JOURNALOFVIROLOGY, Nov. 1969,p.777-796
Copyright ® 1969 AmericanSociety for Microbiology Vol.
4, No. 5 Prinzted in U.S.A.
Structure and Development
of
Viruses
as
Observed
in
the
Electron
Microscope
X. Entry and Uncoatino of Adenovirus
COUNCILMAN MORGAN, HERBERT S. ROSENKRANZ, AND BAIBA MEDNIS DepartmenttoJMicrobiology, College ofPlhysicianisanidSurgeonls, Columbia Uniiversity,
New York, New York 10032
Received forpublication 10July1969
Stages in the direct penetration of adenovirus through the cell membrane are
illustrated. Phagocytosis with ruptureof the vacuole and release ofvirus into the cytoplasm may also account for entry of some particles. Uncoating by digestion
within phagosomeswasnotobserved.Rather,alteration ofcapsidandcoreoccurred to virions free in the cytoplasm. Nucleoprotein released from virus close to the nucleus was transported to the nuclear matrix by a unique mechanism. These
events were notprevented by puromycin and hence werenot dependent upon the
synthesisofnewenzymes.Theywere, however, energy-dependent.
A previous electroin microscopic study of type
7 adenovirus (2) suggested that
phagocytosis
isthe mechanism by which adenovirus gains entry
tocells. Itwas stated,however, thatthe presence of freevirus within thecytoplasmraises the possi-bilitythat"afraction oftheadsorbed virus
pene-trates directly through the (cell)
membrane,"
aphenomenon that could not be documented at
thetime intervals (1 and 2hr) chosen for
exami-nation. The present study, which examines the entry of adenovirus in
detail,
differs from the preceding communication in four particulars. First, adsorption and penetration of the virustook place in monolayer cell cultures while the
cellswerestill in situ.
Second,
thevirusinoculum waspurifiedbybandingin cesiumchloride.Third, the initiation ofviralentry wassynchronized
by quickly warming thecells to 37C after aperiod
of viral attachment in the cold.
Fourth,
andperhaps most
important,
major
attention wasdevoted to the events
occurring
within the first 45 min after warming thespecimens,
because it wasduring thisperiodof time that Lawrence and Ginsberg (5) andPhilipson (9)reported
that the viral DNA becomes sensitive todeoxyribonu-clease.Theresults to bereportedshow thatvirus isphagocytosed but thatdirect entryto the cyto-plasmalsooccurs. Stagesin the
uncoating
of the virus and entry into the nucleus ofnucleoprotein
areillustrated and described.MATERIALS AND METHODS
Virus.The stock of type 7 adenovirus waskindly
suppliedbyH. M. Rose.
Tissue culture. HeLa cells were grown in Eagle
minimal essential medium (MEM) with
10%/
fetalcalfserum.
Preparation of inocula. The cells were infected with
a 10-1 dilution ofstock virus. After incubation for
3days, the cellswerescraped intoMEMand exposed
to sonic vibration for 5 min; cellular debris was
re-moved by low-speed centrifugation for 10 min. The
supernatant fluid was then centrifuged for 3 hr at
35,400 X gin aSpinco model L ultracentrifuge, and
the resulting pellet was resuspended in 3 ml of Earle
solution by exposuretosonicvibrations for 5 min.
Further purification of the virus was achieved by
centrifugation in gradients of cesium chloride. For
this purpose, the virus was prepared as described
above, except that the pellet wasresuspended in 10
mlof Earle solution.To3.5-ml portions of this
sus-pension, 1.86 g of CsCl (Penn Rare Metals) was
added and the sampleswerespunfor24hr at 30,000
rev/min in a SW 50 rotor of a Spinco model L-2
ultracentrifuge. The viral bands were collected as
described previously (3), and the CsCl was removed
before use by dialysis against several changes of
MEMwithoutserum.
General experimental method for attachment and
entry. After washing the tissue culture cells with
MEMdevoid of serum, 0.3 ml of the inoculum was
added to sparse monolayers in 30-ml Falcon plastic
flasks. These flasks were maintained at 4C for 30
to 60min; thentheywerequicklywarmed by
replac-ing the inoculum with MEM at 37 Cand incubated
atthat temperature for 1 to 180 min. In an attempt
toslow down processes whichmightoccurrapidlyat
37C, some samples were initially incubated at 37 C
for20 or40min andthenwereheldat room
tempera-turefor30minbefore fixation.
Preparation of cells for electron microscopy. After
777
on November 11, 2019 by guest
http://jvi.asm.org/
MORGAN, ROSENKRANZ, AND MEDNIS
cooling thetissue culture to 4C for 5 min, thecells
werefixedfor20min insituwith %loglutaraldehyde
bufferedatpH7.2.Theywerethenthoroughly washed
in Sorenson buffer, scraped off the plastic, pelleted
gently, fixed for 30 min in osmium tetroxide,
de-hydrated in ethyl alcohol, and embedded in epoxy
resin (Epon 812). The sections were stained with
uranyl acetate followed by lead hydroxide and were
examined inaPhilips 200electron microscope.
Metabolic experiments. The ability of metabolic
inhibitorstointerfere withprotein synthesiswas
mon-itored by supplementing bottles containing HeLa
cells (5 X 105) with 14C-algal protein hydrolysate
(New England Nuclear Corp); final concentration,
0.48,ug in 3ml of MEM,0.22 uc,/ml.The cellswere
exposedtotheradioactive precursorsfor30min, and
then the cell sheets were washed thoroughly with
chilled 0.15M NaCl and removed from the plastic
surface bytrypsinization
(0.025%0
trypsin inethylene-diaminetetraacetic acid, 1:5,000). The detached cells
wereresuspended in5%20trichloroaceticacid,and, after
standing in the cold, the insoluble residues were
ex-tractedat 90 Cfor40minand collected onMunktell
no. I F filter discs. The residues were washed
thor-oughly with cold 5c% trichloroacetic acid, and the
dried filter discs were placed in vials containing 10
mlof omnifluor(4g perliteroftoluene).Theretained
radioactivity was determined in a liquid scintillation
spectrometer.
RESULTS
Viral entry. In the initial experiments, tissue cultures were inoculated with suspensions of pellets, which had been obtained by ultracentri-fuging the sonic-treated material of infected cells after first clarifying it by low-speed
cen-trifugation. It soon became apparent that such preparations were of little use for the study of viral uncoating, since viral particles, even be-fore entry at the surface of cells, were found in various stages ofnonspecific disintegration (Fig.
1 and 2). Moreover, massive clumping of virus
was common. Virus obtained by banding in
cesium chloride, however, presented an entirely different picture (Fig. 3). The virions exhibited remarkable uniformityinsize, shape, anddensity
and were not aggregated. Accordingly, fresh,
unfrozen inocula of this type were used for all
subsequent experiments. After an adsorption period at 4C of 30 to 60 min, the cells were in-cubated at 37 Cfor0, 1, 5,10, 20, 30, 40, 45, 60,
120, and 180 min. At time 0, no intracellular
virionswereencountered. (It mightbe notedthat there was remarkable variation from one cell
to the nextin the numberof virions attached to
the surface. The reason for this is not known.) By the 5th min, a few virions were observed within phagocyticvacuoles nearthe surface.The
first free virus in thecytoplasm was found at 10
min (Fig. 10). Although at later times the entry
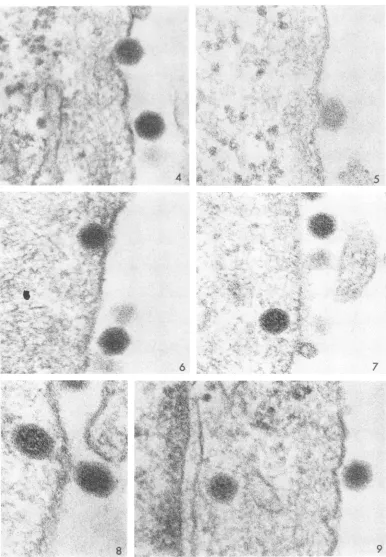
of virus was not synchronous (a point that will be discussed below), the sequence of events could be arranged in what seems to be alogical order. Figures 4 to 6 show initial stages of viral pene-tration. (The difference in appearance of the virus is due to the marked contrast in section thick-ness, the section illustrated by Fig. 5 being un-usually thin.) The fine fibrils in the cytoplasm coursing just beneath the surface were commonly encountered and are unrelated to entry of the virus.
Viral "uncoating." Figures 7 to 13 illustrate virions which have passed further into the cyto-plasm. Itis important to note that in Fig. 7 to 9
and 11 the core of the virion within the cell ap-pears more granular than that of the virion on the surface. The clarity with which this change could be observed depended upon the resolution of the micrograph and the thickness of the
sec-tion. In thicker sections(Fig. 12 or
13),
the granu-larity of the core was less clearly visualized. Another interesting changein the virion, which occurred at about the sametimeastheincreased granularity of the core, was that the surface became indistinct or fuzzy and the crystalline faces were lost, the particle thus appearing to be spherical. To illustrate this clearly, section thick-ness is again important, but, in addition, the level of the virus in the plane of section also playsa role.Thus, the alteration of the capsid is difficult to ascertain with certainty for any given particle, but, when viewed repeatedly and care-fully compared with virus on the surface, the phenomenon is unmistakable. The change is less obvious at 10 min (Fig. 10). It might be noted how close the nucleus at the left margin of Fig.11 is to thesurface of the cell. It was not unusual for this distance to measure less than 200 nm
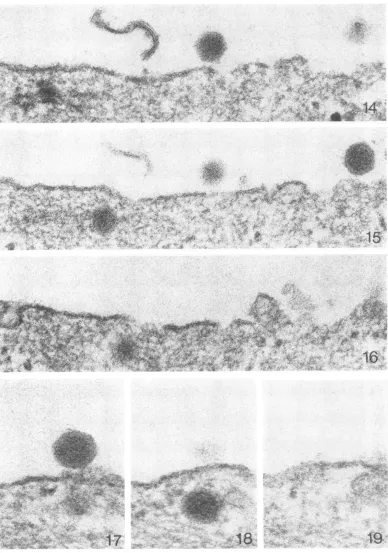
(Fig. 33). Figures 14 to 16 show three serial sections. When central to the plane of section, the two virions on the surface are sharply de-fined and exhibit (particularly in Fig. 14) crys-talline faces, whereas the virion in the cyto--plasm (Fig. 15) is poorly defined with a diffuse peripheral coat. The same phenomenon can be seen very clearly in the serial sections illustrated
by Fig. 17 to 19. In Fig. 20, it will ba noted
that virions nearest to the cell surface (on the right) generally appear less altered than those further in. The foregoing subtle, yet extraordi-nary, changes in the virion have been illustrated anddiscussed insomedetailsimply because their recognition is essential for an understanding of theinitial stages in theuncoatingprocess.
Phagocytosis ofvirus. It was hardly surprising
to find virus within vacuoles, since phagocytosis is an active mechanism by which cells ingest
778 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
e
FIG. 1. Virusfrom resuspended pellet;60mini. X 150,000.
FIG. 2. Virusfrom resuspended pellet;10 min. X 150,000.
FIG. 3. VirusafterbandinginCsCl. Virions aretrappedamongcytoplasmicextensionsofadisintegrating cell;
40min. X 150,000.
779
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.492.52.447.45.592.2]* $ .:.
:,:,
N it
'.4
.,k
8
'4
4t
a
.I
w-9
FIG. 4-6. Passage ofvirus into thecytoplasm. Note that the extracellularparticlesin these andotlher micro-graphsareseldom inappositionwith thecellmembrane.Figure5 isaverythin section. X 150,000.
FIG. 7. Entrywithalterationofthecore. X 150,000.
FIG. 8. Entrywithalterationofthecore. X 200,000.
FIG.9. Virus closeto the nucleus(attheleftmargin).Notethegranularcoreandpoorlydemarcatedperiphery comparedtothe extracellularvirion on1 theright. X 150,000.
780
*\4v A:
,., :, .,> s
J,,;
sr t
4 tr.
.
*:swi.
%*
5
6
7
W'a
iA~
A.-Z
p'~~~4
., ,
.? ,
i.
-.4--Z:...
11 1:
1.1i. .i
i. "
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.492.58.444.30.587.2]~ ~ ~
..7SJaA
*.c
Qt.4W,-~~ ~~~~~4
pESt § eR wJt.
4?
I
- S
iL .fl.Jt
at
4
V
*44 t
S
. I
7'~~ ~ ~~A
4-1
-.1w
44;R
11
12
4
r :13
¢
FIG. 10. Virus close to thenucleus; 10mini. X 150,000.
FIG. 11-13. Intracytoplasmic virionswith alteredcoresandcapsids;20mim. X 150,000. 781
; tt :4p*i.
10
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.492.49.444.42.608.2]%cmab'
1k ;r
iti.;;0X.
Xi' .;.irT.-e i :. / S t}
xe s t:
^w 9
>O^.14t
,..f;.st.:4w<S..a,.e
Aw
A....i,*,
[image:6.492.60.448.41.594.2]~17
19.
FIG. 14-16. Serialsections. Note the diffuse appearance of theintracytoplasmic virion on the left of Fig. 15.
Thisparticlemustbe central to theplane of section since it is only seen once. Adifferent virion begins toappear
inFig.16. X 150,000.
FIG. 17-19. Serial sectionsdemonstrating that, although the virion in Fig. 18 is nearly central to theplanieof section,thecapsidisdiffuseinappearance. X 200,000.
782
1
8'.
on November 11, 2019 by guest
http://jvi.asm.org/
ENTRY AND UNCOATING OF ADENOVIRUS
large amounts of particulate material, viral or otherwise. Itwassignificant, however, that virions within phagocytic vacuoles were generally
un-altered, neitherappearing granular norexhibiting diffusemargins. InFig.21, the virionatthe upper right isextracellular, whereas theone belowlies withina vacuole.They are similar in appearance and do not show thechangesvisiblein thevirion free within thecytoplasm atthe lower left. Other examples of unaltered virions in vacuoles are
shown inFig. 22and 23.
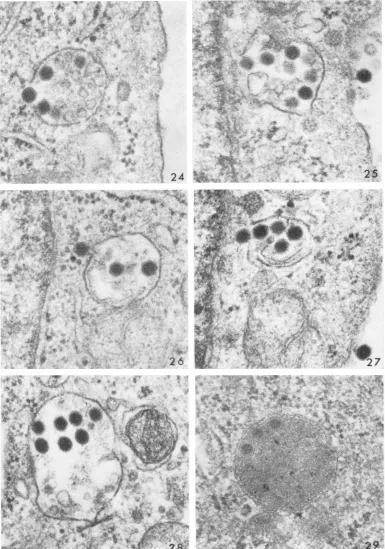
Occasionally, ruptured vacuoles were seen
with virus in the process of release to the cyto-plasm (Fig. 24-27). There did not seem to be a
purposefulmigration of
vacuoles,
sincetheyneverappeared in ordered arrays as though marching from the cell surface to the nucleus.
Rather,
discharge of virus from vacuoles appeared to be at random, occurring at any distance from the cell surface to thenucleus. It should be
empha-sized that not only was it unusual to encounter
virions in the process of release from vacuoles but also that the phenomenon, with rare excep-tions (Fig. 24), was observed only after incuba-tion for 60 min.
Sequestration of virus. A significant proportion ofthe virus became
sequestered
withinvacuoles. Thus, at 3 hr, although most of the free virus within the cytoplasm haddisappeared,
struc-turallyintact virions werefound within vacuoles enclosingmatrix material of low
(Fig. 28)
orhigh
(Fig. 29) density. The relative
proportion
offree virus, incomparison
with that invacuoles,
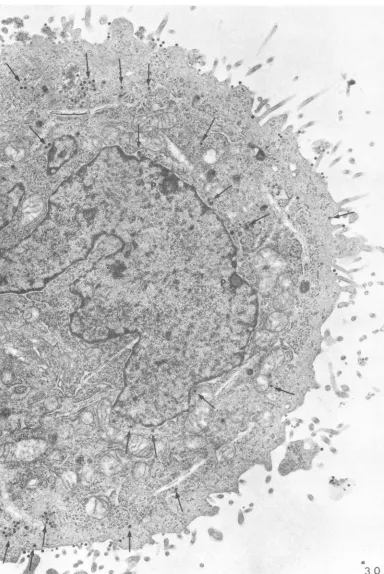
atdifferent intervals after infection can best be illustratedatlow
magnification.
InFig. 30,
which shows a cell after 1 hr of incubation at 37C, there are numerous free virions(marked by
ar-rows). In Fig. 31, which shows a cell2 hrlater,
all of the remaining intracellular virus is present within vacuoles and noneis
lying
free within the cytoplasm. A fewparticles
werealways
found attached to the surface.Presumably
such virions haddefectivecapsids
whichprevented
penetration
(see also under Formalintreatment).
Entry of nucleoprotein into the nucleus. Ulti-mately, muchof the free viruscametolie inclose proximity to the outer nuclear membrane. It is ofinterest inthis connection that the virus could
traverse a considerable distance
through
thecytoplasm, not
infrequently
coming
to rest onthat
portion
of the nucleusopposite
thefreesus-face of the cell. The association of virus with nuclear pores seemed to be quite random. In Fig. 32, for example, one virion (on the right) is closeto a pore,whereas the othertwo arenot.
(The dense intranuclear body at the lower right
cannot beidentified as altered virus or viral
nu-cleoprotein since similar structures also occur in control cells.) In some instances, viral nucleic acid appeared to be discharged from the virus directly througha poreinto the nucleus (Fig. 33 and 34). This phenomenon was- sufficiently rare, however, as to make documentation exceedingly difficult. The events to be described below were much more commonly encountered, reaching their greatestfrequency at about 45 min.
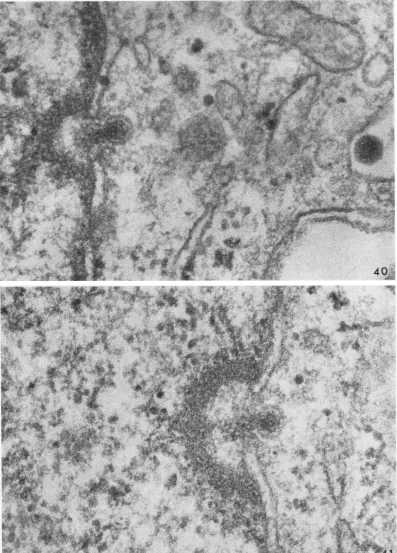
Figure 35 illustrates a virion within a cyto-plasmic protrusion, which indents a nucleus. Several nuclear pores are evident, but there is noabnormal alteration of either nuclear or cyto-plasmic membrane. Figure 36, however, shows the cytoplasmic membrane in proximity to the virus bulging toward the nucleus, while in Fig. 37 a small channel appears to connect the cytoplasm with the nuclear matrix. In Fig. 38, dense material resembling chromatin has col-lected so as to form a pocket extending into the nucleus, and the virus has undergone a striking change. The core is contracted, the diffuse outer coat has disappeared, and a sharply defined membrane-like structure is apparent. This mem-brane may be composed of viral protein(s) (P antigen) thought to be present within the virus but in amasked form (11) andhence not clearly observed heretofore. In Fig. 39, the cytoplasmic membrane curves inward to the nuclear pocket where it vanishes. The pocket is demarcated by granular, chromatin-like material. and the viral membrane proximal to the pocket appears to haveruptured. Oneevengains the impression that the dense viral core material mayhave begun to escape. Figures 40 and 41
streingthen
this im-pression. Occasionally, theentirie
virion seemed todisintegrate (Fig. 42).Usually, after discharge of nucleoprotein, the viral remnant could not be identified, but in
rare instances the ruptured, empty, viral mem-brane remained within thepocket, as seenin the exceedingly thin section illustrated by Fig. 43. The fate of the pocket varied. The side con-tiguoustothe cytoplasmwasgenerally closed off by a rim of chromatin, the nuclear and cyto-plasmic membranes re-forming to close the nu-cleardefect. Theopposite side of the pocket then opened so that, in a manneranalogous to locks in a canal, theviral nucleic acid passed into the nuclear matrix. Stages in this process are shown in Fig. 44 and 45. Sometimes the pocket de-tached and migrated from the margin of the nucleus, presumably opening with release of contents at a later time. In other instances, the pocket opened so that direct continuity was
established between the cytoplasm and the
nu-clear matrix. Figure 46 is an example of this.
VOL. 4, 1969 783
on November 11, 2019 by guest
http://jvi.asm.org/
'it~~~~~~~~~~~~~~~~~~~~~~~~o
*~~~~~~~*
~~~~~~~~~~~~~~~~~~~~~~~~~w
:f :
4. ':U; ?" 'fXWX*
'N.~~~~~~~~~~~~~~~~~~~~~~~R
'4
af
;;
...",''':
'
...,4 ~~~~~~~~~~~~~~~~~~~Z...i ...> r;t
j~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~Ax;.f
mo X + - 0i' S ;-:
e AC
.,"!n,.; -k!l'!:-;,
.:, 4? !ik.
I:1
,
if
:,:I. t*
..4
N171
igV
'
3
FIG.20. Intracytoplasmc virions at 20mihz. Thoseto theleft are niearerthenucleuis alndappear
X 150,000.
FIG. 21. Virus oni the surjice (upper rig ht), within a vacuole (lower right), andfree within7 cytoplasm
(lowerleJi);40miii. X 150,000.
FIG.22 and23. Viriontswithinivacuoles. X 150,000.
784
~~~~~~~~~~~~~~. i.;
.Ss
4go
.t
a
~~~A
pt*S
12
2
-ii4,..,,,
,.
.o.
.r.v
1-21 1
110on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.492.61.448.38.611.2]Amb
4. 4~~~~~~~~~~~~~~~~~~~O
.3I
I.
4.
24
A0
4~~~~~~~~~~~~~~~~~~~~~~~~~%
Jr.1
[image:9.492.54.439.38.587.2]FIo.24. Virionintheprocessofreleasefroma vacuole;45min. X 75,000.
FIG. 25-27. Virusatstagesofreleasefrom vacuolesnearthe nucleus (visibleonitheleftineachcase); 60min.
X 75,000.
FIG. 28. Intactviruswithiniavacuoleafterincubationfor3hr. X 75,000.
FIG. 29. Unusually thin section showing apparently intact virus within a lysosome after incubationi for2 hr. X 75,000.
785
N
i '
_lw~2
on November 11, 2019 by guest
http://jvi.asm.org/
54
,4 lSk'&
.
-Al.Uhis ,
SW
ay,a~
_,*
4r;
.*S
Et1.
i
jeb~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~1.-0
*}rb.
.Sa~~~~~~~~~~V
; F) f , 4, WW ..30,
-FiV.
44aMd45.) X 15,000.~~~X 4;71~~~~~~~~~~~~~~~~~,
V!4-~
~~~ ~ ~ ~ ~ ~ ~~~~~~SA~~~
4 <
,~~~~~W
?<YW
3'~~~~~~-.
fl~~~~~~~~~
~~~~~~~~~~A-.-,4.,t
Al,li
'W~1
S'.~
~~~~~~~~~~~~~t*4~~~~~~~~~
St9.~~~~~~~~~~~~~~~~~~~~~LK
FIG. 30. Free virus (marked byarrows) withinthecytoplasm at1 hr. Notetwo
niuclear
pockets (P). (SeealsoFig.44anid45.) X 15,000.
786
on November 11, 2019 by guest
http://jvi.asm.org/
[image:10.492.57.441.36.610.2]r.W4>,g ,, r f e # ^^;< I
S
j
t;
*-~~~~~~Al
4
;> w
?4,* * '' > S}N'il_
4<~~~~~~~~~~~i
JN.
O,. 4 a,,,
.49,t,',.Ed, .
4, . 69
SC
Cs
.4 r
.
"ga~~~~~~~~~~~f
Art, , 4A
.4' 7$.^>* ,_6 tX 3
Wz~~ ~ ~ ~ ~ ~ ~ ~ C
..4
S. ,sS .:.,, 4'
,CC,MS , Sa t
3
3
' A';' : 'gr ...
X i*~«< ^ . -a
FIG. 31. Virus stillsequestered within vacuolesat3hr.Nofree intracellularvirus remains. X 15,000.
FIG.32. Virus nearthenucleus; 45min. X 150,000.
FIG. 33 and 34. Very thin sections showing probable release of viral nucleoprotein through apore into the
nucleus;45 min. X 150,000.
787
41-Alk
on November 11, 2019 by guest
http://jvi.asm.org/
[image:11.492.52.444.24.614.2]P"~~ ~ ~
7~~~~~~~~~~~~~~~~~~~~~~~k
C., :
z
S! &
37
FIG. 35. Virionlyin2g within2afinger of cytoplasm, which hasindenited theniucleus. Nuclear poresareevident;
20miti. X 150,000.
FIG. 36-39. Earlystages inthe releaseof
nucleoprotein.
Fig. 36,20min;Fig. 37-39,40or45min. X 150,000.788
:.1
4.R
l
I JI,
ziv
Or ANAN
IAl
If
'.. 11
C. i4l, I
i.
'k:-"
"Ir ?. :-.7:
-.
PM7R.
:T.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:12.492.59.445.31.592.2]40
.oes
oryr
N-1~*
*tw
s
oR'
...
¢S
s./N owov £.
..
X-*-r
-.
wf
$] '-e3f>
Ws ! t v-i;- i_ = -Ns,, s. it-.
v rf ;3ttZfsj ;t
Ak
&>aL-( \.yXt$'* {
Ft
X
h>f#s.s
#
re
e
w
sse
.. ,M5L'^' X{e
f 5
'
v
B
'RS
X
W s. <.W ;i;R
#s:,t
' .s *s @w.w
t%0*
.t7b
},t
htt sPM s $t tS
*t\
ea
ou
j2;
..IX
XX'
at5
tC
st.
r
,*,,
i tt 'w'' B .*Jt,. \X ., Uph i
E o < gw f f :
E^i.
*
...
;
....^
\<r::
m2'o.i > 9' j$S.
L S * CX @ 4;t*\g;Ah -b b
1
W@bi.'
d7o'1
FIG. 40. Core material (nucleoprotein) in theprocess
of
release toa nuclearpocket.
Forcomparison, anun-alteredvirion isshownonthesurfaceofthecell;40 min. X
150,000.
FIG. 41. Thinnersection showingcorematerial
(nucleoprotein)
in theprocess ofrelease to a nuclearpocket;
40min. X 150,000. __ _, i3 i
789
on November 11, 2019 by guest
http://jvi.asm.org/
[image:13.492.52.449.34.587.2],t,-4:
7
Si A
4..
*
;t ti
4¶2
*'Yr.
'4..
"'.4~~~~~~4
.4r
:
1X.vt.,. . ,.e '
e
S
.,
4 4, 4-~~~~~~~4
..-;
4, V,"'t~~~~~~~~~V
r4.~~~~~~~~~~~~~~~~~~j4
Z.~~~~~.
4 V
All
'A?;D* < ;
ne,2dJ,!S,p, -i A W t"fl\;jt~~~~~~~~~~-.kek+ X
4K4|S,i'h ag'; ;w \i;\< jV1Zitsfi W4
[image:14.492.58.444.24.595.2]i
FIG. 42. Probablealtered virionatthenuclearmargin;30min. X 150,000.
FIG. 43. Unusuallythin sectionshowinganemptyviral membranewithinanuclearpocket.Note how closethe
ntucleusistothesurface ofthecell;40 miii. X 150,000.
790
43
on November 11, 2019 by guest
http://jvi.asm.org/
ttS
<-r
~~~~~~~~~~~.1l
Imm ; ¢i-44JE tafikXi} .s ,4 . <+' X%*st.sw ^;-8e, n~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~1
>
SJ
<-*'
vf ;8
~~~~~;AYR4e
f ^ . r.. .t .Ae ~ ~ A
;54~~~~~~~~~~~
Sr.~
791~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~j
on November 11, 2019 by guest
http://jvi.asm.org/
-5~ ~ ~ ~~~~~~. *1
Aw
lbi
Att
in~~~,It
.
*..6
4 a
*0 ~ ~ a
SIT
W4~~~~~~~~~~~~~~~~~~~
[image:16.492.50.442.40.604.2]','
FIG. 46. Nucleoproteinatastage in passagefrom aviral remnzant througha nuclearpocket into the n2uclear matrix;40miM. X 150,000.
792
a.
.4
.$."i
.-..e
..%sW,
ji-4
4..
46
Ak z,
.A 'P'
k -Ws%,
I
i.
,4;.
.r4
.:
-j.
-!k.,
.r
.".
..'S«:
:'i
* ', W .
on November 11, 2019 by guest
http://jvi.asm.org/
ENTRY AND UNCOATING OF ADENOVIRUS
The nucleoprotein is observed just as it escapes from the viral shell. Thepresenceof virionswithin
nuclei (4) couldnotbeconfirmed.
Formalintreatmentof theinoculum. Inaneffort
to determine whether alteration of the capsid interfered with penetration of virus through the
cell membrane, resuspended pellets of viruswere
incubated with 1% Formalin for 1 hr at 37 C
before bandingonCsCl. Afterdialysis, the virus was applied tothe cells, which wereheld at 4C
for 30min. The preparations werethen warmed
to 37C and fixed 1 hr later. There were
con-siderably fewerattached viralparticlesat thecell
surface and only moderate numbers within
phagocytic vacuoles. Virtually no virus was
foundfree in thecytoplasm.
Attemptstopreventphagocytosis.Pretreatment
with India ink in aneffort to blockade the cells was unsuccessful; 0.5% India ink in MEM was
addedto the tissuecultures 19 hr before addition
ofvirus. It was of interest that there was great
variabilityin theamountof ink takenupby
differ-ent cells. However, cells that were actively
phagocytic invariably engulfed some viral
particles.
Metabolic inhibition with colchicine, puro-mycin, dinitrophenol (DNP), and arsenate was
also studied for the effects of these agents on
phagocytosis and viral entry. Cells were pre-treated for 19 hr with 5 ,ug of colchicine per ml
of medium. Eventhough viral adsorption inthe
cold andincubationat37 C for 1 hrwerecarried
out in the presence of the drug, phagocytosis still tookplace. Pretreatment with 50,ugof
puro-mycin permlfor2hr, with continued treatment
duringviraladsorptionand incubation, reduced,
but failed toprevent, phagocytosis. In the
pres-ence of puromycin, occasional virions were
found free in the cytoplasm, and virus in the process ofdischarging nucleoproteinintonuclear
pocketswasseen.
DNP (2 X 10-3 M) wasappliedtothecellsfor
2 hr, and treatment was continued during viral
adsorption inthe cold for 30 minandincubation
at 37 C for 45 or 60 min. Although the cells
showed a striking loss ofpolyribosomes, phago-cytosis of virus occurred. Moreover, free virus was found in the cytoplasm, and, in a few
in-stances, viral nucleoprotein was observed at
stagesofpassagetothenuclearpockets. In view
ofthe likelihoodthat cells in the presence of DNP
couldutilizeenergyobtainedbyglycolysis, 10-2M
arsenate, an inhibitor of both oxidative and
fermentative energy metabolism (1, 16; see also
references 10, 14, 15), was applied to the cells 1
hr before andduringtheperiod ofviral
adsorp-tioninthe cold for 30min and incubationfor 1
hr at 37 C. This agent caused severe damage to the cells, as manifested by beginning autolysis with loss of ribosomes, discontinuity of endo-plasmic reticulum, and fragmentation of mito-chondrial cristae. Yet, even under these circum-stances, some phagocytosis of virus resulted. No freevirus was encountered in the cytoplasm. In another series of experiments, the cells were not pretreated, but 10-2M arsenate was added with the viral inocula. Aftera 30-min period of adsorptionat 4 C,the preparations were warmed to 37 C for 1 hr. Although moderate phagocy-tosisand occasional freevirions in the cytoplasm wereobserved, the viral particles were not altered in appearance and nuclear pockets were not formed. Attemptstorevive thecells by removing thearsenate after the period of incubation, with the hope of initiating viral uncoating, were un-successful, the cells appearing to be too badly damagedto recover.
Tomonitortheeffectivenessof these agents on HeLa cells, their
ability
to interfere with protein synthesis was determined. It was found that 50jig
of puromycin per ml prevented protein synthesis, whereas 2 x 10-1 M DNPallowed
some synthesis to occur (12% of the control; Table1).Arsenate (10-2
M)
inhibited metabolism completely, provided that the cells were pre-treatedforatleast30min (Table2).TABLE 1. Effect ofpuromycin and DNP on the
metabolismofHeLacelisa
Addition Radioactivity(counts/5Xincorporated106scells)
None... 21,531
Puromycin (50
Ag/ml)
... 0DNP (2 X 10-3M)...
2,561
aMonolayers of HeLa cells (5 X 106/bottle)
wereexposedtothepuromycinorDNP in 3 ml of
Earle solution for 2 hr at 37 C and then were
washed with cold Earle solution also containing
the agent.Aftermaintainingthecellsat4 Cfor 30
min, they were incubated in the presence of the
inhibitors for 20 min at 37 C, whereupon
radio-active amino acids were added and the
prepara-tions were incubated for another 30 min. The
incorporated radioactivity was determined as
described. The general procedure was similar to
the one used when cells were actually infected
with virus. Theresultswerecorrectedfor the
non-specific adherence of radioactivity to the cells.
This was calculated in a control experiment, in
which radioactive amino acids were added to a
chilled culture and thecellswereprocessed
imme-diately for measurement of the retained
radio-activity.
VOL.4, 1969 793
on November 11, 2019 by guest
http://jvi.asm.org/
MORGAN, ROSENKRANZ, AND MEDNIS
TABLE 2. Effect ofarseniate on the metabolism of
HeLacellsa
Radioactivity Part Conditions Radioactivepulse (min) incorporated(counts/5X
106cells)
I Preincubated with 0-30 0
0.1 M arsenate for
1 hr at 37C
II As in I 30-60 0
III No pretreatment; 0-30 2,682
arsenateaddedat
T= 0
IV As in III 30-60 0
V Untreated cells 0-30 8,258
VI Untreated cells 30-60 10,875
a Before adding the pulse of radioactive amino
acids, the cellswereheldat4 Cfor30min andthen
werequicklywarmedto37Cinamanneridentical
to that employed for the viral adsorption and
entryexperiments.InpartsI, III,andV,the
radio-active labelwasaddedimmediatelyuponwarming
the cells (T = 0) and the preparation was
har-vestedat 30 min, whereas in parts11, IV, and VI
the radioactive label was added after 30 min of
incubation at 37 C and the cellswere harvested
30minlater.
DISCUSSION
This study has shown that unlike ether-sensi-tive viruses, such as
herpes simplex
(8),
in-fluenza (7), and Sendai(6),
which appeartofuse with thecell membrane and release theircontentsinto the
cytoplasm,
adenovirus can passdirectly
through the cell membrane.
Presumably
this mode of entry requiresanunalteredcapsid,
sinceit was not observed to occur after treatment of
the virion with Formalin. It should be
empha-sized that virus in passage
through
the cell membrane was far lesscommonly
encountered thanwasviruswithinphagocytic
vacuoles. How-ever, inattempting
to assess the relative im-portanceoftwodiffering
mechanismsfrom micro-graphs of fixedspecimens,
it doesnotnecessarily
followthat aneventwhich is seldom seen
rarely
occurs. Stages in the direct
penetration
of viruswere not often
encountered,
but this can beex-plained
by
assuming
that the process israpid
rather than uncommon. Since
nonenveloped
virus was found deep within the
cytoplasm
asearly as 10 minafterthe initiation of entry
(Fig.
10), it is not unreasonable to conclude that the actual passage of the virion
through
the cell membranemight
occurinasmallfraction of this time. On theotherhand,
phagocytosis
is acon-stant process
resulting
in theslow, progressive
accumulation of
large
numbers ofparticles
within vacuoles, such as were observed in the studieswith India ink. It is hardly surprising then that many virions were found within phagocytic vacuoles. Philipson (9) alluded to this situation by making the following statements. "The present results are, however, not entirely com-patiblewith electron microscopic data on adeno-virus penetration and eclipse (2). Thus the rapid disintegration of 65 to 85% of the virions ob-servedin this study is in contrast to the slow pene-tration and accumulation of intact particles within the cellobserved byelectron microscopy."
Noevidence was found to support the hypoth-thesis that uncoating occurs within phagosomes. Virus sequestered in vacuoles actually appeared to be protected, since structurally intact virions were still present even after 2 to 3 hr of incuba-tion at 37 C. In rare instances, vacuoles contain-ing virus were encountered at stages of rupture with release of virus into the cytoplasm. Whether such rupture is fortuitous or whether virus can penetrate thelimiting membrane of a vacuole in the same way as the surface of the cell is not clear. However this may be, some virus un-doubtedly does reach the cytoplasm upon the disruption of phagocytic vacuoles. In view of the difficulty encountered in blocking phagocytosis, it is not possible to determine the proportion of virus released from vacuoles as opposed to that entering directly through thecell membrane. The question naturally arises as to why others have failed to observe direct entry of virus into the cytoplasm (2, 4). Presumably, as stated above, this phenomenon occurs so quickly that the eventcould easily escapedetection if some degree ofsynchronywere notattainedby rapidly warm-ing thecells afteradsorption of virus in the cold, and if intensive study were not devoted to the ensuing 30 min. An interesting question raised by the present investigationis why all of the at-tached virus doesnot entersimultaneouslywithin, say,thefirst 10 min. Onecanonly surmise that for admissionto occur thecapsid and the cell
mem-branemust becontiguous. As was mentioned in thestudy of influenza virus (7) and as isevident from examination of the accompanying micro-graphs,thevirions may lodge in the proteinaceous coating of the cellat some distance from the cell membrane. Presumably a variable interval of
time ensues before virus and cell achieve actual
contactandpenetration results.
Once within the cytoplasm, the virion very quickly underwent subtle but significant altera-tion. The core became granular while the surface assumed a diffuse appearance, and the
crystalline faces,socommonlyexhibitedby
extra-cellular virus,were no longerseen. One can
con-794 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
ENTRY AND UNCOATING OF ADENOVIRUS
clude that both the configuration of the capsid as well as that of the core changed. It is of in-terest in this connection that Lawrence and Gins-berg found that "though the viral capsid was altered to expose the viral DNA to deoxyribo-nuclease activity, the capsid proteins were not degraded by the process" (5). They commented ontherapidityof this event bystating: "Uncoat-ing followed penetration closely; both processes occurred at similarrates, and bothwerecomplete by 60 min." [Philipson (9) reported completion within 30min.] Inaddition, theynoted that "the uncoated viral DNA wasstable within thecell;ie.. it was notdegradedtoacid-soluble material." One is led to suggest, therefore, that the afore-mentioned changesin structureof the viralcapsid and core probably correspond to
uncoating.
In other words, "uncoating" is not the actual re-moval of the coat but refers "tothe intracellular alterationsof the viralcapsidwhich render paren-tal DNA accessible to the action ofdeoxyribo-nuclease," to quote Lawrence and
Ginsberg
again. Perhaps the alterationis
analogous
tothat observed uponheating,
wherein the pentons and neighboringfivehexonsarelost with accompany-ing development ofsusceptibility
of deoxyribo-nucleic acid to deoxyribonuclease (12). Such achange,oronesimilarto
it,
inconfiguration
ofthe capsidmayaccountfor the diffuse appearance of theviralsurfaceseeninthin section.The events
occurring
atthe nuclearmembrane were truly remarkable. Afterpassing
a variable distancethrough
thecytoplasm,
the virus cameto lodge in
proximity
to thenucleus. Thecapsid
then was either
markedly
altered or actually strippedoff,
leaving
athin,
well-definedmem-brane
enclosing
thenucleoprotein.
Quite
pos-sibly thesechanges
are, infact,
the "further al-terations"whichLawrenceandGinsberg
deduced mustoccur "before free viral DNA isliberated"(5). At
approximately
the same time, chromatinaccumulated inthenucleus
adjacent
to the virus so that a pocket wasformed;
thecytoplasmic
and nuclear membranes
bulged
inward toward the nucleus and gave way. The viral membrane ruptured and thenucleoprotein
passed directly from thevirusinto thepocket. Thelatter,
either at the same time orshortly
thereafter,
opened and the nucleoprotein passed to the nuclear matrix. [Dales (2) seems to have encountered this phenomenon, asillustrated in Fig. 22 ofhis paper, butheinterpretedthemicrographas prob-ably showing a stage in the actual entry of the virion intothenucleus.]
As was noted in the description of the micro-graphs, viral particles were occasionally found
near nuclear pores, and in rare instances dis-charge of nucleoprotein seemed to occur through these structures.There was no evidence, however, ofanypredilection by thevirus to lodgeclose to pores and little to indicate that they served as the main portal of entry, a suggestion which occurs in reports of other systems [see the re-view by Stevens (13)].
In view of the morphological observations recorded above, the effects of metabolic in-hibitors were investigated. It was thus found (Table 1) that, although puromycin blocked protein synthesis, this property was not shared
by
DNP. Presumably cells exposed to DNP are able to obtain energy for proteinsynthesisthrough fermentative processes. Arsenate, on the other hand, halted protein synthesis completely, pro-vided that the cells were pretreated for at least 30 min (Table 2).Entry
ofvirus, alteration of the virion, and discharge of nucleoprotein to the nucleus were not prevented by pretreatment of thecells withpuromycin, indicating, asobserved by Lawrenceand Ginsberg (5) and byPhilipson (9), thatuncoatingof thevirus isnot dependent upon the synthesis of new enzymes. When cell energy was blocked by arsenate, however, entry of small numbers ofvirions into the cytoplasm was seen but alteration of virions, formation of nuclear pockets, and release of nucleoprotein did not ensue.Phagocytosis wouldseemtobeone of the lastmanifestations of cellularactivity that was lost, since, in cells which had been treated with arsenate and were obviously undergoing autolysis, a fewvacuoles were found to contain structurallyintactvirions.The purpose of this communication has not been to excludephagocytosis as ameans of viral entry but ratherto indicate that direct penetra-tion can also occur. It was repeatedly observed thatphagocytosistended to sequesterstructurally intact virions, often for considerable periods of time, a phenomenon which could be interpreted as suggesting a defensive mechanism ofthe cell rather than a specific modeofviral entry. How-everthismaybe, only virus free inthecytoplasm was found to become altered and to release nucleoprotein to the nucleus. Penetration of recognizable virions into the nuclearmatrix was neverencountered.
Illustrations. To facilitate direct comparison most of the micrographs are reproduced at x
150,000. In each figure, with the exception of the sixserials (Fig. 14to19) and the lastmicrograph (Fig.46),the free surface of thecellistothe right and the nucleus to the left, whether these struc-turesdo or donotappear inthefieldchosenfor reproduction. When significant, the time of incu-bation at 37 C is indicated.
795
VOL. 4, 1969
on November 11, 2019 by guest
http://jvi.asm.org/
MORGAN, ROSENKRANZ, AND MEDNIS
ACKNOWLEDGMENTS
This investigation was supported by Public Health Service grantAI-06814from theNationalInstituteofAllergy and Infec-tious Diseases and grant GB-6013 from the National Science Foundation. Oneof theauthors (H.S.R.) isaResearchCareer Development Awardeeofthe Division ofGeneral Medical Sci-ences, United States Public Health Service (5K3-GM29, 024). Thisinvestigation wasconducted under thesponsorship of the CommissiononInfluenza,Armed ForcesEpidemiological Board, and was supported by the U.S. Army Medical Research and Development Command, Department of the Army, under
re-searchcontractno. DADA 17-67-C-7141.
ADDENDUM
After submission of this report we were informed of the study of Karl Lonberg-Holm and Lennart Philipson entitled"Early Events ofVirus-Cell
Inter-action inanAdenovirusSystem,"whichwaspublished
in vol. 4, p. 323-338, Oct. 1969, of this journal.
The results appearto becomparable, inspite ofthe
fact that different types of adenovirus were used.
Presumably, their B component corresponds to the
structures shown in Fig. 4, 5, and 6 ofthis paper.
Their C component is probably the spherical form
withthe granular core illustrated in Fig. 7 through
21;their D componentmaybethe densecorevisible
in theparticlesofFig.38through 41.Whether theE
component hasbeenvisualized is uncertain. Itseems
likely that forms C and D, rather than being
in-tranuclear as they suggest, were adherent to the
nuclear membrane and thusappearedinthe nuclear
fraction.
LITERATURECITED
1.Clifton,C. E. 1946. Microbialassimilation. Advan. Enzymol. 6:269-308.
2. Dales,S.1962. An electronmicroscope studyoftheearly
asso-ciationbetween two mammalianviruses andtheir hosts.J. Cell Biol. 13:303-322.
3. Ellison, S. A.,and H.S. Rosenkranz.1963. An apparatusfor samplingafter cesium chloridedensitygradient
centrifuga-tion in the preparative ultracentrifuge. Anal. Biochem. 5:263-265.
4. Fong, C. K. Y., K. G.Bensch, and G. D. Hsiung. 1968. Productive and abortive infections of simian and nonsimian cells withasimian adenovirus SV15. 1. Microscopic ob-servations. Virology 35:297-310.
5. Lawrence, W. C., and H. S. Ginsberg. 1967. Intracellular uncoating oftype 5adenovirusdeoxyribonucleic acid. J. Virol. 1:851-867.
6. Morgan, C., and C. Howe. 1968. Structure and development ofviruses as observed in the electron microscope. IX.
Entryofparainfluenza I (Sendai) virus. J. Virol. 2:1122-1132.
7. Morgan, C., and H. M. Rose. 1968. Structure and develop-ment of virusesas observed in the electron microscope. VIII. Entryof influenza virus. J. Virol. 2:925-936. 8. Morgan, C., H. M. Rose, and B. Mednis. 1968. Electron
microscopy of herpes simplex virus. I. Entry. J. Virol.
2:507-516.
9. Philipson, L. 1967. Attachment and eclipse of adenovirus. J. Virol. 1:868-875.
10. Reiner, J. M. 1948. The inhibition ofenzymeformationand nitrogenassimilation byarsenate.Arch. Biochem. 19:218-228.
11. Russell, W. C., and B. E. Knight. 1967. Evidence fora new
antigenwithin theadenovirus capsid. J. Gen. Virol. 1:523-528.
12. Russell, W. C.,R.C.Valentine, and H. G. Pereira. 1967. The effect ofheatontheanatomy of the adenovirus. J. Gen. Virol. 1:509-522.
13. Stevens, A. R.1967. Machinery of exchangeacrossthenuclear envelope,p.189-271. In L. Goldstein (ed.), Thecontrolof nuclear activity. Prentice-Hall, Inc., Englewood Cliffs, N.J.
14. Sussman, M., and S. Spiegelman. 1950. Theuseofarsenatein theanalysis ofenergyrelationsamongstsynthetic reactions. Arch.Biochem. 29:85-100.
15. Ter Welle, H. F., and E. C. Slater. 1964. Uncoupling of respiratory-chain phosphorylation by arsenate and evi-dencefor theexistence ofastableX - Pintermediate of
oxidative phosphorylation. Biochim. Biophys. Acta 89: 385-388.
16. Warburg, O., and W. Christian. 1939. Isolierung und Kris-tallization des Proteins desoxydierenden Garungsferments. Biochem. Z.303:40-68.