0022-538X/78/0026-0468$02.00/0
Copyright©1978 AmericanSocietyforMicrobiology PrintedinU.S.A.
Size
Analysis and Relationship of Murine
Leukemia
Virus-Specific
mRNA's: Evidence for Transposition of
Sequences
During Synthesis and Processing of Subgenomic mRNA
HUNG FAN*AND INDERM.VERMA
TumorVirologyLaboratory, The Salk Institute, San Diego, California 92112
Receivedforpublication 28 December 1977
Virus-specific mRNA from purified polyribosomes
of mouse cells infected withMoloney
murine leukemia virus(M-MuLV)
wasanalyzed by
electrophoresisin
agarose
gels,
followedby hybridization
ofgel slices
withM-MuLV-specific com-plementary DNA (cDNA). The size resolution of the gels was better than that of sucrose gradients used in previous analyses, and twovirus-specific
mRNA's of38S and
24S
weredetected.
The
24Svirus-specific mnRNA is
predominantly
derived from the 3' half of theM-MuLV genome, sincecDNA,fiaga,po
(complemen-tary to the 5'halfof the M-MuLV
genome) could
notefficiently
annealwith this
mRNA. However, sequences complementary to cDNAsynthesized
fromthe
extreme 5' end of M-MuLV 38S RNA(cDNA 5')
are present in the 24S virus-specific mRNA, since cDNA 5' (130nucleotides) efficiently
annealed withthis
mRNA.
The annealing of cDNA 5' was not due to repetition of 5'terminal
nucleotide
sequences atthe 3' end of M-MuLV 38S RNA,
sincesmaller
cDNA 5' molecules(60
to70nucleotides), whichlikely
lack the terminal repetition,also
efficiently
annealed with the 24S mRNA. The sequences in24S
virus-specific
mRNA
recognized
by
cDNA 5' arenotpresent
in 3'fragments
of virionRNAthat
are the samelength. Therefore, it appears that RNA sequences from the extreme 5'end
of the M-MuLV genome may betransposed to sequences from the 3'half
of
the M-MuLV 38S RNAduring synthesis
andprocessing
of the 24Svirus-specific mRNA. These results
mayindicate
aphenomenon similar
tothe
RNAsplicing
processes that occur during synthesis of adenovirus andpapovavirus
mRNA's.RNA tumor viruses
contain
asingle-stranded
used toelucidate
therelationships
ofintracel-diploid
RNAgenome which ispositive
stranded lularpolyadenylic acid[poly(A)]-containing
vi-and
approximately 9,000
nucleotideslong
(sedi-
rus-specific
RNAs(19, 35).
In the resultsre-mentation value
38S) (3,
4,10). This
38SRNA
ported here,
wehaveused
specific cDNA
probes codes forthree classes
ofviral
structural
pro- tostudy theintracellular virus-specific
mRNA's teins:the
internal structural
proteins (products
of
mousecells infected with Moloney
murine ofthe
gaggene),
envelope
glycoproteins (prod-
leukemia virus (M-MuLV),
anRNA tumor virus uctsofthe
envgene), and
reversetranscriptase
which
does notmorphologically
transform
cells.
(product of
thepol gene) (3).
Inaddition,
virusesThe
relationship of the viral mRNA's
wasdeter-that
morphologically
transform fibroblasts en- mined, and indications of transposition of virus-code aprotein
responsible
for this transforma-specific
RNA sequences insubgenomic
mRNA tion(a
product
of the srcgene) (3).
No other wereobtained.
viral
proteins
have yetbeenidentified, although
it is
possible
that nonstructuralviral
proteins
MATERLS AND METHODScould exist. Cells, viruses,andmaterials. M-MuLV clone4A
Studies of the
virus-specific
mRNA's in cellscells,
alineof NIH-3T3 cellsclonally
infected withM-productively
infected with RNA tumorviruses MuLV,were grown onmonolayer in Dulbecco-modi-haveindicated thatvirus-specific
mRNA existsfled
Eagle medium supplemented with 5% calfserum. in twogeneral
size classes: mRNAequivalent
in Inall
experiments, exponentially growingcells
fromlengthtothegenomisui
3subconfluent
cultures were used. Purified M-MuLVlengthtothebgenomic
subunit(38S2729).
RnA,
aend
wasobtained
fromtissue
culture supernatants ofM-subgenomic
mRNA (11,15,23,i27,
29).Inrecent
MuLV clone 1cells(14),anotherlineofNIH-3T3cells studies on aVian sarcoma viruses, complemen- infected withM-MuLV, as described previously (11). tary DNA(cDNA) hybridization
probes specific The deoxyribonucleoside triphosphates were ob-for differentregions
of the viral genome were tained from P-L Biochemicals.[3H]dTTP
(specific468
on November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26,1978
activity, 60 Ci/mmol) was obtained from Schwarz/ except thatdetergent concentrationwas0.02% Noni-Mann, and
[32P]dGTP
waspurchased from ICN, Ir- detP-40, andapproximately 2 mg of calf thymus DNA vine, Calif. Both oligo(dT)1o and oligo(dT)-cellulose primerper ml wasincluded. After 3 h ofincubation at(T-3) were purchased from Collaborative Research. 37°C, maximalincorporation had occurred. The re-Purified avian myeloblastosis virus reverse transcrip- sultant cDNAprobe was then processed for hybridi-tasewaskindly givento usby M. T. Lai. zationasdescribedpreviously(11).Specificactivity of Fractionation of cells and preparation of the cDNA was 1.6 x 107 cpm/ug, as calculated from mRNA. For cell fractionation, cells were removed theinput specific activity of the labeled deoxynucleo-fromthe tissueculture dishes by trypsinization on ice, sidetriphosphate (TTP). The cDNA wasquite rep-as described previously (12). Purified polyribosomes resentative of the 38S viralRNA, sinceatanRNA to wereprepared as describedpreviously, be pelleting of DNA ratio of1:1 50% ofthe cDNAcouldhybridize, polyribosomes fromcytoplasmic extracts through1M and ata2:1 ratioall of the cDNAcould hybridize (D. and2M sucrose (13,15). Thepelleted polyribosomes Dolberg, personalcommunication).
weredissolved in2mlof SDS buffer(0.1 M NaCl,0.01 ThecDNA(,gpIoonwaspreared in the following man-MTris[pH
7.4],
1mMEDTA) containing 0.5% sodium ner. 3H-labeled Moloney murine sarcomavirus-spe-dodecyl sulfate (SDS), and extracted twice with cific cDNA(1.7 x 107 cpm/,ug), which had been pre-phenol-chloroform (26). The extracted RNA was then pared from purified Moloney murine sarcoma virus precipitated with2volumes of ethanol and storedat clone 124(2, 8) using exogenously added calf thymus -20°C until use. In some cases, polyribosomalRNA DNAprimer,waskindly providedby DavidDolberg. was bound to oligo(dT)-cellulose essentially as de- The cDNA(106 cpm) wasannealed with 15 ugof3 scribedpreviously (12),exceptthatbindingwasto 1 poly(A)-containingfragments of M-MuLV virion less ml of oligo(dT)-cellulose T-3 (Collaborative Re- than1,500nucleotideslong per ml inatotalvolume of search), andwashings and elutionswereperformedin 0.1 ml for5 h at68°Cin 0.6MNETES(0.6MNaCl,
acolumn instead ofbycentrifugation. 0.01 M N-tris(hydroxymethyl)methyl-2-aminometh-Preparationof poly(A)-containing size classes anesulfonic acid [pH
7.5],
1 mM EDTA) plus 0.1% of viral RNA. TheproceduredescribedbyWang et SDS. After annealing, the unhybridized cDNA was al. (34) wasused with slight modifications. Purified selected out bybinding tohydroxylapatite in 0.01 M virionsof M-MuLVwerelysedwithSDS (finalcon- phosphate buffer containing 0.3 M NaCl and 0.1% centration1%), and RNAwasextractedbydeprotein- SDS, followed by elution in 0.14 M phosphate buffer ization withphenol-chloroformandprecipitated with plus 0.1% SDS at 60°C. Approximately 70% of the 2volumes of ethanol. Theprecipitatewaspelleted by cDNAwasrecovered in this fraction.ThecDNAwas centrifugation and dissolved in SDS buffer containing then concentratedby ethanol precipitation and freed 0.1%SDS and sedimentedon 15 to30%linear sucrose ofresidualphosphate by passageoveraSephadex G-gradients(11). Material sedimentingat 60 to70Swas 50column. The cDNA was thenagain concentrated pooled andprecipitatedwithethanol. The RNAwas by ethanolprecipitation and finally stored in hybridi-dissolved in buffercontaining0.01MTris-hydrochlo- zation bufferat-20°C.ride(pH 7.4),0.01M NaCl,and0.001 M EDTA and cDNA 5'. cDNA 5' was synthesized by using puri-incubated at50.5°C for 1 min. A 1.0 M solution of fied M-MuLV virions. The reaction mixture (1.0 ml) sodium carbonatewasaddedtothissolutionto afinal contained50mMTris-hydrochloride (pH 8.3), 10 mM concentration of0.06M(pH 11)and incubated further dithiothreitol,6mMmagnesiumacetate,60mMNaCl,
for 3.5min at 50.5°C.The solution wasneutralized 1 mM each ofdATP, dCTP and dGTP, 50
itM
of withaceticacid,diluted with buffer containing 0.4 M [3H]dTTP (specificactivity of dTTP in the reactionNaCl, 0.01 M Tris-hydrochloride (pH 7.4), 0.001 M was4,900cpm/pmol),80ug ofactinomycinD,Nonidet EDTA, and 0.5%SDS,and adsorbedto anoligo(dT)- P-40toafinal concentration of0.01%, and2.5mg of cellulose (T-3) columnasdescribed (12). The bound virus. The reaction mixturewasflushedwithN2 and material was eluted with buffer containing 0.01 M incubatedat37°Cfor6h.About5 to6% of theinput Tris-hydrochloride (pH7.4),0.001MEDTA,and 0.5% nucleotides of viral RNA (assuming that 70S viral SDS. The salt concentration of the eluted material RNArepresentsabout 1% of total viralprotein)were wasraisedto 0.4MNaCl and the materialwasread- transcribed. The reactionwasstopped byadditionof sorbed to oligo(dT)-cellulose. Material eluting from SDSto afinal concentrationof1%,andnucleic acids the secondcycleofchromatographywasfractionated wereextractedbyphenol-chloroformasdescribed(26) onneutralsucrosegradients alongwith28S and 18S and precipitated with ethanol. The precipitate was rRNA markers. Varioussize classes of RNAranging suspendedinasolutioncontaining0.01M
Tris-hydro-from 4Sto38Swerepooled,theiramounts werede- chloride (pH 7.4) and0.001 MEDTA, and theviral termined bymeasurementofabsorbancy at260nm, RNAwashydrolyzed byaddition of NaOHtoafinal andtheywerestored under ethanolat-20°C.About concentration of0.3N,followedbyincubationat37°C 15%ofthestarting60 to 70S RNA wasrecovered after for 18h. ThecDNAwasneutralized by acetic acidand twocycles ofoligo(dT)-cellulosechromatography. separated from low-molecular-weight material by Preparation of cDNA probes. 3H-labeled M- chromatography on aSephadex G-75 column equili-MuLVcDNAprobe, whichwasapproximately repre- brated with 0.05 M triethyl ammonium bicarbonate. sentativeof the entire M-MuLV 38SRNA, waspre- Thematerialelutingin thevoidvolumewascombined,
paredbyprimingtheendogenousreversetranscriptase lyophilized to dryness, and suspended in 0.3 ml of reaction ofpurifiedM-MuLV virions withexogenously buffercontaining0.01MTris-hydrochloride (pH 7.4) added random oligodeoxynucleotides obtained from and 0.001 M EDTA.Thematerialwastransferredto DNase-digested calfthymus DNA (32). Briefly, the siliconized1.5-ml conicalpolypropylene tubes (Eppen-reaction mixtures were as described previously (11), dorf), and, after addition of LiClto afinal
on November 10, 2019 by guest
http://jvi.asm.org/
tion of 0.2 M and10,ug of carrierRNA, the cDNA was 30 min at 37°C, and a sample was withdrawn to precipitated with 2volumesof ethanol and stored at determine acid-precipitable material. In this case, -70°C for 15 min. The ethanol precipitatewascentri- about 1.2 to 1.5% of the input nucleotides of viral RNA fuged in a Brinkman centrifuge attop speed for 10 were transcribed into cDNA. The reaction mixture min, and the pellet wassuspended in 20
pl
of 0.01 M was boiled in 0.3 N NaOH for 5 min, neutralized withTris-hydrochloridebuffer(pH7.4). HCl, andseparatedfromlow-molecular-weight mate-The cDNA 5' (bands 1 to 7) wasseparated from rial by chromatography on aG-75 column. Material total cDNAbyelectrophoresison10%polyacrylamide eluting in the void volume wascollected and either gels. Included in thesamplewere traceramountsof lyophilizedorprecipitated withethanol.
32P-labeled cDNA 5' bands 1 to 7. The 32P-labeled Analysisof cDNA3'oneitherpolyacrylamidegels cDNA 5'(bands 1 to 7) wasprepared by labeling the oralkalinesucrosegradients revealedtwosize classes. 5' end of the total cDNA with y [32P]ATP and T4 About 30% of the cDNA 3'sedimented with an average polynucleotide kinase. Bands 1to7wereisolated by size of about 150 nucleotides, and the rest of the fractionating the total cDNAon 10%polyacrylamide material sedimented with an average size of about 500 gels, and their nucleotide sequencesweredetermined. nucleotides. Hybridization analysis showed that about Bands2to 7appeartobesubsets of cDNA 5' band1. 30 to 40% of cDNA 3' of small size class could be cDNA 5' band 1 appears to be initiated on tRNA protected bypoly(A),whereasonly about 8 to 10%of primer,asit shows the standarda-32Patomtransfer the large class of cDNA 3' could be protected by fromdAtorA(31). Details of nucleotide sequencing, poly(A). The high degree of hybridization of the small 5'-endlabeling, anda-32P atomtransfer experiments size cDNA 3' topoly(A) suggested that the material will be published elsewhere in collaboration with A. contained largeamounts of polydeoxythymidylic acid
Ohtsukaand M. McKennett. The 3H-labeled cDNA- synthesized during the reaction. In the experiments containing 5' 32P-labeled bands 1 to7 weresubjected reportedhere,only the large size classes of cDNA 3' to electrophoresis inTris-borate buffer (90 mM Tris were used.
base,2.5mMEDTA, and 89 mM boricacid) for1hat Gel analysis and hybridization techniques. Size 200 V (about25V/cm). The wet gel was exposed to analysis by electrophoresis in agarose gels was as X-ray film (Kodak NST 54) for several hours. The described previously (9). Briefly, RNA was suspended film was developed and used as a replica toexcise inelectrophoresis sample buffer (4 mM Tris base[pH
bands1 to 7.Thepolyacrylamide gel slices were dis- 7.2], 2 mM sodiumacetate, 2 mM EDTA, 0.2% SDS, aggregated manually with a spatula to a very fine 10%glycerol) and boiled for 1 min, followed by rapid mesh and incubated at 45°C overnight in a buffer cooling in ice.Bromophenol blue dye and 10% glycerol containing0.01 MTris-hydrochloride (pH 7.4),0.001 wereadded, and the sample was layered onto a 10-cm MEDTA,and0.002MNaCl. Thegel suspensionwas 1% agarose tube gel in Tris-acetate buffer.After elec-filteredthrougha0.45-,ummembranefilter(Millipore trophoresis at the times andvoltages indicated, the gel Corp.). The filtratewasadjustedto afinalconcentra- wasstained with ethidium bromide, and the location tionof 0.2 M LiCl and
20,ug
of carrier yeast RNA per of the 28S and18S rRNA's was determined by obser-ml and precipitated with 2.5 volumes of 95% ethanol. vationunder UV light.TheDNAwascollectedby centrifugationandresus- Hybridization across the agarose gels wasperformed pendedinhybridizationbuffer. essentially as described previously (9). Briefly, the Inatypical experiment, westarted with about12 regions of the gels calculated to contain 15 to 45S mgofpurified virions, and the reaction wascarried RNA were cut out and divided into 1-mmslices. Each outin5.0ml. After6h of incubationat37°C,the total slice (approximately 25
AJ)
was placed in a 1.5-ml amount of cDNA synthesized was 7.2jig
(about 6% plastic tube(Eppendorf), and 25p1
of cDNA (100 toincorporation). Afterfractionationonpolyacrylamide 1,000 cpm) in 0.6 M NETES plus SDS (0.6 M NaCl,
gels,theamountsofcDNA 5' bands(1to7)recovered 0.01 M N-tris(hydroxymethyl)methyl-2-aminometh-were as follows: band 1,0.034,ug (about0.5% of the anesulfonic acid [pH 7.5], 1 mM EDTA, 0.1% SDS) totalcDNA);bands2and3,0.014ug
(0.19%);
band4, was added. After addition of 0.1 ml of mineral oil, the 0.0078jig
(0.1%);bands5to7,0.008,ug(0.1%). tubes were sealed and boiled for 3min. The tubes were The cDNA 3' was synthesized by using purified thenimmediately transferred to a 68°C water bath, viral 70S RNA,oligo(dT) primer,andpurifiedreverse andannealing wasperformed for the times indicated. transcriptase from avian myeloblastosis virus. The Afterannealing, the samples were digested withSi
reaction mixture in 0.1 ml contained 50 mM Tris- single-strand-specific nuclease, and trichloroacetic
hydrochloride (pH 7.4), 10mMdithiothreitol, 6mM acid-precipitable radioactivity was determined as
de-Mg2+,15mMNaCl,40
,uM
each ofdATP,dCTP,and scribed before (9).dTTP, 20 gM of a-[32P]dGTP (specific activity of Solution hybridizations were performed in micro-dGTPinthereactionwas12,000cpm/pmol),10,g of capillarypipettesasdescribedpreviously (11). Reac-actinomycin D, 75 U of avian myeloblastosis virus tion volumeswere 5 to10
pl,
andhybrid assay was by reversetranscriptase (1 U=incorporation of100pmol digestionwithS1 nuclease.of dGMPin 15minat37°C),0.5,ug ofoligo(dT)1o,and 10
,ug
ofviralRNA.The viral70S RNAwaspreparedfrompurified virions byphenol-chloroformextraction RESULTS asdescribed above. The RNAwasboiledin 0.01 M
Tris-hydrochloride (pH 7.4) and 0.005 MNaCl for 2
Identification
of virus-specific mRNA. min and selected forpoly(A)-containingRNAbychro-Previous
sLze
measurements
ofvirus-specific
matography on oligo(dT)-cellulose columns as de- mRNAinMuLV-infectedcellsweremadebased
scribedabove. The reaction mixture was incubated for onsedimentationinsucrose
gradients,
sinceon November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 SIZE ANALYSIS OF MuLV-SPECIFIC mRNA'S
471
sequent
analysis
wasrelatively simple (11, 15).
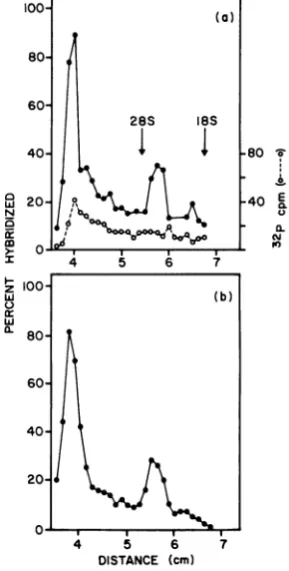
100- ) However, the size resolution of suchgradients
was rather limited. Therefore we performed a
80-more refined sizeanalysis usingelectrophoresis
inagarose gels. Purified polyribosomes were
pre-pared
from M-MuLV clone 4A cells (a line of60-NIH-3T3 cells clonally infected with M-MuLV), 28S
1BS
and extracted RNA wasseparated according to 40-
-80
S'size by
electrophoresis in an agarose gel. It haspreviously
been
shown that
essentially all virus-
a 20R
40specific RNA in
apurified polyribosome
prepa- Nration
isfunctionally mRNA, although
intotal
, Rcytoplasm it represents a minority component 0 K
l,
(13, 15).After
electrophoresis, the gel
wasfrac-
I 6 7tionated
into 1-mmslices, and the
amountof
00- (b)virus-specific RNA in each slice
wasdetermined
by
annealing with radioactively labeled
cDNA a.80-prepared
by the endogenous
reversetranscrip-tase
reaction of M-MuLV
virions. InFig.
1the
60 lM-MuLV
cDNA wasprepared by incubation of
60M-MuLV
virions inthe
presence of randomcalf
thymus deoxynucleotide primer; such cDNA
40-contains nucleotide
sequencescomplementary
to all
regions
of the M-MuLV 38S RNA
in 20approximately equal
concentrations
(32). The
20hybridization
of the
gel
slices indicated
twoma-jor
virus-specific
mRNA
species (Fig.
la).
The
0o
larger mRNA
comigrated
with
32P-labeled
M- 4 56
7
MuLV
38S
virionRNA and
wasverynear, if
not .DISTANCE (cm)
identical,in
size'to38Svo
FIG. 1. Size analysis ofM-MuLV-specific
mRNA.identical,
i szeto 38Sviron RNA,asreported (a) Purified polyribosomes from 1015-cm tissuecul-previously (11). The smaller mRNA migrated ture dishes of M-MuL V clone 4A cells at three-quar-between28S and 18SrRNA,and wasdesignated ters confluency were prepared, and the RNA was
24S
(according
toits sedimentation
in aneutral
extracted. One twentieth of the RNA was combinedsucrosegradient) althoughitsapparentmobility withapproximately1,OOO) cpm of32P-labeledM-MuLV in the agarose gelwas somewhat larger. Other 38Svirion RNA, denaturedby boiling, and
layered
investigators have reported that even smaller onto a 1% agarosegel. Electrophoresis was for 165virus-specific
mRNAmight
also be present in mm at 65 V/cm. After staining with ethidium bro-MuLV-infected cells(15),
but no evidence for mide to visualize the rRNA, the region of thegel suchmRNA'swasfound in the results shown in indicatedwascut outanddivided into 1-mmslices.Fgorsbeunexet.
HThe
'P
radioactivity in
eachslice
wasdetermined
byFig. 1 or subsequent
experlments.
owever,
an Cerenkovcounting, and then each slice wasannealed mRNA smaller than 105 would not have been with 850 cpm of 3H-labeled calf thymusDNA-primeddetected. M-MuLV cDNA probe. Annealing was for 15 h at
Fig. lb
shows that bothvirus-specific
mRNA's 68°C. After annealing, the amount of cDNAhybrid-contain
poly(A)
sequencesasmeasuredby
bind- ized in eachsample
wasdeterminedby digestion of
ing
tooligo(dT)-cellulose.
Both mRNA's bound unhybridized cDNA with Sl nuclease. The locationto the
oligo(dT)-cellulose equally
well
(better
of the 28S and 18S rRNA is indicated in the figure.
than 75%). If the
24S
virus-specific RNA was()
32P
radioactivity;(0)
percentage of3H-labeled produced by artifactual degradation of 38S M-MuLV cDNApolyribosomal hybridized.(b)
One fifteenth of the RNApreparation
described in(a) wasmRNA,
then not all 24S mRNA molecules bound tooligo(dT)-cellulose,
andthepoly(A)-contain-would
containpoly(A).
The fact thatboth
ingRNA was eluted and analyzed in a parallelgel.
mRNA's boundequally
well to theoligo(dT)-
No 32P-labeled385
RNA was added, and only thecellulose indicates that 24S virus-specific
mRNA hybridization with3H-labeled
M-MuLV cDNA is didnotariseby degradation. Furthermore,
since shown. Conditionsof
annealingwerethesame as(a).poly(A)
sequences arelocated
atthe 3' ends ofvirion RNA
(4,
33), aswell
asmany hostcellular tionof 38S RNA might also bepolyadenylated.
mRNA's,
theseresults suggest that the24S
vi-Relationship of the 24S and 38S
virus-rus-specific
mRNAmight
bederived from the 3'specific mRNA's.
To investigate therelation-portion
of the 38S viral RNA.However,
it is ship of the 24S and 38S virus-specificmRNA's,
possible
that sequences derived from the 5'por- acDNA specific for the 5' half of the M-MuLVon November 10, 2019 by guest
http://jvi.asm.org/
[image:4.509.281.426.74.360.2]472
VERMA J. VIROL.genome was
prepared,
asdescribed
in detail in100
Materials and Methods.
Briefly,
acDNA
probecDNACT
(a)was
prepared from
Moloney
murine
sarcoma)0
virus
clone
124,which has been shown
tohave 80 sequencehomology with the 5' half of the
M-MuLV
genome, aswell
as somesequences atthe
60-
l2
3'
end
(8,
20). The
sequencescomplementary
to 2the
3'end of the M-MuLV
genome were re-40-moved from the murine
sarcomavirus
cDNA byannealing
with short
poly(A)-containing
3'
frag-
a20-
oments
of M-MuLV virion
RNA,
yielding
acDNA
Nprobe that
would
only recognize
sequences cor-0
responding
tothe 5' half of M-MuLV RNA.
2 34
5 6 7Heteroduplex
analysis and protein analysis in-
100-dicate that the
gag geneis 5' terminal for M-
cDNAuggpoV
(b)MuLV
(20), similar
tothe
geneorder
deduced
S80-for avian RNA
tumorviruses
(gag
pol
env srcfrom 5'
to3')
(33).
Therefore,
the cDNA
probe
60-described above
wasdesignated
cDNAg<g,,o)
to28S
18Sindicate that it likely contained
sequencesfrom
24S
the
gagand
possibly
aportion
of
the
pol
geneof
404mRNA4
M-MuLV. When
cDNAgog(p,o
wasannealed with
fractions
from
agel of M-MuLV clone 4A poly-
20-ribosomal RNA,
hybridization
of the 38S
virus-specific mRNA
wasobserved, but
nohybridiza-
0otion
by the
24Svirus-specific
mRNA
wasde-
2
3 4 56
7
tected
(Fig.
2b).
Some
annealing with the
DISTANCE (cm)
cDNAg.g(po,
was observed intheregion
of the FIG. 2. Size analysis of M-MuLV-specific mRNAgel
corresponding
tosubgenomic
RNA; this
may with specific cDNA. (a) One tenth of the clone 4Ahavebeen due todegradd38Sv polyribosomal RNA
preparation
from Fig. 1 washRNA boleencdulestohegpradedara38 virunpecic
layered
ontoanagarosegel,
andelectrophoresis
wasmRNA moleculesinthepreparation, since this performed for 105
min
at 10V/cm.
Analysis with 3H-RNA was not selected on oligo(dT)-cellulose.labeled
calfthymus DNA-primed M-MuLV cDNA Forcomparison, annealing
across agel
run in (1,600 cpm per sample) wasperformed
as in Fig. 1. (b)parallel using
calfthymus-primed
cDNAprobe
Anequal
amountof polyribosomal
RNA wassepa-is shown
inFig. 2a;
the24S
M-MuLV-specific
rated by electrophoresis in a gelparallel to thatmRNA
isreadily
evident. Theseresults indicate
shown in (a). The gelfractions wereannealed withthat the 24S
virus-specific
mRNAis
notderived
3H-labeledcDNAgagpo
(375 cpm per sample). Thefrom the 5' half of M-MuLV 38S
RNA,
but location of the24S
virus-specific mRNA is indicated;rather from the 3' half. It is therefore
likely
thatno
specific annealing
of
the 24S mRNAwasevident.the
24Svirus-specific
mRNA contains
sequencesfor
only
the
env gene, andadditionally
somefrom the
5'end
of the38S
RNA, and that sequences whichdo
notcode for viral
structural
transcriptionproceeds toward the 5' end of theproteins.
viral
RNA (31). In conditions oflimiting
deox-Presence
of
transposed
5'
sequencesin
ynucleoside
triphosphate precursor, reverse24S
virus-specific
mRNA. Recent results
transcription proceeds through
a series ofhesi-studying
mRNA's from avian
sarcomavirus-in-
tations,
with
a stronghesitation
as the DNAfectedcells indicate that
sequences
correspond-
polymerase
reachesthe5'
end of the38S
RNAing
tothe
extreme5'endof the 38S viral
genome(17). The product of such
areaction is a series ofarepresenton
mRNA's
containing
3'portions
of short DNAmolecules
with overlapping se-the38SRNA
(35).
The24Svirus-specific
mRNA quences,which
contain the same 5' end and fromM-MuLV clone
4Acellswasfurther inves-progressively
moresequences complementary totigated
todetermine whether
asimilarphenom-
theRNA between the tRNAprimer binding
siteenonwas
evident.
and the 5' end of the genome. For the experimentTo
perform
theseexperiments,
cDNA corre-reported here,
cDNA wasprepared
fromM-sponding
totheextreme5'end ofthe
M-MuLV MuLV virions inconditions where
one triphos-38S virion RNA wassynthesized.
It has been phate waslimiting
and then separated on a shownthat
the site ofbinding
forthe
tRNApreparative
polyacrylamide
gel.
Individual
primer molecule
used in reversetranscription
of cDNAbandscorresponding
todifferent lengths RNAtumorvirus RNA is100 to 150nucleotides of cDNA5' were cutoutandeluted,
andon November 10, 2019 by guest
http://jvi.asm.org/
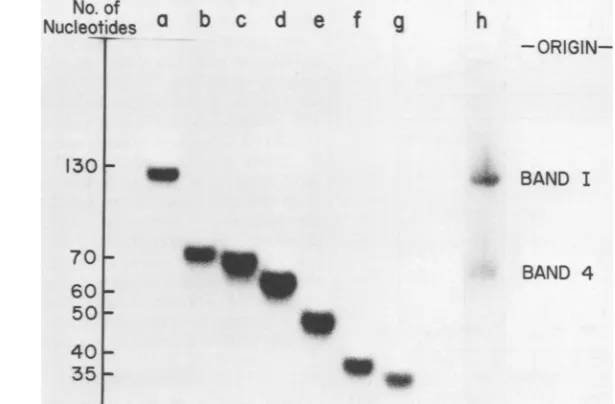
[image:5.509.294.441.63.332.2]VOL. 26, 1978
SIZE ANALYSIS OF MuLV-SPECIFIC mRNA'S
473
acterization
of the bands is shown in Fig.
3.fore
possible
that the
annealing
with cDNA
5'
Seven discrete bands,
varying inlength from 35band
1 was atleast
partially
due
tothese
re-to 135nucleotides (the full length between
thepeated
sequences atthe 3' end of the 24S
mRNA.
tRNA
primer binding site and
the 5' end of M-To
testthis
possibility,
annealing
wasalso
per-MuLV 38S RNA)
wereobtained, and the
lengthformed with cDNA 5' band 4 (approximately
60of each
fragment
innucleotides
is indicated innucleotides). The cDNA 5' band
4is
approxi-the
figure. DNA sequencing of these
bands hasmately 60
to70 nucleotides shorter than cDNA
confirmed that they
represent overlapping se-5' band
1,and therefore
maylack
some orallof
quences(I. M. Verma,
A. Ohtsuka, and M. the sequencesthat
arecomplementary
tothe
McKennett, manuscript in preparation).
terminally
redundant 3' RNA
sequences.Fig.
5When
cDNA5'
band
1(approximately
130shows
annealing
with cDNA 5'
band
4,which
nucleotides
inlength)
wasannealed with
frac-also indicates that
complementary sequences
tions from
agel
of M-MuLV clone
4Apolyribo-
werepresent in both
38S and 24S
virus-specific
somal RNA, both 38S and 24S
virus-specificmRNA.
Therefore, it is unlikely that the
an-mRNA's
effectively
annealed the cDNA,
evennealing observed with the cDNA 5'
preparations
though the 24S virus-specific
mRNA was de- wasexclusively
due
toterminally repeated
se-rived from the 3' end of
38S viral
RNA (Fig. 4). quencesatthe 3'
end.
These results
suggestthat
nucleotide
sequencesSince
sequencessimilar
oridentical
tothose
similar oridentical
tothose found
at the extremefound
atthe
extreme5' end of viral 38S RNA
5'
end of M-MuLV 38S RNA
arealso
present in were present inintracellular 24S
virus-specific
intracellular 24S virus-specific
mRNA.mRNA,
wetested
whether similar
sequences areIt
has been
recently demonstrated
that both presentin the
samelocation in 38S virion RNA.
avian and murine RNA
tumor virus 38S RNAsPartially
degraded
RNA
wasextracted from
M-contain
sequences atthe 3' end that
areidenticalMuLV
virions,
and the
portions covalently
at-tothose
atthe 5' end. In
the
caseof
aviantached the 3'
poly(A)
sequences wereselected
viruses, the terminal repetition
is approximatelyby
binding
tooligo(dT)-cellulose.
The
poly(A)-21nucleotides, whereas the repetition in MuLV
containing fragments
werethen sedimented in
amay
be
longer (18, 28, 30; J. Coffin and W.
sucrosegradient, and
twosize classes of
mole-Haseltine, personal communication). It is there-
cules, corresponding
tothe 400 to 600
nucleo-No.
f
b
d
f
h
Nucleotides
0
-ORIGIN-130 _
AND I
70__
70_
-BAND
4
60
-50
-40
-35
-FIG. 3. Polyacrylamide gel electrophoretic patterns of cDNA 5' bands1to7. (a-g) Electrophoretic mobility
ofisolated cDNA 5' bands1to 7,respectively. (h)Agel where 3H-labeled cDNA 5'wasmixed with 32P end-labeled cDNA 5' bands 1 and4. The excised bands contained both 32P marker cDNA and 3H-labeled experimentalcDNA. The size of thecDNA5' bands 1 to 7 wasdeterminedbyusing restriction endonuclease HaeIII-digested
4X-174
single-strandedDNAasmarker. Theentire nucleotide sequence ofqLX-1
74 DNA has beendetermined,and hence themarkersgivethe exact numberofnucleotides. The HaeIII-digestedXX-174 DNA wasvisualizedby stainingwith ethidiumbromide.Furthermore, the complete nucleotide sequence of bands2to7hasbeendetermined(Verma,Ohtsuka,andMcKennett, manuscriptinpreparation).on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.509.89.397.375.577.2]100- tides and the 4,000 to6,000nucleotides nearest
cDNA5
(D
to the 3'end, werepooled
foranalysis. To deter-minetotal virus-specific
RNA concentration in80- these
preparations,
annealing was performed* .
with
cDNAcomplementary
tothe
3'end of
M-A1\ MuLV RNA(cDNA
3').
ThecDNA 3'wassyn-co 60- 28S 18S
thesized
by using
denatured 70S viral
RNA asI it1 l l
template,
anoligo(dT)
primer,
and
exogenously
z l l il
added
avianmyeloblastosis
virus reversetran-w
-u 40- scriptase. Sequences
complementary
to cDNAa. l 3' would be present in each
poly(A)-containing
I \ ,. ;'! a
M-MuLV RNA
fragment.
Anappropriate
dilu-20- tion of 400- to 600-nucleotide 3' M-MuLVRNA
/T
'/'wXfragment could
completely anneal
the cDNA 3',0___________________________________ .
.but
extremely
little
of the cDNA 5'(bands
2and2 4 6 3 or band 4) couldanneal to the same
concen-DISTANCE (cm) tration of RNAfragment (Fig. 6). It should be FIG. 4. Annealing of M-MuLV-specific mRNA notedthat cDNA 3' (approximately 500 nucleo-with cDNA 5'. An equal amount ofpolyribosomal
tides)
waslarger
than the cDNA 5'preparations,
RNA described inFig.2 wasanalyzedinparallelto sothat in fact the cDNA 5'would anneal four-thegels of Fig. 2, exceptthatannealingwaswith3H- to eight-fold slower than the cDNA 3' if3'and
labeled cDNA 5' band1 (160cpmper
sample).
An- 5' sequences were present in the same concen-nealing andprocessingwereasinFig.2. tration (36). However, an even greater difference inannealing
rate for the two cDNA's was ob-100-cDNACT
(a) served in Fig. 6a, and it can be concluded that less than 10% of the 400- to 600-nucleotide 3'80- virion RNAfragments also contained sequences
complementary to cDNA 5'. The results ofthis
60-
figure
alsosupport
the conclusion that the28S 18S cDNA 5' fragments (bands 2, 3, and 4) do not 40-
recognize
the
terminally
repeated
sequences atthe 3' end of 38S virion RNA.
20-< t An
experiment
similar toFig.
6a wasalso
20
performed
with the4,000-
to6,000-nucleotide
a
0 poly(A)-containing M-MuLV fragments (Fig.
ta
0-
5 6 7 8 96b).
Again,
it canbe concluded
that lessthan
10% of the virion RNA molecules of
thislength
;-100-z cDNA54 (b)
contained
sequencescomplementary
to cDNAir 5'. These
poly(A)-containing
M-MuLV virionX80- RNA
fragments
were, infact,
aslarge
orlarger
than
the24S intracellular
mRNA.Therefore,
60- 28S 18S the presence of sequencescomplementary
to I X 1 cDNA 5' in intracellular24S
virus-specific
40- mRNA cannot be
explained by
thepresence ofthesesequences in a similar location in M-MuLV
20- virion RNA. Rather, it appears that these
se-quences must be
transposed
ontothe 24Svirus-0
specific
mRNAduring synthesis
andprocessing.
5 6 7 8 9
DISTANCE (cm)
DISCUSSION
FIG. 5. AnnealingofM-MuLV-specificRNA with Two major
M-MuLV-specific
mRNA's. shortcDNA 5'.(a) One tenthoftheclone 4Apolyri- Theseexperiments
indicate that there are twobosomalRNA
preparation of Fig.
1waslayered
ontomajor
virus-specific
mRNA's in cels infected anagarosegel, andelectrophoresiswasfor2.5hat majovirus-speifimRnasin cellsifecte
6.5V/cm. Thegelwasprocessedand annealed with w3H-labeledcalf
thymus-primed
M-MuLVcDNA(950 and 24S. The 38Svirus-specific mRNA, whichcpm per sample)asabove, exceptthatannealingwas is extremely similar if not identical in size to
for16.5 h.(b)Agelparalleltothatof(a)wasanalyzed virion 38S RNA, contains the nucleotide se-withcDNA 5' band 4(110 cpm per sample). quencesfor all three M-MuLV structuralgenes.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.509.63.255.58.246.2]VOL. 26, 1978 SIZE ANALYSIS OF MuLV-SPECIFIC mRNA'S
475
0.4 - 0.6 Kb
poly
A+(a)
Several lines of evidence indicate that the
38S
0-
virus-specific
RNA is
responsible
for
synthesis
cDNA 5
89
of
the
internal
structural
proteins (products
of
cDNA5sE)
the gag
gene). Immunoprecipitation
of
polyri-CDNA\ED
bosomes with antiserum
monospecific for the
internal structural protein p30 selectively
en-z 50-
riches the
38S
virus-specific mRNA (23).
Fur-0
thermore, in vitro translation
experiments of
intracellular mRNA
aswell
asisolated virion
N \ RNA indicate that
38S
viral RNA can code forthe
large-molecular-weight
precursors
(pr78
and
100- \cDNA3' pr65) of the internal structural proteins, while
RNAs
ofsmaller
size do not efficiently code forthese
proteins
(10, 16).
The
experiments
re-_
4-6 Kb polyA+
(b)
ported here also support this notion, since the
x
24S
virus-specific
mRNA
lacks
sequencesof the
>~cDNA5a() Ogag
(and
possiblypol)
gene.Thus the 38S
virus-clNAV
specific
mRNA appears to be the onlyvirus-z
'-cDNA5s
specific
mRNA
that
cancode for the
internal
o
structural
proteins.
W 50
*
Although the 38S virus-specific mRNA also
contains the
sequencesfor
the
env gene, it islikely
that these sequences are not translated in
the 38S
virus-specific
mRNA. This is because in
vitro
translation studies of
intracellular
RNA
100-
-
cDNA3indicate that
38S
virus-specific RNA directs the
l0
l
synthesis of the gag
gene-related
precursor
0O
100
000o
polyproteins,
but
nosynthesis
of
envelope
gly-TIME (min.)
10coprotein
polypeptides
occursfrom RNA of this
TIME
(min.)
size(16,
25).
Thishypothesis
canbe
directly
FIG. 6. Annealing of cDNA 5' with3'
fragments Oftested
by analyzing nascent chains of polyribo-virion RNA.(a)
Anappropriate dilution ofM-MuLVsomes
immunoprecipitated
withanti-p30
anti-virionpoly(A)-containing
fragments, 400to600nu-serum,
orby
determining
the size of virus-spe-cleotideslong,
which could anneal'P-labeled
cDNA cificmRNA frompolyribosomes
immunoprecip-3',
wasempirically found.
This concentrationcould itated withanti-glycoprotein
antiserum. be calculatedto beequivalent
toapproximately
2.5iLg
offull-length
38SRNAper
ml(from
thetll2
value).
The 24Svirus-specific
mRNA islikely
trans-3P-labeled
cDNA3'(360 cpm per reaction)
was an- lated togive
theenvelope glycoproteins,
since nealedwith the RNAfor different lengths of
time. 3H-the
only
M-MuLV structural gene contained is
labeled cDNA 5' bands 2 and 3(140 cpm per reaction)
probably the env gene. In addition, in vitro
was alsoannealed with the same concentrationof
thetranslation of intracellular RNA from
Rauscher
RNA. Percentage of
maximalannealing
is shown asMuLV-infected
cells indicated that envelope
gly-a
function of
incubation at theannealing
tempera-
coproteins are coded by an mRNA of
approxi-ture.Maximnal annealing forthe cDNA 3'was51% a
and
for
the cDNA 5' was 55%. At the maximummately
this
slze
(16). Cellfractionation
experi-annealing
time,
cDNA5' band 4(80 cpm)
was alsoments
also support this
hypothesis,since
mem-annealed withthe RNA.(0)
Annealing of
cDNA3';
brane-bound polyribosomes, which are
respon-(0) annealing of
cDNA 5' bands 2 and3; (A)
an- sible for thesynthesis
of membraneglycopro-nealing of
cDNA 5' band 4.(b)
Anexperiment
similarteins,
are enriched for the 24Svirus-specific
to that describedin(a) wasperformed, usingpoly(A)-
mRNA (15). It is also possible that additional
containing
M-MuLVvirion RNAfragments of 4,000
nonstructural viral proteins coded by the
24S
to
6,000
nucleotidesinlength.
virus-specific mRNA might also be synthesized,
but no suchproteins
haveyet
beenidentified.
The24Svirus-specific
mRNA ispredominantly
Athirdvirus-specific
mRNAlikely
tocode forderived
from the 3'end
ofthe
38S viralRNA,
theviral
reversetranscriptase
wasnotidentified
and
likely
contains sequences
forthe envgene
in theseexperiments.
The
amount of reverseas
well
as3'RNAsequences which
donotcodetranscriptase synthesized
ininfected
cells
is ap-for known virusstructural
proteins.
Sequences
proximately
10-fold less
than theinternal
struc-transposed
from theextreme5'end
of38S
viriontural
proteins
orenvelope glycoproteins,
and it RNA arealso present
in the 24Svirus-specific
is therefore
likely
thataputative
mRNAcoding
mRNA. forreverse
transcriptase
alone
wouldbe present
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.509.55.248.58.368.2]476
in an
approximately 10-fold lower
amountthanmRNA's.
In theseviral
RNAs it has been showneither the 38S
or24Svirus-specific mRNA's. An
that shortnucleotide
sequences arepresent atmRNA
present atthese levels would
very pos- the5'
endsof the mRNA's
which are derivedsibly
havebeen missed in these
experiments,
fromregionsof the viral
genomephysically
sep-especially if its size
wereveryclose
tothe size of
aratefromthe region coding
themRNA, and it oneof the
major
virus-specific mRNA's. There-
has beenproposed that these sequences may before these
experiments
cannotbe taken
asevi-
transposed by RNAligation
during processingdence that
aunique mRNA coding only for
of viral RNA
molecules into functional
mRNA reversetranscriptase does
notexist.
Analterna-
(1, 5, 6, 22). The results reported here suggesttive
explanation of the mechanism of synthesis
that
asimilar situation
may exist for M-MuLVof
reversetranscriptase has been
suggested by
24Svirus-specific mRNA, although the locationthe
finding of
small amountsof
a verylarge
of the
transposed
5'sequencesin
the 24S
RNAmolecular
weight (approximately 180,000)
pro- has not yetbeen identified.
It is interesting totein, pr180, with
antigenic determinants for both
notethat
alkali
degradation of 38S virion
RNAinternal structural
proteins and
reverse tran- tothe
samesize as24S intracellular
mRNA doesscriptase
(7,
21).
Ithas been
proposed that pr180
notproduce
an mRNAactive
intranslation
ofarises
by
read-through
translation of the
gagthe 3'
terminal
env gene (K. Beemon and T. geneinto the pol
gene,the
prl80
protein being
Hunter,personal communication).
It may becleaved
toproduce the
reversetranscriptase.
that
transposition of the 5'
sequences to the24S
Recentevidence
supporting this
hypothesis has
virus-specific
mRNA may be necessary foracti-been
reported in which increased
amountsof the
vation of the initiation site for
envgenetrans-prl80
protein
weresynthesized
in
vitro if
ayeast lation.amber
suppressortRNA
wasadded
tothe
reac-Quantification of mRNA's. In
the gelhy-tion mixture
(L. Phillipson, P. Andersson, R.
bridization
experiments shown here,
data wereWeinberg,
D.Baltimore,
andR.
Gesteland, sub-
presented
as percentageof cDNA hybridized. Inmitted for
publication). Synthesis
of the prl80
previous
reports,these
values
wereconverted
toprotein would
notrequire another virus-specific
relative
virus-specific RNA
concentration (11).mRNA in addition
to38S
mRNA,
sothat
only
Forthis
conversion
tobe
performed, it
was nec-twovirus-specific mRNA's could
accountfor
essary to know themaximum
hybridization level
synthesis of all of
theviral structural
proteins.
of
cDNA for eachRNA
sample;
in sucrose gra-However, ithas
still
notbeen
rigorously
estab-
dientanalyses,
everyfraction maximally
hybrid-lished that
prl80
is the in vivo
precursorfor
ized the cDNA
probe. However,
in
the
higher-biologically
active
reversetranscriptase.
resolution
experiments
reported here, it is likely
We
recently proposed
amodel
ofM-MuLV-
thatthe
relatively
pure24S virus-specific
specific mRNA
organization
and function in
mRNA could
notmaximally hybridize
arela-which the
virus-specific
mRNA's consisted of
tively
representative calf thymus-primed
cDNA.three
(or
possibly only
two) molecules with
se-Therefore,
conversion
torelative
virus-specific
quencesoverlapping
from the
3'end
(24).
In RNAconcentration could
notbe
readily
per-each
case,only
the
5'terminal
genewould
beformed. The
hybridization
of the
gel with
cDNAactive
inprotein
synthesis,
and theinternal
5'band
4 inFig. 5b
wasnoteworthy
inthis
geneswould
notbe
translated. The results
pre- respect.The
cDNA 5'represented only
about 60sented
here,
showing
that the 24S mRNA
rep-nucleotides,
whichareperhaps completely
pres-resents sequencesfrom the
3'end of
the
M- entin both 24Sand 38S
virus-specific mRNA.
MuLV genome,
support thismodel.
Asimilar
Therefore,
therelative concentrations of the
twoorganization
forthe
virus-specific
mRNA's
inmRNA's could be inferred from the levels of
avian
sarcoma viruses hasalso
recently
beenhybridization
in the 38S and 24Speaks. Thus,
proposed (35).
Fig.
5b indicates that the concentrations of 38STransposition of
RNA sequences in24S
and 24Svirus-specific
mRNA inpurified
poly-mRNA. Our results indicate
that sequencessim-
ribosomes
areapproximately equal.
Thisresult
ilar oridentical
tothoseatthe
extreme5'end
of was notimmediately
obvious from thegel
hy-theviral 38S
RNAarepresent in the24S
virus-bridization
ofFig.
5a, in which calfthymus-specific
mRNA,
similar toresultsreported
for primed cDNAwasused. This result is in agree-avian RNA tumor viruses. The data ofFig.
6 ment with the observation thatthe
amountoffuthermore
indicate thatequivalent
sequencessynthesis
of gag and envrelatedprotein
is ap-arenotpresent in the 3'half
of38S
virion RNA.proximately
equal,
asmeasured
by
pulse-label-Thepresence of
RNA
sequences fromnoncon-ing
andimmunoprecipitating
viralproteins
fromtiguous
regions
of the viral genome in 24S virus-infected
cells(S.
Edwards,
unpublished
obser-specific
mRNA may besimilar
tothesituationvation).
observed
for adenovirus andpapovavirus
Results similar to these have also beenon November 10, 2019 by guest
http://jvi.asm.org/
VOL. 26, 1978 SIZE ANALYSIS OF MuLV-SPECIFIC mRNA'S
477
tained
by
Rothenberg
etal.
(E.
Rothenberg,
D. sequence complexityof cloned Moloney murineleuke-J.
J.
Dngu,and D. Baltimore, Cell, in press). miavirus60 to708
RNA: evidence forahaploid ge-Donoghue,and D.
Baltimore,
Cell, in
press).
nome. J.Virol.14:421-429.In
addition, these workers have obtained elec-
15.Gielkens,
A.L J.,M. H.L.
Salden,and H.
Bloemen-tron
microscopic
heteroduplex
evidence for the
dal. 1974.Virus-specificmessengerRNA onfreeandtransposition
of virion RNA 5' sequences in membrane-bound polyribosomes from cells infected subgenomicvirus-specificpolyAcontainwith
Rauscher leukemia virus. Proc. Natl. Acad. Sci.suRgenomic
virus-specic
poly(A)-containing
U.S.A. 71:1093-1097.RNA. 16.Gielkens, A. L. J., D. van Zaane, H. P. J.Bloemers,
ACKNOWLEDGMENTS and H. Bloemendal. 1976. Synthesis of Rauscher mu-rineleukemiavirus-specificpolypeptides in vitro. Proc. Wethank PaulMacIsaac, Marianne McKennett, and Rich- Natl. Acad. Sci. U.S.A.73:356-360.
ard Swanson for excellent technicalassistance. Mei-Huei Lai 17.Haseltine, W. H., D. G.Kleid,A.Panet, E. Rothen-and DavidDolberg kindlyprovided purified reversetranscrip- berg, and D.Baltimore. 1976. Orderedtranscription tase and Moloney murine sarcoma virus cDNA. We thank ofRNA tumor virusgenomes. J. Mol. Biol.106:109-131. CarolynGoller fortypingthemanuscript,and themembers of 18. Haseltine, W. A., A. M. Maxam, and W.Gilbert.1977. the TumorVirologyLaboratoryforsuggestions and discus- The Roussarcoma virusgenome isterminally
redun-sion. dant: the 5' sequence. Proc. Natl. Acad. Sci. U.S.A.
This workwassupportedbyresearch grants no.CA-16561 74:989-993.
andCA-21408toI.M.V. from theNational Cancer Institute, 19. Hayward, W. S. 1977. Size and genetic content of viral grantno.CA-15747toH.F. from theNational CancerInstitute, RNAs in avian oncovirus-infected cells. J. Virol. and core grantno. 14195from theNationalCancerInstitute. 24:47-63.
LITERATURE CITED 20. Hu,S.,N.Davidson,and I. M.Verma.1977.A hetero-duplexstudy of the sequencerelationships between the 1.Aloni, Y.,R. Dhar, 0. Laub, M. Horowitz, and G. RNAs ofM-MSV and M-MLV. Cell10:469-477.
Khoury.1977.Novel mechanism for RNAmaturation: 21. Jamjoom,G.A., R. B. Naso, and R. B.Arlinghaus. theleader sequences of simian virus40mRNA are not 1977.Furthercharacterizationofintracellular precursor transcribed adjacent to the coding sequences. Proc. polyproteins of Rauscher leukemia virus. Virology Natl. Acad. Sci.U.S.A.74:3686-3690. 78:11-34.
2. Ball, J.K., J. A.McCarter, andS. M. Sunderland. 22. Klessig, D. F. 1977. Two adenovirus mRNA's have a 1973.Evidence forhelper independentmurine sarcoma common 5'terminal leader sequence encoded at least virus.1.Segregationofreplication-defectiveandtrans- 10 kB upstream from their main coding regions. Cell formation-defective viruses.Virology56:268-284. 12:9-22.
3. Baltimore, D. 1974.Tumor viruses: 1974. ColdSpring 23. Mueller-Lantzsch,N., and H.Fan. 1976. Monospecific HarborSymp. Quant.Biol. 39:1187-1200. immunoprecipitation ofmurineleukemia virus polyri-4. Beemon, K. 1977. Oligonucleotide fingerprinting with bosomes:identification ofp30protein-specific
messen-RNAtumorvirus RNA. Curr.Top.Microbiol.Immu- gerRNA. Cell9:579-588.
nol.76:73-110. 24. Mueller-Lantzsch, N., L. Hatlen,and H. Fan. 1976.
5. Berget, S.M.,C.Moore,and P.A.Sharp.1977.Spliced Immunoprecipitationof murineleukemiavirus-specific segmentsatthe 5'terminus ofadenovirus2late mRNA. polyribosomes: identificationofvirus-specific messen-Proc. Natl.Acad. Sci. U.S.A. 74:3171-3175. gerRNA, p. 37-53. In D.Baltimore, A. S. Huang, and 6. Chow, L. T.,R.E.Gelinas, T. R.Broker, and R. J. C.F.Fox(ed.),Animalvirology. Academic PressInc.,
Roberts. 1977.Anamazing sequence arrangementat NewYork.
the5'ends ofadenovirus2messenger RNA. Cell12:1-8. 25. Pawson,T.,G. S.Martin,and A. E.Smith. 1976. Cell-7.Dina, D.,and K.Beemon. 1977.Relationship between free translation of virion RNA fromnondefectiveand Moloney murine leukemia and sarcomavirus RNAs: transformation-defective Roussarcoma viruses. J. Vi-purification andhybridization map ofcomplementary rol.19:950-967.
DNAsfrom definedregionsofMoloneymurine sarcoma 26. Penman, S. 1966. RNA metabolism in the HeLa cell virus124.J. Virol. 23:524-532. nucleus. J. Mol. Biol. 17:117-130.
8. Dina,D.,K.Beemon,and P. H.Duesberg.1976.The 27. Schincariol, A.L,and W. K. Joklik. 1973.Early syn-30S Moloney sarcoma virus RNA containsleukemia thesis ofvirus-specific RNA and DNA incellsrapidly virusnucleotide sequences. Cell 9:229-309. transformed with Rous sarcoma virus. Virology 9. Fan,H.1977.RNA metabolism of murineleukemiavirus: 66:532-548.
size analysis of nuclear pulse-labeled virus-specific 28. Schwartz, D. E., P. C. Zamecnik, and H. L Weith. RNA. Cell 11:297-305. 1977.Rous sarcoma virus genome isterminally redun-10. Fan,H.1977.Expressionof RNAtumorvirusesattrans- dant: the 5' sequence. Proc. Natl. Acad. Sci. U.S.A.
lation and transcriptionlevels. Curr. Top. Microbiol. 74:994-998.
Immunol.79:1-41. 29. Shanmugam, G.,S.Bhaduri,and M.Green.1974.The 11. Fan,H.,and D.Baltimore. 1973.RNA metabolism of virus-specific RNA species in free and membrane-murine leukemia virus:detection ofvirus-specificRNA bound polyribosomes oftransformed cell replicating sequences ininfected and uninfectedcellsand identifi- murinesarcoma-leukemia viruses. Biochem. Biophys. cation ofvirus-specific messenger RNA. J. Mol. Biol. Res. Commun.56:697-702.
80:93-117. 30. Stoll,E.,M. A.Billeter,A.Palmenberg,and C.
Weiss-12.Fan, H., and P. Besmer. 1975. RNA metabolism of mann. 1977.Avian myeloblastosis virus RNA is ter-murine leukemia virus. II. Endogenous virus-specific minallyredundant: implicationsforthemechanism of RNA in theuninfected BALB/ccell line JLS-V9. J. retrovirusreplication.Cell12:57-72.
Virol.15:836-842. 31. Taylor, J. M. 1977. An analysis ofthe role oftRNA 13. Fan, H.,andH.Mueller-Lantzsch. 1976.RNA metab- speciesas primersforthetranscription into DNA of olism of murineleukemiavirus. III.Identificationand RNA tumorvirus genomes. Biochim. Biophys. Acta quantitationofendogenous virus-specificmRNAinthe 473:57-71.
uninfected BALB/c cell line JLS-V9. J. Virol. 32. Taylor, J. M., R. Ilmensee, and J.Summers. 1976.
18:401-410. Efficient transcription ofRNA into DNA by avian
14. Fan, H., and M. Paskind. 1974. Measurement ofthe sarcoma virus polymerase. Biochim. Biophys. Acta
on November 10, 2019 by guest
http://jvi.asm.org/
442:324-330. tive viruses are at the poly(A) end. J. Virol. 16: 33. Vogt, P. K., and S. S. F. Hu.1977.Thegenetic structure 1051-1070.
ofRNAtumor viruses.Annu. Rev.Genet. 11:203-238. 35. Weiss, S. R., H. E. Varmus, and J. M. Bishop. 1977. 34. Wang, L.-H., P. Duesberg, K. Beemon, and P. K. The size and genetic composition of virus-specific Vogt.1975.Mapping RNase T,-resistant oligonucleo- RNA's in the cytoplasm ofcells producing avian sar-tides of aviantumor virus RNAs: sarcoma-specificoli- coma-leukosis viruses. Cell 12:983-992.
gonucleotides are near thepoly(A) end and oligonucle- 36. Wetmur, J. G., and N. Davidson. 1968. Kinetics of otides commontosarcoma andtransformation-defec- renaturation of DNA. J. Mol. Biol. 31:349-370.