0022-538X/79/02-0726/09$02.00/0
Interruption-Deficient
Mutants
of
Bacteriophage
T5
II.
Properties of
a
Mutant
Lacking
a
Specific
Interruptiont
STEPHEN G. ROGERS,: NANCY V. HAMLETT,§ANDMARC RHOADES*
BiologyDepartment, The Johns HopkinsUniversity, Baltimore,Maryland21218
Receivedfor publication15August1978
An examination wasmade of the properties ofT5HA4, a mutantof
bacterio-phageT5that lacks thesingle-chaininterruption thatoccurs at7.9% fromthe left
endof the genome. The DNAs of T5HA4 andthe wild type werecomparedby
electrophoresis in agarose gels of both single-stranded fragments produced by
denaturation and duplex fragments generated by sequential treatment with
exonuclease III and SI nuclease. These studies demonstrated that T5HA4 also
lacksan interruptionthat occurs at99.6% in wild-type DNA. The interruptions
at 7.9 and 99.6% thereforeoccur within the 8.3% of T5 DNAthat isterminally
repetitious. Evidence on the location of other interruptions within the terminal
repetition was also obtained. Analysis of T5HA4 DNA witha restriction
endo-nuclease indicated thatthe interruption deficiency is not due to a deletion or
addition mutation. Theinjectionof T5HA4DNAintoahost bacteriumwasfound
tooccur, as with the wild type,inatwo-step manner.Theinterruptionat7.9% is
thereforenotrequired for stoppingDNAtransferafter the initial 8%segmenthas
beeninjected.
The genome ofbacteriophage T5 contains a
number of site-specific single-chain
interrup-tions within one strand of its linear
double-stranded DNA. As described in the preceding
paper (18), mutants of T5 have been isolated
that eitherlackspecificinterruptionsorare
de-ficient forinterruptionsthroughout thegenome.
The latter class of mutations appears todefine
twogenes that participate in the formation of interruptions in T5 DNA. In contrast, the single-site mutants appearto haveundergone
altera-tions of therecognitionsequencesthat mark the
positions of the interruptions. Mutants have
been obtainedthat lack each of the four
princi-palinterruptions, whichoccur at7.9, 18.5, 32.6,
and64.8% from the left end ofT5DNA,aswell
as onesecondaryinterruption,locatedat45.3%.
The existence ofsingle-site
interruption-defi-cient T5 mutants permits direct testing of the
functionalsignificance ofspecific interruptions.
Thepresentreportisconcerned with the
prop-erties ofa mutantthatlacks theinterruptionat
7.9%. This site is of interest for several reasons. It occurs close to the boundary between the
repeated andnonrepeated segmentsatthe left
end of the genome. The terminal repetition in
t Contributionno.973 from the DepartmentofBiology, TheJohnsHopkinsUniversity,Baltimore,Md.
tPresentaddress:DepartmentofMicrobiology,TheJohns HopkinsUniversity School of Medicine,Baltimore,MD 21205. §Presentaddress:Department ofBiological Sciences, Tow-sonStateUniversity, Baltimore,MD 21204.
T5 DNA has been estimated tocomprise 8.3%
ofintact DNA (16, 17). The 7.9% interruption
also occurs close tothe boundary between the
first- and second-step transfer segments of T5
DNA. T5 is unique in that injection of its
ge-nomeintoahostbacteriumoccursbyatwo-step
process. Initially, an 8.0 to 8.5% segment, the
first-step transfer (FST) DNA, is injected (14).
Expression of at least two FST genes is then
required before the remaining 92% of thegenome
istransferred (11). The FSTsegment appears to
be at the left end of the T5 genome (10, 20).
Because of itslocation, the interruptionat 7.9%
is therefore a possible candidate for the signal
that terminatesthe firststageof DNA transfer.
The results obtained in this study have
con-firmed that the 7.9%interruptionlies within the
terminal repetition in T5 DNA. Mutants that
lack the 7.9%interruption are alsodeficientfor
an analogous site at 99.6%. In addition, these
studies haveprovided evidence onthe location
of various secondary interruptions that occur within the terminalrepetition.Theinterruption
at7.9%,however,doesnotappear toplayarole
in DNAtransfer.In theabsence of this
interrup-tion, injection occurs in the normal, two-step
manner.
MATERIALS AND METHODS
Bacteria and bacteriophage. The bacteria and
wild-type T5 phage used in this study have been
describedpreviously (18). T5st(102) isaviable,
heat-726
on November 10, 2019 by guest
http://jvi.asm.org/
stable deletion mutant that lacks 10.2% of T5st(+)
DNA (22). T5HA4,which lacks the 7.9%interruption,
was originallyisolated in T5st(102) as described in the
preceding paper (18). The original isolate wasable to
grow on Escherichia coli B23 and B40sul, but was
not able to grow on E.coli F. The inability to grow on
E. coli F was eliminated by backcrossing against
T5st(+). The recombinant obtained, however,
ad-sorbed poorly on E. coli F. A fast-adsorbing, HA4
recombinant was obtained by performing a second
backcrossagainst T5st(+). This stock was usedfor the
experimentsreported here.
Analysis of T5HA4 DNA. The procedures for electrophoretic analysis of T5 DNA have been
de-scribedpreviously. Single-strandedDNAwasanalyzed
in 5-mm-thick horizontal agarose slab gels (16, 19).
Double-stranded DNA underwent electrophoresis
similarlyorincylindrical vertical gels (16). Treatment
of T5 DNA with Aexonuclease, HpaIrestriction
en-donuclease, orexonuclease III and SI nuclease was
carried out as described (5, 16).
Measurement of two-step transfer of T5 DNA.
Theprocedure used to measure two-steptransfer of
T5 DNA was that of Lanni (11), except that shear
treatmentof infected cellswascarried outfor6minat
"HIGH"speed in a VirTis 45 homogenizer.32P-labeled
phage were purified onCsCl step gradients.
Isolation of intracellular DNA from FST
com-plexes.FST complexes (bacteria to which T5 phage
have been adsorbed and haveinjectedonly the FST
DNA), prepared and sheared as above, were
centri-fuged, washed, and resuspended in0.1MTris-0.1 M
NaCl-0.1 MEDTA, pH8.0. DNAwasextracted from
thecomplexes with sodium dodecyl sulfate (SDS) and
Pronase (2) and was dialyzed against 10 mM
Tris-hydrochloride-0.1 mM EDTA (pH 8.0). In one
exper-imentanattemptwasmadetoseparate theFST DNA
from the host DNA by using the
lysozyme-EDTA-Triton X-100 procedure used for plasmid DNA (9).
The FST DNA, however,wasfound associated with
the membrane-host DNA fraction. The FST DNA
wasreleased from thiscomplex by theSDS/Pronase
procedure used for intact cells (2) andwasseparated
from the bulk of the host DNA by sedimentation
through6 to20%neutralsucrosegradients.Thepooled
fractions were concentrated by pressure dialysis
against 10 mM Tris-hydrochloride-0.1 mM EDTA
(pH 8.0) beforeelectrophoretic analysis.
RESULTS
Thesearch forinterruption-deficientmutants
of T5describedintheprecedingpaperresulted
in theisolation ofseven mutantsthatlacked the
single-chaininterruptionat7.9% (18). These
mu-tants were identified because, after
denatura-tion, their DNAs lacked the single-chain
frag-mentsthat haveoneendat7.9%. These include
fragments 18, 21, and 22 (Fig. la and b). The
first mutant isolated in this category, T5HA4,
waschosen for furtherstudy.The HA4 mutation
hasalreadybeen shown to be neither dominant
norrecessivetothewild type(18). Phage
carry-ingthismutation are viableand have thesame
(a) 5
(b)
(C )
08 24 3.541 79 165 32 6
21 Is is3
39 22 i 4A 7 IA
29 26
27 29
A
391 .Ol
33 C
29 D
T
[image:2.505.257.452.77.204.2]27 i E so
FIG. 1. Maps of single-chain interruptions and
fragments attheleft endof wild-typeT5 and T5HA4
DNAs.(a) Positionsof interruptionsinT5st(+) DNA,
expressedas apercentage of wild-type lengthfrom
theleft endofthe molecule. Thestrandpolaritiesand
interruptionsare shown asdetermined inreference
16. (b)Locationsofsingle-chain fragmentsattheleft
endof T5st(+) DNA. With theexceptionof fragment
30, whichoccurs between 0and3.3%o, allpreviously
mapped (16)fragments areshown. Thest(102)
dele-tion occurs between the18.5and 32.6ointerruptions.
(c) Locationsofsingle-chainfragmentsattheleftend
of T5HA4 DNA. The positions ofthe left ends of
fragments AthroughEwerecalculatedby
subtract-ing their molecularweights, giveninFig. 3, fromthe
molecularweightofthe intervalfrom0to 18.5%, as
determinedelectrophoretically (16).There isasmall
discrepancy between this value, 18.0%o of T5st(+)
DNA, and the valueof18.5%determined byelectron
microscopy(21).
averageburst sizeasthewildtype.
Analysis of single-stranded fragments of
T5HA4 DNA. As described below and in the
preceding paper (18), loss of fragment 18 and
manyof the small fragmentsatthe left end of
the genomeclearly indicatesthat T5HA4lacks
the interruption at 7.9%. However, since the
terminal repetition inT5 equals8.3% of intact
DNA, thesequencethatcontainsthe7.9%
inter-ruption should also occurclosetothe right end
of the molecule. Evidence has been obtained
that interruptions occur at analogous sites
within thetwocopies of the terminalrepetition
in T5 (16). Thus, if the previous analyses are correct, T5HA4 should lack twointerruptions,
one at 7.9% andone atapproximately99.6%.
The existence ofan interruption at 99.6% in
wild-type T5 DNA, and its absence in T5HA4
DNA, can be demonstrated by comparing the
single-chain fragments of the two DNAs. Only
the wild type should have a short fragment of
approximately450bases that occurs at the right
end of the interrupted strand of T5 DNA. To resolve the smaller single-chain fragments, we
subjecteddenaturedT5st(+) andT5st(+) HA4
DNAs to electrophoresis in 1.4% agarose slab 29,1979
on November 10, 2019 by guest
http://jvi.asm.org/
ROGERS, HAMLETT, AND RHOADES
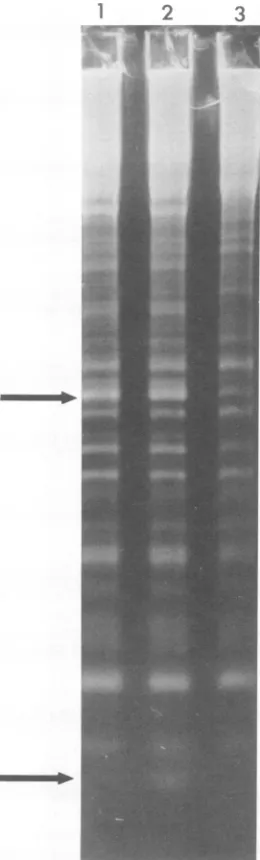
gels. The results (Fig.2) showthat, forfragments
of less than 4% ofan intact strand, the single
chains of thetwoDNAs wereextremely similar.
However, thefastest migratingofthe detectable
1 2 3
TI'
FIG. 2. Electrophoresis ofthe small single-chain fragmentsof T5 DNA. Electrophoresis wascarried
outinahorizontal 1.4%agaroseslabgel.Thelength
ofthegelwas 15 cm. Each slot contained 15[Lg of
alkali-denatured DNA. Slot1,T5st(+)DNAafter1%
digestion with A exonuclease; slot2, T5st(+) DNA;
slot3, T5st(+)HA4DNA. Theupperarrowindicates
theposition, ingels1 and2, offragment28, which
equals 3.8% ofan intact strand. The lower arrow
indicates the 5'-terminalfragmentthat ismissingin
T5HA4DNA.
wild-type fragments appeared to be absent in
HA4 DNA. Since the external 5' end of the
interrupted strandoccurs attherightend ofthe
molecule (16),theterminallocation of this
frag-ment canbedemonstrated by limited digestion
ofwild-type DNA withAexonuclease,anenzyme
that isspecific for external5'termini(4,15). The
result ofdigestion of T5st(+) DNAto 1%
acid-soluble material withAexonuclease is shown in
slot 1 ofFig. 2. The only effect of exonuclease
treatment wasloss of the smallfragmentthat is
missingin T5HA4 DNA. Thisfragment
there-fore occurs atthe external 5' end of the
inter-ruptedstrand. Digestionto0.5% with A
exonu-cleasewas sufficient toyield the resultsshown
inFig.2.Identical electrophoreticpatternswere
also obtained with up to 5%digestion. The
re-sults with higher levels of digestion were not
surprising,since,withtheexception of thesmall
terminalfragment, allof thefragments that
oc-cur within the right terminal repetition also
occur atthe left end of theDNA. The size of the
5'-terminalfragmentidentified in Fig.2hasnot
been accurately measured. Its electrophoretic
mobility, however, is compatible with the
pre-vious estimates of450bases.
Analysis ofduplex fragments of T5HA4
DNA. Loss of theinterruptionsat 7.9and99.6%
should resultinalterations ofmanyofthe
single-chainfragments produced by denaturation of T5
DNA.Asnotedabove, fragments that haveone
terminusateitherinterruption should disappear
while newfragmentsthat spanthesesitesshould
become evident. Analysis of these new
frag-ments, manyofwhichwillhaveoneterminusat
a secondary interruption, can provide further
information on the structure of T5 DNA. In
principle, this informationcanbe obtainedfrom
analyses ofsingle-chain fragments as shownin
Fig. 2. However, the clearest results have been
obtained fromanalysis of the analogous duplex
fragments generated by sequential treatment
with exonucleaseIII ofE. coli and SI nuclease
of Aspergillus oryzae. Limited hydrolysis by
exonuclease III at the 3' side of each
interrup-tion, followed by digestionof thesingle-stranded
regions by SInuclease, resultsincleavageof the
duplexmolecule at the site of eachinterruption
(16). Since theelectrophoretic mobilitiesof
du-plexmolecules arealmostentirely dependenton
molecularweightinagarosegels (24), analysisof
these fragments yields an accurate mapofthe
interruptions.
The results of sequential exonuclease III-SI
nuclease digestion of T5st(+) and T5st(102)
DNA in thepresence and absence of the HA4 mutation are shown in Fig. 3. Electrophoresis
was carried out in 0.5% agarose gels toresolve
thelargerduplexfragments. Thefragmentsare
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.89.219.137.567.2]2
8-9-.-..
10,A
-
B-
11- C-
D - 15-
16-3
-8
-9 -10 -11
-7
-15 -16
4
'-3 -4 -6
-7
-8
-9
-10
-11
-13 -15
-18
FIG. 3. Electrophoresis ofthelargeduplexfragments generated byexonucleaseIII-SI nucleasetreatment
ofT5 DNA.Electrophoresiswascarriedoutincylindrical0.5% agarosegels. Gel1, T5st(102)HA4DNA; gel
2,T5st(102) DNA; gel 3, T5st(+)HA4 DNA; gel 4, T5st(+) DNA. The fragments thatoccurin T5st(+) DNA
arenumbered in orderof decreasingsize(see 16). Electrophoresisin the presenceof referencemoleculeswas
employedtoidentifythe numberedfragmentsin the othersamples. The lettersAthroughEdenotefivenew
fragments that appear inT5st(102)HA4DNA. Theirmolecularweights,expressedas apercentageofT5st(+)
DNA,are17.4%for B, 15.8%for C, 14.6%for D,and14.0%forE.FragmentAcomigrateswithfragment10,
andtherefore equalsapproximately18.4%of T5st(+)DNA.
numbered in order of decreasing size as they
occurinT5st(+) DNA (gel 4). The deletionin
T5st(102) DNA does not remove an
interrup-tion.Therefore,theonlydetectable alterationin
thisDNAisloss oftwofragments,7and 13,that
span the deletion. Fragment 7', the shortened
version offragment7, canbeseeninFig.3 (gel
2). Fragment 13' istoosmalltobeseen inthis
experiment.
The HA4 mutation resultsinloss offragments
7, 18, 21, and 22 of those visible in Fig. 3. A
number ofnewfragmentsare evident.Analysis
ofT5st(102) DNAis most useful inthisregard
since severalof thenewfragmentsareobscured
by fragment 13in T5st(+) DNA. The five new
fragments detectableinT5st(102)HA4DNA are
labeledAthroughE inFig.3 (gel 1). The
exist-ence of fragment A is implied because of the
1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.104.390.72.482.2]730 ROGERS, HAMLETT, AND
increasedintensity ofthebandcontaining
frag-ment 10. Since no interruptions have been
lo-catedbetweentheprincipal interruptionsat 7.9
and 18.5% (16, 21),fragments Athrough E
pre-sumably extend leftward from 18.5% to various
sites near the left end of the molecule. The
positions of thesefragments, basedonthe above
assumption and their molecular weights, are
shown inFig. 1. FragmentA appears toextend
fromtheinterruption of 18.5%totheleft end of
the molecule. The other four fragments
termi-nate at 0.6, 2.2, 3.4, and 4.0%. These positions
agreewell with the locations of 0.8, 2.4,3.5,and
4.1%previously determined for secondary
inter-ruptions within the left terminalrepetition in T5
DNA(16).
Thesmaller fragments generated by
exonucle-ase III-SI nuclease treatment of T5st(+)HA4
DNA were analyzed in 1.0% agarose gels (Fig.
4). T5HA4 DNAwould be expected to lackall
shortfragments that extend leftward from 7.9%
inwild-type DNA. Fragments that extend
right-wardfrom the left end of theDNA tosecondary
interruptions in the terminal repetition should
not be altered by the HA4 mutation. As
indi-cated in Fig. 4, T5st(+)HA4 DNA lacks
frag-ments 21,22, 24, 26,and28andretainsfragments
27, 29, and30. This resultagrees with the
pre-viouslydetermined locations of thesefragments
(Fig. 1).
T5st(+)HA4DNAcontainsanumber ofnew
small duplexfragments (Fig. 4). Theseare
pre-sumablyderivedfrom therightend of the DNA
where loss of the 99.6% interruption will result
in fusion of the short terminal fragment
de-scribed above(Fig. 2)tothefragmentscontained
within the right terminal repetition. Thus,
slightlylonger versions of fragments 22,24, 26,
and 28should bepresentinT5HA4DNA. Bands
that could correspondtomostof the predicted
fragmentswereseen;however,itwasdifficultto
analyze theright end of T5 DNAbyuse ofthe
exonucleaseIII-SI nucleaseprocedure. Since the
3'end of the intact strand occurs atthe right
end of theDNA, fragments that extendto the
right end will be degraded from both sides by exonuclease III. Thiseffect, especiallyif
exonu-cleaseIII actspreferentiallyatduplex3'termini,
precludes accurate measurement of molecular
weights.
Analysis witharestriction endonuclease.
T5HA4 was isolated after hydroxylamine
mu-tagenesis,whichinducespointmutations.
How-ever, the procedure used for mutagenesis in-volved heating in the presence of a chelating
agent (23). Since T5 deletionmutations are
se-lected in asimilarmanner (8), theinterruption
deficiencyof T5HA4 could beduetoloss of the
region containing the 7.9 and 99.6%
interrup-J. VIROL.
18
21
22
24
26
27
28
29
30
FIG. 4. Electrophoresis of the small duplex
frag-mentsgeneratedby exonucleaseIII-SInuclease
treat-mentofT5 DNA.Electrophoresis was carried out in
ahorizontal1.0%oagaroseslab gel. The length of the
gel was 8.5 cm. Slot 1, wild-type T5 DNA; slot 2, T5HA4 DNA.
tions. Althougha deletion in HA4 DNAmight
have been detected by the studies described
above,thispossibilitycan bedirectlytestedby
restriction enzyme analysis. The mostdetailed
restriction map of T5currently availableisthat obtainedbycleavagewithanenzyme from
Hae-mophilusparainfluenzae, HpaI. The 26
frag-ments generated by HpaI cleavage ofT5st(+)
DNA have been ordered (5). The 11thand 17th
largestfragmentsare knownto contain the 7.9
and 99.6% interruptions, respectively. The
re-sults of HpaI digestion of T5st(+) and
-1
2
.""IF
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.329.401.74.483.2]SPECIFIC 731
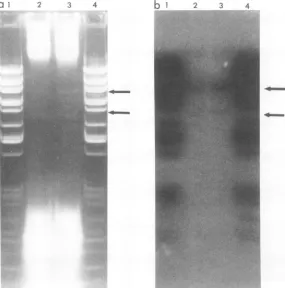
T5st(+)HA4 DNA are shown in Fig. 5. Each
digestwasanalyzedin0.7 and 1.5%agarosegels
toresolvethe entire spectrum offragments.The
mobilitiesof all 26HpaI fragments, including11
and 17, didnotappeartobe altered in T5HA4
DNA. A small alteration infragment 11might
be difficultto detect since thisfragment
comi-1 2 3 4
2 3
20
21,22
[image:6.505.249.440.502.627.2]1H112,
FIG. 5. Digestion of wild-type T5 and T5HA4
DNAs withHpaIendonuclease. Electrophoresiswas
carried out in 0.7%o(gels 1 and2) and 1.5% (gels3
and4) cylindricalagarosegels.Gels1and3,T5st(+)
DNA; gels2 and4,T5st(+)HA4 DNA.Fragment 11,
visibleingels1 and2, spansthe 7.9% interruption.
Fragment17,visible ingels3 and4,spansthe 99.6%
interruption.
grateswith fragments 12 and 13. Fragment 17,
however, is well separated from fragment 16,
which is only50basepairs larger. Consequently,
adeletion of20to30base pairs from fragment
17shouldhavebeeneasilydetected. Itis
there-fore likely that the HA4 mutation is a single,
hydroxylamine-induced base transition.
Two-step DNA transfer by T5HA4. The
interruptionat7.9%has been proposedtobea
signalormechanical barrierthatprevents
injec-tion beyond the FST (first-step transfer)
seg-ment of T5 DNA (1, 3). This hypothesis has
been questioned since mostT5DNA molecules
have additional interruptions between 0 and
7.9% (13, 16). It is also possible that, although
the 7.9%interruptionhasnoroleintermination
of further DNA transfer, it is the preferential
site ofbreakage when infectedcells aresubjected
toshearand thereforerepresentsoneend ofthe
FST segment as isolated. The HA4 mutation
provides a direct means to test both ofthese
hypotheses.
Two-step injection was demonstrated by
in-fecting E. coli F cells athigh concentration in
nutrient-free buffer. Under these conditionsonly
the FSTsegmentistransferredtothe host cell
(14). One portion of the FST complexes was
then subjected to shear forces to remove the
phage capsidand the remaining 92%ofthe
ge-nome. Only a small fraction of the infective
centers survived the shear treatment. Table 1
shows thatthefractionof infectivecenters
sur-viving shearing ofT5HA4 FST complexeswas
identical to that obtained for wild-type
com-plexes.Asecondportionof theFSTcomplexes
was not immediately sheared, but was diluted
into warmnutrientmedium and aerated for 15
min to allow transfer of the total DNA. With
thistreatmentboth theinfectivecentersand the
TABLE 1.
Transfer
ofT5wild-typeand HA4 DNAinto host cells
Conditions for transfer of: Expt Phagegenotype FST DNA Total DNA
no
IC%a 32p%b IC%,, 32p%b 1 Wildtype 7.5 - 100
HA4 5.3 - 111
-2 Wildtype 11.7 10 90 63
HA4 7.3 16 79 66
3 Wild type 0.2 14 106 71
HA4 2.4 12 100 67
aInfective centers after blending, expressed as a
percentage of infective centersinan unblended
con-trol.
b32P associated with thehostcells after blending, expressedas apercentageof32p initiallyadsorbed.
on November 10, 2019 by guest
http://jvi.asm.org/
majority of the 32P became resistant to shear in
bothwild-type andHA4infection.
Characterization of the FST DNA from
T5 wild-type and HA4 infections.
Intracel-lular DNA isolated from shear-treated FST
complexeswasanalyzed by electrophoresis (Fig.
6). T5st(+) HpaI fragmentswereused as
stan-dardsformolecular-weight determination. The
identity of the FST DNA was confirmed by
autoradiography. Thisanalysis showed that the
FST fragments from wild type- and
HA4-in-fectedcellsareidentical. Themajor 32P-labeled
fragment comigratedwith HpaIfragment4,
in-dicatingamolecular weight of6.0 x 106 (5) or
7.9% of T5st(+) DNA. This value agrees well
withpreviouslypublished values of6.3 x 106 for
duplex FST DNA (14) and 3.0 x 106 for FST
single strands (7). Therewas alsoaless
promi-nent 32P-labeled band that migrated ahead of
HpaI fragment 8 and thus has a molecular
weightof 3.8 x106(5). McCorquodaleandLanni
a l 2 3 4
(14) also observed a smallerfragment in DNA
isolated fromFSTcomplexes,but estimated its
sizetobe 1.5 x 106by sedimentation insucrose
gradients. McCorquodale (13) has attributed
thisfragmenttoshearing ofDNAextruded from
phage heads. We do notknow the significance
ororigin of thesmallerfragmentseenhereorits
relationship to the smaller fragment seen by
McCorquodaleand Lanni. Thelarge 32P-labeled
band appears to be intact T5 DNA which
es-capedshear.
The size of the major FST fragment is
ap-proximately equal to 7.9% of the T5 genome.
Thus,inwild-type infection, transferof the FST
DNA must terminate very near to the 7.9%
interruption, and sheartreatmentof the infected
complexes results in breakagenearthis
interrup-tion. The results obtained with T5HA4, which
lacks the 7.9% interruption, show thatthis
inter-ruption neither signals termination of FST
transfernorprovidesashear-sensitive site.
b
1
2 3 4
_-FIG. 6. Electrophoretic analysis ofintracellular DNAfrom FST complexes.(a)Appearance of gelstained
with ethidium bromide; slots 1 and 4, 32P-labeled HpaI-digested T5st(+) DNA; slot2, 32P-labeled DNA
isolatedfrom T5st(+)HA4 FST complexes;slot3,32P-labeled DNA isolatedfrom T5 wild-type FST complexes.
(b)Autoradiograph of thesamegel. Thearrowsindicate thepositions of the6.0x106- and3.8x106-dalton
phageDNAfragments. Thegelwasdriedundervacuum onfilterpaper and thenclamped againstKodak
RoyalBlueX-ray film for autoradiography.
.;.I-% ;%, n..t,v
I&',.
i. 7;.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.505.123.408.309.597.2]SPECIFIC INTERRUPTION-DEFICIENT 733
DISCUSSION
This report describes the properties of
T5HA4,amutantthat lacks oneof thespecific
sites where single-chain interruptions occur in
T5DNA.Analysis ofthesingle-chain fragments
produced by denaturation of T5HA4 DNA
clearly indicates that thismutantlacks the
prin-cipal single-chain interruption that occurs at
7.9%. Sincethis siteoccurswithin the 8.3% ofT5
DNA that is terminally repetitious, the HA4
mutation also results in loss of an analogous
interruptionat99.6%.
T5HA4wasisolated after hydroxylamine
mu-tagenesis andappears tobefree from detectable
deletionoraddition mutations. The interruption
deficiencyis therefore presumably dueto abase
substitution mutation thatoriginally occurredat
either 7.9 or 99.6%. As is the case for other
terminalredundancy heterozygotes, segregation
to homozygosity would be expected to occur
during subsequent growth. Alternatively, it is
possible that the HA4 mutation itself is not
located at 7.9and 99.6%. The observation that
HA4 is neither dominant nor recessive to the
wildtype only indicates thatthisparticular
in-terruption deficiency is not due to lack of a
diffusiblegeneproduct.Otherpossibilities,such
asinactivation ofadistalessential site, havenot
been eliminated. The only direct evidence on
this question has been obtained with mutants
thatlack the interruptionat32.6% (18). Several
mutations that resultinloss of the 32.6%
inter-ruption have been foundtobetightly linkedto
the nearby st(102) deletion (unpublisheddata).
Loss of the interruptionsat 7.9and99.6% in
T5HA4 DNAresults inanumber of changes in
the fragments produced by denaturation or
digestion with exonuclease III and SI nuclease.
Thesechangesreflect the loss of fragments that
normally terminate at 7.9 and 99.6% and the
appearance ofnew fragments that span these
sites. T5HA4 isparticularlyuseful for this kind
of analysis since the terminally repeated seg-ments of T5 DNA contain a high density of
secondary interruptions. These sites appear to
be clustered in the left half of the repeated
region between 0 to 4.1% and 91.7 to 95.8%.
Consequently, T5HA4 DNA contains virtually
all of thefragmentsof less than 4% ofanintact
strand(Fig. 2). These fragments extend from the
left end of the DNA to interruptions located
between 0.8 and 4.1%. Numerous fragments
whichequal4to8%ofT5st(+)DNA aremissing
in T5HA4 DNA. Thesefragmentsnormally
ex-tendleftwardfrom7.9%and 99.6%arereplaced
atthe left end of HA4 DNA, by a new set of
fragmentsthat extend left from 18.5% (Fig. 3).
The four secondary interruptions mapped by
thissetof fragments hadbeen previously located
byanalysis of restrictionfragments of T5 DNA
(16). The redundant regions contain additional
interruptions which couldbe mapped by more
extensiveanalysis ofT5HA4 DNA.
Itshould be noted, however, that at present
none of the new fragments that appear in
T5HA4 DNA have been unambiguously
mapped. It ispossible, for example, thatsome or
all of fragments A through E (Fig. 3) do not
extend as far rightward as the 18.5%
interrup-tion. The construction ofdouble mutants that
lack both the7.9and18.5% interruptions should
resolve thisquestion.
The observation that T5HA4 transfers its
DNA inanormal, two-step manner
unambigu-ously excludesarole for the 7.9%interruption in
this process. Because of its location, the 7.9% sitewas acandidate for thesignalthatinterrupts
transfer,provided that the left end is transferred
first. Although direct evidenceonthe direction
of T5 DNAinjection hasnotbeenobtained, the
available evidence ismostconsistent with initial
transfer of the left end (10, 20). If the rightend
istransferred first, a role forinterruptions can
also be excluded since the region immediately
around 92% islargelyfree fromevensecondary
interruptions (21). Normaltwo-steptransferhas
also been observed with T5HA23,amutantthat
iscompletelyfree frominterruptions(S. Rogers,
unpublisheddata).
The size of the FST segment isolated from
sheared T5HA4-infected cells is identicaltothat
obtained for wild-typeT5. Moreover, the band
containingthe HA4FSTsegmentisassharpas
thatseen forwildtype. Thus,eventhough the
FSTfragment equals7.9% of T5 DNA, the
in-terruption at7.9% doesnotprovideanessential
shear-sensitive site. This observation is
surpris-ing considering the known shear sensitivity of
single-chain interruptions (6) andsuggeststhat
the actual size of the initially transferred
seg-ment equals 7.9% of intact DNA. If the 7.9%
interruption had providedashear-sensitivesite,
the actual size of the initially transferred
seg-ment could have been substantially larger or
smallerthan 7.9%.
Avalue of 7.9%for T5 FST DNAimpliesthat
asmall portionof theterminalrepetitionisnot
included in the initially transferred segment.
Oneofthe functions oftheFST DNA is
exten-sivedegradationofhost DNA(12).Presumably,
the FSTDNA isprotected fromsimilar
degra-dationduring normal infectionalthough the
pro-tectivemechanismhasnotbeenidentified.Even
ifsuch amechanismexists,somedegradationof
FSTDNAcouldoccur.Therefore,byholdinga
portion oftherepeatedregioninreserve,T5has
on November 10, 2019 by guest
http://jvi.asm.org/
the potential for restoring, by recombination, a continuousgenome once DNA transfer has been completed.
ACKNOWLEDGMENTS
Thisresearch wassupported by grant PCM76-24036from theNational Science Foundation.S.G.R.wassupported bya training grant (HD-139) from the National Institutes of Health.
LITERATURE CITED
1.Abelson, J.,and C. A.Thomas,Jr.1966.The anatomy of the T5bacteriophage DNA molecule. J. Mol. Biol. 18:262-291.
2. Berns, K. I., and C. A.Thomas,Jr.1965.Isolation of high molecularweight DNA from Haemophilus influ-enzae.J. Mol. Biol. 11:476-490.
3. Bujard,H.1969.Locationofsingle-strandinterruptions in the DNA ofbacteriophage T5+. Proc. Natl. Acad. Sci. U.S.A. 62:1167-1174.
4. Carter, D.M.,andC. M.Radding. 1971.The role of exonuclease and ,Bprotein of phage A in genetic recom-bination. J. Biol. Chem. 246:2502-2510.
5. Hamlett, N. V., B.Lange-Gustafson,and M. Rhoades. 1977. Physical map of the bacteriophage T5 genome basedonthecleavage productsof the restriction endo-nucleasesSalI, SmaI, BamI, andHpaL J. Virol. 24: 249-260.
6. Hayward, G.S.1974.Uniquedouble-strandedfragments ofbacteriophageT5 DNAresulting from preferential shear-inducedbreakageatnicks. Proc.Natl. Acad. Sci. U.S.A.71:2108-2112.
7. Hayward, G.S.,and M. G. Smith.1972.The chromo-someofbacteriophageT5.II.Arrangement of the sin-gle-stranded DNA fragmentsin theT5+ and T5st(0)
chromosomes. J. Mol. Biol. 63:397-407.
8. Hertel, R., L. Marchi, and K. Muller. 1962.Density
mutantsofphageT5.Virology18:576-581.
9. Katz,L., D. T.Kingsbury,andD. R.Helinski.1973. Stimulation by cyclic adenosine monophosphate of plasmiddeoxyribonucleicacidreplicationand catabo-literepression of the plasmid deoxyribonucleic
acid-proteinrelaxationcomplex.J. Bacteriol. 114:577-591. 10. Labedan, R., M.Crochet,J.Legault-DeMare, andB. J. Stevens. 1973.Location of the first step transfer
fragmentandsingle-strand interruptionsinT5st(0)
bac-teriophage DNA. J. Mol. Biol. 75:213-234.
11. Lanni, Y. T. 1969. Functions of two genes in the first-step-transfer DNA of bacteriophage T5. J. Mol. Biol. 44: 173-183.
12. Lanni, Y. T., and D. J. McCorquodale. 1963. DNA metabolismin T5-infectedEscherichia coli: biochemi-cal function of a presumptive genetic fragment of the phage.Virology 19:72-80.
13.McCorquodale, D. J. 1975.The T-odd bacteriophages. Crit. Rev.Microbiol. 4:101-159.
14.McCorquodale, D. J., and Y. T. Lanni. 1964. Molecular aspects of DNA transfer fromphage T5 to host cells. I. Characterization of first-step transfer material. J. Mol. Biol. 10:10-18.
15. Masamune,Y., R.A. Fleischman, and C. C. Richard-son.1971.Enzymatic removal and replacement of nu-cleotides at single-strand breaks in deoxyribonucleic acid. J. Biol.Chem. 246:2680-2691.
16. Rhoades, M. 1977.Localizationof single-chain interrup-tions in bacteriophage T5 DNA. II. Electrophoretic studies. J. Virol.23:737-750.
17. Rhoades, M., and E. A. Rhoades. 1972. Terminal repe-tition in the DNA ofbacteriophage T5. J. Mol. Biol. 69:187-200.
18. Rogers, S.G., E. A. Godwin, E. S. Shinosky, and M.
Rhoades.1979.Interruption-deficientmutantsof bac-teriophage T5. I. Isolation and general properties. J. Virol. 29:716-725.
19.Rogers, S. G., and M. Rhoades. 1976. Bacteriophage T5-induced endonucleases that introduce site-specific single-chain interruptions in duplex DNA. Proc. Natl. Acad. Sci. U.S.A. 73:1576-1580.
20. Saigo, K. 1975.Tail-DNA connection and chromosome structureinbacteriophageT5.Virology 68:154-165. 21. Scheible, P. A., E. A. Rhoades, and M. Rhoades. 1977.
Localization of single-chain interruptions in bacterio-phage T5 DNA.I.Electron microscopic studies. J. Virol. 23:725-736.
22. Scheible, P. A., and M. Rhoades. 1975. Heteroduplex mapping of heat-resistant deletion mutants of bacterio-phage T5. J. Virol. 15:1276-1280.
23. Tessman,I. 1968. Mutagenic treatment of double- and single-stranded DNA phages T4 and S13 with hydrox-ylamine. Virology 35:330-333.
24. Thomas, M., and R. W. Davis. 1975. Studies on the cleavage of bacteriophage lambda DNA with EcoRI restrictionendonuclease. J. Mol. Biol. 91:315-328.
on November 10, 2019 by guest
http://jvi.asm.org/