JOURNAL OFVIROLOGY, Nov. 2011, p. 11274–11282 Vol. 85, No. 21 0022-538X/11/$12.00 doi:10.1128/JVI.05864-11
Copyright © 2011, American Society for Microbiology. All Rights Reserved.
A Conserved Peptide in West Nile Virus NS4A Protein Contributes
to Proteolytic Processing and Is Essential for Replication
䌤
§
R. L. Ambrose and J. M. Mackenzie*
Department of Microbiology, La Trobe University, Melbourne VIC, Australia 3086
Received 2 August 2011/Accepted 19 August 2011
The West Nile virus strain Kunjin virus (WNVKUN) NS4A protein is a multifunctional protein involved in
membrane proliferation, stimulation of cellular pathways, and evasion of host defense and is a major
com-ponent of the WNVKUN RNA replication complex. We identified a highly conserved region (
120
P-E-P-E123
) upstream of the viral protease dibasic cleavage site and investigated whether this motif was required for
WNVKUN replication. Single point mutations to alanine and a PEPE deletion mutation were created in a
full-length infectious WNVKUNmolecular clone. All mutations drastically impaired viral replication and virion
production, except that of the P122A mutant, which was slightly attenuated. These mutations were
subse-quently transferred to a WNVKUNreplicon to specifically assess effects on RNA replication alone. Again, all
mutants, except P122A, showed severely reduced negative-sense RNA production as well as decreased viral protein production. Correspondingly, immunofluorescence analyses showed a lack of double-stranded RNA
(dsRNA) labeling and a dispersed localization of the WNVKUNproteins, suggesting that replication complex
formation was additionally impaired. Attempts to rescue replication via conservative mutants largely failed except for substitution of Asp at E121, suggesting that a negative charge at this residue is equally important. Analysis of viral protein processing suggested that cleavage of the 2K peptide from NS4A did not occur with the mutant constructs. These observations imply that the combined effects of proline and negatively charged residues within the PEPE peptide are essential to promote the cleavage of 2K from NS4A, which is a prerequisite for efficient WNV replication.
West Nile virus strain Kunjin virus (WNVKUN) is an envel-oped, positive-sense RNA virus that can infect multiple bird species as well and human and horses via a mosquito vector. The replication cycle of WNVKUNinvolves a number of
spe-cific protein-protein and protein-membrane interactions that facilitate efficient genome amplification. We and others have arguably provided the most comprehensive interrogation of the WNVKUN replication complex (RC) and have provided
insight on the composition and interactions that occur during intracellular WNVKUNreplication (20, 21). The WNVKUNRC
itself is composed of vesicles that have invaginated from the ER membrane (3), whereas a second WNVKUN-induced
mem-brane structure (convoluted memmem-branes/paracrystalline arrays [CM/PC]) is proposed to partake in polyprotein translation and processing (11).
One of the WNVKUNsmall hydrophobic proteins,
nonstruc-tural protein 4A (NS4A), has been identified as a key regulator of membrane proliferation and inducer of cell signaling path-ways and immune evasion as well as forming part of the RC (1, 9, 12–14, 16). NS4A is a 16-kDa protein that has an N-terminal cytoplasmic region, three transmembrane regions, and one membrane-associated region at its C terminus (Fig. 1A) (2, 14). It is cleaved from the viral polyprotein by the action of two proteases: (i) the NS2B-3 viral protease at the N terminus and
upstream of a region termed “2K” and (ii) at the C terminus by a luminal endoplasmic reticulum signalase (6). Some studies have indicated a requirement for NS2B-3 cleavage at the 2K site before the host signalase proteolysis downstream in the ER lumen (6, 16).
NS4A is localized to ER membranes during WNVKUN
replication (12) due to its highly hydrophobic nature. Pro-tein interaction studies have shown that it can interact with NS5, NS3, and NS2A as well as forming homodimers (12), leading to a proposed role during viral RNA replication (5). Additionally, a genetic interaction between NS4A and NS1 was identified in yellow fever virus (YFV) (8), revealing further associations between NS1 and NS4A. The topology of NS4A has been predicted using glycosylation studies (14) which showed that the N terminus is located on the cyto-plasmic face, with a transmembrane helix spanning the membrane to the lumenal side and a membrane-associated helix followed by a second transmembrane region which releases the NS2B-3 cleavage site back out to the cytoplasm. The proposed 2K region is also highly hydrophobic and is proposed to act as a signal peptide for correct NS4B cleav-age and topology (6).
In this paper, we identify a small region (PEPE) in the hydrophobic C terminus of NS4A that is almost identical among the flaviviruses surveyed. We also show that mutation of this region in the full-length WNVKUNgenome completely
abolishes replication and virion production and that this is most likely due to defects in negative- and positive-sense RNA transcription. Additionally, mutations in this region perturb NS4A proteolytic processing, in particular at the NS2B-3 cleav-age site upstream of the 2K transmembrane region. Specifi-cally, the first proline and downstream glutamic acid residues * Corresponding author. Mailing address: Department of
Micro-biology, La Trobe University, Melbourne VIC, Australia 3086. Phone: 613 9479 2225. Fax: 613 9479 1222. E-mail: j.mackenzie @latrobe.edu.au.
§ Supplemental material for this article may be found at http://jvi .asm.org/.
䌤Published ahead of print on 31 August 2011.
11274
on November 7, 2019 by guest
http://jvi.asm.org/
are critical for virus replication and, we hypothesize, are re-quired to project the cleavage site from the embedded mem-brane region, thus allowing NS4A(⫺2K) generation and its roles in replication.
MATERIALS AND METHODS
Cells, viruses, and antibodies.HEK 293T and BHK cells were maintained in high-glucose Dulbecco modified Eagle medium (DMEM) (Gibco) supplemented with 5% fetal bovine serum (FBS) (Serum Supreme; Lonza), 1% penicillin/ streptomycin (Gibco), and 1% Glutamax (Gibco). Puromycin selection was per-formed at 5 or 10g/ml for 293T and Vero cells, respectively, in the presence of 2% FBS and 1% Glutamax. West Nile virus Kunjin strain MRM61C (WNVKUN) was propagated in Vero cells for 40 h, and tissue culture fluid was centrifuged at 1,600 rpm to remove cell debris. Virus titer was determined by plaque assay as previously described (1). Mouse anti-NS1 and NS5 monoclonal antibodies were a kind gift from Roy Hall (University of Queensland). WNVKUN-specific rabbit anti-NS3 and anti-NS4A polyclonal antisera were kindly provided by Alexander Khromykh (12, 19). Mouse anti-dsRNA (clone J2) and rabbit anti-actin antibod-ies were purchased from English & Scientific Consulting Bt. (Hungry) and Sigma, respectively.
Site-directed mutagenesis of WNVKUNinfectious clones and replicons.Ala mutations were generated in the WNVKUNcDNA infectious clone, FLSDX (4), using site-directed mutagenesis (Stratagene) with the primers described in Table S1 in the supplemental material. These mutations were transferred to pKUNrep4 (17) by restriction digest withBstBI(NEB) and AgeI (Pro-mega). Conservative mutations were generated in P120 and E121 using site-directed mutagenesis as described above, except that an extra silent mutation
was introduced in the primers for a unique restriction site (AseI) to aid in clone selection. The equivalent mutation was incorporated in pKUNrep4 (pKUNrep4 AseI) in the absence of other mutations to control for effects on replication. All clones were sequenced prior to maxiprep amplification (In-vitrogen) and transfection.
In vitrotranscription and electroporation.FLSDX clones were linearized by XhoI and DNA purified using phenol-chloroform extraction. RNA was tran-scribed from the FLSDX vector using SP6 RNA polymerase (NEB) and elec-troporated into 2⫻106
BHK cells using an exponential protocol (2⫻750-V pulses, 25F, infinite resistance) on the Bio-Rad Genepulser II. Cells were recovered in growth medium (as described above) at room temperature for 5 min and then plated out into six-well plates. Cell lysates and tissue culture fluid samples were collected at 48, 72, and 96 h postelectroporation (hpe) as described below.
[image:2.585.43.537.66.386.2]Transfection of mammalian cells.pKUNrep4 DNA was transfected into 293T cells and Vero cells using Lipofectamine 2000 (Invitrogen). Briefly, 4g of each pKUNrep4 clone was precipitated using 5 M sodium acetate and 100% ethanol and resuspended in 10l of elution buffer (Tris, pH 8.0; Qiagen). DNA was combined with DMEM and 10l of Lipofectamine to a total volume of 500l and incubated at room temperature (RT) for 40 min. 293T cells were washed once in DMEM and transfected with the DNA-Lipofectamine complexes and incubated at 37°C and 5% CO2for 24 h. Expression of the replicon was selected for by treatment with 5g/ml or 10g/ml puromycin (Sigma) in 2% FBS (Lonza) and 1% Glutamax (Gibco) in DMEM for 293T and Vero cells, respectively. At 3 days posttransfection (dpt), adherent cells were resuspended in fresh medium as described above and split equally into three wells of a six-well plate to further select for transfectants. At 6 dpt, adherent cells were collected for RNA, protein, and immunofluorescence analysis.
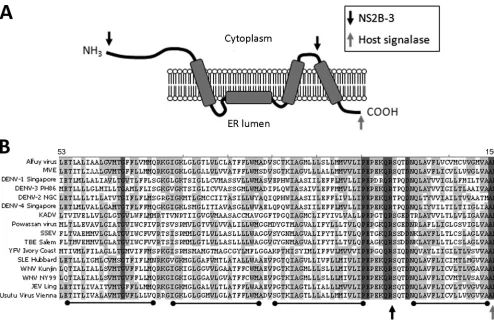
FIG. 1. NS4A120P-E-P-E123region is highly conserved in theFlavivirusgenus. (A) Predicted topology of NS4A in the endoplasmic reticulum membrane as described by Miller et al. (15). NS4A is cleaved from the viral polyprotein at the N terminus and at the C terminus upstream of the 2K transmembrane region by NS2B-3 (cytoplasmic face, indicated by the black arrow). The NS4A-4B junction is cleaved by a host signalase in the ER lumen (indicated by the gray arrow). (B) Various NS4A sequences from theFlavivirusgenus were aligned using ClustalW software (Vector NTI; Invitrogen). Dark gray shading indicates identical residues, while light gray residues are conservative. Transmembrane regions are shown by black bars below the alignment, and cleavage sites are indicated as for panel A.
on November 7, 2019 by guest
http://jvi.asm.org/
Immunofluorescence analysis.Cells were fixed by treatment with acetone-methanol (1:1) at⫺20°C for 7 min. Following aspiration with phosphate-buffered saline (PBS), cells were incubated with primary antibodies in 1% bovine serum albumin (BSA)-PBS for 1 h at RT. Cells were washed four times in 0.2% BSA-PBS and then incubated with species-specific secondary antibodies conju-gated to either Alexa Fluor 594 or 488 in 1% BSA-PBS at RT for 45 min. Cells were washed in 0.1% BSA-PBS and then PBS, followed by a 5-min incubation with 4g/ml DAPI to counterstain the nuclei. Coverslips were mounted with Ultramount (Fronine); immunofluorescent staining was visualized on a Zeiss confocal microscope, and figures were assembled using Adobe Photoshop.
Western blot analysis.Cells were washed once with PBS to remove debris and dead cells and then lysed with SDS lysis buffer (0.1% SDS, 0.5 mM EDTA, 10 mM Tris, pH 8.0, 150 mM NaCl) containing protease inhibitors (protease inhib-itor cocktail III; Astral Scientific) and pipetted multiple times to reduce viscosity. Lysates were loaded on a 12% Tris-glycine polyacrylamide gel and separated at 100 V for 2 h. Separated proteins were transferred to a Hi-Bond ECL nitrocel-lulose membrane (Amersham) at 100 V for 75 min and blocked at RT in 6% skim milk-TBS (Diploma) with 0.05% Tween (TBS-T). Following blocking, mem-branes were washed for 15 min in TBS-T and then incubated overnight in primary antibodies diluted in 6% BSA–TBS-T at 4°C. Membranes were washed in TBS-T three times and then incubated with species-specific secondary anti-bodies conjugated to either Alexa Fluor 488 or 647 at RT for 2 h. Following two washes in TBS-T and one wash in TBS, membranes were viewed on the Storm fluorescent scanner (Amersham) on either a 635 nM or a 430 nM emission channel.
RNA extractions and qPCR analyses.Cells were rinsed in PBS and then lysed in 1 ml TRIzol reagent (Invitrogen), and RNA was extracted according to manufacturers’ specifications. Total RNA concentration was quantified using a Qubit fluorometer (Invitrogen), and 4g was treated with RQ1 DNase (Pro-mega) at 37°C for 30 min. cDNA was reverse transcribed from treated RNA with either WNVKUNsense or antisense primers and an internal control (see Table S2 in the supplemental material for sequences) using Superscript III (Invitrogen) at 50°C for 50 min. Following heat inactivation at 65°C, RT reactions were diluted in diethyl pyrocarbonate (DEPC)-treated deionized water. cDNA levels were quantified by quantitative PCR (qPCR) with Sybr GreenER (Bio-Rad) (cycling conditions: 50°C for 8 min, 95°C for 2 min, 40 cycles of 15 s at 95°C, 1 min annealing/extension at 60°C, followed by a final extension of 10 min), and ana-lyzed with ICycler software (Bio-Rad). Fold induction of sense or antisense RNA was calculated compared to results for the housekeeping gene RPL13A, and error bars indicate duplicate experiments. Studentttests were used to determine significant changes between samples (Microsoft Excel), and asterisks indicate a
Pvalue of⬎0.01.
RESULTS
NS4A120P-E-P-E123region is highly conserved in the
Flavi-virusgenus.To identify regions of NS4A important for repli-cation, we undertook an alignment study, with the rationale that vital sequences would be highly conserved across the fla-viviruses. At least two NS4A sequences for each species within
the Flavivirus genus were aligned using ClustalW, and
con-served areas were compared to the published topology regions (14) (Fig. 1B and A, respectively). Several highly conserved residues were identified throughout the cytoplasmic and hy-drophobic regions, such as glycine 68 and prolines 13 and 48. Strikingly, a small region of four residues (proline 120 [P120], glutamic acid 121 [E121] proline 122 [P122], and glutamic acid 123 [E123], referred to as PEPE from here on) was found to be almost completely conserved among all flaviviruses surveyed. This region was of particular interest for two reasons: it was found to be upstream of the predicted 2K cleavage site but just downstream of a predicted transmembrane helix, and it contained two conserved prolines (Pro), which have been previously reported to be important for the signal peptidase site at the C-prM junction (10). Given its high level of conservation and close proximity to the NS2B-3 cleavage
site, we hypothesized that this PEPE sequence could be important for WNVKUNreplication.
Alanine mutations within the PEPE motif abolish
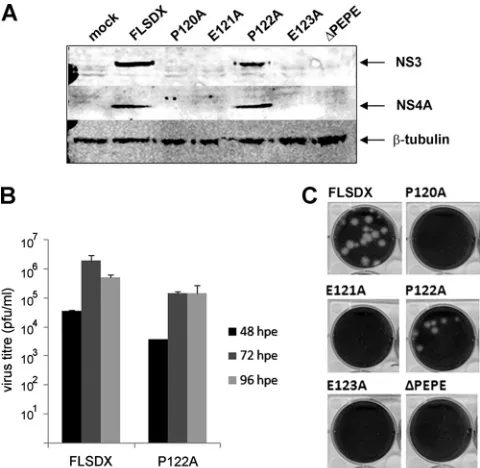
replica-tion from a full-length WNVKUNinfectious clone.To assess the
importance of the PEPE region in WNVKUNreplication, sin-gle-nucleotide substitutions were introduced in a full-length infectious clone, FLSDX, to change each residue separately to alanine (Ala), or the region was deleted entirely (⌬PEPE).In
vitrotranscribed RNA was then electroporated into BHK cells,
[image:3.585.300.540.69.303.2]and lysates and tissue culture fluid samples were collected at 48, 72, and 96 hpe. Interestingly, all mutants but one (P122A) showed complete inhibition of replication compared to the wild-type (FLSDX), as assessed by Western blotting at 72 hpe (Fig. 2A). Accordingly, no virus plaques were observed after reinfection of collected tissue culture fluid onto Vero cells at any time points analyzed (Fig. 2B). Attempts made at later time points (i.e., 7 days postelectroporation) also failed to detect secreted virions (data not shown). The P122A mutant did produce plaques, although plaque size was smaller (Fig. 2B) and viral titers at 48 and 72 hpe were reduced by approx-imately 1 log compared to FLSDX. This result was not entirely unexpected, as P122 was not as tightly conserved as P120, suggesting that this residue was not critical for WNVKUN rep-lication or virion secretion.
FIG. 2. Alanine mutants within the PEPE motif abolish replication from a full-length WNVKUN infectious clone. Single Ala mutations were generated in the PEPE region in FLSDX using site-directed mutagenesis, or the region was deleted entirely (⌬PEPE). RNA was transcribed and electroporated into BHK cells, and tissue culture fluid and protein lysate samples were collected at 48, 72, and 96 hpe. (A) Protein lysates from 72 hpe were analyzed by Western blotting using the viral antibodies NS4A and NS3 compared to the internal control-tubulin. (B) Tissue culture fluid samples at each time point were analyzed by plaque assay, and visible plaques were used to cal-culate titers for the WT and P122A mutant. Error bars indicate results of replicate experiments. (C) Plaques in the 10⫺5dilution were ana-lyzed for size difference.
11276 AMBROSE AND MACKENZIE J. VIROL.
on November 7, 2019 by guest
http://jvi.asm.org/
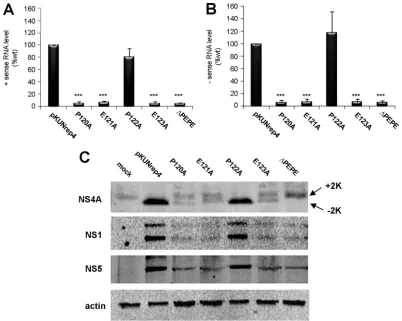
Mutation of the PEPE region affects WNVKUN RNA
repli-cation and NS4A proteolytic processing.Although the
conse-quences of mutating the PEPE region on WNVKUN repli-cation were clearly evident, it was not known if the defect was at the transcriptional or translational level or even at the stage of RC formation. To determine this, the mutations made in FLSDX were transferred to a WNVKUN replicon (pKUNrep4), which contains the essential RNA elements for replication and the coding region of the nonstructural proteins but not that of the major structural proteins E and prM, all under the control of a CMV promoter, as well as the puromycin resistance gene for selection. Expression from this cassette allows robust WNVKUNRNA replication, viral protein translation, and RC formation but not virus assembly. 293T cells were transfected with the cDNA con-structs with or without the specific mutations described above, and expressing cells were selected with puromycin treatment from 24 hpt onwards. RNA samples at 6 dpt were analyzed for negative- and positive-sense viral transcripts using complementary and reverse complementary primers with qPCR. Compared to levels in the pKUNrep4 control, both negative and positive transcript levels were
dramati-cally reduced in all mutants except P122A, which was slightly attenuated but not significantly so (Fig. 3A). To confirm that the detection of negative-sense transcripts was not due to mis-annealing of primers or background ampli-fication, RNA was treated with DNases and no RT controls were included in the analysis, with all controls having at least seven CT values lower than the test samples (data not shown).
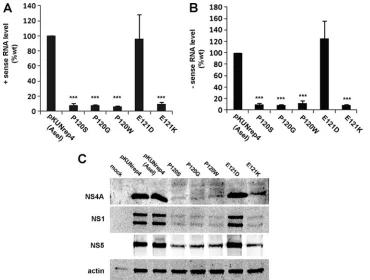
Given that RNA replication in the mutant samples was sig-nificantly impaired, it was also expected that viral protein levels in expressing cells would correspondingly be reduced. Western blotting for the WNVKUNNS1, NS5, and NS4A proteins was
[image:4.585.90.492.68.389.2]utilized to determine whether these proteins were being ex-pressed properly despite the mutations. As shown in Fig. 3B, overall viral protein levels were drastically reduced in all mu-tants except P122A. Interestingly, two bands were detected by the NS4A antibody in the mutant constructs that displayed defective replication. The size difference between these two NS4A protein species was estimated to be 2 kDa. In compar-ison, pKUNrep4 and P122A samples had only one protein band at the lower molecular size (⬃14 kDa; Fig. 3B). Although it cannot be definitively proven, given the specificity of the FIG. 3. Mutations in the PEPE region affect WNVKUNRNA replication and NS4A proteolytic processing. PEPE mutations in FLSDX were transferred to a WNVKUNreplicon, pKUNrep4, from which transcription is driven by the CMV promoter. 293T cells were transfected with the mutant plasmids and placed under puromycin selection (5g/ml) at 1 dpt, and RNA and protein lysates collected at 6 dpt. (A and B) RNA was extracted and analyzed for sense (A) and antisense (B) viral RNA levels using qPCR and compared to the internal control RPL13A. Transcript levels were normalized to WT levels, and error bars indicate⫹1 standard deviation from duplicate experiments.*,Pⱕ0.01. (C) Protein lysates were analyzed by Western blotting for viral proteins NS1, NS4A, and NS5 and compared to the internal control actin. Arrows indicate different species of NS4A (⫹2K,⫺2K).
on November 7, 2019 by guest
http://jvi.asm.org/
anti-NS4A antibody and the 2-kDa size difference observed, we are confident that the two bands observed correspond to NS4A(⫹2K) and NS4A(⫺2K). This suggested that processing at the “NS4A-2K” site was affected by mutations in the up-stream PEPE region, resulting in two different forms of NS4A [i.e., NS4A(⫹2K) and NS4A(⫺2K)] being present in mutant replicon-expressing cells.
Ala substitutions within the PEPE motif prevent RC
forma-tion.Although both viral RNA and protein production were
severely restricted by mutations in the PEPE region, we were intrigued to observe whether the low levels of viral proteins still resulted in formation of RCs, or whether the replication defect could be attributed to a lack of RCs. Vero cells were transfected with the same mutant replicons as above, selected with puromycin, and then fixed and labeled with antibodies to dsRNA or the WNVKUNproteins (Fig. 4A and B). In
corre-lation with the qPCR data, no dsRNA was detectable in the Ala mutants except for P122A and pKUNrep4 replicons, sug-gesting that replicative RNA intermediates were not present when PEPE was mutated. Additionally, NS3, which usually colocalizes with dsRNA during replication in vesicle packets (19), accumulated in discrete cytoplasmic foci in the P120A, E121A, E123A, and⌬PEPE mutants (Fig. 4A). Interestingly, when we determined the localization of NS4A in these cells, we also observed accumulation in the cytoplasm, suggesting that replicative proteins are redirected or mis-located in the ab-sence of RNA replication. NS1 localization remained relatively unchanged between samples (Fig. 4B) and could still be visu-alized at the periphery of the cell. However, NS1 and NS4A did not colocalize in the P120A, E121A, E123A, and⌬PEPE mu-tants, compared to results in pKUNrep4- and P122A-trans-fected cells (Fig. 4B) Overall, our results indicate that Ala FIG. 4. Ala substitutions within the PEPE motif prevent RC formation. Vero cells were transfected with the mutant pKUNrep4 plasmids and selected with 10g/ml puromycin for 6 days. Fixed cells were then labeled with either (A) NS3 (in green) and dsRNA (in red) or (B) NS4A (in green) and NS1 (in red). Images were collected on a confocal microscope; yellow in the merge images shows colabeling.
11278 AMBROSE AND MACKENZIE J. VIROL.
on November 7, 2019 by guest
http://jvi.asm.org/
mutations to P120, E121, and E123 within the PEPE region resulted in decreased RNA and protein levels as well as dis-association of replicative components, leading to a severe de-fect in replication.
Conservative mutations at E121 partially restore WNVKUN
RNA replication and RC formation. We were interested to
investigate whether conservative changes within the PEPE re-gion could be tolerated and thus restore WNVKUNRNA rep-lication. Previous studies have shown that Pro residues have vital␣-helical breaking properties in transmembrane helices (18) as do Gly and Ser. To identify whether this particular activity is important for replication, P120 was mutated to Ser (P120S) or Gly (P120G) as well as Tyr (P120W) as a control. Additionally, E121 was mutated to Asp (E121D) or Lys (E121K) to determine if a particular charge is required at this site. Transfected cells were analyzed at 6 dpt by qPCR, West-ern blotting, and immunofluorescence as described for the Ala mutants described above. We observed that neither the P120S, the P120G, the P120W, nor the E121K mutant demonstrated a restoration in positive- or negative-sense RNA transcription (Fig. 5A and B), and only the conservative change to Asp (E121D) displayed wild-type (WT) levels of RNA replication. Similarly, viral protein synthesis was dramatically decreased in all mutants except E121D (Fig. 5C). Immunofluorescence as-says showed that RC formation was also inhibited in the con-servative mutants; dsRNA labeling was absent in all mutants except E121D, and NS3 localization was very distinct from that
of the WT, forming aggregates in the cytoplasm, as was ob-served in the Ala mutants (compare Fig. 6A and Fig. 4A). The colocalization of NS1 and NS4A was also not observed in the P120S, P120G, P120W, or E121K mutant (Fig. 6B), indicating, as with the Ala mutants, that RC formation presumably did not occur.
DISCUSSION
NS4A is a highly multifunctional protein during WNVKUN replication, with roles in replication, membrane proliferation and recruitment, and regulation of cell and immune signaling. This is surprising given its small size (16 kDa) and high level of hydrophobicity, which ensures that a large part of the C ter-minus is embedded in reticular membranes. Accordingly, this protein also has a high level of sequence similarity between various members of theFlavivirusgenus, suggesting that the functional regions of NS4A are quite closely con-served. We identified a strongly conserved tetrapeptide se-quence, 120PEPE123, in the hydrophobic C terminus of
NS4A, a few residues upstream of the internal 2K site, which is cleaved by the viral protease NS2B-3. Site-directed mu-tagenesis or complete removal of this region had severe effects on WNVKUN replication. Our results suggest that correct processing of NS4A is a critical step in the biogen-esis of the WNVKUNRC on the ER membrane.
The location and sequence of the PEPE peptide are inter-FIG. 5. Conservative mutations at E121 partially restore WNVKUNRNA replication and RC formation. Conservative changes to the PEPE motif in pKUNrep4(AseI) were generated using site-directed mutagenesis. 293T cells were transfected with the mutant plasmids and placed under puromycin selection (5g/ml) at 1 dpt, and RNA and protein lysates were collected at 6 dpt. (A and B) RNA was extracted and analyzed for sense (A) and antisense (B) viral RNA levels using qPCR, compared to levels in the internal control RPL13A. Transcript levels were normalized to WT levels, and error bars indicate⫹1 standard deviation from duplicate experiments.*,Pⱕ0.01. (C) Protein lysates were analyzed by Western blotting for viral proteins NS1, NS4A, and NS5 and compared to the internal control actin.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.108.478.68.348.2]esting for a number of reasons; transmembrane prediction studies suggest that PEPE is located on a cytoplasmic loop between two highly hydrophobic transmembrane helices (Fig. 1A). Also important is this region’s close proximity to the downstream NS2B-3 cleavage site, which must also be pre-sented to the cytoplasm for interaction with the viral protease. Finally, the presence of two Pro residues interspersed with the highly charged Glu residues, combined with the almost iden-tical conservation throughout the genus, suggested that this region would be important for NS4A function and warranted further study. Our mutational analyses revealed that all the Ala mutations generated completely inhibited WNVKUN replica-tion, except for a single mutant, P122A. This mutant was slightly attenuated at earlier time points, as demonstrated by the viral titer and plaque size, and showed reduced viral
pro-tein levels. This may be due to the position of this Pro in the cytoplasmic loop, flanked as it is by two highly charged resi-dues. Interestingly, when we reexamined the alignment from Fig. 1, we found that this “mutation” was naturally occurring in the tick-borne encephalitis group, reinforcing our data and suggesting that P122 is not essential for WNV replication.
[image:7.585.79.503.70.493.2]However, the remaining mutants were severely attenuated and showed no protein or virus production whatsoever, sug-gesting that the defect occurred during replication. To study this further, the mutations were transferred to a replicon sys-tem (pKUNrep4) from which viral RNA transcription is driven from a CMV promoter. As we hypothesized, both negative-and positive-sense RNA transcription processes were inhib-ited, with equally low levels of viral protein production and a lack of RC formation. As there were low levels of viral proteins FIG. 6. Conservative mutations at E121 partially restore WNVKUNRNA replication and RC formation. Vero cells were transfected with the mutant pKUNrep4 plasmids and selected with 10g/ml puromycin for 6 days. Fixed cells were then labeled with either (A) NS3 (in green) and dsRNA (in red) or (B) NS4A (in green) and NS1 (in red). Images were collected on a confocal microscope; yellow in the merge images shows colabeling.
11280 AMBROSE AND MACKENZIE J. VIROL.
on November 7, 2019 by guest
http://jvi.asm.org/
present, it could be assumed that some transcription/transla-tion of viral genes was occurring, although this is most likely due to direct transcription from the pKUNrep4 plasmid. There were also comparable reductions of negative- and positive-sense RNA, suggesting that only a small amount of negative-sense RNA was being transcribed from a positive-negative-sense tem-plate. However, we believe this was only occurring in minute amounts, as dsRNA replicative intermediates, indicative of replication, were not observed by immunofluorescence in the mutant replicon-expressing cells. The defects in RNA replica-tion described may have been due to the low overall level of viral protein production, which in turn would have been caused by low RNA levels. Or it may be a direct effect of mutating NS4A, which impeded RNA RC formation, reducing RNA transcription and thus viral protein translation.
Surprisingly, NS4A processing at the C terminus was also affected by the Ala mutations, resulting in the detection of “full-length” NS4A (with the 2K peptide) and NS4A (without the 2K peptide, compared to the FLSDX and P122A mutants, which showed only NS4A(⫺2K). This suggested that NS2B-3 cleavage downstream of PEPE was altered by these mutations. There are two possible explanations for this; either the chron-ically low levels of viral proteins were inefficiently processing the equally low levels of NS4A or the mutations were prevent-ing NS2B-3 protease accessibility and thus activity due to the proximity to the cleavage site. We are inclined to believe the latter, as NS1, and NS5 processing was unaffected, and to believe that NS3 and NS4A subcellular localization was also affected by mutations in the PEPE region. This is an interesting result, as it implies that correct and efficient processing of NS4A into NS4A(⫺2K) is required for efficient RNA
replica-tion, which has not been clearly assessed before. It also sup-ports our previous work that showed that the two different NS4A protein species can localize to separate subcellular com-partments (16). Extrapolating these data could also suggest a mechanism by which NS4A can have multiple roles during WNVKUNreplication, via differential cleavage of NS4A.
Formation of RCs was also not observed in mutant repli-cons. Instead, protein aggregates of NS3 and NS4A accumu-lated in the cytoplasm, away from their usual perinuclear/ reticular location observed during WNVKUN replication (12, 19). Additionally, colocalization between NS1 and NS4A was not observed in the Ala or conservative mutants, although NS1 localization did not dramatically change (Fig. 4B and 6B). Together with the replication data, these results suggest that correctly processed NS4A is likely responsible for the assembly of the RC, possibly via interaction with NS1 on the ER mem-brane, which is followed by RNA replication (Fig. 7). Based on a model that had been only proposed previously (5, 20), we suggest now that processing of NS4A to NS4A(⫺2K) is a necessary step to allow association of the various replicative proteins to enable efficient replication. Thus, we propose that in the presence of NS4A(⫹2K) (or absence of NS2B-3 pro-cessing), the interaction between NS4A and NS1, and poten-tially between NS4A and the RC, does not occur (Fig. 7). Two previous observations support this model: (i) we have observed that NS4A and NS2A appear to interact only when replicating WNVKUNRNA is available (12), and (ii) the Rice laboratory
demonstrated a genetic interaction between NS4A and NS1 (7). We are currently investigating the protein-protein interac-tions that are occurring during this scenario.
[image:8.585.112.471.68.300.2]As the Ala mutations in the PEPE region had such severe FIG. 7. Mutations in the PEPE region of NS4A inhibit NS2B-3 binding, cleavage, and subsequent replicative function. We predict that the helix-breaking properties of Pro and downstream negative charges in the PEPE region may be important in promoting the KQRS cleavage site to the viral protease, NS2B-3. Once processed, NS4A(⫺2K) is able to interact with NS1 within the ER lumen and the other cytoplasmic replicative components to form the replication complex. When these residues are mutated, we propose that the KQRS cleavage site remains associated with the membrane and is inaccessible to NS2B-3. The uncleaved NS4A(⫹2K) protein is not able to associate with components of the replication complex, thus preventing RNA replication and high-level viral protein synthesis.
on November 7, 2019 by guest
http://jvi.asm.org/
effects on replication, attempts were made to generate more conservative mutants, with the expectation that an intermedi-ate phenotype would be observed. Interestingly, only one mu-tation could rescue replication, and that was an equal substi-tution of negative charge at E121. Surprisingly, none of the changes to the initial P120 (Ser, Gly, or Tyr) were able to replicate efficiently, and they showed replication defects simi-lar to those of the P120A mutant. Pro is a unique amino acid, in that the␣-nitrogen of the peptide is incorporated into a rigid ring structure, which introduces a kink in alpha helices (18, 22). This characteristic makes it a vital component in protein fold-ing, as well as providing a “helix-breaking” function for trans-membrane segments (18). Given that P120 in NS4A is directly adjacent to a strongly hydrophobic helix, we suggest that it may play a role in “breaking” the transmembrane helix and releas-ing the downstream peptides from the membrane into the cytoplasm. As this is also where NS2B-3 processing occurs, this Pro in combination with the adjacent negatively charged Glu residue would thus be vital for exposure of the KQRS cleavage site to the viral protease. This hypothesis is supported by the incorrect processing of NS4A observed in the mutant-express-ing cells.
Overall, we have shown the importance of the PEPE region in NS4A for WNVKUN replication complex formation and
RNA replication. Given the high degree of conservation be-tween the various members of the Flavivirus genus, it could also be proposed that this sequence and its role in NS4A cleavage would be essential for many other viruses, such as Japanese encephalitis virus or dengue virus. This work pro-vides a unique insight into the essential roles of NS4A during WNVKUNreplication and the requirement for correct process-ing in the assembly of the replication complex.
ACKNOWLEDGMENTS
We thank Alexander Khromykh for generously providing the WNVKUN infectious clones and replicons and for the rabbit NS3 and anti-NS4A antibodies. We also thank Roy Hall for providing mouse mono-clonal anti-NS1 and anti-NS5 antibodies.
This research was supported by a Project Grant (no. 1004619) to J.M. from the National Health and Medical Research Council of Australia.
REFERENCES
1.Ambrose, R. L., and J. M. Mackenzie.2011. West Nile Virus differentially modulates the unfolded protein response to facilitate replication and im-mune evasion. J. Virol.85:2723–2732.
2.Coia, G., M. D. Parker, G. Speight, M. E. Byrne, and E. G. Westaway.1988. Nucleotide and complete amino acid sequences of Kunjin virus: definitive
gene order and characteristics of the virus-specified proteins. J. Gen. Virol.
69:1–21.
3.Gillespie, L. K., A. Hoenen, G. Morgan, and J. M. Mackenzie.2010. The endoplasmic reticulum provides the membrane platform for biogenesis of the flavivirus replication complex. J. Virol.84:10438–10447.
4.Khromykh, A. A., M. T. Kenney, and E. G. Westaway.1998.trans -Comple-mentation of flavivirus RNA polymerase gene NS5 by using Kunjin virus replicon-expressing BHK cells. J. Virol.72:7270–7279.
5.Khromykh, A. A., P. L. Sedlak, K. J. Guyatt, R. A. Hall, and E. G. Westaway.
1999. Efficienttrans-complementation of the flavivirus kunjin NS5 protein but not of the NS1 protein requires its coexpression with other components of the viral replicase. J. Virol.73:10272–10280.
6.Lin, C., S. M. Amberg, T. J. Chambers, and C. M. Rice.1993. Cleavage at a novel site in the NS4A region by the yellow fever virus NS2B-3 proteinase is a prerequisite for processing at the downstream 4A/4B signalase site. J. Vi-rol.67:2327–2335.
7.Lindenbach, B. D., and C. M. Rice.1999. Genetic interaction of flavivirus nonstructural proteins NS1 and NS4A as a determinant of replicase function. J. Virol.73:4611–4621.
8.Lindenbach, B. D., and C. M. Rice.1997.trans-Complementation of yellow fever virus NS1 reveals a role in early RNA replication. J. Virol.71:9608– 9617.
9.Liu, W. J., et al.2005. Inhibition of interferon signaling by the New York 99 strain and Kunjin subtype of West Nile virus involves blockage of STAT1 and STAT2 activation by nonstructural proteins. J. Virol.79:1934–1942. 10.Lobigs, M., and E. Lee.2004. Inefficient signalase cleavage promotes efficient
nucleocapsid incorporation into budding flavivirus membranes. J. Virol.78:
178–186.
11.Mackenzie, J.2005. Wrapping things up about virus RNA replication. Traffic
6:967–977.
12.Mackenzie, J. M., A. A. Khromykh, M. K. Jones, and E. G. Westaway.1998. Subcellular localization and some biochemical properties of the flavivirus Kunjin nonstructural proteins NS2A and NS4A. Virology245:203–215. 13.Mertens, E., et al.2010. Viral determinants in the NS3 helicase and 2K
peptide that promote West Nile virus resistance to antiviral action of 2⬘,5⬘ -oligoadenylate synthetase 1b. Virology399:176–185.
14.Miller, S., S. Kastner, J. Krijnse-Locker, S. Buhler, and R. Bartenschlager.
2007. Non-structural protein 4A of Dengue virus is an integral membrane protein inducing membrane alterations in a 2K-regulated manner. J. Biol. Chem.282:8873–8882.
15.Miller, S., S. Sparacio, and R. Bartenschlager.2006. Subcellular localization and membrane topology of the Dengue virus type 2 non-structural protein 4B. J. Biol. Chem.281:8854–8863.
16.Roosendaal, J., E. G. Westaway, A. Khromykh, and J. M. Mackenzie.2006. Regulated cleavages at the West Nile virus NS4A-2K-NS4B junctions play a major role in rearranging cytoplasmic membranes and Golgi trafficking of the NS4A protein. J. Virol.80:4623–4632.
17.Varnavski, A. N., P. R. Young, and A. A. Khromykh.2000. Stable high-level expression of heterologous genes in vitro and in vivo by noncytopathic DNA-based Kunjin virus replicon vectors. J. Virol.74:4394–4403. 18.von Heijne, G.1991. Proline kinks in transmembrane␣-helices. J. Mol. Biol.
218:499–503.
19.Westaway, E. G., J. M. Mackenzie, M. T. Kenney, M. K. Jones, and A. A. Khromykh.1997. Ultrastructure of Kunjin virus-infected cells: colocalization of NS1 and NS3 with double-stranded RNA, and of NS2B with NS3, in virus-induced membrane structures. J. Virol.71:6650–6661.
20.Westaway, E. G., J. M. Mackenzie, and A. A. Khromykh.2003. Kunjin RNA replication and applications of Kunjin replicons. Adv. Virus Res.59:99–140. 21.Westaway, E. G., J. M. Mackenzie, and A. A. Khromykh.2002. Replication and gene function in Kunjin virus. Curr. Top. Microbiol. Immunol.267:323– 351.
22.Williams, K. A., and C. M. Deber.1991. Proline residues in transmembrane helices—structural or dynamic role. Biochemistry30:8919–8923.
11282 AMBROSE AND MACKENZIE J. VIROL.