h postinfection was mapped by primer
extension with a CAT-specific oligonucleotide probe. The
extended products from this primer display a number of
characteristics that are common to vaccinia virus late tran-scripts (Fig. 6b). As can be seen from the population of sizes whose diversity appears to begin within the TAAAT motif of
box IA (lanes 2 through 5), the 5' end is heterogeneous in
lengthandis likely to be polyadenylated in a fashion similar
tothat of late orthopoxvirus genes (3, 45). The length of the
apparent 5' poly(A) head of the TRT-CAT RNA is about 20
nucleotides, again consistent with other late poxviral
tran-scripts.The last5' sequence of the TRT-driven RNA which
is complementary to the DNA sequence is within the
TAAATcorrespondingtoTRTbox IA.Thus, in thiscontext
within the TK gene, the SFV TRT behaves as a rather
typical strong latepromoter.
Another late-promoter sequence can function to resolve
replicative intermediates. The observation that the minimal
TRT sequencecan act as alate promoterledus to
investigate
the possibilitythat other late promoters could, if
presented
in an inverted repeat configuration, function as a viral
telomere to be resolved into hairpins. To test this, we first
constructeda series oflarge inverted repeatsofSFV
telom-ere sequence (Fig. 7a). Plasmids pSAB-67, pSXB-102, and
pSCB-la all contain the native TRT within progressively
larger inverted repeats and can be efficiently resolved into
linearminichromosomes in vivo(Fig. 7b,lanes 1, 2,and3).
However, intheconstruct shown in lane4, thecentral-axis
XhoI fragment of pSCB-la was removed to generate
pSCX-1,whichcontainsa0.9-kbperfect palindromeof viral
DNAthat lacks the native central axis, includingthe entire
TRT sequences. Despite the removal of the SFV TRT
domain, pSCX-1canbereplicatedandresolved in vivo into
monomer and multimer linear minichromosomes (Fig. 7b,
lane4). Although the resolution of pSCX-1 is less efficient
than for constructs which possess the wild-type TRT, this
resultremainsparticularlystriking whenoneconsiders that
all of the palindromes of nonviral origin which have been
tested in theresolution assay havealwaysbeen inactive(31).
Compare, for example, the ability of this "cryptic" TRT
(cTRT)tocatalyzeresolutiontominichromosomes with the
completeablation of resolutionbyasingle pointmutation in
- ,-i; ,t,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.135.486.75.365.2]CAT
FY7Y~I NW
I i20 I
---XI.
VAR= :Ii Elt I(AT AC'lVITY
---N.
3*4. pSD77-CAT (A49) .L. . ---L-L-- f n-5. pSD)48-CAT (ASI)
6. pSD53-CAT (A91)
7. pSD56-CAT (Al12) -_I
8. pSD5-10-CATr (A57)
9. pCST-1 (CR) v----..Z JI r ==I S.
[image:5.612.109.514.81.368.2]10. pCST-l/A-{1Cl--- -X-- 8
FIG. 4. CATassaydemonstrating the minimal TRTsequencesresponsible forpromoteractivity. CATassaysin infected/transfected cells
wereperformedasforFig. 3. pSD19-CAT (lane3)contains all of thesequencesin pCST-1 (lane 2) but also includes46bpof 3'sequencewhich
extendstothe axis ofsymmetryof thereplicative intermediate invertedrepeat.This 3'sequencecontains severalstartandstopcodons which result inadecreased expression of CAT activity in relationtopCST-1. Theconstructs in lanes4through7are 5' deletionsoftheparent pSD19-CAT.pSD5-10-CAT (lane 8) isa3' deletion of pSD19-CAT which completelyremovesTRTbox IA. Thenumber of nucleotides deleted ineachcaseisindicated in brackets. The cloning of pCST-1 resulted in the deletion of nucleotide A16,whichwas restored bysite-specific
mutagenesistowild-type in pCST-1 (CR) (lane 9). In
pCST-1/A2,-.C,
the TAAATsequenceof boxIAwasmutatedtoTAACT. Fortelomere resolution, + indicates that the plasmid containing this inverted repeat was resolved in vivo into monomer and multimer linear minichromosomesand-indicates that the plasmidwasreplicated butnotresolved. Theresolution data shown in lanes1through 9 havebeenpresentedpreviously (14, 16), and resolution data for
pCST-1/A26--a'C
areshown inFig. 7c.the TRT box IA (Fig. 7c, lane 3) which had been shown
previously to destroy promoteractivity ofthe TRT(Fig. 3,
lane10).Apositive control for efficientinvivo resolution of
inverted repeats is provided by pSD-19, which possesses twoinvertedcopies of the SFV TRT (Fig. 7c, lane 1). The
replicated andresolved forms ofpSD-19maybe compared
Promoter Virus
( I) pCST-I TRT SFV
(2) pCST-1 tlRT Vaccinia
(3) pTI-CAT Iearly
CAT
D)rug Activity
* s
SFV araC
(4) pCST-I T,RT SFV araC'
0 to
0
FIG. 5. The TRTsequencefunctionsas apromoteronlylatein infection. Lanes 1and 2 illustrate thepromoterfunction of theSFV TRT when it istransfected into cellsinfected with either SFV or
vaccinia virus. Atransfected CATgeneunder theregulation ofan
earlypromoter(SFV T1) isexpressedwhenviral DNAreplication andlate-geneexpressionareblockedwitharaC (40 ,ug/ml).
with pUC19, which is replicated butnot resolved by
virus-infected cells (Fig. 7c, lane 2). Since the remaining SFV
sequence in pSCX-1 had not been assayed for promoter
activity,wefusedonehalf ofthepSCX-1inverted repeatto
a CAT reporter gene in both orientations, as was done
previously with the native SFV TRT, and used this as a
vectorfor transientassayinSFV-infectedcells.ThepSCX-1
fragment was also found to be capable of directing CAT
expression inan orientation-specific manner (Fig. 7d).
Ex-amination of the SFV sequence in pSCX-1 yielded no
regionswhichexactly fitthe TRTconsensus sequence, but
5' deletionanalysisidentified the sequenceTACGTTTACA CCTATATAAAT as a strong late-promoter elementin the transientexpressionassay(datanotshown).The 3'Tof the TAAAT of this sequence maps 96 nucleotides upstream of the XhoI site which forms the symmetry axis ofpSCX-1,and
sothepositionandorientation of this cTRTarevery similar
to those of the native TRT in the viral replicative interme-diate. Notice that since thetwostrandsatthesymmetryaxis of the resolution target segregate intothedaughter hairpins
(16, 34), the minichromosome generatedfrompSCX-1 will
represent viral telomeres with theXhoI sequences now at
the hairpin turnaround. Furthermore, since pSCX-1 is a
perfect palindrome,therecanbenoflipandflopisomersand
CON STRUC-JT 1. pUC-CAT 2. pCST-1 3. pSD19-CAT
Tis I DM FER E.
RESQLUTI1N ND
+
.4.
is.
I rda B., I I==kb-. i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.56.296.570.673.2]a
TTTTTTTCTAGGG TTATAAATTA
fVV
S TK
- ,primer 4
.-~
I-I
CAT
O. TK4
b
Heterogeneous 5' polyadenylation
.;i
7l.
I_i
9
., v3' IA
T A T T A A T A. T T 5
FIG. 6. Arecombinant vaccinia virus (VV)containingthe TRT sequenceof SFVdirects theexpression ofaCATreporter gene. (a)The plasmid pVCST-1wasused to constructtherecombinant vaccinia virus vCST-2awith theTRT-regulated CATgeneinsertedintothevaccinia virusTKgene. The relative position and orientation of the primer used for the RNAanalysisareindicated, and the apparent RNA start sites aremarked withasterisks. (b) Primer extension analysis oftheTRT-specificRNAsproduced late in infection with vCST-2a. End-labeled primerswereextended in thepresenceofdeoxynucleotideordideoxynucleotide ddCTP(lane 1),ddATP (lane2),ddTTP (lane 3), orddGTP (lane4) or inthe absence ofdideoxynucleotides(lane 5). Thenucleotide sequence of the DNA template is shown, with + 1indicatingthefirst nucleotide complementarytothe RNA. Lane 4 wasunderloaded,butthe sequenceofinterest isstillvisible.
all ofthe termini of pSCX-1 minichromosomes would be
identicalin sequence.
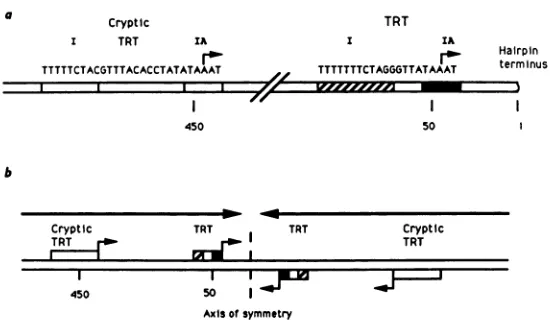
The relative positions of the TRT and the cTRT in the
matureandreplicative intermediate forms of the SFV telom-ere areillustrated inFig. 8. To date, we have been unable to
observe RNAtranscripts initiating at the TRT or the cTRT
sites when these sequences are at their normal telomeric
locations. It remains to be determined whethertranscription
and resolution are interdependent or possibly mutually
ex-clusive activities of the same sequence.
DISCUSSION
We have undertaken an investigation of the relationship
between viral transcription, which synthesizes RNA, and the resolution of poxvirus replicative intermediates, which is
asite-specific DNArecombination event. The involvement
oftranscriptionaleventsin theprocess of DNAreplicationis
well known from work on the mammalian papovaviruses
such as simian virus 40 and polyomavirus, in which
tran-scriptionalelementsareknown to berequiredfor the
initia-tionofDNAreplication(17, 23). Similarly,transcription has
been shown to "silence" replication origins (41) and in
eucaryoticcells may be involved in the selectionof
replica-tionorigins (24) and the termination or blocking of
replica-tion forks (7). To aid our investigation, we have taken
advantage of the previous observationthat clonedinverted
repeat versions of poxvirus telomeres are replicated and
resolved into linearhairpin molecules when transfected into
virus-infected cells (16, 33). Similarly, the regulation of viral promoters can be studied by transient assays in infected cells
and in recombinantpoxviruses. Both transcriptionand
res-olution are uniqueto infected cells, and it is believed that
virus-encoded factors areresponsible for bothactivities.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.160.475.74.472.2]Symmetry
Axis
Acc TRT TRT Acc
I. .m..I _ l
1.pSAB-67
2.pSXB-102
Cia
(ho I Xn0
. .=
Cryptic
TRT Xho
m-l
CryofIc
Xho TRT I X
0 8Kb
3.PSCB-Ia
Cia N I l "la
4.PSCX-I0l45K
b
2 3 4
-T
,V.'w-~~~~40
_- 40 -D
0
C
CA"'ac:I----, cTRT rAT
__
S
_ -M
wt A--C
20
FIG. 7. Identification ofacTRT sequence in SFV which is capableof acting as a late promoter and a resolution target. (a) Plasmids in which the SFV termini have been cloned in an inverted repeat configuration to mimic the replicative form of the virus. pSAB-67,pSXB-102, andpSCB-la consist of various lengths of viral telomere sequence, and all include the conserved TRT near the axis of symmetry of the inverted repeat. pSCX-1 was generated by the removal of a 0.7-kbXhoIfragment from the central axis of plasmid pSCB-la. Ligation of the resultingClaI-XhoIfragments produced a 0.9-kb perfect palindrome of viral DNA with a new central-axisXhoIsite andlacking the native
TRT.(b)Southernblot ofDpnI-resistantDNAharvested from SFV-infected cells which were transfected with pSAB-67 (lane 1), pSXB-102
(lane 2), pSCB-la (lane 3),orpSCX-1 (lane 4). The blot was hybridized with nick-translated pUC1932P-labeledDNA sothat the monomer andmultimer forms of only the plasmid DNA constructs are visualized. (c)Southernblot illustratingthe lethal effect of a point mutation within the criticalTRTboxIAsequence.DNAwasharvested from infected cells which had been transfected with pSD-19 (wild-type TRT [wt];lane 1), pUC-19 (no TRT) (lane 2), orpCST-1/A207*C (lane 3). (d) CATassay demonstrating the ability of a single copy of cTRT to act as a promoter. Lane1,ClaI-XhoIfragment of pSCX-1 fused to CAT such that theClaIsite forms the 5'end of the insert; lane 2, same vector with thepSCX-1 fragment intheopposite orientation.
A significant relationship between late transcription and
telomere resolution can be drawn from a number of
obser-vations. In this work we show that the TRT sequence
responsible fortelomere resolution in SFV is alsocapableof
acting as a promoter at late times during infection. This
capacity to drive transcription has been observed both in
transient assays and when the sequenceisintegratedwithin
the TK gene ofa recombinant viral genome. All mutated
versions of the TRT thus far investigated (point mutations
and 3' and 5' deletions) showa direct correlation between
the ability of a single copy of the TRT to function as a
promoter and theabilityof the inverted repeatconfiguration
to supporttelomere resolution intohairpin termini. Domain
analysis indicatesthatthe conserved regions of the known
TRTsequencesfrom SFV and severalorthopoxviruses(Fig.
lb) arevery similartothe consensus sequenceof viral late
promoters (Fig. 2). The overall variation in the catalog of
sequences of characterized late promoters may indicatethe
needfor the genes which they regulate to be expressed at
different levels (12). On the other hand, the apparent
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.138.470.69.493.2]Cryptic
I TRT IA
roT
TTTTTCTACGTTTACACCTATATAAAT
TRT
TTTTTTTCTAGGGTTATAAAT
450 50 1
b
Cryptic TRT TRT Cryptic
TRT r ~ r~ I TRT
_I
II~p450 50 1
Axlsotsymmetry
FIG. 8. Organization of the TRT and cTRT within the telomere of SFV in the mature hairpin form as packaged in the virion (a) and in the inverted repeat replicative intermediate configuration (b). Thearrows indicate the RNA start sites which have been determinedby primer extension (TRT) from constructs containing fusions to the reporter CAT gene. Whether these RNAs are synthesized from their telomeric locationinSFV during resolution in vivo remains to be established.
quenceconservation of the TRTdomainsmaybeareflection
of therequirement foraspecificdegree ofpromoteractivity
to obtain an optimal amount of telomere resolution at the
propertimeduring the viral replicative cycle. Thisargument
is supported by the observation that another late promoter
sequence, denoted cTRT, that maps over 0.4 kb from the
native viral terminus will also function to resolve inverted
repeats. The cTRT is less than 50% identical to the native TRTintermsof primarysequenceandyet supportstelomere
resolution at an appreciable efficiency, whereas all other
control palindromes tested are uniformly negative. Thus
telomereresolution mightbe atleastpartly regulated bythe
affinityof the TRTsequencefor transcriptionfactors which
areavailableatlate times duringinfection. Thesimilarityof the TRT sequence to late promoters thus suggests a novel
mechanism for limiting DNA processingevents toa
partic-ulartemporal stageofthe viral lifecycle.
The terminal regions ofthe vaccinia virus DNAgenome contain alarge region ofnoncoding repeats. Nevertheless,
vaccinia virus transcripts oriented towardtheterminus have
been detected in thisregion (40).The SFV terminicontaina
series of nine tightly packed open reading frames, all of whichareexpressedatearlytimes,with transcription being directed toward the telomeres (29). The Ti open reading frame is thegene mostproximalto the SFV terminus, and three signals to terminate early transcription have been identifiedbetweenTi and thehairpin (49).Thisclustering of termination signals may be a fortuitous observation or it
could be interpreted as an indicator of the importance of
preventing early transcription from entering the noncoding
region of thetermini.
Transcription of the viral telomeres alone is clearly not
sufficient for the resolution ofinverted repeats. The struc-tural requirements that the replicative intermediates be
nearly perfect palindromes with the TRT sequences in a
particular orientation and within a specific distance ofone
anotherhave been previouslydemonstrated (31, 32).Thisis furtherillustrated by the observation that the cTRT identi-fied in SFVis ahomolog ofa vaccinia virus sequencethat
directs transcription into the telomere at late times (40).
However, the SFV cTRT apparently has no effecton
telo-mereresolution untiltwoopposing copiesaremoved within
100 bp ofone another in an inverted repeat configuration.
Genetic studies of vaccinia virus which have attempted to
determine thespecificallelesimportantfor telomere resolu-tion have identified only mutants which are defective in someaspectoflate-gene expression (13, 35). This hasbeen
interpretedas anindication thatsomelate-geneproduct(s)is
essentialfortelomereresolution(35).Mostofthesemutants show defects in RNA polymerase, and it could also be
arguedthat the lack of telomere resolution results fromthe
defectin either late-RNAsynthesisorevensimplypromoter
recognition byelementsof the late-transcription apparatus.
One of themutantswhich isdefective in telomere resolution butcompetentfor latetranscription, ts22, has been charac-terized elsewhere (13, 35, 39). This mutation showsa
reso-lution-negative phenotype and is defective for the
produc-tion oflate-gene productsbecauseofdegradationof the viral mRNA. Another mutant, ts9383, is defective for resolution andhas been mappedto oneof the subunits of thecapping
enzyme (13).
While late transcription of the telomeres appears to be linked insomefashion totheresolution ofreplicative
inter-mediates, the question of the precise role played by RNA
synthesisremainsopen.It has beenshown thattranscription
in vivo can transiently alter the local topology ofthe
tem-plate DNA (21, 27, 48). Specifically, it has been
demon-strated that themovement ofan RNApolymerase complex
along duplex DNA can inducetwo supercoiled domains of
opposite signwhich balanceoneanother. Thegeneration of
positive superhelicaldensity whichoccurs aheadofa
tran-scription complexispresumably relaxedbytheactivityofa
topoisomerase. Indeed, it has recently been shown that in
eucaryotic cells, topoisomerase I may operate in close
conjunction with RNA polymerase II and is localized to
actively transcribed genes (46). However, an absolute
re-quirement for the relaxation of tension accumulated in
transcribed DNA hasyet tobeestablished,and
topoisomer-aseinhibitionstudiessuggestthatsuchanabsolute needmay notexist(43). Itis known thattranscriptionwith its
associ-ated superhelicaltensioncan proceed through palindromes
without causing the extrusion ofcruciform or other major
topologicalalterations (36). However, theeffect of creating
transcriptsfrompromoterswhicharearrangedheadtohead,
such as occurs at poxvirus telomeres, has not been ad-dressed.
The detailedsignificanceofconvergent promoters to
telo-mereresolution remainstobeworkedout.One model which
Hairpin terminus
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.174.449.72.232.2]canbe envisaged involves theformation of open complexes by an RNA polymerase complex simultaneously on either side of an inverted repeat. This could result in sufficient unwinding of the DNA helix to allow the formation of a
secondary structure such as a Holliday junction analog
which could be cleaved by recombinationactivities encoded
by poxviruses (19). A model for cruciform extrusion has
been proposed to include site-specific unwinding by a
TRT-specific helicase activity (31). The present data suggest an
alternative activity, namelypromoter-dependentunwinding,
which could accomplish the same goal. Confirmation of the
mechanism for resolution will require an in vitro resolution
system,which this work indicates must include at least some
elements of the late-transcription machinery.
The utilization of promoter sequences to resolve
replica-tive intermediates is an elegantexampleof howviruses have
evolved to utilize a preexisting element for multiple
func-tions. It has been shown that in some eucaryotes, actively
transcribed DNA is preferentially involved in homologous
recombination (47). Similarly a strong correlation between
active transcription and site-specific recombination of the
immunoglobulin kappa genes hasbeendemonstrated (6, 44).
However, webelieve that this is the first example of
tran-scriptional events implicated in activating a site-specific
recombination event in eucaryotic viruses.
ACKNOWLEDGMENTS
We thank C. Upton, R. Condit, and R. Morgan for useful and stimulating discussionsand R. Maranchuk for technical assistance. This work was supported by salary stipends from the Alberta Heritage Foundation for Medical Research and by an operating grant to G.M. from the MRC ofCanada.
REFERENCES
1. Baroudy, B. M., S. Venkatesan, and B. Moss. 1982. Incom-pletely base-paired flip-flop terminal loops link the two DNA strands of the vaccinia virus genome into one uninterrupted polynucleotide chain. Cell28:315-324.
2. Berns, K.I. 1990. Parvoviridae and theirreplication, p. 1743-1763. In B. N. Fields (ed.), Virology, 2nd ed. Raven Press, New York.
3. Bertholet, C., E. V. Meir, B.Heggeler-Bordier, and R.Witteck. 1987. Vaccinia virus produces late mRNA by discontinuous synthesis. Cell50:153-162.
4. Blackburn, E. H. 1990. Telomeres: structure and synthesis. J. Biol. Chem. 265:5919-5921.
5. Blackburn, E. H., and J. G. Gall. 1978. A tandemly repeated sequence at the termini of the extra chromosomal ribosomal RNA genes inTetrahymena. J. Mol. Biol. 120:33-53.
6. Blackwell, K. T., M. W. Moore, G. D.Yancopoulos, H.Suh, S. Lutzker, E. Selsing, and F. W. Alt. 1986. Recombination be-tween immunoglobulin variable region gene segments is en-hanced bytranscription. Nature (London) 324:585-589. 7. Brewer, B. J., and W. L. Fangman. 1988. A replication fork
barrier at the 3' end of yeast ribosomal RNA genes. Cell 55:637-643.
8. Cavalier-Smith, T. 1974. Palindromic base sequences and the replication ofeukaryotic chromosomes. Nature (London)250: 467-470.
9. Chambers, S. P.,S.E.Prior, D. A.Barstow, and N. P. Minton. 1988. The pMTLnic- cloning vectors. I. Improved pUC poly-linker regions tofacilitate the use of sonicated DNA for nucle-otidesequencing. Gene68:139-149.
10. Chirgwin, J. M., A. E. Przbyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
11. Condit, R. C., and A.Motyczka.1981. Isolation andpreliminary characterization oftemperature sensitive mutants of vaccinia
virus. Virology113:224-241.
12. Davison, A. J., and B. Moss. 1989. The structure of vaccinia virus late promoters. J. Mol. Biol. 210:771-784.
13. Delange, A. M. 1989. Identification of temperature-sensitive mutants ofvacciniavirusthat aredefective intheconversion of concatemeric replicative intermediates to the mature linear DNA genome.J. Virol. 63:2437-2444.
14. Delange, A. M., andG.McFadden.1987. Efficient resolution of replicated poxvirus telomeres to native hairpin structures
re-quirestwo invertedsymmetrical copies of a core target DNA sequence.J. Virol. 61:1957-1963.
15. Delange,A. M.,andG. McFadden. 1990.Therole of telomeres inpoxvirus DNAreplication. Curr. Top. Microbiol. Immunol. 163:71-92.
16. Delange, A. M., M. Reddy, D. Scraba, C. Upton, and G. McFadden. 1986.Replicationandresolution of clonedpoxvirus telomeresinvivogenerateslinearminichromosomes with intact viral hairpin termini. J. Virol. 59:249-259.
17. Depamphilis, M.L. 1988. Transcriptional elements as compo-nentsof eukaryoticorigins of DNA replication. Cell 52:635-638. 18. Ellis, T. H. N.,and A. Day. 1986. Ahairpin plastidgenome in
barley. EMBOJ. 5:2769-2776.
19. Evans, D. H., D.Stuart, and G.McFadden.1988.High levels of genetic recombination among co-transfected plasmid DNA in poxvirus-infected mammalian cells. J. Virol. 62:367-375. 20. Franke,C. A.,C. M. Rice, J. H. Strauss, and D. E. Hruby. 1985.
Neomycinresistanceas adominant selectable markerfor selec-tion and isolation ofvaccinia virus recombinants. Mol. Cell. Biol. 5:1918-1924.
21. Giaever,G. N., and J.C. Wang. 1988. Supercoiling of intracel-lular DNA can occurin eukaryotic cells. Cell 55:849-856. 22. Gonzalez, A., A. Talevera, J. M. Almendral, andE. Vinuela.
1986. Hairpin loop structure of African swine fever virus. Nucleic Acids Res. 14:6835-6846.
23. Hassell, J. A., W. J. Muller, and C. R. Mueller. 1986. Thedual role of the polyomavirus enhancer in transcription and DNA replication. CancerCells4:561-569.
24. Hatton, K. S., V. Dhar, E. H. Brown, M. A. Iqbal, S. Stuart, V.T. Didamo, and C. L. Schildkraut. 1988.Replicationprogram of active and inactive multigene families in mammalian cells. Mol. Cell. Biol. 8:2149-2158.
25. Kelly, T. J., M. S. Wold, and J. Li. 1988.Initiation of viralDNA replication. Adv. Virus Res. 34:1-42.
26. Kunkel, T. A., J. D. Roberts, and R. A. Zabour.1987.Rapid and efficientsite-specific mutagenesiswithoutphenotypic selection. Methods Enzymol. 154:367-382.
27. Liu, L. F., and J. C. Wang. 1987. Supercoiling of the DNA template during transcription. Proc. Natl. Acad. Sci. USA 84:7024-7027.
28. Macaulay, C., and G. McFadden. 1989. Tumorigenic poxvi-ruses: characterizationofanearlypromoterfromShope fibroma virus. Virology 172:237-246.
29. Macaulay, C., C. Upton, and G. McFadden. 1987.Tumorigenic poxviruses: transcriptional mapping of the terminal inverted repeats of Shope fibroma virus. Virology 158:381-393. 30. Mackett, M., G. L. Smith, and B. Moss. 1984. General method
forproductionand selectionof infectious vaccinia virus
recom-binants expressingforeigngenes. J. Virol. 49:857-864. 31. McFadden, G., D. Stuart, C. Upton, P. Dickie, andA. Morgan.
1988. Replication and resolution of poxvirus telomeres, p. 77-85. In T. Kelly and B. Stillman (ed.), Cancer cells 6. Cold Spring HarborLaboratory, Cold SpringHarbor, N.Y. 32. Merchlinsky, M. 1990. Resolution ofpoxvirustelomeres:
proc-essing ofvaccinia virus concatemerjunctions by conservative strand exchange. J. Virol. 64:3437-3446.
33. Merchlinsky, M., and B. Moss. 1986. Resolution of linear minichromosomes with hairpin ends from circular plasmids containing vaccinia virus concatemer junctions. Cell 45:879-884.
34. Merchlinsky, M., and B. Moss. 1989. Nucleotide sequence required for resolution ofthe concatemerjunction of vaccinia virus DNA. J. Virol. 63:4354-4361.
35. Merchlinsky, M., and B. Moss. 1989. Resolution of vaccinia
on November 10, 2019 by guest
http://jvi.asm.org/
virusDNAconcatemerjunctionsrequireslate geneexpression. J. Virol. 63:1595-1603.
36. Morales, N. M., S. D.Cobourn,and U.Muller.1990. Effect of in vitro transcription on cruciform stability. Nucleic Acids Res. 18:2777-2782.
37. Moss, B. 1990.Poxviridaeandtheirreplication,p.2074-2111. In B.N. Fields (ed.), Virology, 2nd ed. Raven Press, New York. 38. Moyer, R. W., and R. L. Graves. 1981. The mechanism of
cytoplasmic orthopox replication. Cell27:391-401.
39. Pacha, R. F., and R. C. Condit. 1985. Characterization ofa temperature-sensitive mutantof vaccinia virus reveals anovel function thatprevents thevirus-induced breakdown ofRNA. J. Virol.56:395-403.
40. Parsons, B. L., and D. J. Pickup. 1990. Transcription of or-thopoxvirus telomeres at latetimes during infection. Virology 175:69-80.
41. Patel,I.,and D.Bastia. 1986. Areplicationorigin is turned off by anorigin "silencer" sequence. Cell47:785-792.
42. Pritchard, A. E., and D. J.Cummings. 1981.Replicationof the linear mitochondrial DNA from Paramecium: sequence and structureoftheinitiation and crosslink. Proc. Natl. Acad.Sci. USA 78:7341-7345.
43. Schaak, J., P.Schendel, and T. Shenk. 1990. Transcription of adenovirus and Helacell genes in the presence of drugsthat inhibit topoisomerase I and II function. Nucleic Acids Res. 18:1499-1508.
44. Schlissel, M. S.,and D. Baltimore. 1989.Activation of immuno-globulin kappagenerearrangement correlates withinduction of
germline kappagenetranscription. Cell 58:1001-1007. 45. Schwer, B., P. Visca,J. C. Vos, and H. G.Stunnenberg. 1987.
Discontinuous transcription or RNA processing of vaccinia virus late messengers results in a 5' poly(A) leader. Cell 50:163-169.
46. Stewart, A. F., R. E. Herrera, and A. Nordheim. 1990. Rapid induction of c-fos transcription reveals quantitative linkage of RNApolymerase IIand DNAtopoisomeraseI enzyme activi-ties. Cell 60:141-149.
47. Thomas, B. J., and R. Rothstein. 1989. Elevatedrecombination ratesintranscriptionally active DNA. Cell56:619-630. 48. Tsao, Y. P., H. Y. Wu,andL. F.Liu.1989.Transcription driven
supercoiling: direct biochemical evidence from in vitro studies. Cell56:111-118.
49. Upton, C., A. M. Delange, and G. McFadden. 1987.Tumorigenic poxviruses: genomic organization and DNA sequence of the telomeric region of the Shope fibroma virusgenome. Virology 160:20-30.
50. Watson, J. D. 1972. Origin ofconcatemericT7 DNA. Nature (London)New Biol. 239:197-201.
51. Wills, A., A. M. Delange, C. Gregson, C. Macaulay, and G. McFadden.1983.Physicalcharacterization and molecular clon-ing oftheShope fibroma virus genome. Virology 130:403-414. 52. Yuen, L., and B. Moss. 1987. Oligonucleotidesequence
signal-ing transcriptional termination of vaccinia virus early genes. Proc. Natl.Acad. Sci. USA 84:6417-6421.
53. Zakian, V. A.1989.Structureand function of telomeres.Annu. Rev.Genet. 23:579-604.