0022-538X/91/031558-10$02.00/0
Copyright X3 1991, American Society for Microbiology
Variable Expression of
Latent
Membrane Protein in Nasopharyngeal
Carcinoma Can Be Related
to
Methylation Status of the
Epstein-Barr Virus
BNLF-1
5'-Flanking
Region
HU LI-FU,tJANOS MINAROVITS,T CAO SHI-LONG,§ BERTHA CONTRERAS-SALAZAR, LARS RYMO,II KERSTIN FALK, GEORGE KLEIN, ANDINGEMAR ERNBERG*
DepartmentofTumorBiology, Karolinska Institute, Box60400, 10401 Stockholm, Sweden
Received 8August1990/Accepted 6 December 1990
Seven virus-coded proteins, the nuclear proteins EBNA-1 to EBNA-6 and the latent membrane protein (LMP), are regularly expressed in Epstein-Barr virus (EBV)-transformed lymphoblastoid cell lines. In
nasopharyngeal carcinoma(NPC),only EBNA-1 is regularlyexpressed; LMP is detected in about 65% of the tumors. In Burkitt's lymphoma tumors only EBNA-1 is expressed. We have recently shown that the methylation patternsoftheEBV genome varied between these cell types. Invirallytransformedlymphoblastoid
cell lines of normal origin, the EBV DNA is completely unmethylated. In contrast, in the Burkitt's
lymphoma-derived cell line Rael and in a nude mouse-passaged NPC tumor, C15, there was an extensive methylation of CpG pairs. The methylation extended into the coding regions of the two expressed genes, EBNA-1(inboth tumortypes)and LMP(inC15).Twopresumptivecontrolregionswereexemptedfrom this overall methylation: the oriP that contains both an origin ofDNA replication and an EBNA-1-dependent enhancer and the5'-flanking regionof the BNLF-1 openreadingframe that codes for LMP. The latter wasonly
exemptedin the LMPexpressing NPC.We have nowinvestigatedthe relation betweenexpressionof LMP and
methylation ofDNA in the5'-flanking 1kbregion of BNLF-1,coding forLMP. LMPwasmethylatedin3 of 12NPC biopsiesthat did not express LMP but waspartiallyortotallyunmethylatedin theremaining9 that
expressedtheprotein. The three BNLF-1 exons werehighly methylatedin allthe tumors. The oriPregionwas unmethylatedin all the tumors, as in thepreviously studiedRael cell line and nudemouse-passagedNPC.Also, theBamHIWenhancerregion involvedin theexpression ofEBNAnuclearproteinswasmethylated. Noneof
thebiopsies expressedEBNA-2. Ourdata show that theEBVgenomesarehighlymethylated inNPCtumors. The strong reversecorrelationbetweenthemethylationof theputativecontrolregionof the LMP gene and the
expression of LMP suggests thatmethylationhas a role in theregulation ofthis gene.
Seven virus-coded proteins are regularly expressed in Epstein-Barr virus (EBV) transformed lymphoblastoid cell
lines(LCLs): the nuclear proteins EBNA-1 toEBNA-6and thelatent membrane protein (LMP) (8, 9,18-21, 24, 30, 32). In nasopharyngeal carcinoma (NPC) only EBNA-1 and
LMP were detected (14, 43), and in Burkitt's lymphoma (BL) tumors and phenotypically representative cell lines
onlyEBNA-1 wasdetected (13,35, 36). Sofar, EBNA-1 has been reported in 75 NPC biopsies, whereas LMP was only detected in 45 of 80 (56%) NPC biopsies. The biopsies showed similar expression patterns, irrespective of
geo-graphical origin (North andEastAfrica, China, Malaysia,or
the United Kingdom), although more of the Chinese NPC
biopsies (71%)thanthoseof EastorNorthAfrican origin (25
to 62% in the two published studies) expressed LMP.
EBNA-2 to EBNA-6 could not be detected in any of the
biopsies.
Wehaverecently shown that the
methylation
of theEBVgenomes differs among the three EBV-carrying cellular
* Correspondingauthor.
t Permanent address: Shanghai Cancer Institute, Shanghai
200032,People's Republic of China.
t Permanent address: Microbiology Research Groupof the Na-tionalInstitute of Hygiene, Budapest, Hungary.
§ Permanent address: ShanghaiMedical University Tumor Hos-pital, Shanghai,People's Republic of China.
brPresentaddress:DepartmentofMedicalBiochemistry,
Gothen-burg University, 40033 Gothenburg,Sweden.
prototypes. InEBV-transformed B-cell lines ofnormal
ori-gin (LCLs), the viralgenomes areunmethylated (12). In the
BL-derived cell line Rael and in the nude mouse-passaged
NPC C15 tumor, CpG pairs of the EBV genomes were
extensively methylated. The exons of expressed genes,
EBNA-1 and LMP, were methylated as well, but two
potential control regions were unmethylated: the oriP that
contains one origin of DNA replication and an EBNA-1-dependent andEBNA-regulating enhancer (31, 38) and the
5'-flanking regionofBNLF-1thatcodes for theLMP(inthe
NPC, which expressed LMP). Rescue of the virus from BL andfrom NPC into LCLs has led to demethylation of the viral DNA in allderived lines.
DNAmethylationcanparticipatein the controlof
eukary-oticgenes(6, 10). The cause-effectrelationshipis
controver-sial (6, 11, 23). Recent studies have shown that the methyl-ation ofspecific CpG pairswithin theDNA-binding motifsof transcription factorscanpreventtheir binding and downreg-ulate thetranscriptionofanadjacentgene(2,28, 41).Hence,
the methylation ofspecific sitescan directly influencegene
expression and hypomethylation of 5' promoter-enhancer
regions is often correlated with expression, whereas
meth-ylation of structuralgenesis not(6).
The cell phenotype-dependent differences in EBV DNA
methylation and the apparent correlation with viral gene
expressionsuggestthatmethylation is involved in viralgene
control in latently infected cells (12). This suggestion was
tested in thepresent paperby focusingonthefact that only about56% of NPCsexpressLMP(14). Wehaveinvestigated 1558
on November 10, 2019 by guest
http://jvi.asm.org/
the methylation of CpGs in BNLF-1 and in 5'-flanking
regions. The putative EBNA-controlling region in oriPand the enhancer in the internal repeat 1 have been studied in parallel. We have found a strongcorrelation between LMP expression and lackof methylation of the 5'-flanking region ofBNLF-1, extending1 kbupstreamof the promoter. This correlation is in line withtherecently discovered regulating
elements in this region involved in LMP control (12a, 17). The correlation between LMP expression and the
methyla-tion status of the LMP regulatory sequence suggests a
modulating role of methylation or, alternatively, that the
methylation status depends on the activity of the LMP
regulatorysequence.
MATERIALS AND METHODS
Cell lines and tissue culture. Rael isaBL-derived cell line
(27) with a stable type I BL phenotype, expressing only
EBNA-1 but not EBNA-2 to -6 or LMP. The CBC- and
NAD-C15-STO cell lineswereestablishedby transfer of the
virus carried by the NPC designated C15, serially
propa-gated in nude mice, to normal B cells (12). All lines were
grownin RPMI 1640 medium(GIBCO Biocult,Long Island,
N.Y.), supplemented with 100 U ofVpenicillin perml, 100
,ug of streptomycin per ml, and 10% fetal calfserum. The
cells werefed every 3 to4 days and cultured in incubators with 5% CO2 atmosphere andhigh humidity.
Biopsies. NPC biopsies were doneby local surgery in the
Shanghaiarea. The tumorswerestored frozen (-70°C) until
prepared for DNA or protein blotting. CAO is a Chinese
EBV DNA-positive NPC tumorpassaged in nude mice (20
passages [5]). C15, C17,andC18 arenudemouse-passaged, EBVDNA-positive African NPC tumors, analyzed after 15 to 50passages (4). The tissues were prepared for
immuno-blottingas describedearlier(14).
Immunoblotting. Immunoblotting was performed as
de-scribed earlier (13, 14). A previously characterized serum
samplefromahealthy EBV-seropositive donorwasusedas the antibody source fordetection of EBNAs (VCA, 1:320; EAD, <1:10; EAR, 1:320; EBNA, 1:320; and EBNA-2, 1:80). Alternatively, wealso used the seraofaCLLpatient PG (VCA, 1:1,280; EA(D), 1:640; EA(R), 1:20; EBNA-1, 1:320; and EBNA-2, 1:80). EBNA-1 was detected by a humanserumthat has beenaffinitypurified on an
EBNA-1-specific polypeptide (no. 107) (14). LMPwasdetectedbythe
S-12 monoclonal antibody (MAb) provided by D.
Thorley-Lawson(DepartmentofPathology, TuftsUniversity School ofMedicine, Boston, Mass.). The humanserumsamplewas used at a 1:20 dilution and the S-12 MAb was used at a
1:1,000 dilution of ascitic fluid. Alternatively, a pool of MAbsagainst LMP, kindly suppliedby Martin Rowe(CRC Laboratories, Birmingham, United Kingdom; CS.1 to CS.4
[34]), wasusedata1:1,000dilution. LMP could be detected in a control sample prepared from B95-8 cells when an
immunoblotting lane was loaded with material
correspond-ingto 5 x 103 cells. LMP could not bedetected in samples
withlower numbersof cells. Normallythe lanesare loaded
with material from 105 to 106 cells; LMP would thus be detected if it were expressed in 0.5% of the cells (given a level ofexpression per cell similar to that of B95-8 cells).
This givesan indication of the levelofsensitivity needed to detect LMP in NPC biopsies, although accurate cell num-bers inbiopsies cannotbe determined.
Analysis of DNA methylation. Eukaryotic and viral DNA
are predominantly methylated in CpG pairs. Methylation
was analyzed by digesting high-molecular-weight cellular
m021
-36
T
916T
- I . I
b.
C.
132151 I 1162871
__ilU
11
ill557r
1llllllllllllllmlllm,!__
d.
111126219
1132 III ,, ,,,, in,529,
1L
.... ..
_:~ .
e.
11671304 -§67i It494 ,,
f.
357, 818 ,
L.6
169424
506 ,,iA.
1722
,,213,, nl .i,i1
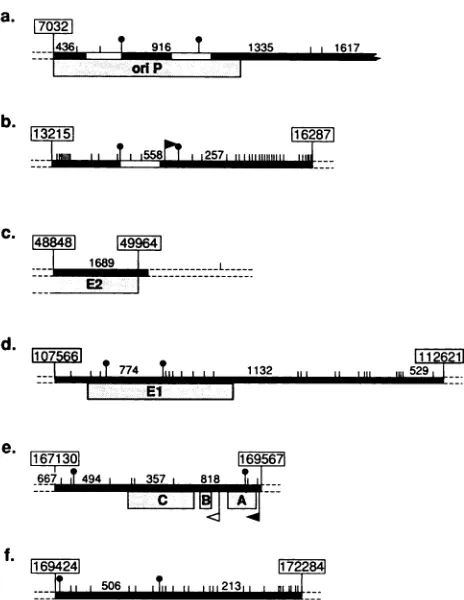
FIG. 1. DNA probes used to study methylation by Southern blotting. Horizontalblacklines(_)between boxeswithnumbers (e.g.,7032)showprobes usedinthisstudy.Inseveral casessmaller probes(t)wereused as an alternative. Small vertical barsindicate restriction sites detectedby theisoschizomerMspIorHpaII (in the latter caseonlywhen theCpGwasunmethylated). Major fragments resulting fromcompletedigestion withthese enzymes areindicated by size (in base pairs). Identified enhancers and EBNA-1-binding regions have been indicated within the probes (IZ1). The major latent promoters (
r)
and the early promoterfor the latentmem-brane protein
(PI)
areindicated. oriP and the exonsforEBNA-1, EBNA-2,and LMP areindicated beneath theprobes (L ).(a)oriP region in theBamHI C fragment. In particularthe916-bpfragment within oriP, cleaved out by MspI (t) was used. (b) BamHI W fragment, with theAvaIfragmentusedas a probetodetectthe W enhancer (t). The figure shows the first of a series of identical BamHI W fragments. (c) Probe used to detect the 3' part ofthe EBNA-2-coding exon. (d) BamHI K fragment, with a smallerMspl-MspI probewithin the repeatregionof BKRF-1(t). (e) Probe recognizingthe LT 3region,obtainedby digestion oftheEcoRIDhet fragment with XhoI. A, B,andC showthethreeexonsforLMP.(f) Twoprobes foranalysis ofthe5'-flankingsequenceofLT3, either as anXhoI-EcoRl fragmentorobtainedbydigestionwithBgIl (t). The maps andcoordinatesgivenare basedonthe B95-8 sequence and onmolecular weight markers included in our blots (HindIII-cleaved lambda DNA anda1-kb ladder(BethesdaResearch Labo-ratories).
and viral DNAs with either HpaII (specificity, CCGG) or
HhaI(specificity, GCGC). For each 1 pgofDNA, 10 U of
enzymewasaddedtocompletedigestion. TheHpaIIdigests were controlledbyparallel digestion withMspI, an isoschi-zomerwhich isnotinhibitedby
methylation.
The restrictionenzymes were obtained from Amersham
(Amersham,
En-gland). Digestion was performed
overnight
or for longertimes in the
enzyme-specific
buffersprovided by
the manu-facturer.9 I F4-99-6-4.
1689
.--T
774Till
lt.c. 1101
:d
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.320.552.67.367.2]r CM CO) f L) to r~- o O7)rYO r-v
00
N ( 0)
rrv-m
C
116.
-#, .
97
-68
-...
-EBNA- I
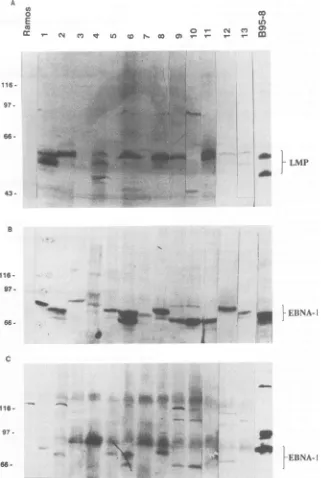
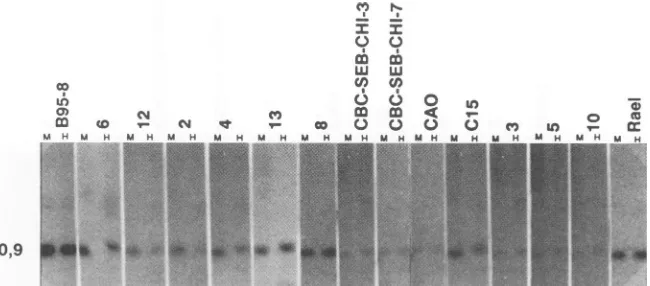
FIG. 2. Immunoblots showing expression ofEBVproteinsinNPCbiopsies. (A) Detection ofLMPwithS-12 MAbs.Biopsies 3, 5, and 10 werenegative, while expressionwasweak butpositivein 7, 12,and 13. (B) Detection ofEBNA-1 byamonospecific human EBNA-1 serum,obtainedbyaffinity purificationwithanEBNA-1polypeptide.(C) Detection ofEBNAswithapolyvalentEBNA-positiveserum(PG).
Ramos wasused as anegative control, and B95-8 was used as apositivecontrol. Thepositionsof standard molecularweightmarkersaswell asthe positions of EBNA-1 and LMP are indicated.
ThedigestedDNAswereseparatedon a1.5% agarose gel
(Bethesda Research Laboratories, Inc., Bethesda, Md.) in
Tris-borate-EDTA buffer (TBE; 89 mM Tris base, 89 mM
boricacid, 2 mM EDTA). Electrophoresis was carried out
for 13hat50 V.Digestionwaschecked by ethidium bromide
staining. Denaturationwascarried out in 0.5 M NaOH with
1.5 MNaCifor1h.The DNA was then neutralized for 1 h in
1 M Tris-1.5 M NaCl, pH 7.5. Transfer to nitrocellulose
(HybondN;Amersham) was done in 20x SSPE (Ix SSPE is
0.18 M NaCl, 10 mM sodium phosphate, and 1 mM EDTA
[pH 7.7]) overnight. Prehybridization was performed with
6x SSPE-1% sodium dodecyl sulfate (SDS)-100 p.g of
herring sperm DNA (Boehringer Mannheim) per ml-lOx
Denhardt solution(0.2% Ficoll, 0.2% polyvinylpyrrolidone, 0.2% bovine serumalbumin) at42°C overnight.
Theprobes were labeled with[32P]dCTP (Amersham) by
random primer oligolabeling with the Klenow fragment of Escherichia coli DNA polymerase (16). Hybridization with
specific probes was performed at 42°Cfor 16 to 24 hin 6x
SSPE-100 ,ug of herring sperm DNA-50% formamide-1%
A
CA
0
E
(U
116 -97.
66
-43.
a
a
.4
.A_
B
- LMP
116-97
-66--!F
ip
lEBNA-Ion November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.151.472.73.551.2]TABLE 1. Designations and EBV protein expression in biopsies and cell lines used inthis study
Expression
Cell(origin)a Reference
EBNA-1 EBNA-2to -6 LMP
1(ChNPC) + - +
2(ChNPC) + - +
3(Ch NPC) + -
-4(Ch NPC) + - +
5(Ch NPC) + -
-6(Ch NPC) + - +
7(Ch NPC) + - +
8(Ch NPC) + - +
9(Ch NPC) + - +
10 (Ch NPC) + -
-11(Ch NPC) + - +
12(Ch NPC) + - +
13(ChNPC) + - +
C15(NuMo NPC) + - + 4
CAO (NuMo NPC) + - + 5
B95-8 (MarmosetLCL) + + +
Rael(BL) + - - 29
CBM1-Ral-STO(LCL) + + + 12
CBC-SEB-CHI-3 (LCL) + + +
CBC-SEB-CHI-7 (LCL) + + +
a Ch, Chinese; NuMo, nude mousepassaged; B95-8, marmoset B cells
transformed by infectious mononucleosis-derived EBV;CBC, cord
blood-derived cells. Rael and CBM1-Ral-STO carrythe samestrainof EBV.
C) Q
wU LuI
04 V) m m
Ut) C
msM U ('_T- 5¢Rt T- o0*
0
M H MH M H M H M H M H M HM H M H
4,0-2,0
1,6-'@ @ *
*
*
1,0-0 aS
a 0
0,50- a
0,39- _ . _ S
0,34
-0,30- - * * 0
dextran sulfate containing the radioactive probe. Subse-quently the filters were washed in 2xSSPE-0.1% SDS for15 min at room temperature, two times in 0.2x SSPE-0.1% SDS for30 min at 65°C, and finally for 5 min in 0.lx SSPE. The filters werefinally exposed to Fuji medical X-ray film for 4 to48 h at 90°C and then developed (12).
Probes. The following probes were used (Fig. 1): the BamHI K fragment; the pMBamH2 fragment (cloned from the M-ABAcell line, kindly provided by Georg Bornkamm), which covers the 3' part of the EBNA-2A exon BYRF-1 (coordinates 48848 to 49964); a MspI-MspI digest from the BamHICfragment(coordinates 8189 to 9105), coveringpart
of oriP;aMspI-MspIfragmentfrom the BamHI K fragment, covering part of the EBNA-1 exon BKRF-1 (coordinates 108161 to 108924); an XhoI-XhoI fragment from the EcoRI
Dhet
fragment covering most of the LMP exons (coordinates 167484 to 169424); an XhoI-EcoRI fragment (coordinates 169424 to 172281) and a BglI-BglI fragment (coordinates 169449 to 170290), both covering the promoter and5'-flanking sequences of LMP; and an AvaI-AvaI fragment
covering the BamHI W enhancer (coordinates 13827 to
14460).
RESULTS
Expression of EBV-encoded proteins. Thirteen Chinese
NPC biopsies fromtheShanghai area were included in this
study. They were EBV DNA positive, as shown by Southern blotting (for details, see below). Expression of the EBNAs and LMP was assayed by immunoblotting by using the polyvalent human serum, PG (Fig. 2C), the affinity-purified
O ul z
4 . o X
o) 0 co n _ a:
M H M H M H M H M H M H
it
4D
d
i
_,_A *
*
-0
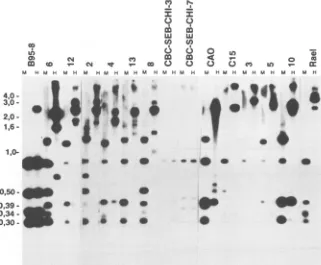
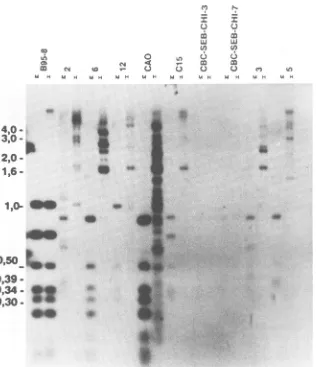
FIG. 3. Methylationpatternofthe BNLF-1exonsinNPCbiopsies.Biopsyandcontrol cell line DNAscleaved withHpaII(H)orMspI
(M) and probed with an XhoI-XhoI probe (Fig. le). Biopsies 3, 5, and 10 were LMP negative. As controls, B95-8 (predominantly
unmethylated), Rael(methylated), andtwoEBV-transformedLCLs(CBC-SEB-CHI-3and-7,unmethylated)wereused. Thepositionsof the molecularweightmarkerfragmentsareindicated.TheeasilyidentifiedmajorB95-8fragmentscanalsobe usedasreferences(357,494, and 818bp).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.149.470.412.677.2]CM
w W- N q'
M H M H MH MH
.,... '...
.W.
CII)
NI-I I
.
m m
0
W
C/)
T- cO Q
MH M H M H MH
t. . : V s^
.&AL l:-sw
d
e4''
*d&Aw_ _w qo
[image:5.612.146.480.76.376.2].1
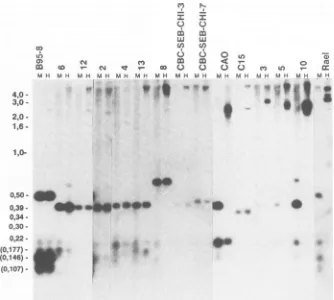
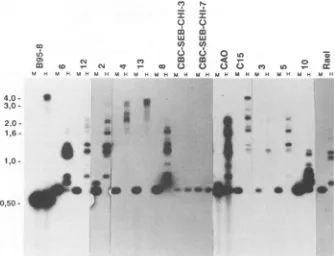
.7FIG. 4. Methylation pattern of the 5' BNLF-1-flanking region. Same blot shown Fig. 3, strippedandreprobedwithaBgIl-BgIl probe
(smallersubfragmentinFig. if),coveringthe promoter andapproximately800bp upstreamoftheLMPexons.SampleDNAs cleaved with
HpaII and MspI. Biopsies 3, 5, and 10 and the control cellline Rael didnotexpressLMP,while all the othersamplesdid. Thepositionsof
the DNAmolecularweight markerfragmentsaswellasthepositionsof three minor B95-8fragments (in parentheses: 177,146,and 107bp; theyare easytoidentify togetherwith thelarger 506-bpfragment)areindicated.
(a) B95-8
5' 3
(b) NPC-12
LT
...t ttt
t
Msp I
LT
...tttT
Hpa II
(c)
NPC-10MspI
Hpa II
0.1 kb
FIG. 5. Schematic drawingof theBNLF-1 region andupstream sequences in twoNPCbiopsies(oneexpressing LMP [b; no. 12] and one notexpressingLMP[c; no. 10]). MspIandHpaIIcleavage sites
areindicated(O).Forreference,the B95-8 map withcleavage sites, LMP exons, andthepromoter isprovided (a).
human serum specificfor EBNA-1 (Fig. 2B), and the S-12
MAbfor LMP(Fig. 2A).The dataonprotein expressionare summarized in Table 1 and are related to those of earlier studies summarized in Table 2. All EBV DNA-positive biopsies expressed EBNA-1, as shownby both the
polyva-lent and themonospecificreagent.Ten of the thirteen(77%)
EBVDNA-positive biopsieswereLMPpositive (no. 1, 2, 4,
6, 7, 8, 9, 11, 12, and 13). Ofthese, no. 7, 12, and 13 gave
relatively weakersignals. Nevertheless, inspectionof these lanes in reflected lightandbytransillumination showed that
theywereclearly positive,whereasnosignalatallcould be detected in no. 3, 5, and 10. Biopsy 7 hada slightly higher LMPmolecularweightthan the others. A small variation in LMP size is common. C15 and CAO have been shown in earlier studies tobe LMPpositive (4, 14).
Methylation of the BNLF-1 readingframes (LMP coding)
and the 5'-flanking region. Twelve of the biopsies were selected for DNA methylation studies. Two nude
mouse-passaged NPC tumors, C15 (4) and CAO (5), were also included. As controls we have included two LCLs
estab-lished from Chinese wild-type EBV (CBC-SEB-CHI-3 and
-7)thatcarryunmethylatedgenomes: thetypeIBL cell line Rael that contains highly methylated EBV DNA and the
virus-producing B95-8 line. The materials and their EBV
antigen expression are listed in Table 1. The biopsy DNA
was cleaved in parallel with HpaII (methylation sensitive)
and MspI (methylation-insensitive isoschizomer) and
hy-bridized with the XhoIprobe (Fig. 1), coveringthe promoter
co
uM)
MH
4,0
-3,0
-2,0
-1,6
-MH
.ift
a)
(a
eI - cc
MHMH MH
.f
W."#t ; v
00Ul
9
-M H M H
1,0-0,50
0,39 -0,34
-0,30
-0,22
-(0,177)
-(0,146)
-(0,107)
-'V.
0
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.61.299.484.680.2]Co
Lh
af)
00
m Co
h Mtb H
eI-
_ m NX~
~T cMCoM. H MW H M H M H M H
;M" S~ |~& .::^l
Mv r%.
I I
CD x C.)Q
w w U)
6
4
Ito
5r
(, U-I
(a
IH
FIG. 6. Methylation of the oriP region in NPC biopsies. Same blot shown in Fig. 3 and 4,strippedandreprobed with the oriP probe (the 916-bp subfragmentdepicted inFig. la). The position of theMspI-MspI916-bp fragment in the control cell line B95-8 is indicated. Sample DNAs werecleaved withHpaII andMspI.
and all three coding exons of BNLF-I and extending over most of the 3'-untranslated region of the LMP-coding
tran-script. The B95-8 DNA showed a largely unmethylated
pattern (Fig. 3, lane B95-8 H) with four major fragments corresponding to the expected lengths of 818, 494, 357, and 253 bp from the sequence data which was comparable with
thefragments obtained withMspI. Therewas also one large
fragmentof around 2.5 kb, suggesting that some genomes in
B95-8 cells were completely methylated over the whole
region covered by the probe. Rael cells, in contrast,
con-tainedonly four large fragments (2.5, 3.0, and 4.0 kb and a
larger one). This reflects a high degree of methylation,
extending to CpG sites outside the region covered by the probe. The four smaller fragments were only seen in the
MspI (M) lane, confirming the absence of any detectable
hypomethylation. TheRael counterpart of the 494-bp B95-8
fragmentwas smaller, about 400 bp. The LCLs
CBC-SEB-CHI-3 and -7carried totallyunmethylated viral DNA in the
whole region. An 818-bp fragment was consistently present, but there was some size variation in the three smaller
fragments. CBC-SEB-CHI-3 had a Rael-like fragment size
distribution, while CBC-SEB-CHI-7 showedfragment sizes
intermediatebetween those of Rael and B95-8 cells. The size
variation of the 357- and 293-bp fragments may conceivably
influence the size of the LMP, since both fragments are
located within the majorBNLF-1 coding exonof LMP. Incontrast toB95-8cells and thetwocordblood-derived
cells, all the NPC tumors showed extensive EBV DNA
methylation.Nudemouse-passaged CAO andbiopsytumors
4, 5, and 10 contained one major unmethylated 357-bp
fragment, corresponding tothe BNLF-1 codingexon. CAO
andtumor4expressedLMP, whiletumors5and 10didnot.
The unmethylated fragment gave a weak signal in three
cases, suggestingthat itwasonlypresentinafraction of the EBV genomes. In biopsy tumor10 the fragment was
equi-molar with the otherfragments. These fourtumors and the
remaining seven showed the presence of much larger
frag-ments, which weretheresultofincompletecleavage due to
methylation protection of restriction sites. The
resulting
fragmentsvaried between0.7and5kb.Mostof themwerein
the size order of 1.5 to 4 kb. They all gave strong signals.
This suggests that theHpaII restriction sitesaremethylated
and thatmethylation extends fromafew hundred base
pairs
to 1kboutside theprobedregion, in most cases. Atleast10
of the tumors showed traces of uncleaved DNA. These
fragments have beenderived from genomes thatwere
meth-ylated over several tens of kilobases outside the BNLF-1 region. This is also supported by the lack of any incomplete cleavage with MspI. Moreover, staining the gels with ethid-ium bromide before blotting showed that good digestion occurred (data notshown). All NPC biopsies contained the 818-bpfragmentafter MspI cleavage; the smaller fragments were of various sizes, as in the cell lines. The 357-bp fragmentwasnotdetectable ineightof the tumors;onlytwo
smallfragments could be seen. Seven tumors contained the
400-bp fragment associated with Rael cells and lacked the
494-bp fragmentassociated with the B95-8 virus.
The LMP-coding exons varied slightly in size between different biopsies.Theywerehighlymethylated in the whole coding region, both in LMP-positive and LMP-negative
tumors. In contrast, agood correlation wasfound between
themethylation statusof the5'-flanking region of the LMP
geneanditsexpression.Theflanking regionwascompletely
unmethylatedinseventumors (tumors6, 12, 2,4, 13,8, and
C15), all of which expressed LMP (Fig. 4). The flanking
region was partially methylated in CAO, which expressed
LMPat alow level. Theregionwasextensively methylated
in tumors 3, 5, and 10, which did not express LMP. The smallestfragmentthat could be obtainedfrom thesetumors
after digestion with HpaII was2.5 kb. Similar tests on the
various control cell lines further corroborated the close
relationship between methylation of the 5'-flanking region
and lackofexpression, orviceversa. The twohumancord blood-derived LCLs and the B95-8 line expressed LMP,
whereas theRael line didnot. TheLMP-flanking regionwas
unmethylated in theformer three but highly
methylated
inRael, asexpected.
The5'-flankingregion oftheLMPgene showed
consider-ablemicroheterogeneitybetween thedifferent virus isolates.
Thelarge,506bpB95-8fragmenthad400-bpcounterparts in
nineof thetumorsandin the cord blood-derived cells.Itwas larger in biopsy 8 and of the same size in Rael. The resolution of the smaller fragments was not sufficient to detect minor variations.The
175-bp
fragment
was about thesame size inallviralDNAsexamined.
Three ofthree LMP-negative
biopsies
showed extensivemethylationof the whole800-bp
region
immediately
5' of theBNLF-1promoterand
extending
outside into theLMPgene.The same region was
completely
unmethylated
in six ofseven LMP-positive tumors. The seventh
(CAO)
waspar-tially methylated around the
large
400-bp
fragment
buton November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.146.471.71.214.2]0
M u MH A H u H
6 6
m
mi
CD U)
UN C)
_ co m
1i H M H Mi H
4,0
-3,0-.
3-2,0
-1,6
-1,0-
am
0,50_
0,39
-0,34
-0,30. A__t
a*
oF 4D [
a.
ggm~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
[image:7.612.158.474.65.432.2]*...e
FIG. 7. Methylation of the BamHI K fragmentandEBNA-1exoninNPCbiopsies. Southern blotshowingNPCbiopsiesand controlcell linesprobed with the whole BamHl K fragment (Fig. ld). Sample DNAs cleavedwithHpaII (H)orMspI(M).Thepositionsofmolecular
weight marker fragmentsareindicated, and themajor B95-8 fragmentscaneasilybe identified(1,132, 774,and529bp).All the NPCsamples show extensivemethylation of all fragments. They allexpressEBNA-1.The control cell lines CBC-SEB-CHI-3 and -7gaveveryweaksignals with thisexposure,although thesameamountof cellular DNAwasloadedinalllanes. The BamHI Kfragmentwasunmethylatedin these
samples.
yielded one or twosmaller fragments, because ofa lack of
methylation of the surroundingrestriction sites.
Figure 5 is a schematic restriction map of the relevant BNLF-1 region and its 5'-flanking sequence in an
LMP-positive (tumor 12) andanLMP-negative (tumor 10) biopsy.
The dashedlines indicate the size variations of these frag-mentscompared with those of B95-8 cells.
Lack of methylationin the oriP region. A916-bp probewas used to analyze methylation in the oriP region. The probe
covers the middle part of the region, with the EBNA-1
enhancerjustupstreamandthe EBNA-1-binding dyad
sym-metry motifin the 3' end (31, 38).
Thesameunmethylated 916-bp fragmentwas detectedin 11tumorsand inthe controls (Fig.6). Inourprevious study
(12) the sameregion was unmethylated in all BL cells and LCLs examined and represented the only known area
ex-empted from high overall EBV DNA methylation in thetype
I BLline Rael.
TheexonofEBNA-1showedahigh degree ofmethylation in alltheNPCtumors (Fig. 7).
Methylationof theBamHI W enhancer.Recently,a
B-cell-specific enhancer in the BamHI W fragments that can influencetheexpressionof EBNA-2wasdescribed(33).It is contained withinanAvaIfragment(Fig. 1).Wescreenedthe DNA from nine NPCtumorsand controls with thisfragment
as alabeledprobeafter MspI andHpaII cleavage (Fig. 8).
All were EBNA-2 negative by immunoblotting. All were
methylatedin this region, althoughnotasextensively as in the LMPregion. Thus,awhole ladder offragments, ranging from 0.9to4 kb, was generated afterHpaII cleavage. The
single 558-bp fragmentfound inunmethylatedDNAwasnot detected inanyofthem,however.AfterMspI cleavage,this
fragment was detected in all tumors. It was slightly larger than that inB95-8cells,around 600bp.As shownearlier,the EBNA-2 exon was methylated in NPC tumors (12). The BamHI W enhancerregion was unmethylated in the LCLs thatexpressedEBNA-2 (Fig. 8).
DISCUSSION
We havefoundan overalldifference in EBV DNA
meth-ylation of NPC tumors and EBV-transformed LCLs of co
u H co U)
U H A H
a.
.s'll
.1
..i;...:...
on November 10, 2019 by guest
http://jvi.asm.org/
Cl XC.
to T- CJ s
-M H M H M H M H M H
C.,)
I
C-)
CD (0)
6
Ca C.)
M H M H
m
C-)
cn
CDO
6
o
m 4: 0 C.)
MN M H
UHM
M HM H MNH
lo
p.
I&
a,
*
.0J6
FIG. 8. Methylation of theBamHI W enhancer. The same blot as in Fig. 3, 4, and 6, stripped and reprobed with an AvaI-AvaI probe covering the BamHI W enhancer (Fig. lb). The DNA samples were cleaved with HpaIl or Mspl. The control cell lines B95-8, CBC-SEB-CHI-3, and -7 expressed EBNA-2 to -6, while all the other samples (NPC biopsies) expressedonly EBNA-1. The 558-bp fragment could easilybeidentified in the control cell line B95-8, and it is slightly larger in all the other samples.
normal B-cell origin. EBV genomes are extensively methyl-ated in theformer and unmethylated in the latter. We have foundasimilar difference previously between the type I BL line Rael and the derived LCLs (12).
In addition to this overall difference, there is a superim-posed, expression-related difference in the 5'-flanking re-gions which is known or suspected to contain regulatory sequences (Table 2).
It is known that methylation of 5'-flanking regulatory sequences mayinfluence gene expression, whereas
methyl-ation ofthecoding exonshas no such effectas arule (3, 11,
25, 26). The recent findings of single, specific methylation sites within the DNA binding motif of several proteins
involvedintranscriptionalregulation show a potential direct
role ofmethylation in gene control (2, 26, 37, 41). A search
for specific critical CpG methylations within the relevant
promoter and enhancer regions in the EBV genome will
obviouslybe of great interest. Our data suggest that
meth-ylation of regulatory regions influences EBNA and LMP control.
The promoter for LMP-coding transcripts has been mapped to the EDL-1 site (coordinate 169546). We have foundacomplete correlation between LMPexpressionand lackofmethylation of the promoter region andsome 800bp upstream of it. This is particularly remarkable since the surrounding EBV DNA is extensively methylated in NPC tumors,includingthe threeLMP-encodingexons. Thesame
region was methylated in all tumors that failed to express
LMP. This suggests that methylation participates in the
regulationof LMPexpression. Direct studies onLMP
tran-scriptional regulation will be necessary to establish any
direct causalrelationship.Theregionthatshowedan expres-sion-correlated methylation pattern coincides with the
re-cently described LMPregulatorysequence (17).
Promotersfor EBNAtranscripts havebeen found in two
[image:8.612.139.474.76.332.2]regions so far, BamHI-C (BCR 2, P11305) and BamHI-Ws
TABLE 2. DNAmethylationand geneexpressionpatterns incodingand control regions
Methylation' Proteinexpression Celltype
oriP EBNA-1 exon BamHI Wenhancer EBNA-2exon BNLF-15'sequence LMPexon EBNA-1 EBNA-2 LMP
NPC(LMP+) - + + + - + + - +
NPC(LMP-) - + + + + + + -
-C15 - + + + + + +
C15 LCL - - - + + +
Rael - + + + + + +
CBM1-Ral-STO - - - + + +
aoriP, Originof latent DNAreplication; EBNA-1exon, BKRF-1 exonfor EBNA-1 protein;BamHI Wenhancer, enhancer5'ofWpininternalrepeat1;
EBNA-2 exon, BYRF-1 codingexonforEBNA-2; BNLF-1 5' sequence, LMPcontrol region;LMP exon, threeLMP-codingexons. +,Methylation; -, no
methylation.
co 0) m
a
0 ci
M H M H
U: a
4,0
-3,0
-2,0
-1,6
-1,0
-0,50
-O
S.
so 1
II
_.
..10lo"m
X,F.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.54.557.609.699.2]TABLE 3. NPCbiopsy protein expressiona
No.expressing:
No.of biopsies Reference EBNA-1 LMP
24 24 9 43
56b 56 36 14
13 13 10 Thispaper
aProtein expressionwastestedbyimmunoblottingwithpolyvalent and/or
monospecific reagents ofEBNA-1 and LMP. EBNA-2 to -6 werealways
negative.
bOnlyEBNA-1-and/orEBVDNA-positivebiopsiesareincluded.
(P14352+nx30739
n = 4 to 11). In the producer line B95-8,several different EBNA transcripts start at the same pro-moter,P11305,whereas EBNA-2transcriptsinanLCL,IB4,
werefound toinitiate at one of the BamHI W promoters. An enhancer that can transactivate after EBNA-1 binding is
localized in oriP 3 kb upstream of the BamHI C promoter (31). The oriP enhancer may be involved in regulation of transcripts from the BamHI C promoter (38).
The oriP region has been unmethylated in all EBV-carrying tissues studiedsofar, includingBL andNPC.This
mayfitwell with thefactthat EBNA-1is the mostregularly
expressed EBVprotein, is found in all EBV DNA-carrying
cells studiedsofar,andisunaffectedbythe cell
phenotype-dependentexpression and regulation.
Recently, it was demonstrated that in a selection of
BL-derived cell lines and invitro-transformed LCLs,either the BamHI W or the BamHI C promoterwasused
irrespec-tive of cell phenotype (42). This study, however, did not
includeanytype I BLcell linesorNPCtumorbiopsies.The
EBNA-1 transcription initiation remains to be identified in these cell types. Hittetal.(22)detected notranscriptsfrom the BamHI W region when screening their cDNA library
derivedfromthenudemouse-passaged NPCC15,suggesting
that EBNAexpressioninthisNPCiscontrolled inadifferent way and without the extensive splicing of the BamHI W
regionsthat has beenseeninB-celllines (Raji, B95-8, IB4,
andJY).
The lack ofmethylation oforiPmay also besecondaryto
its involvementin DNAreplication or may evenresultfrom
theprotection by DNA-binding regulatory proteins,
includ-ing EBNA-1. Recently, an enhancer that is 50 to 400 bp upstreamof the BamHI Wpromoter(33) and thatfunctions
in B-cell lines but not in HeLa cells has also been demon-strated.This enhancer is likely to beimportant for regulation of EBNA expression. The BamHI W promoter-enhancer
was hypomethylated in the EBNA-2-expressing LCLs but
methylated in all the nonexpressing NPCs.
Our study has further confirmed the specific pattern of
EBVprotein expression in NPC: EBNA-1 expressionin all tumors that are EBV DNA positive, and LMP expression
only in a large fraction. This fraction has been repeatedly
higherin Chinese NPCs than in NPC materials from other
high or medium-high endemic regions. Earlier we detected LMP in 71% of Chinese tumors (12), and now we have
detected LMPin 10 of 13 (77%) tumors in two consecutive,
randomly chosen groups (Table 3). This compares with 25 to
62% in earlier investigated tumors from North Africa and
EastAfrica (14, 43).
LMP can transform established strains of rodent
fibro-blasts (1, 40).
F?ahraeus
et al. (15) have recently found thatexpression of LMP in an EBV-negative epithelial cell line
(RHEK-1) changes the morphology of the transfected cells
andinhibits keratinization. Dawsonet al. (7) demonstrated
simultaneously
that LMP inhibitsdifferentiation inahuman keratinocyte cell line. NPC cells are low differentiated oranaplastic.Whethertheexpressionof LMPcanberelatedto
the level ofdifferentiation andtothe clinical course of the
tumorremainsa question.
ACKNOWLEDGMENTS
Weare grateful toDavid Thorley-Lawson and Martin Rowe for providing LMP MAbs, to Georg Bornkamm for providing EBV DNAclones, toMaria GraziaMasuccifor fruitful discussions,and to Eva Ragnar for technicalassistance.
This study was supported by the Swedish Cancer Society, the SwedishMedical ResearchCouncil (project 5667),andPHSgrant5 RO1CA28380-03 from theNational CancerInstitute. H.L.-F. and J.M. are the recipients of visiting scientist fellowships from the Swedish CancerSociety. H.L.-F. is alsosupportedbytheNational NatureScience Foundation andNationalOncogeneResearch Cen-ter in China.
REFERENCES
1. Baichwall,V.R.,and B.Sugden. 1989. The multiple membrane-spanning segments of the BNLF-1 oncogene from Epstein-Barr virus are required for transformation. Oncogene4:67-74. 2. Becker, P. B., S. Ruppert, and G. Schultz. 1987. Genomic
footprinting reveals cell type-specific DNA binding of ubiqui-tous factors. Cell 51:435-443.
3. Bonnerot, C., M. Daeron, N. Varin, S. Amigorena, P. M. Hogarth,J. Even, and W. H. Fridman. 1988. Methylation in the 5' region of the murine beta FcgammaR gene regulates the expression of Fcgamma receptor. J. Immunol. 141:1026-1033. 4. Busson, P., 0. Ganem, P. Flores, F. Mugneret, B. Clausse, B.
Caillou, K. Braham, H. Wakasugi, M. Lipinski, and T. Tursz. 1988. Establishment and characterization of three transplant-able EBV-containing nasopharyngealcarcinomas. Int. J. Can-cer42:599-606.
5. Cao, S.-L. 1987. Studies of the biological behaviour and flow cytometric characteristics ofanasopharyngeal carcinomatumor
passaged in nude mice. Chin. J. Oncol. 9:6-9.
6. Cedar, H. 1988. Minireview: DNAmethylation. Cell 53:3-4. 7. Dawson, C. W., A. B. Rickinson, and L. S. Young. 1990.
Epstein-Barr virus latent membrane protein inhibits human epithelial cell differentiation. Nature (London) 344:777-780. 8. Dillner, J., B. Kallin, H.Alexander, I. Ernberg, M. Uno, Y. Ono,
G. Klein, and R. A. Lerner. 1986. An Epstein-Barr virus (EBV)-determined nuclear antigen (EBNA 5) partlyencodedby the transformation associated BamWYH region of EBV DNA: preferential expression inlymphoblastoidcell lines. Proc. Natl. Acad. Sci. USA 83:6641-6645.
9. Dillner, J., B. Kallin, B.Ehlin-Henriksson, L. Rymo, G. Henle, W. Henle, and G. Klein. 1986. The Epstein-Barr virus-deter-mined antigen is composed of at least three different antigens. Int. J. Cancer 37:195-200.
10. Doerfler, W. 1983. DNA methylation and gene activity. Annu. Rev. Biochem.52:93-124.
11. Dynan, W.S. 1989.Understanding the molecular mechanism by which methylation influences gene expression. Trends Genet.
5:35-36.
12. Ernberg, I., K. Falk, J. Minarovits, P. Busson, T. Tursz, M. G. Masucci, and G. Klein. 1989. The role of methylation in the phenotype-dependent modulation of Epstein-Barr virus nuclear antigen 2 and latent membrane protein genes in cells latently infected with Epstein-Barr virus. J. Gen. Virol. 70:2989-3002. 13. Ernberg, I., B. Kallin, J.Dillner, K. Falk, B. Ehlin-Henriksson,
and G. Klein. 1986. Lymphoblastoid cell lines and Burkitt lymphoma derived cell lines differ in the expression of a second Epstein-Barr virus encoded nuclear antigen. Int. J. Cancer 38:729-737.
13a.Fahraeus, R., A. Jansson, A. Ricksten, A. Sjoblom, and L. Rymo. 1990. Epstein-Barr virus encoded nuclear antigen 2 activatesthe viral latent promoter by modulating the activity of
on November 10, 2019 by guest
http://jvi.asm.org/
a negative regulatory element. Proc. Natl. Acad. Sci. USA 87:7390-7394.
14. Fahraeus, R., H. LiFu, I. Ernberg, J. Finke, M. Rowe, G. Klein, K. Falk, E. Nilsson, M. Yadav, P. Busson, T. Tursz, and B. Kallin. 1988. Expression of Epstein-Barr virus proteins in nasopharyngeal carcinoma. Int. J. Cancer 42:329-338. 15. Ffahraeus, R., L. Rymo, J. S. Rhim, and G. Klein. 1990.
Morphological transformation of human keratinocytes express-ing the LMP gene of Epstein-Barr virus. Nature (London) 345:447-449.
16. Feinberg, A. P., and B. Vogelstein. 1983. A technique for radiolabelling DNArestriction nuclease fragments to high spe-cific activity.Anal. Biochem. 132:6-13.
17. Ghosh, D., and E. Kieff. 1990. cis-Acting regulatory elements near the Epstein-Barr virus latent-infection membrane protein transcriptional start site. J. Virol. 64:1855-1858.
18. Hennessy, K., S.Fennewald,and E. Kieff. 1985. A thirdnuclear protein in lymphoblasts immortalized by Epstein-Barr virus. Proc.Natl. Acad. Sci. USA 82:5944-5948.
19. Hennessy, K., S.Fennewald, M. Hummel, T. Cole, and E. Kieff. 1984. A membrane protein encoded by Epstein-Barr virus in latent growth-transforming infection. Proc. Natl. Acad. Sci. USA 81:7207-7211.
20. Hennessy, K., and E. Kieff. 1985. A second nuclear protein is encoded by Epstein-Barr virus in latent infection. Science 227:1238-1240.
21. Hennessy, K., F. Wang, E. W. Bushman, and E. Kieff. 1986. Definitive identificationof a memberoftheEpstein-Barr virus nuclearprotein 3family. Proc. Natl. Acad. Sci. USA 83:5693-5697.
22. Hitt, M. M., M. J. Allday, T. Hara, L. Karran, M. D. Jones, P. Busson, T.Tursz, I. Ernberg, and B. E. Griffin. 1989. EBV gene expression inanNPC-relatedtumour. EMBOJ.8:2639-2651. 23. Holler, M., G. Westin, J. Jiricny, and W. Schaffner. Spl
transcriptionfactor binds DNA and activates transcription even when the binding site is CpG methylated. Genes Dev. 2:1127-1135.
24. Kallin, B., J. Dillner,I.Ernberg, B.Ehlin-Henriksson, A. Rosen,
W.Henle, G. Henle, and G. Klein. 1986. Fourvirallydetermined antigensareexpressedinEpstein-Barrvirustransformedcells. Proc. Natl.Acad. Sci. USA 86:1499-1503.
25. Kelley, D. E., B. A. Pollok, M. L. Atchison, and R. Perry. 1988. Thecoupling between enhancer activityandhypomethylation of
K immunoglobulin genes is developmentally regulated. Mol. Cell. Biol.8:930-937.
26. Keshet, I., J. Yisraeli, and H. Cedar. 1985. Effect ofregional DNAmethylation ongeneexpression. Proc. Natl. Acad. Sci. USA 82:2560-2564.
27. Klein, G., L. Dombos, andB. Gothoskar. 1972. Sensitivity of Epstein-Barr virus (EBV) producer and non-producer human lymphoblastoidcelllines. Int.J.Cancer10:44-57.
28. Kovesdi, I., R. Reichel, and J. R. Nevins. 1987. Role ofan
adenovirusE 2 promoterbinding factorinElA-mediated
coor-dinategenecontrol.Proc. Natl. Acad. Sci. USA 84:2180-2184. 29. Masucci, M. G., B. Contreras-Salazar, E. Ragnar, K. Falk,J. Minarovits, I. Ernberg, and G. Klein. 1989. 5-Azacytidine up regulatestheexpression ofEpstein-Barrvirus nuclearantigens 2 (EBNA-2)through EBNA-6and latentmembrane proteinin the Burkitt'slymphoma line Rael. J. Virol. 63:3135-3141.
30. Petti, L., J. Sample, F. Wang, and E. Kieff. 1988. A fifth Epstein-Barr virus nuclear protein (EBNA3C) is expressed in latently infected gorwth-transformed lymphocytes. J. Virol. 62:1330-1338.
31. Reisman, D., and B. Sugden. 1986. trans activation of an Epstein-Barr viral transcriptional enhancer by the Epstein-Barr viral nuclear antigen 1. Mol. Cell. Biol. 6:3838-3846.
32. Ricksten, A., B. Kallin, B. H. Alexander, J. Dillner, R. Fahraeus, G.Klein,R.Lerner, and L. Rymo.1988. The BamHI Eregionof theEpstein-Barr virus encodes three transformation associated nuclearproteins. Proc. Natl. Acad. Sci. USA 85:995-999. 33. Ricksten, A.,A.Olsson,T.Andersson, and L. Rymo. 1988. The
5'flanking region ofthe genefortheEpstein-Barr virus-encoded nuclearantigen2containsacell typespecificcis-acting regula-toryelementthatactivates transcriptionin transfected B-cells. Nucleic Acids Res. 16:8391-8409.
34. Rowe, M., H. S. Evans, L. W. Young, K. Hennessy, E. Kieff, and A. B. Rickinson. 1987. Monoclonal antibodies to the latent membraneprotein of Epstein-Barrvirus revealheterogeneityof theproteinand inducibleexpressionin virustransformed cells. J. Gen. Virol.68:1575-1586.
35. Rowe, M., and C. Gregory. 1989. Epstein-Barr virus and Bur-kitt'slymphoma. Adv. Viral Oncol. 8:237-259.
36. Rowe, M., D. T. Rowe, C. D. Gregory, L. S. Young, P. J. Farrell, H. Rupani, and A. B. Rickinson. 1987.Differencesin B cell growth phenotype reflect novel patterns of Epstein-Barr viruslatent gene expression in Burkitts lymphoma cells. EMBO J. 6:2743-2751.
37. Saluz, H. P., I. M. Peavers, J. Jiricny, and J. P. Jost. 1988. Genomicsequencingand in vivo footprinting ofan expression-specific DNase I-hypersensitive site of avian vitellogenin II promoter reveal a demethylation ofa mCpGand a change in specific interactions of proteins with DNA. Proc. Natl. Acad. Sci. USA85:6697-6700.
38. Sugden, B., andN. Warren. 1989. ApromoterofEpstein-Barr virus that canfunctionduringlatentinfection can be transacti-vated by EBNA-1, a viral protein required for viral DNA replication duringlatentinfection. J. Virol.63:2644-2649. 39. Tugwood, J., W.-H. Lau, S.-K.0,S.-Y.Tsao,W. M.C.Martin,
W. Shiu, C. Descranges, P. H. Jones, and J. R. Arrand. 1987. Epstein-Barr virus-specific transcription in normal and
malig-nantnasopharyngealbiopsiesand inlymphocytesfromhealthy donors and infectious mononucleosis patients. J. Gen. Virol. 68:1081-1091.
40. Wang,D.,D.Liebowitz, and E. Kieff. 1985. An EBV-membrane proteinexpressed inimmortalized lymphocytes transforms
es-tablished rodent cells.Cell 43:831-840.
41. Watt, F., and P. L.Molloy.1988.Cytosinemethylationprevents bindingtoDNAofaHeLacelltranscriptionfactorrequiredfor optimal expression of the adenovirus major late promoter. GenesDev. 2:1136-1143.
42. Woisetschlaeger, M., J.L. Strominger, andS.H. Speck. 1989. Mutually exclusiveuseof viral promoters inEpstein-Barrvirus latently infected lymphocytes. Proc. Natl. Acad. Sci. USA 86:6498-6502.
43. Young,L.S., C.W.Dawson,D.Clark,H.Rupani,P.Busson,T.
Tursz, A. Johnson, and A. B. Rickinson. 1988. Epstein-Barr virus gene expression in nasopharyngeal carcinoma. J. Gen. Virol.69:1051-1065.