0022-538X/89/104234-08$02.00/0
Copyright © 1989, AmericanSociety for Microbiology
Characterization of
Enhancer Elements and
Their Mutations in
the
Long Terminal Repeat of Feline
Endogenous
RD-114
Proviruses
ANANTA K. GHOSH' ANDPRADIP ROY-BURMANl12*
Departments ofPathology' andBiochemistry,2 University ofSouthern California SchoolofMedicine, LosAngeles, California 90033
Received 3 April1989/Accepted 16June 1989
Tolocate theenhancer regionsof the felineendogenousRD-114long terminalrepeat(LTR),we examined expressionof thechloramphenicol acetyltransferasegenedrivenbyvarioussegmentsof theU3regionfromtwo different proviral loci(CRL3 and CR1). Transientexpression assays demonstrated that the primarysignal
sequencefor transcription enhancement waslocated within the63-base-pair (bp)element of the CRL3DNA occurring betweenpositions -184 and -121 from the CAP site(+1), whereas the similarregionof CR1was
almost inactive. This element from both CRL3 and CR1 contained a single 30-bp sequence (direct repeat [DR]-B2) foundinduplicate tandemcopiesinthe LTR of the infectiousRD-114provirus. Two9-bpinverted repeatsmarked the DR-B unit ofthe activeelement,andaprominentbasedeletioninoneof theserepeats in CR1 DNAappearedtobe relatedtoloss of enhanceractivity. AnothersegmentofCRL3(-296to-184), also displaying enhancer function, contained tandem repeated sequences(DR-Al and DR-A2). The DR-A2unit, which lacked the 5' 20-bp sequenceof the 47-bp DR-Al, couldnot function as an enhancerby itself,but it contributed to enhancer effects in cooperation with either the DR-Al or DR-B2 region. The CR1 LTR contained asingle DR-Al sequencewith extensivemutations, and theregion (-313to-181) containing this DR-Al unit was nonfunctional, similar to the DR-B2 region ofCR1. Site-directed mutagenesis analysis of anotherenhancerelement,anoctamermotifoccurringbetween CAAT and TATA boxes of all RD-114LTRs
sequenced, revealed thatthis element was necessary for full enhancer function of the U3region but witha
variable effect, depending on the cell types in which chloramphenicol acetyltransferase expression was
determined.
There are 15 to 20 copies of stably inherited RD-114
retroviralDNA perhaploidgenomeofthedomesticcat(1, 3, 27, 28, 32, 38). Apparently, most are defective in one or moreviralgenes,andit isprobablethat there isonly asingle
locus with the capacity to code for a viable xenotropic
RD-114retrovirus(27, 38).However, severalofthepartially deleted RD-114 proviruses in the cat genome are strongly expressed incertainfelinetissuesand cell lines (5, 23-25, 30,
37). Previously, we showedthatthe 5' long terminal repeat
(LTR) ofan endogenous RD-114 locus (CRL3), partly de-leted inthe envgene, retained strong activity in promoting thetranscriptionof a linked gene,althoughthis activity was
approximately threefold lower than that of the infectious RD-114provirus (37). In contrast, the 5' LTR from another
similarly deleted proviral locus (CR1) lacked this ability to support transcription. DNA sequencing of these LTRs re-vealedinteresting differences in the U3 region, especially in the two setsof tandemdirect repeats (DRs) found in the LTR
of the infectious RD-114 provirus (37). None of the se-quences within any of the RD-114 DRs or anywhere in U3 contained regions homologous to the core enhancers of similar virus 40 (SV40), mouse leukemia virus, or mouse mammary tumor virus (6, 7, 14, 18, 19, 41). In addition to these novel DRs or their mutated forms, all RD-114 proviral LTRs sequenced showed the presence of a conserved
oc-tameric sequence identical to the sequence functionally
implicatedin transcription regulation of several mammalian genes (9, 22, 26, 29, 35, 37).
In this report, we extend our endogenous RD-114 LTR
studiestocharacterizetheenhancer elements present in the
*Corresponding author.
U3region. A series of constructs linking various regions of the active (CRL3) or inactive (CR1) LTR to the bacterial chloramphenicol acetyltransferase (cat) gene has been gen-erated. Study of these constructs derived from these two functionally opposite but structurally closely related variants has aided in the identification of important domains by
correlating sequence changes with differences in activity.
Here, we demonstrate the presence of transcriptional en-hancer in domains of the U3 region of the CRL3 LTR and show that the similar domains of the CR1 LTR have been inactivated by naturally occurring mutations. We also pro-videevidence that the octamer motif,occurring between the CAAT and TATA boxes of this provirus, functions like its mammalian gene counterpart in enhancingtranscription ina
cell-type-dependent manner.
MATERIALS ANDMETHODS
Cell lines. The mouse fibroblast cell line NIH 3T3 and felineembryo fibroblast cell line H927 (30) were maintained in high-glucose Dulbecco modified Eagle medium supple-mented with 10% fetal bovine serum. The human B-lym-phoid tumor cell line Raji was cultured in RPMI 1640 mediumsupplemented with 10% fetal bovine serum.
Plasmid constructions. Plasmid pSVIXcat (12), provided by D. Celander, is identical to pSVIcat except that it contains anXhoI linker at the siteofrecircularization. This plasmid has the SV40 promoter, but not enhancer, se-quencesupstreamof thecatgene(Fig. 1B). The endogenous RD-114 LTR putative enhancer regions were inserted up-stream of the promoter sequences in pSVIXcat. Four 5' LTRs, clones CRL3, CR1, CR5, and CRL17 (38), were used. The 309-base-pair (bp) Sau3AI-HindIII fragment of the
4234
on November 10, 2019 by guest
http://jvi.asm.org/
A CRL 3
-3410 -430 -121 .1 *290
3U R u5
f/ I Il
E Sa H Sm E
-278 -121
Aval
j>2bpZp- 2 bp_ 2
K DR-Al1 DR-A2
CR1
-1810 -313 -120o 1 .289
13 R U5
I
,-- ---l
E P H Sm E
-249 -120
Aval
49bp2Sb
DR-A l DR-B 2
B
CAT activity relative
topSV1XCAT
J pCRL-3fSX-.-4
H
-' pCRL-3/SVIX 11.2 4.9
P H
L ---- I pCR-1,SYVX
P H
P H
1.2
pCR-5, SV1X 0.9 pCRL-17 SVIX 1.0
1.2 0.7 1.0
CRL3 5' LTR, containing DR units of
as a portion of 5' flanking cellular DNA sequences, was inserted into the XhoI site of pSVIXcat (Fig. 1A and B). Similarly, the 193-bp PstI-HindIII fragment of each of the other three 5' LTRs (CR1, CR5, and CRL17) was cloned into that plasmid. The plasmid derivatives, named pCRL-3/SVIX, pCR-1/SVIX, pCR-5/SVIX, and pCRL-17/SVIX, were isolated in both orientations of the insert, the correct orientation being designated ->and the inverted orientation
being designated
*-(Fig.
1B).Plasmids pL3 and pCR1 (Fig. 2A) were constructed by inserting the 411-bp HindIII-EcoRI fragment of CRL3 or the corresponding 409 bp of the CR1 LTR into the HindlIl site of
pSVOcat(12). These constructs, lacking DR units but con-taining the promoter region (CAAT, octamer, and TATA boxes), were used as recipient plasmids for introduction of various enhancer sequence domains. The correctly oriented pL3 and pCR1 plasmids were cut at the HindlIl site for insertion of either the 309-bp (-121 to -430 of CRL3) or 193-bp (-120 to -313 of CR1) fragment, using HindlIl linkers. The resulting plasmids, containing homologous and mix-and-match sequences between CRL3 and CR1 LTRs, weretermedpCRL-3/L3, pCR-1/L3, and pCRL-3/CR1 (Fig. 2A).
To generate DR deletion mutants, pCRL-3/L3 was first linearized by NdeI digestion, and the 5' upstream DNA was
trimmedback by BAL 31 exonuclease. HindlIl linkers were then added to the deletion ends and excised by Hindlll
digestion,and theisolated 5' deletion DNA fragments were inserted into theHindlIl siteof pL3. Two deletion mutants were isolated (Fig. 3A): one lacking most of the sequences upstreamofthe DRregion(pL-3/L3)and the otherwith most of the DR-Al sequence eliminated [pdL-3(-221)/L3]. For making otherdeletion mutants, pL-3/L3 was digested with both AvaIl andHindlll, and two ensuing fragments (-296 to -184 and -184 to -121) were modified by Hindlll linkers
and ligated into the HindIII site of pL3. This process
generatedplasmidspdL-3(-184)/L3andpdL-3(-296)ID/L3. Similarly, pdCR-1(- 181)CR1 and pdCR-1(-313)ID/CR1
were obtained frompCR-1/CR1 (Fig. 3A).
Two additional DR deletion mutants, pdCR-1(-181)/L3
and pL-3(-221)ID/L3, were also obtained. The Hindlll
L0.1kb
C
3-Cm-1-Cm- *
Cm-
-0
a
E I I I
b c d e f g h i
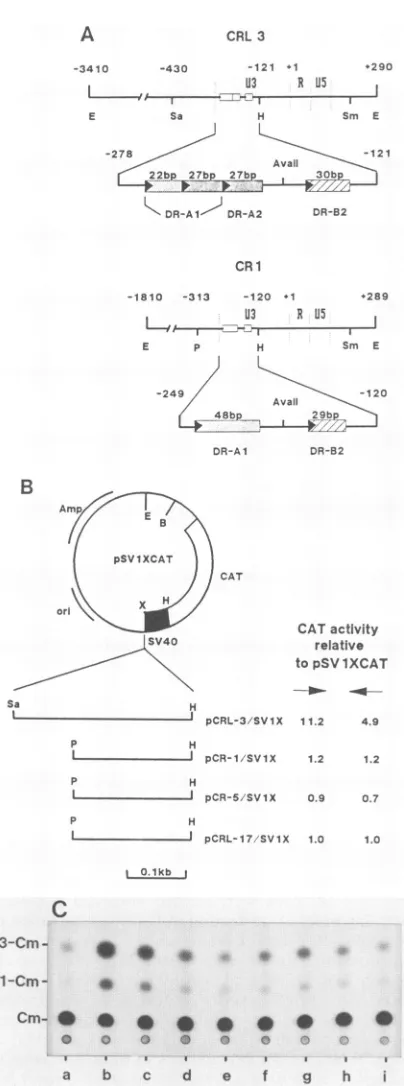
FIG. 1. Differential activity of the enhancerregion from the 5' LTRsoftwoendogenousRD-114 proviruses,CRL3 and CR1. (A)
Schematic offeatures, including U3, R,andU5 regionsand certain
restriction sites withinoradjacenttoeach LTR. The bottom line for
each LTRstructurerepresentsanexpandedversion of the 5' half of
U3, with DR units shown as boxes. The LTR of the infectious
RD-114 provirus contains two sets of tandem DRs (37). The first
47-bpDR(DR-Al)isfollowedbyasecondcopy,DR-A2.Similarly,
a30-bpDR(DR-B1)isrepeatedas DR-B2. InCRL3,the5'portion
of DR-A2 is missing, but the 3' 27-bp region isretained. CR1 has
onlyone DR-Al-related region, and both CRL3 and CR1 lackthe
firstcopyof the 30-bpDR. The DR boxesareshadeddifferentlyto
emphasizesequencenonidentitybetweenDR-A and DR-B domains,
and the 27-bprepeatsin CRL3 areindicatedbydense stippling.A
solidarrowrepresentsthestartofeachrepeatunit;the actuallength
of the unit(inbasepairs)isindicated above the box.(B)
Construc-tion ofcatplasmids, showingthepSVIXcat circle containingSV40 earlypromoters(U)andsequencesencodingthecat gene(L-).The plasmidsequences of the circle, although notshown, alsocontain
SV40intronandpoly(A)additionsitesimmediatelydownstreamof
thecatgene.Thediagramsbelow the circledepictLTRsegments of
differentRD-114proviruses (CRL3, CR1, CR5, andCR17) andthe
relevant restriction endonuclease cleavage sites. Abbreviations for
restriction sites:B, BamHI; E, EcoRl; H, HindlIl; Sa,Sau3A1; Sm,
SmaI; P, Pstl; X,XhoI. Results of CATassaysafter transfectionof
theseplasmidswith correct(-*) orinverted(<-) orientation ofthe
insert into NIH 3T3 cells are shown at the right. CATactivity is
expressed as the level of conversion of chloramphenicol to its acetylatedforms relativetothatobserved withpSVIXcat.Each value
represents theaverageof threetransfections;the variationbetween
experiments was less than 30% of the average. (C) Representative autoradiogramof the CATassay.Lanescontainedtransfected DNA
from:a,pSVIXcat; b,pCRL-3/SVIX (->); c,pCRL-3/SVIX();d,
pCR-1/SVIX (--); e, pCR-1/SVIX (<-); f, pCRL-17/SVIX (>;g,
pCRL-17/SVIX(<-); h, pCR-5/SVIX (-*); i, pCR-5/SVIX (<-).
Posi-tions ofmigrationofchloramphenicol (Cm)and itsacetylatedforms
(1-Cmand3-Cm)areindicated hereandinFig.2B to 4B.
Sa
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.76.278.69.612.2]A
Construct
121
L
pL3
pCRL-3/L3 CRL-3
-430 DR-Al DR-A2 DR-BZ
-313 CR-1
D1-s-DRB2
CRL3
Relative CATactivity
-E E
.290
CAAT octamerTATA I
J/ CAT 1.0
--_
_---CRL 3
CAT 7.5 2.9
CRL 3
CAT 1.3 0.9
-120 CR1 *289
I---- -_-- CAT 0.5
CRL-3
-DR--Al
-I *; I.-_ X
CR1
CAT 4.6 1.e
B
3-Cm1
1-cm-i
*1*
Cm- 99tt t
, , , ,
a b c d e f h
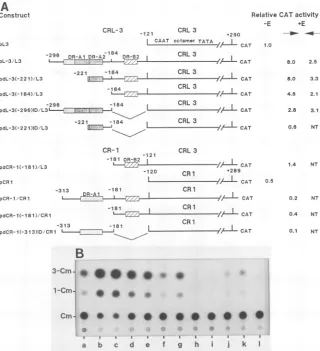
FIG. 2. Effect ofenhancer regiononhomologouspromoters.(A)Linedrawingsofcatplasmidscontaining onlythepromoterregion from theCRL3orCR1 LTR (pL3orpCR1)orbothpromoterandupstreamenhancerregions(pCR3/L3). Of thetwo mix-and-matchconstructs, pCR-1/L3 contains promoters from CRL3 and upstream sequences from CR1, and pCRL-3/CR1 contains promoters from CR1 but the enhancerregion from CRL3. The boxed regions mark DRunits, and the relativepositionsof theCAAT,octamer, and TATA elementsare indicated withinthepromoterregion. CATactivity dataareaveragevalues obtained from three transfectionexperimentsin NIH 3T3cells when the orientation of the enhancer (+E) region wasinnatural(-*)orinverted orientation(<-)with respecttothe promoterregion. -E, Valuesobtained without the enhancer region. All valuesareexpressed relativetotheactivityofpL3 (normalizedto1.0). (B)Autoradiogram fromatypical CATassay. Lanes containedplasmid DNAs from:a,pSVOcat; b, pL3;c,pCRL-3/L3(-*); d, pCRL-3/L3 (<-);e,pCR-1/L3 (--); f, pCR-1/L3 (<-);g,pCR1;h, pCRL-3/CR1 (->);i,pCRL-3/CR1(<-).
fragment (-181to -120), isolated frompdCR-1(-181)/CR1, when cloned into the HindIII site of pL3 provided pdCR-1(-181)/L3. Togenerate pL-3(-221)ID/L3, theNdeI-AvaIl fragment ofpdL-3(-221)/L3wasblunt ended intoNdeI- and
HindlIl-cleaved pL3 (Fig. 3A). Deletion endpoints of all constructs and theirorientations were determined by
dide-oxy sequencing aftercloning oftheadjoining region intoan
M13 vector(31).
Mutagenesis of the octamer sequence ATGCAAAT was
donebythemethod of Zoller andSmith(43) asmodifiedby
Geisselsoder et al. (11). Briefly, two oligonucleotides,
oc-tamerA (ATTCAACT; OA) andoctamer B (ATTAAAAT; OB), were synthesized and purified by high-performance
liquid chromatography. The HindIII-SmaI (-121 to +200) fragmentofCRL3, containing theoctamermotif,wascloned
into the same restriction-digested M13mpl8 vector and transformed into Escherichia coli CJ236 (dut ung strain) to
yield uracil-containing single-stranded bacteriophage DNA (16). The phage DNA was then annealed with the
kinase-treated mutant oligonucleotide, the complementary strand
was extended by T4 DNA polymerase, and the ends were
joined by T4 ligase. The heteroduplex DNA was used to
transform MV1190,anE. coli strain containingactiveuracil
N-glycosylase (16). Thepresenceof thisenzymereduced the
biological stability ofthe parental DNA strand and
conse-quently led to accumulation ofthemutant phage. Afterthe mutations in the progeny phage were confirmed by DNA
sequencing, the HindIII-SmaI fragment was excised from
each mutant phage DNA and cloned into
HindIII-SmaI-cleaved pL3 orpL-3/L3 togenerate pL3(GA) andpL3(GB)
orpL-3/L3(GA) and pL-3/L3(GB), respectively (Fig. 4A).
Allplasmidswere purifiedbytwoCsClgradient centrifu-gationsbefore use intransfection analyses.
Transfections and CATassays.FelineH927ormouseNIH
3T3 fibroblasts were transfected by a modification ofthe
calcium phosphate method of Grahamand Van der Eb(13). Briefly, 5 x
105
cells were seeded onto 100-mm-diameter dishes 1 daybefore transfection in Dulbecco modifiedEagle medium plus 10% fetal bovine serum. Calcium phosphateDNAprecipitateswereprepared from 20 p.gofplasmid DNA
alongwith30,ugof shearedcalfthymuscarrier DNAin 1ml of HEPES (N-2-hydroxyethylpiperazine-N'-2'-ethanesulfo-nicacid)-bufferedsalineby constantlybubbling nitrogengas
through the solution while adding calcium chloride. After allowing20minfor theprecipitates toform, the suspension
was added to each dish and incubatedat 37°C for 4 h. The cells were thenglycerol shocked (15%glycerol in
HEPES-buffered saline) for 4 min, washed withfreshmedium, and incubatedfor 48 h at 37°C before harvesting (4).
The human Raji cells were transfected with the plasmid
DNAbyamodifiedDEAE-dextran protocol(2).Briefly,2x 107 cells were washed twice with serum-free medium and oncewithTBS(25mMTrishydrochloride [pH 7.4], 137 mM
NaCl,5 mMKCI,0.6 mMNa2HPG4, 0.7mMCaCl2, 0.5mM MgCl2) by gentle centrifugation.Thecellswereincubatedat
room temperature in 2 ml of TBS containing 500 jig of DEAE-dextran (Pharmacia, Inc.)per mland 20 ,ug of plas-mid DNAfor 45min.Theunabsorbed DNAwasremoved by
pCR I
pCRL-3/CRI
-430
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.153.476.87.347.2]A
Construct
CRL-3
pL3
1Z
-296 DR-Al DR-A2 DR-B21
pL-3.L3 Z ii
-221 -184
pdL-3(-221)2 =/L3
-184 _ pdL-3(-184).L3
-2 96 -184
pdL-3(-296)IDI/L31 :--- D
-22 1 -1 84 pdL-3(-221ID,/L3 -2 8 L
! C 3 290
CAAT octamer TATA
CR3' CAT
CRL 3
CRL 3
CRL 3
-L CAT
1 CAT
CRL 3 /J C CRL-3-- If CAT
CRL3 e
--- ~~if CAT
RelativeCATactivity
-E +
1.0
8.0 2.5
8.0 3.3
4.6 2.1
2.8 3.1
0.6 NT
CR-1
_ _-121
-1
-313 &A
CRL 3
8l OR-B21
810 '- -- /A-'- CAT
-120 CR1 +289
I_IF_I_f- CAT
181 CR1
._, t
---7d1 71/{4 CAT
CR1
pdCR-1(-181),'CRI -//-< CAT
-313 -181 CR11
pdCR-1(-313)1D'CR1 - _ 1 II/ CAT
B
1.4 NT 0.5
0.2 NT 0.4 NT 0.1 NT
3-Cm-. *
1-C * * *
Cm!
l l I l l l l l
a b c d e f g h i j k
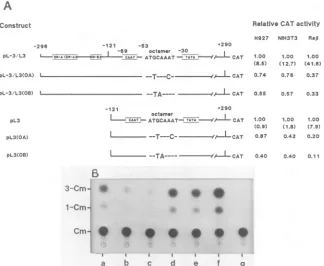
FIG. 3. Structures and activities of mutants bearing 5' unidirectional deletions ordeletions in the central region. (A) Line drawings numbered with deletion endpoints. The sourceof the enhancerregion is indicated above each line by CRL-3orCR-1. Similarly, the origin
ofthepromoterregion is marked by CRL3orCR1. DR domains, promoterelements, and othersymbolsare asforFig. 2. ID inconstruct
designations indicates internal deletion. CAT activity data are the averages ofat least three but in most cases upto eight independent transfection experiments. NT, Notransfection done. (B)Representative CATassay autoradiogram. Lanescontained plasmid DNAs from:
a,pL3; b,pL-3/L3 (-);c,pdL-3(-221)/L3(->); d,pdL-3 (-184)/L3(-);e,pdL-3 (-296)ID/L3(p); f,pdL-3 (-221)ID/L3();g,pdCR-1 (-181)/L3( +*); h,pCR-1/CR1();i, pdCR-1(-313)ID/CR1(--);j,pdCR-1(-181)/CR1 (-->); k,pCR1. Lanel, Untransfected celllysate.
two washes with TBS and one wash with serum-free
me-dium, and the cells were replated in 20 ml of
serum-containing medium. After 48 h of incubation at 37°C, the
cells were harvested for theCATassay.
Transfected cells were washed three times with cold
phosphate-bufferedsalineandlysedin100
RI
of250 mM Tris hydrochloride (pH 7.8) by three cycles of freezing andthawing. The cellular debriswaspelleted by centrifugation,
andportionsof thesupernatantfluidequivalentto150,ug of protein were assayed for CAT activity as described by
Gorman et al. (12). The percent acetylation of
[14C]chlor-amphenicolwasdeterminedbyseparatingtheacetylatedand unacetylated forms by thin-layer chromatography, followed
by liquid scintillation counting of spots scraped from the
silicagel plates. The positions of the spotswere located by
autoradiography.
RESULTS
Activityof theenhancerregions fromendogenous proviral
DNAs. To characterize the enhancer elements, we
intro-ducedthe LTRandadjoining cellular DNA region upstream
of the CAAT box and containing the DR units into the
parental test plasmid pSVIXcat (Fig. 1A and B). Plasmid
pCRL-3/SVIX comprised the 309-bp HindIII-Sau3Al (-121
to -430) fragment of the CRL3 clone placed 5' to SV40
promotersequences. Similarconstructions were made with
the analogous 193-bp HindIII-PstI fragment occurring in
eachoftheCR1,CR5,andCRL17clones.Theseconstructs,
containing the insert in either correct or opposite
orienta-tion, were transfected into NIH 3T3 mouse fibroblasts and
assayed for transient CAT expression. Average CAT
activ-ities relative to that ofpSVIXcataregiven in Fig. 1B, anda
representative autoradiogram shown in Fig. 1C.The309-bp
region of the CRL3 DNA induced a severalfold increase in
cat geneexpression.In thecorrectorientation,the
enhance-ment was 11-fold; the opposite orientation of the insert
resulted inanapproximately5-fold increase. Incontrast,the
corresponding fragments ofCR1, CR5, and CRL17 cloned
DNAs were inactive in inducing cat gene expression over
the background level ofpSVIXcat. Therefore, it appeared
that theinactivity of the CR1 fragmentwas notan isolated
case, asthere were otherloci that were similarlyinactive. pdCR-1(-181)'L3
pCR 1
pCR-1. CR1
CRL3
-.1.
-1I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.142.462.79.430.2]A
Construct
-296 -121 -53 *290
X -69 octamer -30 CAT
pL-3. L3 L.D-.0R-APA-A AT -ATGCAAAT CAT
pL-3/IL3(0A) t -_- -T--C----1 CAT
--TA--- - CAT
'290
I octamer -eAA
1 CAAT ATOCAAAT-'._ --{ I-CAT
l --T---C- if CAT
I --TA-- i-f, I CATr
Relative CATactivity
H927 NIH3T3 Ral
1.00 1.00 1.00
(8.5) (12.7) (41.6) 0.74 0.76 0.37 0.55 0.57 0.33
1.00 1.00 1.00
(0.9) (M1.) (7.9)
0.87 0.42 0.20 0.40 0.40 0.11
B
3-Cm-1-Cm.
Cmj
[image:5.612.156.480.77.343.2]a b c d e f g
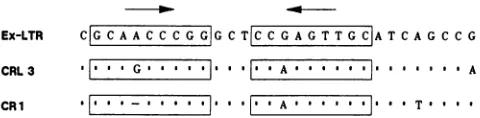
FIG. 4. Structures and activities ofmutantsbearingaltered octamericsequences.(A)Theoctamer sequence(ATGCAAAT)wasreplaced
byeitherOA(ATTCAACT)orOB(ATTAAAAT),andplasmids pL-3/L3 andpL3 containingthese mutationswereisolated. Thewild-type
andmutantplasmidsweretransfected into feline H927fibroblasts,mouseNIH3T3fibroblasts,orhumanRaji B-lymphomacells forcatgene
expression analyses. CATactivityvaluesareaveragesof the data obtained from three transfectionexperiments.Actualpercent conversion ofchloramphenicoltoitsacetylatedforms isgiveninparentheses.(B)AutoradiogramfromatypicalCATassaywithRajicell extracts.Lanes contained transfected DNAs from:a,pL3; b, pL3(GA);c,pL3(OB);d,pL-3/L3(GA);e,pL-3/L3(OB);f, pL-3/L3.Laneg,Untransfectedcell lysate.
These experiments with heterologous promoters were
then extendedto studythe effect of the enhancerregionon
homologous promoters. Since the nucleotide sequences of the CRL3 and CR1 LTRs were known (37), all subsequent
workwasdevotedto defining the enhancer elements within the309-bp regionof CRL3 andcomparingthem withthose in the similarregionof thefunctionallyinactiveCR1 LTR. The vectors containing the homologouspromoters werecreated
by inserting the 411-bp HindIII-EcoRI fragment (-121 to +290) of CRL3orthecorresponding 409 bp (-120to +289) of CR1 5' ofthe cat gene in pSVOcat (12), which did not contain any promoteror enhancer elements. The resulting
plasmids, pL3 and pCR1, were used to construct mix-and-match sequences between individual promoterand
en-hancerregions(Fig. 2A). Transfection and CATassayswith theseconstructsrevealedthat CRL3 andCR1 DNAs differed
in both promoter and enhancer activities. The promoter
ability ofpCR1 was approximately 50% of that observed
with pL3 (Fig. 2). This reproducible drop in activity could notbeattributedtothesequencesof the functional domains
such as CAAT, octamer, and TATA boxes, since CR1
sequences in these domains were identical to either the CRL3 LTRorthe LTR of the infectiousRD-114virus (37).
In fact, the entire 409-bp (-120 to +289) segment ofCR1
was highly homologoustothe corresponding portion of the other two DNAs, differing in only 22 scattered nucleotide changes uniquetoCR1. However, it should be noted that the 119-bp regionimmediatelyupstream of the CAP site(+1)of theCR1 LTRhadfive unique changes, includingadeletion
40bp upstreamof the CAAT box (37).
Thedegreeof inactivation of the enhancer function ofCR1 seemed much higher than the loss of its promoter ability.
The CR1 region in plasmid pCR-1/L3 did not significantly alter expressiondriven by the CRL3promoters,whereas the CRL3 enhancer region stimulated expressionfrom the CR1 promoters by approximately ninefold in the sense
orienta-tion and threefold in the antisense orientaorienta-tion (Fig. 2). Considering the relative strengths of the CR1 and CRL3 promoters, this level of enhancement was similar to that observed with plasmid pCRL-3/L3, the values obtained beingsevenfoldandthreefold, respectively. Together, these results indicated that the inability of the CR1 LTR to promotetranscriptionwas primarilycaused byits enhancer weakness. The results also revealed that the active CRL3 enhancers were partially dependenton orientation, the
an-tisenseorientationbeing threefold lowerinactivity than the native order.
Borders of the enhancer region. The DR domains fre-quently occurring in the U3 region of the retroviral LTR have been implicated as transcriptional enhancer elements
(6, 8, 15, 17, 20, 21). Since the CRL3 LTR containedtwo copies of partly homologous DR sequences (DR-Al and
DR-A2)andasinglecopyof DR-Bthat isfoundinduplicate copiesin theinfectious RD-114virus LTR(37),wewishedto
assessthe relativestrengthof the DRunitsandexamine their
cooperativity. Several deletion mutants were constructed.
The first productisolated by 5' unidirectional deletion was
pL-3/L3, which lacked most of the flanking cellular DNA
sequence. This plasmid expressed the cat gene with effi-ciencysimilartothat of thepreviousconstruct,pCRL-3/L3,
pL-3/L3(OB) L
pL3
pL3(OA)
pL3(0B) F
on November 10, 2019 by guest
http://jvi.asm.org/
Further 5' deletion of the insert to -221 removed DR-Al
sequences
completely
exceptfor the last3 nucleotides. Thismutant,
pdL-3
(-221)/L3, also expressedthecatgene at the samelevelseenwiththepL-3/L3
control, indicatingthattheDR-Al-containing
region
added very little to the totalen-hancer
activity.
Additional deletion resulted in plasmidpdL-3 (-184)/L3,
with anendpointat -184. However, thismutant,
lacking
both DR-Al and DR-A2 regions, showed areduction in
activity
of 42% of the control level. Thus, it seemed that themajority
of the enhancereffectwas associ-ated with the sequences(-184
to -121) that included the30-bp
DR-B2 domain. Thispoint
was strengthened by theobserved
significant
drop
inactivity
when the DR-B2region(-184
to -121) wasinternally
deleted. PlasmidpdL-3(-296)ID/L3
showedonly
35%
of the control level ofactivity.
Additional 5' deletion of thisplasmid,
lacking the DR-B2region,
resulted inelimination ofmostof the DR-Alsequences and consequent loss of all enhancer
activity
inplasmid
pdL-3(-221)ID/L3.
Takentogether, theresultsim-plied
that themajor
enhancer domain residedin the -184 to -121 DR-B2region
and that theDR-Al and DR-A2regions had acooperative
enhancereffect,
although the DR-A2region
by
itself could not function as an enhancer in thetransient
expression
assay system.Totest whether the
DR-B2-containing
region of the CR1LTRcould stimulate
transcription
when separated fromthe restofthe upstream sequences,we linkedthe -181 to -120region
of CR1 to the CRL3 promoters, as indicated in thepdCR-1(-181)/L3
construct(Fig.
3A). This CR1 region showedonly
aslight
increase(40%)
overthat of thepromot-er-alone construct, whereas the same
region
from CRL3displayed
an almostfivefold enhancement ofactivity.
Dele-tion
plasmids
were also made with CR1 DRregions
linked upstream of thehomologous
CR1 promoters. Three such constructs(bottom
ofFig.
3A),
containing
either theDR-Alor DR-B2
region
orboth,
did not exhibit anypositive
enhancereffect.
Rather,
itappeared
that the DR-Alregion
of CR1
might
have anegative
action ontranscription
pro-motion. For
example,
whereaspdCR-1(-181)/CR1
(retain-ing
only
the DR-B2region)
had nodetectable effect on cat geneexpression by
CR1 promoters,pCR-1/CR1 (containing
both DR-Al and DR-B2
regions)
andpdCR-l(-313)ID/CRl
(containing
only
theDR-Alregion)
showed reducedactivity
(40
and20%, respectively)
compared
withthe valuefortheCR1promoteralone
(Fig.
3A).
Thiseffectmight
berelatedtoextensive mutations in the CR1
47-bp
DRdomain relativeto theCRL3 orexogenous LTR element.Analysis
of octamer deletion mutants. The octamer en-hancerATGCAAAT,
conservedasisorinits invertedformin several mammaliangenes, alsooccursbetween theCAAT and TATA boxes of RD-114 andbaboon
endogenous
retro-virus but not in any other known retroviruses
(37).
To examine the function of this viral octamer, a set of con-structs was made in which the octameric sequence was alteredby oligonucleotide-directed
mutagenesis.
The mu-tants,OAandOB,
eachcontaining
twonucleotidechanges,
had the sequences ATTCAACT and
ATTAAAAT,
respec-tively
(Fig. 4A).
The mutated octamers inplasmid
pL3
orpL-3/L3
were testedfor catgeneexpression
inmouseNIH 3T3 and feline H927 fibroblasts and in humanRaji
B-lymphoma
cells. cat geneexpression
was reducedby
the octamer mutation toavariabledegree,
depending
onthe cellline used. Octamer mutants in the
pL-3/L3
frame reduced thecontrolactivity
to55to75%in thefibroblastic cells(Fig.
4A).
In thepL3
plasmids,
themutations lowered the levelofOA) caused significant reduction of cat gene expression in H927 cells. Of the three cell lines tested, Raji cells showed the most pronounced effect of the octamer mutations. In this cellline,pL3(OA) and pL3(OB) displayed only 20 and 11%, respectively, of the control level of activity (Fig. 4). Simi-larly, a reduced activity of 30 to 40% was seen with pL-3/L3(OA) and pL-3/L3(OB). These results implied that the viral octamer sequence, like the mammalian cellular gene counterpart, was necessary for efficient transcriptional
ac-tivityandthat thelevel of activity was dependent on the type
of cell in whichexpressionwas measured.
DISCUSSION
Wehave analyzed the enhancer function of the U3 region of the LTR of endogenous RD-114 proviruses by using acat gene expression system driven by either heterologous or
homologouspromoters. This analysishasyielded new
infor-mationonthe location of the enhancer comprising multiple functional elements. Although the boundary of each element has notbeen rigorously determined, the approximate
loca-tion ofeach isdeduced from analysis ofthe various deletion mutants. The element with the most pronounced activity that can function independently is located within a 63-bp sequence occurring between nucleotide positions -184 and -121. This region alone of the proviral clone CRL3 can stimulate the level of cat gene expression driven by its homologous promoters by fivefold. The similar region of anotherclone, CR1, located between -181 and -120, could notinduceexpressionabove the levelobtained only with its promoters.The extentof inactivationof this CR1 enhancer element is apparently quite striking, especially in the pres-enceof extensive homology betweenCRL3 andCR1 in the
respective 63- and 61-bp stretches ofsequence (37). There are only six base substitutions and two base deletions scattered in this region. The element, however, contains a copy of the 30-bp DRunit (DR-B2) (37). Ofthe eightbase
mutations,twodistinguish the CR1DR-B2domain fromthat of CRL3: one deletion 5 bp 3' of its 5' border and one transversion mutation (A-* T) 5
bp
5' of its 3' border. The base deletion is positioned near the middle of a 9-bp se-quence,which occurs againin invertedform, with3nucle-otides between copies (Fig. 5). In the LTR of infectious RD-114(Ex-LTRin Fig. 5), the inverted repeats within the DR-B2 domain are perfectly matched except for a single base mutation. These repeats have three base mutations in
CRL3,but thedeletion in CR1
disrupts
the sequence of thefirst domain, whereas its inverted repeat retains the same sequenceasin CRL3. Sincecrucialsequencesfor retroviral
transcriptionalenhancersresidelargelyinthe DRs of the U3
region (42),it isprobablethatDR-B domainhasasignificant
role in the observed function. The palindromic sequence present within DR-B may be essential for the
binding
ofatransactivatingdimericprotein factor,andwe
speculate
thatpartial
disruption
of thisdyad
symmetry in this sequence ofCR1 maybe a
major
determinant of its considerable loss of enhanceractivity.
Two other segments of the CRL3 LTR that display en-hancer activity also contain DR sequences. The
DR-Al-containing
region,
located betweenpositions
-296 and-221, has all of the
47-bp
DR-Al sequence except for the last 3 nucleotides. Whereas the DR-Al motif istandemly
repeated as DR-A2 in the LTR of the infectious RD-114
DNA,the 5'
portion
of DR-A2 is deleted inCRL3,
resultinginan
imperfect
DR-A2of 27 nucleotides. The DNAsegmenton November 10, 2019 by guest
http://jvi.asm.org/
Ex-LTR CGCAACCCGGGCTCCGAGTTGCATCAGCCG CRL 3 ' G . . . . A A CR1 . . . .' A . . . T
FIG. 5. Comparisonof the DR-B2 nucleotide sequence present
in all three RD-114 LTRs. The sequence of the infectious or
exogenous proviral LTR (Ex-LTR) as well as the sequences of
CRL3 and CR1 LTRs were reported previously (37). The boxed
areasrepresentthe9-bpinvertedrepeats; arrowsindicate orienta-tion.Symbols:0,sequenceidentitybetween theDNAs;-,deletion
found in CR1.
(-296 to -184) containing both DR-Al and DR-A2
se-quences ofCRL3, when separated from the DR-B2 region (-184 to -121) and linked upstream of the cat gene with CRL3 promoters, increases cat gene expression by
three-fold. Thus,the -296 to -184regionhassignificantenhancer
activity. Further deletion of this construct to remove the
-296 to -221segment, however, results in almostcomplete
loss of enhancer function, indicating that the
DR-A2-con-taining
region (-221 to -184) has little or no enhanceractivity byitself.However,when it isplaced upstreamof the
DR-B2-containing element, asinconstructpdL-3(-221)/L3,
it increases the DR-B2 enhancer effect by almost another
twofold. It is also noteworthy that deletion of the DR-Al
region (-296 to -221) from the entire enhancer does not
reduce enhancerfunction.
Takentogether,these resultsimplythat the DR-A2region
contributes to enhancer effect when it pairswith either the
DR-Al orDR-B2region but thatas anindividual element it
lacks enhanceractivity. It alsoappearsthat the presenceof
both DR-Aland DR-A2regionsmaynot berequiredforfull
function of theentireenhancer.However,this observation is
limited to theassaysystem used,and it remainspossiblethat
DR-Aland DR-A2togetherarecritical forexpressionofthe
endogenous proviral sequences in specific tissues of the
domestic cat. Since the DR-A2 domain lacks the first 20bp
of the DR-Al sequence,the 3' halfof thisrepeatedsequence may be functionally moreimportant than the 5' half. How-ever, the issue is also complicated by the demonstrated cooperativity between the DR-Al and DR-A2 regions of CRL3, and it is unclear whether the 5' region present in DR-Al butnot in DR-A2playsaroleintheenhancementof activity by DR-Al and DR-A2 together. Incontrast to the
enhancerability of the DR-Aregionof theCRL3clone, the
DR-Al region ofCR1appearsto be completelyinactivated
bythenaturalmutations.Themutations in this DRinvolvea
6-bp insertion near the 5' end and a 5-bp deletion of the
3'-terminal sequence(37).
The octamer sequence conserved in all RD-114 LTRs
sequenced constitutes another enhancer element. Distinct
elementscontainingthisoctamer motif havebeenimplicated
in both ubiquitous and tissue-specific enhancerfunction of
mammalian genes (10, 33, 34, 36, 39, 40). The octamer
sequenceisboundbytheubiquitous octamer-binding factor,
aswellasbyaB-cell-specificfactor foundonlyincellsof the B-lymphoid series. Sen and Baltimore (33) have proposed
that in vivo, a given segment of DNA is perhaps initially
activatedtomake thechromatinregionaccessibletobinding
factors. The tissue-specific binding proteins may play the
role ofactivation and thereby openthe DNA to interaction
with ubiquitous transcription-enhancing proteins. Our
site-directedmutagenesis experiments revealadifferentialeffect
of mutations in the proviral octamer sequence. Similar to
findings for immunoglobulingene octamer, mutations
maxi-mally affect the expression of the linked gene in B cells.
Thus, it seems likely that this octamer motif plays an
important role in tissue-specific expression of the RD-114 endogenous loci (5, 23-25, 30). This observation also indi-cates apotential utility of RD-114vectors forgenetransfer andexpression in targetcells. The newly defined enhancer elements, combined with the built-inoctamermotif and the unusual glycine-tRNAprimer-binding site(37)in the RD-114 vector, may add to the choice ofvectors forgene transfer into desired cell types to augment or replace a defective gene.
The mechanismsbywhich the RD-114 enhancer elements function to stimulate transcription remainto be elucidated. Additionalstudies will be required to characterize the cog-natesitesin the enhancer elementsthat interact with viralor cellular DNA-bindingproteins and to understand how pro-tein-protein interactionsmayberesponsiblefor the enhancer sequenceelements that function in combination.
ACKNOWLEDGMENTS
Wethank B. Bachman for expert technical assistanceaswellas for allcomputer-generated artwork. We also thank D. V. Kumarfor helpfulcomments,F.Miyagawa for typing, and E. Mader for critical reading ofthemanuscript.
This study was supported by Public Health Service grant CA 40590from the National Institutes of Health.
LITERATURECITED
1. Baluda, M. A., and P. Roy-Burman. 1973. Partial characteriza-tion of RD-114 virus by DNA-RNA hybridization studies. Nature(London)New Biol.244:59-62.
2. Banerji,J., L. Olson,and W. Schaffner. 1983. A lymphocyte-specific cellular enhancer is located downstream of the joining region in immunoglobulin heavy chaingenes. Cell33:729-740. 3. Benveniste,R.E., andG.J. Todaro. 1974.Evolution ofC-type
viral genes: inheritance ofexogenously acquired viral genes. Nature(London)252:456-459.
4. Berry, B. T., A. K. Ghosh, D. V. Kumar, D. A. Spodick, and P. Roy-Burman.1988.Structure and function of endogenousfeline leukemia virus long terminal repeats andadjoining regions.J. Virol. 62:3631-3641.
5. Busch, M. P., B. G. Devi, L. H. Soe, B. Perbal, M. A. Baluda, andP.Roy-Burman.1983.Characterization of the expression of cellular retrovirusgenesandoncogenesin feline cells. Hematol. Oncol. 1:61-75.
6. Celander, D., and W. A. Haseltine. 1984. Tissue-specific tran-scription preference as a determinantof cell tropism and leu-kaemogenic potential of murine retroviruses. Nature(London) 312:159-162.
7. DeFranco, D., and K. Yamamoto. 1986. Two different factors act separatelyortogethertospecify functionally distinct activities at asingle transcriptional enhancer.Mol.Cell. Biol. 6:993-1001. 8. DesGroseillers, L., and P. Jolicoeur. 1984. Thetandem direct repeats within the long terminal repeat of murine leukemia viruses are the primary determinant of their leukemogenic potential.J. Virol.52:945-952.
9. Falkner, F. G., and H. B. Zachau. 1984. Correct transcriptionof animmunoglobulin kappagenerequires an upstream fragment containingconserved sequenceelements.Nature (London) 310: 71-74.
10. Fletcher, C., N. Heintz, and R. G. Roeder. 1987. Purification and characterization ofOTF-1,atranscription factor regulating cell cycle expression ofahuman histone H2B gene. Cell 51:773-781. 11. Geisselsoder,J., F. Witney, and P. Yuckenberg. 1987. Efficient site-directedinvitromutagenesis. BioTechniques 5:786-791. 12. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.
Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol.2:1044-1051. 13. Graham,F.L.,and A.J. Van der Eb. 1973. A new technique for
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.64.304.74.133.2]14. Gruss, P., R. Dhar, and G. Khoury. 1981. Simian virus 40 tandemrepeated sequences as an element of the early promoter. Proc. Natl. Acad. Sci. USA 78:943-947.
15. Ishimoto, A., M. Takimoto, A. Adachi, M. Kakuyama, S. Kato, K. Kakimi, K. Fukuoka, T. Ogiu, and M. Matsuyama. 1987. Sequencesresponsible for erythroid and lymphoid leukemia in the long terminal repeats of Friend-mink cell focus-forming and Moloney murine leukemia viruses. J. Virol. 61:1861-1866. 16. Kunkel, T. A. 1985. Rapid and efficient site-specificmutagenesis
without phenotypic selection. Proc. Natl. Acad. Sci. USA 82:488-492.
17. Laimins, L. A., P. Gruss, R. Pozzatti, and G. Khoury. 1984. Characterization of enhancer elements in the long terminal repeatofMoloney murine sarcoma virus. J. Virol. 49:183-189. 18. Lenz, J., D. Calender, R. L. Crowther, R. Patarca, D. W. Perkins, and W. A. Haseltine. 1984. Determination of the leukaemogenicity ofa murine retrovirus by sequences within thelong terminalrepeat. Nature (London)308:467-470. 19. Levinson, B., G. Khoury, G. Vande Woude, and P. Gruss. 1982.
Activation of SV40genomeby 72-base pairtandem repeatsof Moloney sarcomavirus.Nature(London) 295:568-572. 20. Li, Y., E. Golemis, J. W. Hartley, and N. Hopkins.1987. Disease
specificity of nondefective Friend and Moloney murine leuke-miaviruses iscontrolled byasmall number of nucleotides. J. Virol. 61:693-700.
21. LoSardo, J. E., L. A. Cupelli, M. K. Short, J. W. Berman, and J. Lenz.1989. Differences in activities of murine retroviral long terminal repeats incytotoxic TlymphocytesandT-lymphoma cells. J. Virol.63:1087-1094.
22. Mason, J. O., G. T. Williams, and M. S. Neuberger. 1985. Transcription cell typespecificity is conferred by an immuno-globulin VHgene promoterthat includesafunctionalconsensus sequence.Cell41:479-487.
23. Niman, H. L., M. Akhavi, M. B. Gardner, J. R.Stephenson,and P. Roy-Burman. 1980. Differential expression oftwo distinct endogenous retrovirus genomes in developing tissues of the domesticcat. J. Natl. CancerInst. 64:587-594.
24. Niman, H. L., M. B. Gardner, J. R. Stephenson, and P. Roy-Burman. 1977. EndogenousRD-114 virusgenome expres-sion inmalignant tissues of domesticcats.J.Virol. 23:578-596. 25. Niman, H. L., J. R. Stephenson, M. B. Gardner, and P. Roy-Burman. 1977. RD-114 andfeline leukemia virus genome expression in natural lymphomas of domestic cats. Nature (London) 266:357-360.
26. Parslow, T.G., D. L.Blair,W.J.Murphy,and D. K. Granner. 1984.Structure of the5'endsofimmunoglobulingenes:anovel conservedsequence. Proc. Natl.Acad. Sci. USA81:2650-2654. 27. Reeves, R. H., W. G. Nash, and S. J.O'Brien. 1985. Genetic mapping of endogenousRD-114retroviralsequencesof domes-ticcats.J. Virol. 56:303-306.
28. Reeves, R. H., and S. J. O'Brien. 1984. Molecular genetic characterization of theRD-114 genefamilyofendogenous feline retroviralsequences. J. Virol.52:164-171.
Multiple functional motifs in the chicken Ul RNA gene en-hancer. Mol. Cell. Biol. 7:4185-4193.
30. Roy-Burman, P., M. P. Busch, S. Rasheed, M. B. Gardner, and M. M. C. Lai. 1980. Oncodevelopmental gene expression in felineleukemia, p. 361-372. In W. D. Hardy, Jr., M. Essex, and A. J. McClelland (ed.), Proceedings of the ThirdInternational Feline LeukemiaVirus Meeting. Elsevier/North-Holland Pub-lishing Co., New York.
31. Sanger, F., A. R. Coulson, B. G. Barrell, A. J. H. Smith, and B. A. Roe. 1980.Cloninginsingle-strandedbacteriophage as an aid torapidDNAsequencing.J. Mol. Biol. 143:161-178. 32. Sarma, P. S., J. Tseng, Y. K. Lee, and R. V. Gilden.1973. Virus
similartoRD-114virus incatcells. Nature(London)NewBiol. 244:56-59.
33. Sen, R., and D. Baltimore. 1986. Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell 46:705-716.
34. Singh, H., R. Sen, D. Baltimore, and P. A. Sharp. 1986. A nuclear factor that binds to a conserved sequence motif in transcriptional control elements of immunoglobulingenes. Na-ture (London)319:154-158.
35. Sive, H. L., N. Heintz, and R. G. Roeder. 1986. Multiple sequence elements arerequired for maximalin vitro transcrip-tion of a human histone H2B gene. Mol. Cell. Biol. 6:3329-3340. 36. Sive, H. L., and R. G. Roeder. 1986. Interaction ofa common factor with conserved promoter and enhancer sequences in histone H2B, immunoglobulin and U2 small nuclear RNA (SnRNA)genes. Proc. Natl. Acad. Sci. USA 83:6382-6386. 37. Spodick,D.A., A. K.Ghosh, S. Parimoo, and P.Roy-Burman.
1988. The long terminal repeat offeline enodgenous RD-114 retroviral DNAs: analysis of transcription regulatory activity and nucleotide sequence. Virus Res. 9:263-283.
38. Spodick,D. A., L. H. Soe,and P. Roy-Burman. 1984. Genetic analysis of the feline RD-114 retrovirus related endogenous elements. Virus Res. 1:543-555.
39. Staudt, L. M., R. G. Clerc, H. Singh, J. H. LeBowitz, P. A. Sharp,andD.Baltimore. 1988. Cloning ofalymphoid-specific cDNAencodingaprotein bindingtheregulatoryoctamerDNA motif. Science241:577-580.
40. Staudt,L.M.,H.Singh,R.Sen,T.Wirth,P. A.Sharp,and D. Baltimore. 1986. A lymphoid-specific protein binding to the
octamermotifofimmunoglobulingenes. Nature(London)323:
640-643.
41. Weiher, H., M. Konig, and P. Gruss. 1983. Multiple point mutations affecting the simian virus 40 enhancers. Science 219:626-631.
42. Weiss, R.,N.Teich,H.Varmus,andJ.Coffin(ed.).1985. RNA
tumor viruses: molecular biology of tumor viruses, 2nd ed.,
supplement and appendixes. Cold Spring Harbor Laboratory, Cold Spring Harbor,N.Y.
43. Zoller, M., and M. Smith. 1983.Oligonucleotide-directed
muta-genesis ofDNAfragments clonedinto M13 vectors. Methods Enzymol. 100:468-500.