Copyright@1975 American Society for Microbiology Printedin U.S.A.
Replication
of
Polyoma
DNA
in
Isolated
Nuclei
V.
Complementation
of In Vitro DNA
Replication
BERND OTTO AND PETER REICHARD*
Medical NobelInstitute, Departmentof Biochemistry, KarolinskaInstitute, S-10401Stockholm, Sweden Received forpublication4October1974
Nuclei from polyoma-infected 3T6 fibroblasts elongate in vitro the progeny
strandsofthe replicative intermediatesofpolyoma DNA. Whenhigh
concentra-tions of such nuclei were incubated, short DNA fragments were formed and
subsequently addedontogrowingprogenystrands. When nucleiwererepeatedly
washed with buffercontainingdetergent and then incubated at low
concentra-tions, DNA synthesis was decreased. In particular, the joining process was
reduced, resulting inanaccumulation of short DNA fragments.Allaspectsofthe
syntheticcapacity ofthe nucleiwererestoredby additionof cytoplasmicextract.
Additions of purified enzymes (polynucleotide ligase from calf thymus or
Escherichia coli togetherwith E. coli DNA polymerase I) increased thejoining
function of the nuclei. The system can be used for the identification of the enzymaticsteps concernedwith polyoma DNA replication.
Wedescribed earlier an in vitro system for the study of polyomaDNAreplication,consisting of
isolated nuclei from 3T6 mouse fibroblasts in-fected withpolyoma virus (15). During
incuba-tion, the nuclei are able tocontinue and
com-plete the elongation of progeny strands which
were initiated in vivo (9). This process was
shown to proceed in a stepwise fashion and to involve the intermediate formation of Okazaki typefragments (11)about 150nucleotides long, which are initiated at their 5' end by short
stretches of ribonucleotides (2, 8).
Experiments by Mueller and co-workers dem-onstrated that the in vitrosynthesis of chromo-somal DNA in nuclei from HeLa cells could be stimulated by cytoplasmic factors (4, 10). Here
we explore the possibility of influencing the synthesis of aviral DNA inisolated cell nuclei
by addition ofcytoplasmic extracts orpurified enzymes. With this system it is possible to
demonstrate effectsondiscretesteps
participat-ingin the overall process, and our experiments shouldprovide a complementation system
sim-ilar to what has been described for
microorga-nisms (7).
MATERIALS AND METHODS
[3H ]thymidine ([3H]TdR; 6.7 Ci/mmol) and [a-32PJdGTPwereobtainedfromNewEngland Nuclear Corp.;
["4C]dATP
was purchased from Amersham. Escherichia coli DNA polymerase I (fraction VII. Jovin et al. [6]) was a giftfrom Lambert Skoog; E. colipolynucleotide ligasewaspreparedessentially by the method of Olivera and Lehman (12) with some minor modifications (B. Heyden, Dipl. Arbeit,Uni-versity ofTiibingen, Tiubingen, Germany, 1970) was
agift from Heinz Schaller. Calf thymus DNA ligase I, purified 1,000-fold by the method of Soderhall and Lindahl(13) and then furtherpurifiedby chroma-tography onphosphocellulose, was a gift from Stefan Soderhall. Most of the methodology including the handling of cells and virus has been described pre-viously (15).
Invivolabeling of DNA.Immediately before the preparation ofnuclei, cellswere incubated for6to7 minwith [3H]TdR (1.0
MM)
intheculture medium. Labeling wasterminated byremoval of the medium and addition of5 ml ofice-cold Tris-buffered saline (15). Under theseconditions,about 90% ofthe radio-activity was recovered in the progeny strands of replicativeintermediates.Preparationand incubation of nuclei. The prepa-ration ofnuclei from fivepetri dishes(15 cm), each containing approximately 107 cells, is described. All manipulations were performed at 4C. (i) Normal nuclei: the Tris-buffered salinewasremoved,and the cellmonolayers were washedtwotimes with5 ml of isotonic
N-2-hydroxyethyl-piperazine-N'-2'-ethane-sulfonic acid (HEPES) (15). Cellswere
scraped
and diluted into one 15-ml centrifugetube with 5ml of isotonic HEPES, and Nonidet P-40 was added to afinal concentration of 0.5%. The suspension was
vortexedat2-minintervals,firstfor 90sand then five
times for 30 s on a Lab Line Super Mixer. Isotonic HEPES (10 ml) wasadded, and the suspension was
centrifugedfor10minat800x g. The
pellet
contain-ing the nuclei (about 0.5 ml) wasresuspended
with1.5 ml of isotonic HEPES and used either
directly
forincubationorstoredat -70C after
quick
freezingin an ethanol-dry ice-bath. (ii) Depleted nuclei:
fro-zensuspensionsofnormal nucleipreparedfromabout
5 x 107cells werethawed and diluted into 20 ml of 259
on November 10, 2019 by guest
http://jvi.asm.org/
isotonic HEPES. Nonidet P-40was addedtoa final
concentration of 0.5%, and the suspension was vor-texed five times for 30 s as described above. After
centrifugation for 10 min at 800 x g, the nuclear
pelletwasresuspended in 1.5 ml of isotonic HEPES. The depleted nuclei were then frozen and storedat
-70C.
Incubation conditions.The conditions previously
described (15)weremodified asfollows:
[a-"32P]deox-ynucleoside triphosphate (4,000 to 8,000 counts per minperpmole)or ["4C ]dATP (700countsperminper
pmole) were used at saturating concentrations (10
MM), and ribonucleoside triphosphates were always includedataconcentration of 60MM (exceptforATP,
whichwaspresent at2mM). Further variations of the incubation conditionsaregivenforindividual experi-ments.All incubationswereat25C. Selective
extrac-tion ofviralDNAaccordingtoHirt(5)wasperformed
asdescribed previously.
Preparations of cytoplasmicextracts.All
manip-ulationswereperformedat4C. Between 26 and 28 h afterinfection,the cellmonolayerswerewashed twice
with5ml ofice-cold Tris-buffered salineperdish and
then covered with 5 ml of isotonic HEPES. After 5 min, buffer was removed and plates were left in a
vertical position foranother5mintodrain. The cells
were scraped off and treated five times at 2-min
intervals with three strokes in aloose-fitting Dounce
homogenizer. This highly concentrated lysate was
centrifugedfor 40 minat25,000 x g.NaClwasadded
to the supernatant to give a final concentration of
0.1 M. Thesupernatantwasdialyzed for24h against
isotonic HEPES supplemented with0.1 MNaCland
was then frozen at -20 C. About 0.5 to 1.0 ml of
cytoplasmicextractwasobtained from5x 107cells.
Determination of DNA synthesis. The in vivo
['H]TdR prelabel described abovewasusedto
stan-dardize the amount of nuclei used in the in vitro
incubations. For each preparation of nuclei, the
specific activity (3H counts permin perAgofDNA)
wasdetermined first. From this value and from the
amount of 'H label in a given experiment we could thencalculate theamountofnuclei used. This value is expressed as Hg ofDNA per 100
Al
ofincubation mixture. The in vitro DNA synthesisiscorrelatedto the in vivoprelabel and expressedasthe ratio of "2Por4Clabelto 'Hlabel.
Alkalinesucrosegradientcentrifugation. These were all run in a Beckman SW 56 rotor for 3 h at
55,000 rpm at 4C. An internal marker (16S) of labeled, linear polyoma DNA obtained by cleavageof
form Iwith E. coli R, restrictionenzyme(1)was usu-allyincluded.
RESULTS
Stimulation of DNA synthesis of depleted nuclei by ribonucleoside triphosphates. Al-though RNA synthesis appears to be required for the elongation of polyoma DNA in isolated nuclei (8), the addition of ribonucleoside
tri-phosphatesother than ATPto incubation
mix-tures stimulated incorporation of radioactive deoxynucleoside triphosphates into DNA only marginally and under special conditions (8). Probably the normal isolated nuclei retain
con-siderablepools of ribonucleosidetriphosphates. When ribonucleoside triphosphates other than ATP were added to depleted nuclei, both the
rate and extent ofDNA synthesis were
stimu-lated (Fig. 1). Incubation mixtures in all further experiments contained ribonucleoside triphos-phates at saturating concentrations unless stated otherwise.
Dependence of DNA synthesis on
concen-tration of nuclei and stimulation by addition ofcytoplasmicextract.Inthefollowing
experi-ment, the concentration of nuclei (normal or
depleted) in the incubation mixturewasvaried,
and the amount of DNA synthesis was
deter-mined. As described above, the replicative in-termediates in nucleiwere prelabeled with
trit-ium, while the in vitro incorporation utilized [a-32P]dGTP. The 32P to 3H ratio is thus a measure of in vitro DNA synthesis. This ratio was drastically decreased at low concentrations of nuclei. The effect was apparent with both types of nuclei, even though it was somewhat morepronounced with the depleted nuclei (Fig.
2). Addition ofhigher concentrations of dithio-threitol (up to 10 mM) had no effect. The
results suggest that at low concentrations of nuclei the activity of the system is limited by
somefactorswhicharelost from the nuclei. The
effectwasless pronounced during short
incuba-tion times (5 min), indicating that factorswere
eluted from nuclei during DNA synthesis in
1.0
I
-.
t
0.5
U)-10 20 30
MINUTES
FIG. 1. Effect of ribonucleoside triphosphates on
DNAsynthesis of depleted nuclei. Depleted nuclei (27
ug ofDNA) were incubatedfor differenttimes with and without 60 MM each of CTP, GTP, and UTP.
Standard conditions weremodified asfollows:
deox-yribonucleoside triphosphates were used at 10 MM,
ATP at 0.4 mM final concentrations. Symbols: 0,
with 60 MM each of CTP, GTP and UTP; and *, without CTP, GTP, and UTP.
260
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.503.293.438.438.563.2]1.5
20 40 60 s0
CONCENTRATION OF NUCLEI(jigDNA/loojA FIG. 2. Dependence of viral DNA synthesis on
concentration of nuclei. The indicated amounts of
nuclei (micrograms ofDNA/100-microliter volumes) wereincubated under standardconditions for30min. Symbols: 0,depletednuclei; and*,normal nuclei. vitro as well as during preparation of nuclei.
Thisinterpretationisalsofavouredbyan
exper-iment (data not shown) in which depleted
nuclei were first incubated for 4 min under
standard conditions but without labeled
deox-ynucleoside triphosphates. After pelleting, dif-ferent amounts of nuclei were resuspended in
standard incubation mixtures containing
"2P-labeled dGTP and DNA synthesis was
mea-sured for 25 min. In such experiments, DNA synthesis (32P to 3H) was low and showed less
dependence ontheconcentration of nuclei.
These results suggested that DNA synthesis of diluted nuclei could be restored by the addition of factors lost from them, and that such factors might be present in cytoplasmic extracts.This approachassumesthat nucleican
exchange proteins with the incubation medium. The effect of cytoplasmic extracts on in vitro
DNA synthesis is shown in Fig. 3. Two
concen-trations of nuclei were incubated for 30 min
with increasing amounts of extract. In both
cases DNA synthesis was stimulated, but the
degree of stimulationwasmuch larger withthe
lower concentration of nuceli. The extent of DNA synthesis obtained with the largest amount of extract was thesame, however, and
wasidenticaltothatobservedwithhigh
concen-trations of nuclei intheabsence of extract(Fig. 2). The amount of extract required to obtain half maximal stimulationwasabout threetimes
larger with the lower concentration of nuclei. Cytoplasmic extracts used in this and all fur-ther experiments describedwerepreparedfrom polyoma-infected 3T6 cells, but similar effects
wereobtained withextractsfromgrowing, unin-fectedcells. Extracts heatedfor5minat 100 C lost their stimulating capacity. Furthermore,
the stimulating capacitycould be fractionated byammonium sulfateprecipitationand precip-itated between 30 and 60% saturation. Both of
these results suggest that the stimulating fac-tors areproteins.
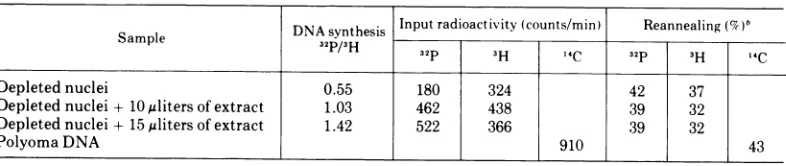
The polyoma-specific nature of the DNA synthesizedindepleted nuclei in the presence of
extracts was
demonstrated
by reannealingex-periments (Table 1). Equal relative amountsof in vitro 32P-labeled DNA were annealed to
polyoma
DNAindependent
of whether DNAwas synthesized in the absence of extracts
(42%)
or in thepresenceof extracts(39%).Sim-ilar results were obtained with thein vivo
3H-labeledDNAwhichhybridizedto 37% and32%, respectively. Inaparallel experiment, authentic polyoma DNA reassociated to about the same extent
(43%).
Sedimentation properties of in vitro syn-thesized DNA.In thefollowingsections we will show that the effect of extract also is of a
qualitativenatureand inparticular reflectsthe
ability
ofthe nuclei tojoinsmall DNApiecestolonger chains. In Fig. 4, two alkaline sucrose
gradient centrifugationsofthe products
synthe-sized by normal and depleted nuclei, respec-tively, arecompared.After a30-min incubationwithnormalnuclei, mostof theincorporated in vitro label wasfoundinlong chainssedimenting
close to 16S while with depleted nuclei about
5 10 15 20
[image:3.503.52.213.52.167.2]CYTOPLASMIC EXTRACT (i's)
FIG. 3. Effect of cytoplasmic protein extract on
DNA synthesis of depleted nuclei. Prelabeled, de-pleted nucleiwereincubatedat twoconcentrationsof nuclei withincreasingamountsof cytoplasmic protein
extracts from polyoma-infected cells (microliters). Incubations were carried out under standard condi-tionsfor30min.Inparallelincubations,DNA synthe-sis of indicated amounts of extracts per se was
measured. DNA synthesis (32P/3H) of nuclei was correctedfor theincorporationvaluesofextractalone. Thelargest correction (20 uliters of extract) amounted
to15%. Theproteinconcentrationoftheextract was
about 10 mg/ml. Symbols: 0, 27 Ag of DNA/100
Alitersfinalvolume;and*, 13 lAgofDNA/100
Mliters
final volume.
,I
Ic
,'
261
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.503.249.444.336.521.2]TABLE 1. ReannealingtopolyomaDNA bound tofiltersa
DNA synthesis Inputradioactivity
(counts/min)
Reannealing(%)bSample 32P/3H 32p 3 14C 32P 3H "C
Depleted nuclei 0.55 180 324 42 37 Depleted nuclei + 10Mlitersofextract 1.03 462 438 39 32 Depleted nuclei + 15 ulitersofextract 1.42 522 366 39 32
PolyomaDNA 910 43
aThe conditionsforreannealing were the same aspreviously described (15). 'Filters contained 0.5ggofpolyomaDNA.
half of the radioactivity appeared as 4 to 5S
fragments. These short fragments were earlier shown tobe precursors of
long
strands (8). Theresults (Fig. 4) suggestthat depleted nuclei are
lacking inthe capacitytojoin thefragments. A second point concerns the amount of 3H
sedi-menting in the position ofform I DNA (53S). When the values for the fraction of prelabeled DNAsedimenting at53Swere corrected for the amountofprelabelinform IDNA
by
theinvivo pulse (12%), an additional 12% of the in vivolabeled viral DNA was transformed to form I DNA after a 30-min incubation. The
corre-sponding valuefordepleted nucleiwasonly2%.
Additionofcytoplasmicextracts restored the
capacity of depleted nuclei to join 4 to 5S
fragments. Figure5shows theeffect of two con-centrations of extract on the sedimentation
profile in alkaline sucrose gradients. In both cases a considerable stimulation of total DNA
synthesis had occurred, and is expressed byan
increased amount of in vitro label
(321p)
appear-ing as long single-stranded chains sedimentingclose to 16S and as form I DNA (53S). In the
absence of nuclei, the extract alone showed a
limited capacity to synthesize DNA (Fig. 5B), most ofwhich sedimented asshortchains. This
activity probably explains the shoulder at the 4to 5Sposition inFig. 5Cand D. Theaddition
of extract also considerably increased the amount of prelabel in form I DNA, which was
stimulated from 2%T (Fig. 5A) to 11% (C) and 19%(D).
Effects of some purified enzymes on DNA
synthesisofdepletednuclei. It appearedtobe ofinteresttoinvestigatewhetherthe additionof certain purified enzymes, either alone or in
combination, could mimic the effect of crude
cytoplasmic extracts. Depleted nuclei clearly are deficient in the ability to join short DNA
chains. Since this deficiency might in turn be the cause of the general inhibition of strand
elongation,
we first investigated whetherpoly-nucleotide ligase might substitute forthe cyto-plasmic extract. Addition of highly purified
ligasefrom either calfthymusorE. coli did not
increase the total amount of in vitro label
incorporated
intopolyoma
DNA. However,there was a
slight
effect on the distribution ofisotope
inalkalinesucrosegradients
involvingashift of some ofthe material
present
as shortDNA
fragments
tolonger
chains(Fig.
6B andC).
Addition of moreligase
did not increase theseeffects.However,
addition of DNApolym-erase I from E. coli
together
with ligase in-creased this shift(Fig.
7D and E). There wasnowalso a
slight
stimulation of total amount of DNAsynthesis
(up
to 125% of the control) but the amount of stimulation observed withcyto-plasmic
extracts(Fig.
5) was definitely notat-tained.
Similarly,
there was only aslight stim-ulation of the amount ofisotope
appearing in form I. Our results demonstrate that theproc-essof
polyoma
DNAsynthesis
in isolated nucleicanbeinfluenced
by purified
enzymes and thatdepletion
ofligase
and DNA polymerase areinvolved in the loss ofsynthetic capacity ofthe nuclei. Since E. coli DNA polymerase I has both
polymerizing
and exonucleolyticactivities(14)
bothmight
be involved in the observedstimulatory
effect. However, eventhough
these enzymes were added inlarge
excess,they
couldby
no means completely restore theac-tivity
ofthenuclei, suggesting thatcytoplasmic
extracts
supply
otherfactorsaswell.The stimulatory effects described above
sug-gestthat
purified
enzymes andeffectorspresent incytoplasmic
extracts can penetrate into iso-lated nuclei and participate in thereplication
process inside the nuclei. Alternatively, DNA and enzymes maydiffuse out from the
nuclei,
and thestimulationsobservedinthepresenceof extractsmaythus represent processesoccurring outside thenuclei. Todistinguishbetweenthesetwo alternatives the following experiment was done. Depleted nuclei were incubated either alone or in the presence of an excess of DNA
ligase
from calfthymus orcytoplasmicextract. At the end of theincubation, DNA synthesizedoutsidethe nucleiwasseparated from theDNA
synthesizedinsidebycentrifugation. About10% of the total in vitro incorporated
radioactivity
OTTO ANDREICHARD J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.503.74.467.87.170.2]100
1I
U
1 1000
II
I
-1500
0I
I_ Io
Il
I
I
I
-t500
20
40
[image:5.503.105.388.67.477.2]FRACTION NUMBER FROM BOTTOM
FIG. 4. Characterizationof viralDNA synthesized in normal and depleted nuclei.Prelabelednuclei (36 ,g of
DNAof normal nucleiand27,g of DNA of depletednuclei) wereincubated under standard conditions with
14C-labeled dATPfor 30 min. Portions (200 Mliters) of Hirtsupernatant fluidwere centrifuged through an
alkalinesucrosegradientasdescribedintext.Panel A, normalnuclei; panel B, depleted nuclei. Symbols:0,
3H;and*, "C.
was found outside the nuclei when
they
wereincubated aloneorinthepresence ofligase.The amount increased to 27% in the presence of extract. Figure 7shows the sedimentation pro-files in alkaline sucrose gradients of the DNA
synthesized in the nuclei. It is clear that sim-ilar effects to those observed in Fig. 5 and 6 were found. A comparison of Fig. 7C and 5D suggests that the radioactive material present as a shoulder in the position of4 to 5S DNA fragments in Fig. 5D was outside the nuclei. In conclusion, the addition of cytoplasmic
extract indeed stimulated the overall
elonga-tionprocess ofDNAreplication insidedepleted
nuclei.
DISCUSSION
The capacity ofnuclei to complete the elon-gation of progeny strands during incubation
depended on the concentration of nuclei. The relative amount ofpolyomaDNA
synthesis
wasgreatly decreased when low concentrations of nuclei were used. This effect was more pro-nounced with nuclei which had been
prepared
263
on November 10, 2019 by guest
http://jvi.asm.org/
264 OTTO AND REICHARD
A
B200
-0
0
C
100
,*
0~~~~~~~~~I~~~~~~
*0
0~~~~~~~~
600~~~~~~~~
*16 S
20
40
0
20
40
FRACTION
NUMBER FROM BOTTOM
FIG. 5. Effect of cytoplasmic protein extracts onDNA synthesis of depleted nuclei (product analysis). Prelabeled,depletednuclei(13,ugof DNA)wereincubated eitherintheabsence(A)orpresenceof20Mliters(C)
or50 Mliters (D) of cytoplasmic proteinextractfrompolyoma-infected3T6 cells. A50-gliteramountofextract was also incubated without nuclei (B). All incubations were for 30 min under standard conditions with [32P]-labeleddGTP. The datawereobtainedfromidenticalportions (200,litersofHirtsupematantfluid with
5 .liters of "'C-labeled linearpolyomaDNA as a marker)run inparallelonalkaline sucrosegradients (see
text).Thearrowsgivetheposition ofthemarker(16S). Symbols:0, 3H;ando, S2p.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.503.72.451.83.525.2]I,
X
I
20
400
20
40
FRkCTION NR
FROM WTT
FIG. 6. Effect of somepurified enzymes on DNA synthesis of depleted nuclei. Prelabeled, depleted nuclei (13 ggof DNA)wereincubated without added enzymes(A);with E. colipolynucleotide ligase (B)orcalfthymus DNA ligase I (C);orwith E. coli DNA polymerase I in the presence of either calf thymus DNA ligase I (D) or E. colipolynucleotideligase(E).Incubationswerefor30min.Thestandard incubation mixture with [82P]-labeled dGTPwassupplemented with NAD+ (10-I M final). Portions (200 /uliters) ofHirtsupernatant fluid together with 5 /Lliters of "4C-labeled linearpolyoma DNA as a marker were centrifuged through alkaline sucrose gradients (see text). Thearrowsgive the position of the marker (16 S). Symbols: 0, 3H;and o, 32P.
265
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.503.61.428.36.604.2]I
I
m
B16S
u B~~~~~~~
50 * 50' * -'50
20 40 20 40
FRACTION NUMBER FROM BOTTOM
FIG. 7. Effect of ligaseorcytoplasmic proteinextractonDNAsynthesis inside depleted nuclei. Prelabeled depleted nuclei (13 ugofDNA) wereincubated under standard conditions with
[32P]-labeled
dGTPfor30min: (A)alone, (B) with DNA ligaseIfromcalf thymus, or(C) with50,liters
of cytoplasmic protein extract from polyoma-infected mouse fibroblast 3T6 cells. DNA synthesis was stopped by adding 100Mlliters
ofice-cold isotonic HEPEStotheincubation mixture.Aftercentrifugation for10minat 4Cat 800 xg, the nuclearpelletwassuspended in 100gliters of isotonic HEPES and viral DNAfromboth the supernatant and the nuclei
sus-pensionwasextracted by the Hirt procedure (5). Identicalportions (200Ilitersof Hirt supernatant fluid from the nucleifraction and5/.liters of'4C-labeledlinearpolyomaDNAas amarker) wereruninparallel in alkaline
sucrosegradients (see text). Thearrowsgivetheposition ofthemarker(16S). Symbols:0, 3H;and ,S2P.
by repeated washing with buffers containing
Nonidet P-40 and which were therefore less contaminated with cytoplasm. The decrease
wasmainlyapparent inthe capacityofnucleito sustain DNA synthesis and did not so much affect initial rates. From our data, it seems likely that part of the enzymes are washed out during preparation ofnuclei and part leave the nuclei during DNA synthesis. Nevertheless,
depleted nuclei incubated at a high concentra-tion retained their capacity to continue the
elongation process.
Earlier results showed that polyoma DNA synthesisinnucleiinitially involved the forma-tion ofshort DNAfragments, 100 to 150 nucleo-tides long, and that these fragments subse-quently were joined to form longer progeny strands (8). With depleted nuclei we now find anaccumulation ofshortfragments also during
prolonged incubation indicating that some of the components being removed are involved in thejoining process.
The synthetic capacity can be fully restored
by addition of cytoplasmic extracts, and the active factors in the extracts appear to be proteins. Restored DNA synthesis leads to the formation oflong progeny strands. The extracts thussupply enzymes required forthejoining of the short DNA fragments as well as for other steps of the elongation process. Extracts also increase the rate oftransformation ofreplicative intermediate-DNA into mature viral DNA (form I DNA), but it is uncertain whether extractsalso supply enzymes for the segregation of completely replicated DNA molecules. For-mation of formI was,however, never as efficient as was reported for crude lysates of polyoma-infected 3T3 cells (3).
266
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.503.123.401.59.360.2]About equal activities were found with
ex-tracts from infectedoruninfected growing 3T6
cells. This result indicates that at least all enzymatic activities which can replace
func-tions in the elongation process are of cellular
rather than of viralorigin.
Polynucleotide ligase is an enzyme which
would be expectedtoparticipate inthe joining
process. Neither purified calf thymus ligasenor
E. coli ligase stimulated the overall capacity of the nuclearsystemtosynthesize polyoma DNA. However, ashift of isotope from short fragments to long progeny strands was observed. This
result indicates that ligase activity probably
wasoneof thedepleted nuclear factors, but that
the cytoplasmic extract clearly supplied other additional missing fractions. E. coli DNA
po-lymerase I, together with ligase, gave only a
small stimulation of the overall synthesis, but
increased the joining to a larger extent than ligasealone.Both the polymerizing activity and the exonuclease activity of this enzyme might
be responsible for the observed effect.
Theexperiments involving addition of highly purified enzymes indicate that proteins with
molecular weights of about 100,000 (6, 13) can
influence a synthetic processwhich apparently
occurs inside nuclei. It seems reasonable to
assumethat thisoccursafterpenetrationofthe
enzyme intothe nuclei.
The nuclearsystemwas originally developed
with the hope that it would be useful in the dissection of intermediate steps of polyoma DNA synthesis. The effects described in this
paperopen upexperimentalpossibilitiesinthat
direction. The demonstration of stimulatory effects on a depleted system must be inter-preted with great caution since clearly neither E. coli ligase norE. coli DNApolymeraseI per se participate in polyoma DNA synthesis. The
demonstrated permeability for proteins of the nuclear in vitro system appears promising,
however,and itnowseemsexperimentally feasi-ble to identify enzymes participating in the intermediate steps ofDNA synthesis either by fractionation ofcytoplasmic extracts orby
im-munological techniques.
ACKNOWLEDGMENTS We thankGunilla S6dermanforexpertassistance. The studywassupported byapostdoctoral fellowshipfrom
theDeutscheForschungsgemeinschafttoB.0.andbygrants
from the Swedish Cancer Society and Magnus Bergvalls StiftelsetoP.R.
LITERATURE CITED
1. Crawford, L. V., C. Syrett, and A. Wilde. 1973. The replication of polyoma DNA. J. Gen. Virol. 21:515-521. 2. Eliasson,R., R. Martin, and P.Reichard. 1974.
Charac-terization of the RNA initiating the discontinuous synthesis of polyoma DNA. Biochem. Biophys. Res. Commun.59:307-313.
3. Francke, B., and T. Hunter.1974.InvitropolyomaDNA synthesis: discontinuous chain growth. J. Mol. Biol. 83:99-121.
4. Hershey, H. V., J. F. Stieber,andG. C. Mueller. 1973. DNAsynthesis in isolated HeLa nuclei. Eur. J. Bio-chem.34:383-394.
5. Hirt, B.1967.Selectiveextractionofpolyoma DNA from infectedmousecell cultures.J. Mol.Biol. 26:365-369. 6. Jovin, T. M., P.T. Englund, and L. L. Bertsch. 1969. Enzymatic synthesis of DNA. XXVI. Physical and chemicalstudies ofahomogenous DNApolymerase.J.
Biol. Chem. 244:2996-3008.
7. Klein, A., and F. Bonhoeffer. 1972. DNA replication. Annu.Rev. Biochem.41:301-332.
8.Magnusson, G., V. Pigiet, E.-L. Winnacker,R.Abrams, and P. Reichard. 1973. RNA linked shortDNA
frag-mentsduring polyoma replication. Proc. Nat. Acad. Sci.U.S.A.70:412-415.
9. Magnusson, G., E.-L. Winnacker, R. Eliasson, and P. Reichard. 1972. Replication ofpolyoma DNA in iso-lated nuclei. II. Evidenceforsemiconservative replica-tion.J. Mol. Biol.72:539-552.
10. Mueller, G. C. 1969. Biochemicaleventsin theanimal cellcycles. Fed. Proc. 28:1780-1789.
11. Okazaki, R., T. Okazaki, K. Sakabe, K. Sugimoto, R. Kainuma, A. Sugino, and N.Ivatsuki. 1968. In vivo mechanismofDNA chain growth. Cold Spring Harbor Symp. Quant. Biol.23:129-143.
12. Olivera, B. M., and I. R. Lehman. 1967. Linkage of polynucleotides through phosphodiester bonds by an
enzymefrom Escherichia coli. Proc. Nat. Acad. Sci.
U.S.A.57:1426-1433.
13. Soderhiill, S., and T. Lindahl. 1973. Two DNA ligase activities from calfthymus. Biochem. Biophys. Res. Commun.53:910-916.
14. Westergaard, O., D. Brutlag, and A. Kornberg. 1972. Initiation ofDNA synthesis. IV. Incorporation ofthe RNAprimerintothephage replicativeform.J. Biol. Chem.248:1361-1364.
15. Winnacker, E.-L., G. Magnusson, and P.Reichard.1972. Replication of polyoma DNA in isolated nuclei. I. Characterization ofthesystemfrommousefibroblast
3T6cells.J. Mol. Biol.72:523-537.
15,