Copyright01976 AmericanSocietyfor Microbiology PrintedinU.S.A.
Replication Process
of
the Parvovirus
H-1
V. Isolation
and
Characterization of
Temperature-Sensitive H-1 Mutants
Defective in Progeny DNA
Synthesis
SOLON L. RHODE III
Putnam Memorial Hospital Institute for Medical Research,Bennington, Vermont05201 Receivedforpublication25August 1975
Twotemperature-sensitive mutants of the parvovirus H-1 have been isolated
and characterized. Thesemutantsaredistinguishable
by
the immunofluorescentstaining of cells infected by them and by the thermolability of their
hemag-glutinins. Under restrictive conditions both mutants synthesize a missense
capsid protein defectiveinhemagglutination. Synthesisofthe viral DNA strand
was shown tobe diminished for both wild-type virus andthese mutants at the
restrictive temperature of 39.5C, but themutants were more defective than the
wild type at the less restrictive temperature of 38 C. Replicative form DNA
replicationwasnot decreased inthesemutants. It is proposed that H-1 requires
one ofthecapsid proteinsforthesynthesisofthe single-strandedprogeny DNA.
Replication ofthesingle-stranded DNA
par-vovirus H-1 involves the synthesis and
replica-tionof adouble-strandedreplicativeform (RF)
DNA(8).Progenyviral strand (V) DNAis then
synthesizedwith thecomplementarystrand(C)
DNA in the RF as template(8, 10).
Viral hemagglutinin, antigen, and the viral
proteins VP1 and VP2 have been shown to
dependonpriorDNAsynthesis, termed
hemag-glutinating (HA)-DNA synthesis,anevent that
takesplace ataspecific time nearthe end of S
phase of the infected cell (5, 9). Viral protein
synthesis and theinitiation ofRF DNA
replica-tion occur concomitantly shortly after
HA-DNAsynthesis (10). Although the initiation of
RF DNA replication was shown to require
proteinsynthesis, it was notdetermined ifviral
protein synthesis was required for RF DNA
replication or for progeny DNA synthesis (10).
The present study describes the isolation and
characterization of two temperature-sensitive
(ts) mutants ofH-i. The mutations occur in a
capsid
protein, a protein which isrequired forprogeny DNAsynthesis butnotRFDNA
repli-cation.
MATERIALS AND METHODS
Cell culture. The cells used inthisstudy are the SV40 transformed human newbornkidney (NB)cells routinely used for the plaque assayofH-1orhamster embryo cellsaspreviously described (6, 9). Thecells werecultivated inEagle minimum essential medium (FlowLaboratories, Autopowminimum essential me-diumEagle, modified) supplemented with10% heat-inactivated calf serum, glutamine, penicillin (10
Isg/
ml), streptomycin (5
,g/ml),
and sodiumbicarbonatetopH 7.6. Parasynchronous cultures, used for some purposes,werepreparedby incubationinculture me-diumMAG containing methotrexate (0.5gg/ml), ade-nine (13
gg/ml),
and Eagle minimum essential me-dium nonessential amino acidsfor 16 to 20hat37C. The methotrexate block was reversedby removing the medium, washing once with Hanks balanced salt solution, and refeeding with culture medium con-tainingthymidine (TdR) at 10-iM.Virus. Wild-type H-1 virus was propagated in hamster embryo cells and purified as previously described (9).
Infection of cells. NB cultures were infected by inoculating the monolayer with 0.1 or 0.2 ml of virus stock per60-mmpetri dish and adsorbing it at 33 C for 30 min. The cultures were washed twice with Hanks balanced salt solution, medium was added, and the cultures were incubated at the desired tem-perature.
Plaque assay. The plaque method ofvirusassay has been described (6). This method was modifiedin thelengthofincubationbefore the neutral red over-lay, both at thepermissive temperature (33 C) and
nonpermissive temperature (39.5 C) used here to
define the conditional mutants ofH-1. The overlay with neutral redwasmadeonday9 forthe assayat 33 C andon day6 forthe assay at 39C. Plaqueswere counted 16 to 24h aftertheoverlay with neutral red. Wild-typevirususually titeredasefficientlyat 39.5C as at 33C, but modest reductionsintiterrangingas highas 75% of the titer at 33 Cwerecommon.
Mutagenesis. H-1 virus (4 x 109PFU/ml), puri-fiedaspreviously described (9),wastreated with0.25 Mhydroxylamine asoutlined byTessman for bacte-riophage S13 (11). The reaction mixture contained onepart H-1 inH,O, twoparts0.1 MP04(pH 6.0),
and one part1 Mhydroxylamine(made pH6.0with NaOH) plus10-3MEDTA. Thereaction wasstopped
659
on November 10, 2019 by guest
http://jvi.asm.org/
660 RHODE
by diluting an aliquot 100-fold in regular medium
with 10% fetal calf serum, 10% tryptose phosphate broth, and 2% acetone at 4 C. In the absence of hydroxylamine, H-1 infectivity decreased only 20% after 4 h at 37 C. Hydroxylamine inactivated H-1 infectivityexponentially ata rate of 10-1/hof treat-ment to a final surviving fraction of 10- the last tested.
Isolation of ts mutants. A preparation of virus inactivatedtoasurviving fraction of10 wasusedas
the sourceofmutagenized virus.Thisvirus
prepara-tionwaspropagatedinhamsterembryo cells infected
atamultiplicity of infection of 0.1 for 48hat33Cto
dilute nonconditional mutants. With this
mutagen-izedvirusstock,diluted togive approximately 1 to5 plaques per 60-mm petri dish, individual plaques
were isolated from assays at 33 C with a Pasteur
pipette. The plaque isolate was stored in 0.5 ml of
N-2-hydroxyethylpiperazine-N'-2-ethanesulfonic
acid-buffered Eagle minimal essential medium and subsequently propagatedinNB cellsina35-mmpetri
dishor ina microtiterplate for 5daysat33C. The virus and cellswerethen harvested anddilutedtoa
final volume of 3 ml in N-2-hydroxyethylpiperazine-N'-2-ethanesulfonicacid-buffered medium. Viruswas
released from cells by freezing and thawing three times. Aliquots of 0.2 ml ofthe isolates were
inocu-latedonduplicate plates andassayedatthe permis-sive andnonpermissivetemperaturesforplaque
pro-duction. Isolates which showed a 100-fold orgreater titerat 33C thanat39.5Cwereselected for further
study. Prepared in this manner, wild-type isolates
generally produced20to100PFUat39.5C and 40or
moreat33C with undilutedinocula. Isolates
appear-ingtobetemperature sensitiveby thescreening test
were assayed again at the two temperaturesand in appropriate dilutions forquantitative determination oftiters.Insome cases,ts mutantswererepurifiedby
one or more plaque purifications. Mutants were
propagated in NB cells at 33 C once or twice to
producevirus stocks with titersof107or moreatthe permissive temperature.
RESULTS
Replication of wild-type H-1at33 and 39.5
C. The kinetics ofhemagglutinin and infectious
virus production in parasynchronous NB cells
for wild-type H-1 at33 and 39.5C were
exam-ined. The growth curves for HA synthesis and
infectious virus production were similarat-the
two temperatures (Fig. 1, Table 1). If the time
axis for HAsynthesisat33C is transformed by
a factorof0.5, the curves are nearly identical.
Theyieldof infectious viruswasreduced about
50% at39.5C comparedto33C.
Isolation ofH-1 mutantstsl and ts2.
Puri-fied wild-type H-1 was mutagenized with
hy-droxylamine to a survival of 10-1, propagated
onceatlowmultiplicity in NB cellsat33C, and
plaqueswereisolated and screened for
tempera-ture sensitivityasdescribed above. Isolates190
16
12
~ ~ C33C
39.5 C/ C
<~ ~ ~ 0
4 / 00
0 8 16 24 32
HoursPI
FIG. 1. H-1 HA synthesis in parasynchronous NB cultures infectedwithwild-typeH-i atamultiplicity
ofinfection of5to 10and incubatedat39.5 or33C
from I h p.i. to the time of harvest. Cultures were
harvested with their mediumatvarious timesp.i.and the HA titers weredetermined in the usualmanner
[image:2.491.261.450.58.247.2]withguinea pig erythrocytes.
TABLE 1. Replication ofH-i attherestrictive and permissive temperatures
Time PFU/culture
(hp.i.) 33C 39.5C
1/2
6.6x106
6.6x106
6 2.6x 106 1.6x 106
12 2.4x 106 2.0 x106
24 1.4x 107 1.6x 107
48 5.4x 107 2.0 x 107
120 5.8 x 10' 2.2x 107
aReplicate cultures of NB cells in exponential growth were inoculated with wild-type H-1 at a multiplicity of infection of 5 to 10. After a 30-min adsorption at 33 C, the cultures were washed twice with Hanks balanced salts, and medium was added andincubatedat 39.5 or 33Cforthetimesindicated. Cultureswere harvested, subjected to freeze-thawing three times, and H-1 infectivity was determined by plaque assayat 33C.
and 389, subsequently termed tsl and ts2,
respectively, produced higher titers atthe
per-missive than the nonpermissive temperatures,
asreported inTable 2. Subsequentpassages of
the virus stock maintained this difference. The
ratio of titer at 33C to that at 39.5 C did decline
with some propagations at 33 C, indicating a
possible selective disadvantage for the mutant
virus. It hasnot been determinedasyet ifthe
plaques appearing at 39.5 C are produced by
J.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.491.257.450.344.442.2]TABLE 2. Isolation ofH-1 mutants tsl and ts2a Virus 33C 39.5C 33C/39.5
PFU/ml PFU/ml C(PFU) tsl
Original-screening CL 0
Original I x 103 0
P2 0
P3 5xlO 1.7x103 2.9.A 104
P4 3.4 x10 4.2x10' 8.1 X 102 Plaque purified two 4.1 x10' 1.2 x 104 3.4 < 103
times P3
Plaquepurified two 3.8 x10' 1.1 X104 3.5x 103 times P3
ts2
Original-screening CL 5
Original 2.5x103 0
P2 5.3 x10' 9.8 x10' 5.6X 10' P3 2.5x 10' 8.5 x 103 2.9x 104 Plaquepurified two 3.1 x10' 1x10' 3.1 x 103
timesP3
aThe infectivity of tsl and ts2 H-1 mutants at the permissive and nonpermissive temperatures at various stages oftheir propagation are presented. Assays used innocula of 0.2ml. but titers are given as PFU/milliliter. CL indicates a complete lysisof the cell layer. The screening test is an assay of the plaque isolate undiluted after its first passage. A passage(P)represents a propagation of the virus for 5 days at 33CinNBcell cultures.
wild-type virus or the mutant, due to a
diffi-cultyinrecovering virus from these plaques.
Characteristicsof thereplicationof tsland
ts2. Anumberof parameters ofH-1replication,
including synthesis of HA-DNA, HA, and H-1
antigen, were examined withtsl and ts2atthe
permissive and nonpermissive temperatures.
Both tsl and ts2 were indistinguishable from
wild-type H-1 for these parameters at 33 C
(Table3).
At 39.5 C, tsl andts2produced little HAby
16hpostinfection (p.i.) inparasynchronous NB
cells (Table 4). When cultures of tsl and ts2
were shifted down to 33 C for various periods,
the HA titer increased at least 16-fold andwas
maximal in2h. The appearance of the HA took
place at 33 C, even when DNA or protein
synthesis was inhibited with arabinofuranosyl
cytosine (araC) (10
Ag/ml)
orcycloheximide (50Ag/ml),
respectively, during
the shiftdown andthe 15 min preceding the temperature shift.
Sincecycloheximide
probably
does notachievea complete inhibition ofprotein synthesis and
viralproteinsynthesisaccountsforonlyasmall
percentage of totalcellprotein
synthesis,
itwasnecessary to confirmthat viral HAsynthesisis
sensitive toinhibition by the drug. To do
this,
the inhibition of wild-type HA synthesis by
cycloheximide (50
gg/ml)
at 37 C in the samecell system wascompared to that of
actinomy-TABLE 3. Synthesis of HA-DNA, HA, and FA by H-I ts mutants
HA-DNA HA FA
Virus
33 39.5 33 39.5 33 39.5
Wildtype + + + + + +
tsl + + + _ + +a
ts2 + + + - +
-aReplicate parasynchronous NB cultures were in-fected with wild-type H-1, tsl, or ts2 and incubated for 16 h at 39.5 C or 20 h at 33 C. Cultures were harvested for HA determination or fixed for FA staining. HA-DNA synthesis at39.5 C for tsl orts2 wasdetermined by adding araC (10/Ag/ml)tocultures at 39.5 Cat 16 hp.i. for 15 min and then shifting the cultures to 33Cfor 3hin the presence ofthedrug.If theHA titerincreasedtothe samedegreeas acontrol culture receiving no araC, then HA-DNA synthesis had occurredat39.5C preceding the addition of araC.
'Nuclei contained FA material in an abnormal pattern asdescribed inFig. 3.
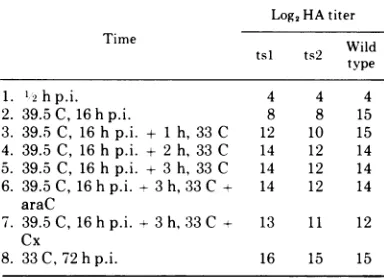
TABLE4. HAsynthesisby tsl, ts2, and wild-typeH-il Log2HA titer
Time tsl ts2 Wild
type
1. '2hp.i. 4 4 4
2. 39.5C,16hp.i. 8 8 15
3. 39.5C, 16 h p.i. + 1h, 33 C 12 10 15 4. 39.5C, 16 hp.i. + 2h,33 C 14 12 14 5. 39.5C, 16hp.i. + 3h, 33 C 14 12 14 6. 39.5C, 16hp.i.--3h,33C+ 14 12 14
araC
7. 39.5C, 16hp.i. +3h,33C + 13 11 12 Cx
8. 33C, 72hp.i. 16 15 15
aReplicate dishes ofparasynchronous NB cultures
were infected with tsl, ts2, or wild-type H-1 and placedat 33 or39.5C. At the times indicated, cultures wereharvestedforHA determinations or shifted from 39.5to 33C for various times before harvest (cultures 3through7). Cultures 6 and 7 were treated with araC (10 ug/ml) or cycloheximide (Cx) (50 gg/ml) for 15 minbefore andduring the temperatureshiftto 33C. All valuesare the log2 HA titer.
cin D at 5
qg/ml
(1 ug/ml was previouslyfound to be a potent inhibitor of viral HA
synthesis [9]). The actinomycin D control was
necessary to establish that viral mRNA was
present at the time of addition of
cyclohexi-mide, thus allowing the inference that any
increment in inhibition by
cycloheximide
com-paredtoactinomycinDwasdueto aninhibition
oftranslation. It was found that cycloheximide
caused immediate inhibition ofthe
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.491.248.443.77.161.2] [image:3.491.249.441.307.446.2]662 RHODE
tion of HA ateither10.5 or12hp.i. On the other
hand, the total HAunitsper culture increased
by eight- and
fourfold,
respectively, by
16hp.i.inthe presenceof 5
,g
ofactinomycin
Dperml.Thus HA synthesis was sensitive toinhibition
by cycloheximide. This indicates that
HA-INA, viral mRNA, and the hemagglutinin
protein had been
synthesized
before theshift-down with tsl and ts2. It appears, therefore,
that the temperature-sensitive
phenotype
istheresult of an altered polypeptide that does not
assume its active structure at 39.5C.
Thestabilityofthe HAtotemperature
inacti-vation wasdetermined
by
using thevirusstocksoftslandts2(produced at33C) and wild type
atpH7.2.The results showeda
sharp
tempera-ture transition forthe inactivationofthe HAat
72 to 78 C. The temperature
completely
inac-tivating tsl was reduced about 3 to 4 C
compared to ts2 and
wild-type
virus(Fig.
2).The temperature for activation of the HA for
the wild-type H-1 agrees with
previously
pub-lished data (2).
To summarize, we have found that the
con-+
0
-I
-4
1--E
7
-8 2
ts
II
.11
1I
oll
I I
I
6I
5 Is
II II
I I
I t
3 4 %
II % II %
7 11 1.
II % II
t
I'll.I
I..a
4 68 70 72 74 76 78
[image:4.491.54.244.315.570.2]0C
FIG. 2. ThermalstabilityofH-i hemagglutininfor wildtype, tsl,andts2.Aliquots(1 ml) of virus stocks diluted 1:10 in Tris-buffered saline, pH 7.2, were
incubated for20minatvarioustemperatures,chilled in ice water, and the hemagglutination titers were
determined. The values shown are the log, of the
surviving fraction of thehemagglutinin relative toa
controlmaintainedat0C.
version ofthe mutant viral
hemagglutinin
pro-teins to an active form is deficient at 39.5 C.
Once the virus is
formed,
the ts2 HA hasa heatstability
similar to that of thewild-type
virusand tsl shows a
slightly
reduced transitiontemperature.
Immunofluorescence of tsl- and
ts2-in-fectedcells. H-1
antigen synthesis
wasstudiedinasynchronousNBcultures incubatedat 33or
39.5 C for 24h afterinfectionwithtsl, ts2, and
wild-type
H-1. At 39.5C,
tsl producedstrongly
positive
fluorescent nuclei (fluorescentantibody
[FA])
with an abnormalglobular
pattern
ofantigen,
illustratedinFig.3B.Cultures infectedwith ts2 at 39.5 C produced little or no FA
staining (Fig. 3D),
asin theuninfected controlculture
(Fig. 3G).
However, infectionwith ts2at33 C resulted in many FA nuclei with the
wild-type
FA staining pattern (Fig. 3A,C).
Cultures infected with ts2 for 24 h and then
shifted down to 33 C for 1 h, with and without
araC or cycloheximide as above, showed a
gradual transitiontoFAnuclei withawild-type
staining
pattern.Sincetsl and ts2 aredistinctly differentwith
regard
to FA staining at thenonpermissive
temperature and the sensitivity oftheir HAto
heat inactivation, they must contain different
mutations.
It should be noted that antisera
prepared
using
guinea pigswith sodiumdodecylsulfate-disrupted purified H-1 as the antigen by the
procedure ofJohnson et al. (3), as well as the
very high-titer antisera obtained from
chroni-cally infected hamsters inoculated with
suble-thal doses of H-1 at birth, produced brightly
positive stainingofts2-infectedcellsincubated
at 39.5 C. Thus ts2 capsid proteins have been
transported to the nucleus under restrictive
conditions. We also observed that nuclei
in-fectedwith tsl underrestrictiveconditionswere
stained in thepatternofwild-type-infectedcells
by
thesodium dodecylsulfate-disruptedwhole-virusantisera andintheabnormal pattern (Fig.
3B) by antisera to purified VP2 (4). Our
anti-VP1serumwas tooweak toestablishitsstaining
patternsintheseexperiments. Thus these
find-ings cannot be usedtodeterminetheidentityof
themutantpolypeptide.
Viral DNA synthesis. The effects of the
mutations of tsl andts2onviralDNAsynthesis
were examined. As described above, HA-DNA
synthesis occurred undernonpermissive
condi-tions for both mutants. It was of interest to
determine ifRF DNA replication and progeny
DNA
synthesis
were affected by these muta-tions.Intheabsenceof apractical, direct,
quantita---1
J. VIROL.
I
on November 10, 2019 by guest
http://jvi.asm.org/
FIG. 3. Immunofluorescentstaining of wild-typeH-i and tsmutant-infected NB cells. NB cells, grown on coverslips, wereinfected withH-i in the usual mannerand incubated at the restrictive (39.5 C) or permissive temperature(33C).Atthe termination of incubationat 24hp.i., the cover slips were quickly rinsed in Hanks balanced salts andfixedwithacetone 15to20minat 4C. The dried cover slips were stained by the indirect immunofluorescent method using anti-H-i hamster sera (hemagglutination inhibition titer of 210) and
fluorescein-conjugated rabbit anti-hamster immunoglobin. (A) tsl, 33 C; (B) tsl, 39.5 C; (C) ts2, 33 C; (D) ts2,
39.5C; (E)wild type,33C;(F)wild type, 39.5 C;(G) uninfected, 39.5 C. (Magnification x460).
tive measureofviralsingle-strandedDNA
syn-thesis, the extent of incorporation of
[3H]bromodeoxyuridine (BUdR) into the V
strandand itsCstrandwasusedtoestimate the
proportion of V strand to C strands newly
synthesized in RF DNA. Thisprovidesa
quali-tative measure ofthe extent of progeny DNA
synthesis. The method employsashort period of
BUdR substitution for TdR to label the newly
synthesized
DNA andtoallowseparation
oftheVand C strands.
5-Fluorodeoxyuridine
(FUdR)
is used to prevent fluctuations in the
specific
activity ofthe BUdR
incorporated
with respecttoTdR.Vstrand
synthesis
inexcessofC strandsynthesis
cannot be ascribed tosemiconserva-tive replication of RF DNA and is thus a
qualitative
measure ofthe presence of progenyDNA synthesis (8). Thismethod does have the
VOL. 17, 1976
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.491.38.434.57.483.2]664 RHODE
advantage over a more directmeasure of
prog-eny DNAaccumulation inthat itdetects
prog-enyDNAsynthesis, evenif the progeny DNA is
rapidly degraded for some reason, such as
defective encapsidation.
Infected cultures were density labeled with
BUdR forsubsequentseparation oftheVandC
strands of RF DNA. The relative rates of
synthesis ofeach strandforwildtype, tsl, and
ts2could then be measuredfor comparison. In
hamsterembryo cells infectedat37C, RF DNA
incorporated BUdR asymmetrically with a
higherrate ofsynthesisfortheVstrand thanfor
the C strand (8, 10). This was the expected
result for anRF DNA pool engaginginprogeny
DNAsynthesisbyastranddisplacement
mech-anism in addition to semiconservativeRFDNA
replication. If a mutant is defective inprogeny
DNAsynthesis butnot in RF DNA replication,
the RF DNA will incorporate BUdR intoequal
numbers of V and C strands andinproportion
to their TdR contents
(V-TdR/C-TdR
= 1.15)(7). A mutant defective in RF replication will
shownoincorporation intoRFDNA.The
exper-iment wascarriedoutby infecting
parasynchro-nous NB cultures withtsl,
ts2,
orwildtype atthe restrictive temperature. At 16 h p.i. the
cultureswere treated with FUdRfor 30min to
exhaust the cell poolsofTdR and then labeled
for 1h with [3H
]BUdR
inthepresence ofFUdR.Viral DNA was extracted by the Hirt method
andanalyzedbyzonalcentrifugationinneutral
sucrose. The results (Fig. 4) showed increased
labelingofRFDNAfor tsl (andts2,notshown)
relative to the wild-type virus. In a similar
experiment, the region of the sucrose gradient
that contains the RFDNA contained lessthan
1% of the radioactivity for a mock-infected
control culture as that recovered from a
tsl-infected culture andnopeak waspresent.Thus
tsl (and ts2) does not show any defect in RF
DNA replication. In addition, there is some
change inthe sedimentationpatternconsisting
of a decrease in the material sedimenting at
fractions 13 and 14, the expected position of
progeny DNA (8). A DNA peak, which is
usually more prominent than shown here, is
commonly seen at this position in
sedimenta-tion patterns of Hirt extracts of H-1-infected
NBcells asopposed to infectedhamsterembryo
cells,whetherBUdRorTdR is used forlabeling.
Todeterminetherelativerates ofsynthesisof
Vstrands versusC strands bymeasuring
incor-poration ofBUdR into the V and C strands of
the RFDNA,the peakfractions of the RF DNAs
(Fig. 4) werepooled, precipitatedwithethanol,
and subjected to equilibrium centrifugation in
Cs2SO4
after heat denaturation as previously0~~~~~~~~
3
ts2
0'
1 5 10 15 20 25
FrOction No. 9
FIG. 4. Zonal sedimentation in neutralsucrose gra-dients ofwild-type H-i and tsl viral DNA synthesized atthe restrictive temperature(39.5 C). Wild type and tsl wereused to infect separate replicate culturesof synchronized NB cellsat amultiplicity of infectionof
5; cultures were treated with FUdR (0.5Ag/ml)16to 17hp.i. and thenlabeled with [3H]BUdR (1 Ci/10-9 mol) 17to18hp.i. Viral DNA wasextractedby the Hirt method.Sedimentationwascarriedoutin
gradi-entsof 5 to 20% sucrose(wt/vol) in50mM Tris-hydro-chloride, pH 8.0,1MNaCI,1mMEDTA, and 0.15% Sarkosyl in the SW25 Beckmanrotorfor 18 h at25,000 rpm at 4 C.Aliquots were measured for radioactivity byliquid scintillationspectrometry aspreviously (8). Theexpectedsedimentation position of virion DNA is indicated by the arrow labeled SS (8).
described (8, 10). The accumulations of
[3H ]BUdRwerenearlyequalforboth strands of
RF DNA for tsl and for ts2 (notshown) at39.5
C. The wild-type virus incorporated
considera-bly less BUdR into the V strand relative to the C
strandin NB cells at 39.5 C than waspreviously
found for wild-type virus in hamster embryo
cells at 37 C (V/C = 3.5; reference 8). Since
wild-type virus shows a twofold reduction in
PFUproductionwhenpropagatedat 39.5C,the
partial defectiveness of V strand synthesis at
the restrictive temperature may account for
this.
To establish that a mutation in tsl affects
progeny DNA synthesis, theabove experiment
wasrepeated at the lessrestrictivetemperature
of 38 C in hamsterembryo cells which permit
more virally related DNA synthesis (8).
Para-synchronous hamsterembryo cultures were
in-fected withwild type or tsl andincubatedat 38
C, from 30 min p.i. The cultures were then
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.491.255.442.63.271.2]labeled with [14C]TdR (1 pCi/ml, 62 M
Ci/
mmol) in the presence of FUdR (0.5 ug/ml),
from 14 to 17 h p.i. At 17 h p.i., the cultures
were washed twice with warm Hanks balanced
salts and incubated from 17 to 18 h p.i. with
FUdRand [3H]BUdR (5
gCi/ml,
2.5Ci/mmol).Viral DNA was extracted by the Hirt method
with Pronase digestion (4) and RF DNA was
preparedby zonal centrifugation as above. The
pooled RF DNA fractions were precipitated
withethanol, redissolvedin50 mM
Tris-hydro-chloride (pH 8.0), 10 mM EDTA, and heat
denatured
by
treating 6 min in a boiling-waterbath.Thetubeswerequenched inice water and
subjected to equilibrium centrifugation in
Cs2SO4 (Fig. 5).
Wild-type
virus shows a cleardisproportionate incorporation of [3H]BUdR
into theV strand ascomparedto the C strand.
The mutant tsl synthesized nearly equal
amounts of V and C strands even at 38 C,
indicating that it is relatively defective in V
strand synthesis in comparison to wild-type
H-1. The RF DNA labeled
by
[14C]TdRpro-vides a reference for light DNA and it can be
seenthatmostofthisDNA was not converted to
hybrid
density
in the period 17 to 18 hp.i.DISCUSSION
Temperature-sensitive mutants of the
par-vovirus H-1 have been isolated and partially
characterized. In this report, the properties of
two mutants are described. Both of these
mu-tants aredefectiveinplaque productionat39.5
C. Theyalso showinhibitionof viral
hemagglu-tininsynthesisandalterationsintheir
immuno-fluorescent antigen productionat therestrictive
temperature. It was found that for both of the
mutants thehemagglutinin, and for onemutant
(ts2), theantigen, appearedininfected cultures
after shiftdown to thepermissive temperature,
even when protein synthesis was inhibited by
cycloheximide. Thus these mutations are
caus-ing the synthesisof amissense capsid
polypep-tide with an alteredstructure atthe restrictive
temperature. This is the expected case for a
temperature-sensitive change in phenotype. In
further supportofthis,the hemagglutininoftsl
was found to have an increased sensitivity to
thermal inactivation.
We have previously shownthat the capsid of
H-1 consists oftwo polypeptides, VP1 and VP2
(5). Which of these two polypeptides contains
the mutation is under study. An earlierreport
8
!
C H-H RF
Hybrid
RFAI
6
/v\
4~~~~~~~~
2~~~~~~~
OO\b~~0 ~ ~ ~ ~ ~ V
0
5
BI
15 0
~15
20 25 30 35 4Fraction
No.FIG. 5. Isopycnic gradientcentrifugationofheat-denatured [3H]BUdR-substituted wild-type RF DNA(Fig.
5A,and tsl RF DNA,Fig. 5B).Parasynchronoushamsterembryocultures wereinfectedwithwild-typeH-1or tsW asdonepreviously (4) and incubatedat38Cfrom 1hp.i. From 14 to17hp.i.thecultureswereincubated withFUdR (0.5 Ag/ml)and ['4C]TdR (1
ACi/ml,
62mCi/mmol).At 17 hp.i. the cultureswerewashed twicein Hanks balanced salt solution and incubatedfrom17to18hp.i.in mediumcontaining FUdR and [3H]BUdR(5 MCi/ml,2.5Ci/mmol).At18h,theviral DNAwasextractedandRFDNAwasisolatedasinFig.4.RFDNAwas heatdenatured andsubjectedtoequilibrium centrifugation inCs2SO4. The centrifugation wasconducted inCsS04, 25mMTris-hydrochloride (pH8.0),5mMEDTA, and0.15%Sarkosyl for48hat35,000rpm at4 C
in the type 40 fixed-angle rotor. The density positions of the viral strand DNA is indicated by V, the complementary strandby C; theheavy,fullysubstitutedRF DNA andhybriddensity RF DNAaresolabeled. Gradientswereprocessedaspreviously (8). Symbols: (0) 14C, (0) 3H.
VOL. 17,
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.491.106.388.341.544.2]666 RHODE
has identified thehemagglutininandviral
anti-gen ofthe closelyrelated parvovirusKRV with
the major capsid component, protein
B,
whichsuggests VP2 may be the affectedprotein (11).
The effect ofthese capsid mutations on viral
DNA synthesis was also examined. H-1 DNA
synthesis has been classified into three
cate-gories: (i) parental RF DNA synthesis, (ii) RF
DNA replication, and (iii) progeny DNA
syn-thesis (8, 10). A DNA synthetic event upon
which subsequent VP1 and VP2 synthesis is
dependent andwhich occurs near the end of S
phase in the infected cell was termed HA-DNA
synthesis (9). It has been shown here that tsl
and ts2synthesize viralproteins atthe
restric-tive temperatureand thus HA-DNAsynthesisis
not defective. If RF replication required viral
protein synthesis, which in turn requires
HA-DNA synthesis (5), then HA-DNA synthesis
probably represents parental RF DNA
synthe-sis.
Similarly, tsl and ts2 produced normal or
increased levels of RF DNA. By using BUdR
substitution for TdR, it is possible to separate
the V strand and C strand of RF DNA and
compare the relative amounts of the density
label incorporated intoeach strand (8). Inthis
manner it was found that tsl and ts2 show no
evidence of asymmetric synthesis of V strand
DNA as a result of progeny DNA synthesis at
39.5C.Wild-typeH-1 was alsorelatively
defec-tive in V strand synthesis versus C strand
synthesis at that restrictive temperature. By
repeatingthecomparison atthe lessrestrictive
temperature of 38
C,
it was shown that themutation in tsl rendered V strand DNA
synthe-sismore sensitive tothermal inactivation than
that ofwild-typevirus.Thusacapsidprotein of
H-1, apparently thehemagglutinin, isrequired
fortheasymmetricsynthesisof Vstrand DNA.
A DNA moiety was observed consistently in
neutralsucrosegradients ofthe Hirt extracts of
NB cells infected with wild-type H-1 thatwas
decreased or absent with tsl infection. This
DNAsedimentedata ratesimilar tothatofH-1
viralDNA, butit has not yetbeenidentified as
H-1progeny DNA. The sedimentation patterns
of viral DNA extracted from hamster embryo
cells infected with wild-typeH-1 do not show a
peakatthat position (8). If that DNA proves to
be viral progeny DNA, then there may be
cellular factors that play a role in the virion
assembly and various cell types may exhibit
different efficiencies for this process. In the case
of adeno-associated virus, a similar form of
defectiveassemblyhas been suggested. Thus, in
thepresence of the partial helper virus, herpes
simplex virus, adeno-associated virus protein,
and DNA synthesis occur, but a late step in
infectious virus production is defective and
requires an adenovirus helper function (1). However, it has not been shown herpesvirus
infection is not inhibitory to adeno-associated
assembly. Minute virus ofmice, another
par-vovirus, has been shown to produce a pool of
viral DNA
readily
extractable by the Hirtprocedure (12). For both minute virus of mice
and H-1, the virions are not
lysed
by sodiumdodecyl sulfate at room temperature, so that a
Hirtextraction without Pronase does not
liber-ate DNA fromfully assembled virions. In
recon-struction experiments, H-1 viral DNA was
added to the sodium dodecyl sulfate lysing
bufferand was recovered in the final
superna-tant with a 50 to 60% efficiency. Therefore, it
canbe concluded thatnosignificantpooloffree
viral DNA is present in infected hamster
em-bryocells. Whetherthe rate of incorporation of
progeny
viral DNA into virions isdependentoncertain cell functions assuggested will require
furtherstudy.
ACKNOWLEDGMENTS
This research was supported by Public Service grants CA13414-03andCA07826-10fromthe National Cancer Insti-tute.
The guinea pig and hamster antisera used in thisstudy werekindlyprovided by Helene Toolan. The author gratefully acknowledgesK.Ellem andH.Toolanforhelpful criticisms of the manuscripts, Jessica Bratton for expert technical assistance, andVirginiaHaasand Janeen Prattforsecretarial duties.
LITERATURE CITED
1. Boucher, D. W.,J. L.Melnick, andH. D. Mayor. 1971. Nonencapsidated infectious DNA of adeno-satellite virus in cells coinfected with herpesvirus. Science 173:1243-1245.
2. Greene, E.L. 1965.Physical and chemicalproperties of H-1 virus. I. pH and heat stability of the hemag-glutinating property. Proc. Soc. Exp. Biol. Med. 118:973-975.
3. Hirt, B.1967.Selectiveextraction ofpolyoma DNAfrom infected mouse cellcultures. J. Mol. Biol.26:365-369. 4. Johnson, F. B., N. R. Blacklow, and M. D. Hoggan.1972.
Immunological reactivityofantisera preparedagainst the sodiumdodecylsulfate-treated structural polypep-tides of adenovirus-associated virus. J. Virol. 9:1017-1026.
5. Kongsvik, J.,J.Gierthy,and S. L. Rhode. 1974. Replica-tion process of the parvovirus H-1. IV. H-1 specific proteins synthesized in human NB kidney cells. J. Virol.14:1600-1603.
6. Ledinko, N. 1967.Plaqueassay oftheeffects of cytosine arabinosideand 5-iodo-2'deoxyuridineonthe synthesis ofH-1virusparticles. Nature(London) 214:1346-1347. 7. McGeoch, D. J.,L. V. Crawford,andE.A.Follett. 1970. The DNA'softhreeparvoviruses. J. Gen. Virol. 6:33-40 8. Rhode, S. L. 1974.Replication process of the parvovirus H-1. II. Isolation and characterizationofH-1 replica-tive formDNA.J. Virol. 13:400-410.
9. Rhode, S. L.1973.Replicationprocess of the parvovirus H-1. I. Kinetics in aparasynchronous cell system. J. J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 17, 1976 Virol. 11:856-861.
10. Rhode, S. L. 1974. Replicationprocessoftheparvovirus H-1. III. Factorsaffecting H-1 RF DNA synthesis. J. Virol. 14:791-801.
11. Salzman, L. A., and W. L. White. 1970.Structural pro-teins of Kilham rat virus. Biochem. Biophys. Res. Commun. 41:1551-1556.
REPLICATION OFPARVOVIRUS H-1
667
12. Tattersall, P., L. V.Crawford, and A. J. Shatkin. 1973. Replication ofthe parvovirus MVM. II. Isolationand characterization ofintermediates in thereplicationof the viraldeoxyribonucleic acid.J. Virol. 12:1446-1456. 13. Tessman, I. 1968. Mutagenic treatment ofdouble- and
single-strandedDNAphagesT4 andS13with hydroxyl-amine. Virology 35:330-333.




![FIG. 5.MCi/ml,5A,HankswithtsWheatGradientsincomplementaryCsS04, the as Isopycnic gradient centrifugation of heat-denatured [3H]BUdR-substituted wild-type RF DNA (Fig](https://thumb-us.123doks.com/thumbv2/123dok_us/1564371.109036/7.491.106.388.341.544/hankswithtswheatgradientsincomplementarycss-isopycnic-gradient-centrifugation-denatured-budr-substituted-dna.webp)