JOURNALoFVIROLOGY,Feb.1968,p.129-148 Copyright © 1968 AmericanSociety for Microbiology
Replication
of
Bacteriophage
Ribonucleic
Acid:
Analysis
of
the Ultrastructure
of
the
Replicative
Form
and
the Replicative Intermediate
of
Bacteriophage R17
NICOLE GRANBOULAN ANDRICHARD M. FRANKLIN
Laboratoire deMicroscopie Electronique, CentredelaRecherchesScientifiquessurleCancer, 94 Villejuif, France,and The Public HealthResearch Instituteofthe City ofNew York, Inc.,
NewYork,NewYork 10016
Receivedforpublication2November1967
A detailed qualitative and quantitative comparison was made of the
ultrastruc-tureofsingle-stranded ribonucleic acid (RNA)frombacteriophageR17and double-strandedreplicative form (RF) and replicative intermediate (RI) fromcellsinfected with this bacteriophage. The nucleic acids were prepared for electron microscopy by the protein monolayer spreading technique of Kleinschmidt. Single-stranded RNAaggregated duringspreadingintheabsenceofurea, whereasRF and RIdid
not. On the other hand, RF and RI appeared to be susceptible to shear during spreading, whereas R17 RNA was not. From the maximal length ofRF, a base
translation of 3.14 A was calculated. This value favors a 10-fold helix model of double-stranded RNA. The same base translation was found for R17 RNA, in-dicating a stacked basestructure for single-stranded RNA spreadin the presence ofurea. RI isa branchedstructure and thebranchesareremovedby ribonuclease
treatment.The branchesarebelievedtobenascentsingle-stranded viral RNA. The
contourlength ofthebranchwasequaltothecontourlength of the main chainup to the branch point, as predicted from theoretical analysis of the replication of
viral RNA. The structure of RF and the main chain of RI was also analyzed by plotting the log (end-to-end distance squared)versuslog (contour length). This dem-onstratedstructuresintermediate instiffness betweenarandom coil andarigid rod. The key role of double-stranded ribonucleic
acid (RNA) in the replication of the genetic materialof RNAbacteriophageshasbeen demon-strated by a wide variety of in vivo and invitro experiments (9, 26, 27). Two species of
double-stranded RNA have been isolated from cells infected with RNA bacteriophage and their properties have been described (1, 10-13). Repli-cative form (RF) isdouble-stranded RNA com-posedofaviralRNAmolecule (+ strand) hydro-gen-bonded to acomplementary RNAmolecule
(- strand). Replicative intermediate (RI) is double-strandedRNAwithnascentsinglestrands.
It is presumably derived from a more complex
structureinwhichthe RNA isprobablyassociated
with RNA polymerase and may sometimes be associated withpolysomes (16).
Studies of the in vitro reaction have demon-strated a conversion of parental RNA to RF, subsequent conversion ofRFtoRI, andfinally a
transfer of label from RI to newly synthesized
si-igle-strandedviral RNA(26).Semiconservative and conservative models for this process have
been published (9). Some of the experimental datafavor thesemiconservative model of replica-tion (7, 8, 24).
Inthispaper, theultrastructural characteristics of RF and RI are analyzed. Since RI contains both single- and double-stranded components, it wasalsonecessarytoinvestigate the ultrastructure of single-stranded RNA. Unfortunately, single-stranded and double-stranded RNA molecules had the same width in shadowed preparations, precluding any unequivocal identification ofthe twocomponentsof RI. However, twoproperties of single-stranded RNA could be exploited in comparative ultrastructural studies. These were
itssusceptibilitytoribonuclease digestionand its
intra- and intermolecular aggregation during
preparation for electron microscopy.
129
Vol.2,No. 2 Printed inU.S.A.
on November 11, 2019 by guest
http://jvi.asm.org/
GRANBOULAN AND FRANKLIN
Some preliminary data have been publishedon certain of the parameters to be discussed in this
paper (14).
MATERIALS AND METHODS
Preparation ofRNA. Single-stranded RNA from bacteriophage R17, double-stranded RF, and RI
were prepared as previously described (11, 14). RI
wasdenaturedby heatingto95 Cfor 3minin10-3M
potassium phosphate buffer (pH 6.6) with 10-s M
ethylenediaminetetraacetic acid (EDTA) or by heat-ing to 37 C for 10 min in phosphate-EDTA con-taining 85.7%7o dimethylsulfoxide (DMSO) as described earlier (13). DMSO lowers the melting
temperature (T,0) of double-stranded RNA so that
it maybedenatured underverymildconditions (18). In some experiments, RI was treated with
ribo-nucleaseto digest the single-stranded branches (12). Ribonuclease A (Worthington Biochemical Corp., Freehold, N.J.) was used at aconcentration of 0.08
to 0.1 ,ug/ml, and the RI in 0.1 M NaCl, 0.001 M
EDTA, 0.05 M
tris(hydroxymethyl)aminomethane-chloride buffer atpH6.98 (25 C) was treatedfor 10 min at 37 C (11). After digestion, the ribonuclease
wasremoved byasingle phenolextraction (11). Controlsweremadeby mixingRF andR17RNA. Some preparations were simply mixed (2.3 optical density units,at 260m,, for each RNA) and others
were mixed and extracted once with phenol after
mixing.
Preparation for electroni microscopy: dilutioni of nucleic acids before spreadinig. The single-stranded
RNA extracted from bacteriophageR17 wasdiluted
inurea atafinalconcentrationofeither 6or 8M, to
avoid aggregation of the RNA molecules due to
intra- or intermolecular hydrogen-bonding (15; Granboulan, Scherrer, and Franklin, in press;
Scherrer and Granboulan, inpreparation). The
con-centration ofthe single-stranded RNA in urea was
about 50 Ag/ml. RF and RI were diluted to a final
concentration of 10 to 30 ag/ml, either in (i) 6 or 8
M ureaunder thesameconditionsas the viral single-stranded RNA or in (ii) 0.02 M EDTA at neutral
pH. Single-stranded RNA preparations spread from 6to8Murea were always made at the same time in ordertocheck the action ofurea ontheultrastructure
ofRI.
RI which had been denatured with DMSOatlow temperature or by heatinginthe absence ofDMSO
was diluted in the same manner as single-stranded RNA (in 6or 8 M urea and at final concentrations of 2.5or5 jug/ml).
In control reconstitution experiments, samples of RF and single-stranded RNA were mixed in equal concentrations and were then diluted in either 8 M
ureaor0.02 MEDTAaccordingtotheprocedureused
for RF and RI. The same procedure was also
fol-lowedoncomparable "reconstituted" sampleswhich
had been treated with phenol aftermixing. The final
concentrationof these sampleswas 20 MAg/ml. Spreading and shadow-casting. All of the various
preparations of nucleic acids described above were
spread bythe protein monolayer technique of
Klein-schmidt and co-workers (19-22). Several proteins weresuccessfully used to form the monolayer: cyto-chrome c (Fluka, Buks, Switzerland), chymotrypsino-gen(Nutritional Biochemicals Corp., Cleveland, Ohio), diisopropylphosphoryltrypsin (Worthington Bio-chemical Corp.), and lysozyme (kindly provided by G. Cohen, Gif sur Yvette). Lysozyme was adopted for routine studies because it seemed to give the best differentiation between RNA molecules and back-ground. Itwasusedat afinal concentration of 0.01% in ammonium acetate buffer (1.0M, pH 8.0) contain-ing0.05%- isopropanol. The solutions of protein and ammonium acetate buffer were always filtered on membrane filters (average pore diameter, 0.22 us;
Millipore Corp., Bedford, Mass.) before use. De-pending on the concentration of RNA, 0.1 or 0.2 mlof the RNA solution was added to 1 ml of lysozyme in ammonium acetate buffer; 0.2 ml of this final mix-ture wasgently dropped onto ameticulouslycleaned glass slide from which it flowed onto the surface of ammonium acetate buffer (0.015 M, pH 8.0) in a glass dish. The glass slides were cleaned by keeping them ina 1:1 solution of nitric acid in distilled water and then rinsing them with distilled water followed by ammonium acetate buffer (0.015 M, pH 8.0) just before use. They were always handled with forceps. After spreading, the film was picked up on 300-mesh grids covered with a carbon-coated Formvar film. The specimens were dried in alcohol and iso-pentane and were then shadowed with uranium oxideat anangle of 7°at a vacuumof5 X 10-5 mm
of Hg. The specimens were rotated at 50 rev/min during shadowing. Some samples of RI were shadowed with platinum-carbonatthe same vacuum, but with-outrotation.
Measurements. Thespecimens were examined with a Siemens Elmiskop I or a Philips EM 200 electron microscope at nominal magnifications between 15,000 and 30,000. The actual magnifications were determined with the aid ofacarbon grating replica. The lengths of the molecules were measured on prints withamapmeasurer atfinal magnifications of 45,000 to 130,000. Theend-to-end distance was also
determined.
RESULTS
Single-stranded RNAfrom bacteriophage R17.
Although the single-stranded RNA was spread from solutions whichwereapproximately two to five timesas concentrated as RFor RI, the fre-quency ofmoleculesperunitareaofgridwasfar
less thanfordouble-stranded RNA. Thismay be due to the loss of single-stranded molecules in theammoniumacetatesolutionduringthe spread-ing process (14). It isnot duetothe presenceof
nucleases in the protein,since ribosomalRNA is
notlostduringspreading (Granboulan, Scherrer,
andFranklin, in press; Scherrerand
Granboulan,
inpreparation).
All ofthemolecules spread from solutions of6
to 8 M urea werelinear, i.e., noncyclic and non-branched (Fig. 1). An analysis of 117 molecules
130 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
REPLICATION OF PHAGE RNA
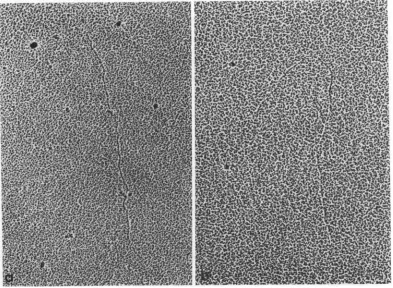
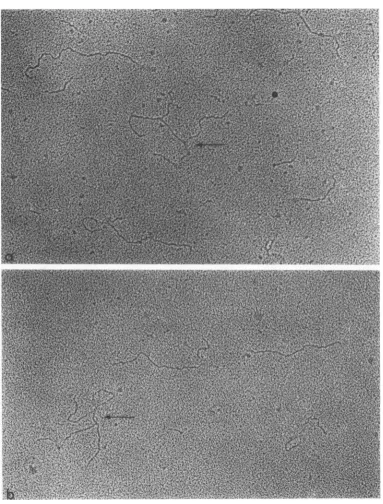
FIG. 1. Single-strandedRNAfrombacteriophage RI7,spread in the presence of urea. Examples of whole mole-cules (a,b) and ofafragment (b). X80,000.
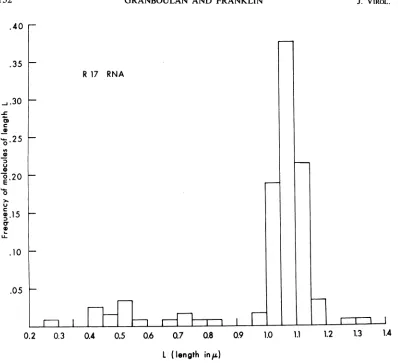
revealed a modal length between 1.05 and 1.10
,l
(Fig. 2)
and a meanlength
of 1.06 i 0.12,u.
RF.There was nodifferencein theshape ofRF spread in the presence orabsenceof
urea.Inbothcasesthe
molecules
werelinear(Fig. 3);
therewasno tendencyto aggregate inthe absence
of
urea(Fig. 3a)andnobranchedmoleculeswere
fotnd.
Besides the prominent population of molecules distributed aroundameanlength
of 1.05 i 0.03 ,u anda modallength
between 1.05 and 1.10A.,
there were smaller molecules with
lengths
cor-responding
toone-half
andone-fourth
thatof
theformer
population
of molecules(Fig. 4).
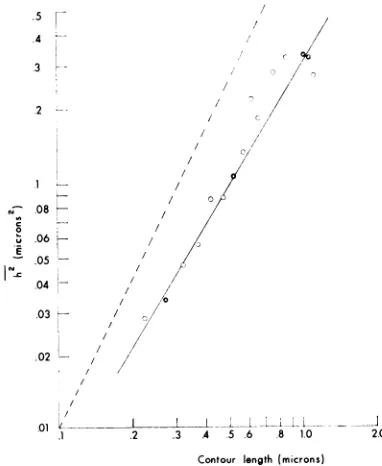
An analysis of the shape of the spread RF molecules was made by means of a double-logarithmic plotof
the mean square end-to-enddistance versus the contour length (19, 20, 22).
Accordingto this analysis, the
molecules
spreadin theabsence ofurea wereintermediate in struc-turebetween a
rigid
rod anda random coil (Fig.5).
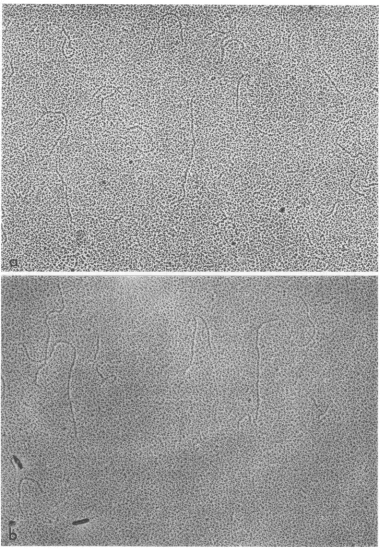
RI.Whether spreadin the presence orabsence ofurea, all
preparations
of RI contained linearmolecules, some of which were branched and othersofwhich were unbranched (Fig.
6-8).
Nocircularmoleculeswerefound inany RI
prepara-tions. Branches were attached at all
possible
positions
along
thepresumed
main chain. Theleng.hof the branch varied accordingto thepoint
of
attachment to the main chain(Fig. 6).
The variation in branch lengthwasverystriking in therare cases
of
RIwithtwobranches(Fig.
7). WhenRI was spread in the absence
of
urea, branches located closeto oneendofachainwerefrequently coiled(Fig.
8c). This configuration was notobservedwhen RI was
spread
in thepresence ofurea.
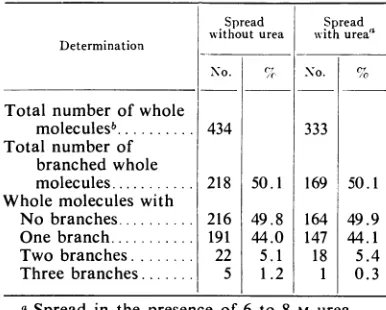
DataonthebranchesofRImolecules are
pre-sented in Table 1. Similar values were
found
for molecules spread in the presence or absence ofurea. Only 50%
of
the molecules had branches, and almost allof
thosewith brancheshad only asinglebranch. Veryrarely,molecules with two or three branches were seen, but none with more than three branches. Inafew instances,RI
mole-culesappearedto have twobranchesat onepoint. Such molecules were examined closely at high
magnification; inevery case,these were two
over-lapping molecules. An example of two
over-lappingmolecules is shown in Fig. 9b, a
prepara-131 VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.490.47.440.77.365.2]GRANBOULAN AND FRANKLIN
.40
r-R 17 RNA
I-I
I I I I I I I Il I __ X
0.2 0.3 0.4 0.5 Q6 0.7 0.8 0.9 1.0 1.1 1.2 1.3 1.4
[image:4.490.61.459.60.423.2]l (length inai)
FIG. 2. Histogram ofthelengthdistributionofR17RNAspreadin thepresenceofurea;atotal of117molecules
wereanalyzed. For 1.45to1.50uandfor1.65 to1.70I,afrequency of0.009wasfound. Thesearenotincludedin
thehistogram forlackofspace.
tion of ribonuclease-treated RI spread in the
presenceofurea.
RI which had been treated with ribonuclease had fewor nobranches(Fig. 9).Ofatotal of 295
molecules spread without urea, only 13 (4.4%)
werebranched; ofatotalof361moleculesspread
with urea, only 7 (1.9%) were branched. These
preparations behaved like RF, since there was
notastrikingloss of moleculesduringthe spread-ing process as there was in the case of
single-stranded R17 RNA. Furthermore, the molecules
spreadintheabsence ofureadidnotform
aggre-gates.
The maximal length of the presumed
double-strandedcomponentofRIwasthesameasthatof RF(Fig. 10,11).Thehistogram for881 molecules
spreadintheabsence ofureahadapeakbetween 1.00and 1.10A (Fig. 10).Themeanlength
calcu-lated for the 333 molecules oflength equalto or
greaterthan 1.00,uwas 1.05 i0.06 ,u. The histo-gramfor 809molecules spread inthepresence of ureaalsohadapeak between 1.00 and 1.10 ,u (Fig. 11). Themeanlength calculated forthe 434
mole-cules of length equal to or greater than 1.00 ,u
was 1.05 + 0.04j,. Therewereshorter molecules
present in both types of preparations and there were peaks corresponding in length to approxi-mately one-half and one-fourththe length ofthe
longest molecules.
The shapeorstiffnessof thepresumed
double-stranded component of RI was analyzed by a
double logarithmic plot of end-to-end distance squared versus contour length. When spread in
the absence of urea, these molecules were more rigidthanarandom coil butnotasrigidas astiff rod (Fig. 12). When spread in the presence of urea, the stiffness of the double-stranded
com-ponentwasincreased,since theplotcontinuedas .35
_.30
C
0
O.25
0 0
u
.20
S
0
u C
T0
U-.10
.05
132 J. VIROL.
---I
on November 11, 2019 by guest
http://jvi.asm.org/
REPLICATION OF PHAGE RNA
FIG.3. Replicativeform(a)spreadintheabseniceofureaanid(h) spreadinthepreseniceofurea. Wlholemolecules (lenigth : IIA),aswellasole-halfanidonie-quiartermolecules,areillustrated. X60,000.
133
VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.490.52.440.74.630.2]GRANBOULAN AND FRANKLIN J. VIROL.
Replicative Form (spread without urea)
496 molecules
0.1 0.2 3 0.4 Q5 0.6 0.7 0,8 0.9 1.0 1.1
L (length in,u)
FIG. 4. Histogram of replicative formspreadinthe absence ofurea; atotal of 496 molecules were analyzed.
.5/ .4
3
D-.2/
/
/
0
N08
0
04 - ,
0
.03
/ /
.02 - ,
.1 .2 .3 A 5 6 8 10 20
Contour length (microns)
FIG. 5. Doublelogarithmic plot ofthemeant
end-to-elid distanice squared versus the contour length (h2'
versuscontourlength) of replicative form spreadinthe
absence of urea. In this plot, contour lengths were
pooled into groups of O.OS-,u steps; variable sample
numbers were available for each group (G = more
thani50samplespergroup; 0 = less thaniSOsamples
pergroup). The dotted line represenits therigidrod.
a straight line up to the maximal length of RI
(Fig. 13). Thiscurve wasalmost identicaltothat of RF (Fig. 5).
According to the semiconservative model of replication, the branches of RI are the partially
synthesized single strands of viral RNA which are displaced from the double strandas newviral
RNAissynthesized (7,9,12). Sincesynthesisand
displacement are presumed to occur simultane-ously,thelengthofthebranchshould be thesame asthe length of the double strandto the branch point, providedthat the mass per unit length of the single-stranded RNA isjust one-half that of
the doublestrand (9). Thisconditionseemstobe
true when one compares the maximal lengthsof R17 RNA and RF (see Discussion). Therefore,
themodelwasinvestigated by measuring the
con-tour length of the presumed double strand up
tothe branchpoint.
Asalready mentioned, it hasnot been possible
to distinguish the single-stranded and
double-strandedcomponentsofRIby theirwidth.
Never-theless,from evidencepresentedhere(see
Discus-sion) and elsewhere (12), it is clear that one of the branches of the branched molecules is single-stranded. Theoretically, the single-stranded
branch could haveall possible lengths up to the
length of the viral RNA (12). The double-stranded chain shouldbeofconstantlength equal
to the length of RF. Since there seemed to be breakage of the presumed double-strand during
134
.24
J
-C
a
e .20 0
0
0 .16
E
0
c . 12
0
0
.08
.04
I
on November 11, 2019 by guest
http://jvi.asm.org/
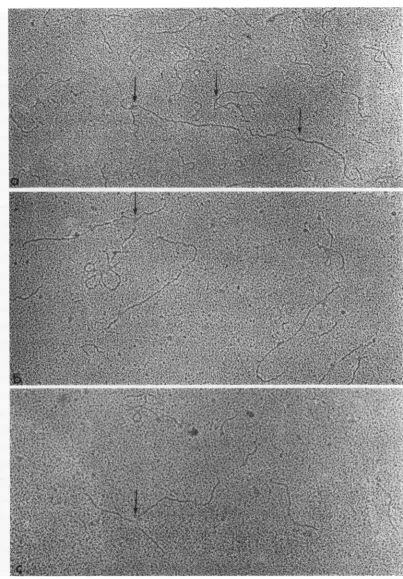
[image:6.490.61.447.48.305.2] [image:6.490.56.247.337.570.2]FIG. 6. Replicative intermediate spreadin thepresence ofurea. The arrows point out whole molecules with variation inthe branch point andin thelengthofthebranch. Rosette structuressuchasthat seenin 6c werevery rarelyfoundinbothRIand RF. Thenatureof thesestructures is notknown. X 60,000.
135
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.490.41.444.44.623.2]GRANBOULAN AND FRANKLIN
spreading, the analysis of the relation between length of branch and its position on the main chain wasdoneonly onmain chainsof maximal length.
Despite the problem of identification of the branch, contour lengths of the presumed branch and
of
thepresumedmain chainuptothe branchpoint
were measured. Often therewas a definiteFIG. 7. Replicative intermediate spreadinthe presence ofurea. Whole molecules withtwobranches (arrows);
variationinthebranch point andinthelength of the branch. X 60,000.
136 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.490.62.445.123.627.2]REPLICATION OF PHAGE RNA
FIG. 8. Replicative intermediate spreadintheabsenceof urea;w,idirectionalshadowingwith PtC; (a, b) whole molecules withsinglebranches; (c) long coiledbranch attachedto oneendofawhole molecule. (a) X 105,000; (b, c) X 102,500.
137
VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
FIG. 9. Replicativeintermediate treated with ribonuclease A (0.08
pg/ml,
10 min, 37C)followed by removal of the enzyme by phenolextraction: (a) spreadinthe absenceof urea;(b) spreadinthepresenceofurea.Nobranches wereobserved in these preparations. There is a pair of crossed molecules in theupperleft-handcornerof 9b. X 60,000.138
on November 11, 2019 by guest
http://jvi.asm.org/
REPLICATION OF PHAGE RNA
break inthedirection ofthemoleculeat the branch
point. Theoretically, the branch contour length (X) shouldequalthelength ofthedouble-stranded
molecule up to the branch point. Extensive measurementsofthese two parameters were made on whole RI molecules spread in the absence or presence of urea.
A scatter diagram ofthe datafor RIspread in the presence of urea is shown iii Fig. 14. There was agoodcorrelation betweenXand y, the
con-touLrlengthof the main chain tothe branch point.
TABLE 1. Ultrastructure of replicative i,ztermediate
Determination
Total number of whole moleculesb... Total number of
branched whole molecules ... Whole molecules with
No branches... Onebranch... Two branches... Threebranches...
Sp)read Spread withouturea with ureaa
No. %c No. %
50.1 49.8 44.0 5.1 1.2 333 169 164 147 18 1 434 218 216 191 22 5 50.1 49.9 44.1 5.4 0.3
a Spread in the presence of 6 to 8 M urea.
IMolecules with contour length of 1.0 to 1.1 ,.
Branches of all possible lengths were found, but
most ofthebranches were 0.2 to 0.4piin length.
Themaximal length of the branches was 1.05 ,u, approximately equal to the length of the single-strandedR17 RNA.
Theapparent correlationi between X and y was confirmed by least-squares analysis from the equation y = bX + c, where b and c are the con-stants to be determined (Table 2). This
analysis
was performed on a LOCI 2a computer with a programwhich handles a sample of 99 at any one time.Therefore, the samples of 266 molecules (RIspread in the absence of urea) and of 210 mole-cules (RI spread in the presence of urea) were
split into three parts. Since the measurements were made at random, thevalues ofband c were very similar for each subsample. Whether spread in the absence or presence of urea, the resulting average values of b and c were the same and very
closetotheexpectedvalues ofb = 1 and c = 0. ThedistributionsoflengthsofXfromRIspread in theabsence and thepresence of urea is shown in Fig. 15 and 16, respectively. Neither
distribu-tion was uniform, but the distribution was more uniform with RI spread inthe absence of urea. When spreadin the presence of urea, there was askeweddistribution with a maximum at 0.20 to 0.25 ,u.Thiscanalsobeseeninthe scatter diagram (Fig. 14). In both preparations of RI, there were
Replicative Intermediate(spread without urea) 881 molecules
.08 H
.04 H
F-r
1 .2 .3 A .5 .6 .7 .8 .9 1.0 1.1 1.2
.28H .24 -.20 e-.16 W-.12 H c C 0 -0 0 0) E 0 c 0) cr 0 LL L(length inp)
FIG. 10. Histogram of thepresumed
maini
chtaint
ofRIspreadinz
theabsentce
ofurea; a totalof881molecules wereanialyzed.
VOL. 2, 1968 139
-I
on November 11, 2019 by guest
http://jvi.asm.org/
[image:11.490.43.236.211.366.2] [image:11.490.45.436.355.627.2]140 GRANBOULAN AND FRANKLIN J. VIROL.
.28 Replicative Intermediate (spread with urea)
809 molecules
. .24
0 .20
u
E
0
u .12
U_ .08
.04
.1 .2 .3 .4 .5 .6 7 .8 .9 1.0 11 1.2
L
(length
inEs)
FIG. 11. Histogram of the presumed mainchlain of RIspreadinthepresenceofurea;atotalof809molecules
wereanalyzed.
.5
.5-c /
/
~~~~~~~~~~~~.4/
.2
.1 / .0 00 '
(>08~~~~~~~~~~~
.0 /
~~~~~~.05~~~~~~~~~~~~~~~~~.8/
bj .05
.04 0
-/04
.03 - /.03K
.02 5 8. .
02 / i10
.1 .2 .3 .4 .5 .6 s 1.0 2.0 _01
l/_I
l_ l_ l_I*l2 .3 A .S .6 .8 l.0 2.0
Cnurlength of double strand (microns)
[image:12.490.59.448.54.334.2]Contour length of doublestrand(microns)
FIG. 12.Double-logarithmicplot
oJf/T2
versus contourlength for thepresumed main chain of RI, spread in FIG. 13.Double-logarithmic plot
oJ'h2
versuscontour the absence ofurea. At allpoints the sample was 20 lengthfor thepresumed main chain ofRI, spread inorgreater. 0 =individuallengths;for lengths within the the presenceofurea. Thenumberof samplesperpoint
ranges indicated,data werepooled to give a sample of variedbetween 26 and 69. The dotted line represents
20orgreater. Thedottedline represents the rigid rod. the rigid rod.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:12.490.56.248.374.615.2] [image:12.490.260.452.376.618.2]REPLICATION OF PHAGE RNA
Number of occurrences
00 00 0 0 0 0
0 00
. A n
oo 0 0 w
00 0
0 0
00 0 00 0 0
-~~~~G
oo e 000 0 o
_0 0 0 00 0
0,&&Q0%00 oo _ 0 0 1I 0.2 0.4 2 3 4 5 6 . 7 8
0.6 0.8 1.0 1.2
CONTOUR LENGTH OF BRANCH (microns)
FIG. 14. Scatter diagram of theconitourlentgth of the presumedmaint chainito thebranich pointt (y) anldthe con-tourlenigthofthebranich (X). Thisisthesample of210whlolemolectulesspreadiln thepresenice ofuireaanldusedfor
the liniearleast-squaresanalysis ofTable 2.
TABLE 2. Correlationt betweeni branich poinlt anld
branzcha
Urea Group Samplesize b c
Absent 1 98 0.978 0.028
2 98 1.009 0.004
3 70 0.959 0.052
Avg 0.98 0.03
Presentb 1 89 1.016 0.018
2 59 0.958 0.039
3 62 0.958 0.036
Avg 0.98 0.03
aFrom the formula Y =
bX
+ c where y =distance to branch point measured on double-stranded molecule and X = length of the branch.
bConcentration,6to 8 M.
branches ofall possible lengths uptothe length ofthe
single-stranded
R17molecules.Denatured RI. Samples of RI denatured with DMSO or by heating could be examined only
after spreading in the presence of urea. One attempt was made to examine the denatured RI
spread in the absenceofurea.There wasa great dealofinter- andintramolecularaggregation,and
thereforethe remainder of thepreparations were
made in thepresenceofurea.Linear molecules of highlyvariablelengthwerefoundinsuch prepara-tions (Fig. 17).No branchedmoleculeswere seen. The length distribution of 860 molecules of RI denatured with DMSO showed a peak
cor-responding to molecules of R17 RNA (1.05 ,u)
and a heterogeneous population with lengths
between 0.02 and 1.05 ,u (Fig. 18). The length
distribution of 389 RI molecules denatured by heatingshowedasimilarheterogeneity (Fig. 19).
Mixedpoptulations of RFandR17 RNA.
Con-trolexperiments were done with 1:1 mixtures of double-stranded RF and single-stranded R17 molecules. In the presence of urea, branched molecules amounted toabout 3 of the
popula-tion (Fig. 20). Therewas noincrease in the
per-centage of branchedmolecules in the absence of
urea, but aggregates were frequently found.
Exactly the same results were obtained with
mixtures which had been extracted witb phenol before spreading and mixtures which had not beenextracted with phenol.
DIscUSSION
Since measurements of base translation are available for double-stranded RNA but not for single-stranded RNA, the length of the
double-1.2 1.0 0.8 0.6 0.4 0.2 U) c 0 E F-z 5
0D
m I 0 z m 0 II 'r H z w -J 0 z 0 0 141VOL. 2, 1968
0 0
I
on November 11, 2019 by guest
http://jvi.asm.org/
[image:13.490.79.402.74.349.2] [image:13.490.41.232.390.543.2]GRANBOULAN AND FRANKLIN .1 2 I .1 0 z w U. 0
(,,.0
8 w 11 Q z,.06
U-0 0 z w04 w U-.0 2
0.1 0.2 0.3 0.4 0.5
RI SPREAD WITHOUT UREA
XILu
[image:14.490.92.424.67.339.2]0.6 0.7 0.8 0.9 1.0 1.1 X(CONTOUR LENGTH OF BRANCH in,u)
FIG. 15. Histogram ofXforRIspread ihtheabselice ofairea;atotalof266wholemolecules ofRIwereanialyzed.
stranded RF will be considered before thatof the single-stranded viral RNA. The histogram ofRF
has three peaks, the peaks atthe shorter lengths corresponding to approximately one-half and one-fourth the longest length. The mean of the
cluster around the longest length was 1.05 ,.
RFshould have 3,342 base pairs, by comparison with R17 (28); therefore, the calculated base translation distance for spread RF was 3.14 A.
According to the original analysis of X-ray diffraction patterns of double-stranded RNA
fromreovirus, the RNAwasthought to bea
10-fold helixwithapitch length of30.0 Aperresidue
(23, 29). More recent data and theoretical analyses of thestructures have revealedan
ambi-guity, since the diffraction patterns could be
accounted for by either a 10-fold or an 11-fold helix (2-4). Since the pitch length is thesamefor
either helix, the base translation may be either 3.00or 2.73 A perresidue. The ambiguity arises
from two possible ways of packing molecules in thecrystallographic unitcell. Sincecontourlength
measurements are made on isolated
molecules,
this ambiguity does notariseinthepresent deter-mination. With abase pair translation of3.14 A
anda pitchlength of30.0A, the number of base
pairsperpitch length would be 9.6, thusfavoring the original 10-foldhelixmodel ofLangridgeand
Gomatos (23).
Thesingle-strandedR17 RNAdistribution has
a single peak, and themeanlength ofR17 RNA is1.06,g.Thenumber of nucleotidespermolecule is 3,342 (28), and therefore the translation per base
residue is 3.17 A for this RNA spread on
mono-layers from solutions of 6to8 M urea.This is the
sameinternucleotide distance determined for RF,
implying that the spreadsingle-stranded RNA has
astacked baseconfiguration. Evidence for stacked
base structures insingle-stranded nucleic acidsin
solutionmay be found in recent literature. Only
onesuchexamplewillbecited.Themassper unit
length of single-stranded DNA, measured by small-angle X-ray scattering, is just half that of double-stranded DNA, implying stacking and
orderingof the base planes inthe single-stranded
DNA (25).
The length distribution of the presumedmain chainofRI is similartothatof RF, withapeak
between 1.00and 1.10 ,u and smaller peaks cor-respondingtoone-half and one-fourth this length. NeitherRFnorRIhad suchpopulations of mole-culespriortospreading, accordingtoanalyses of
the molecular weight distributions by
ultra-centrifugation (12). The presence of short
seg-ments ofRFfromRNA bacteriophage M12 has
also been noted inelectron micrographs, but no
distributionshave beenpublished (1). Populations
of "one-half" and "one-fourth" molecules of
142 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
REPLICATION OF PHAGE RNA 143
.2 0
.18
.16
.14
.12
RI SPREAD WITH UREA
z
.0a
z
co.06
UA.
0
W.04
w
2
0.1 0.2 0.3 0.4 0.5 0.6 0.7 0.8 0.9 1.0 .
X(CONTOUR LENGTH OF BRANCH in/)
FIG.16. HistogramofXforRIspreadihthepreseniceofurea;atotalof2JO whiole molecuiles ofRIwerealnalyzed.
bacteriophageT2 DNA have alsobeen observed inspreadpreparations, butit isnotclearwhether these preparations already contained such mole-culespriortospreading (19).The double-stranded RNA has a greater rigidity than the single-strandedRNA(12), andismaybethat theformer
RNA issubject toshearing forces during spread-ing on monolayers. Breakage of the double strands byshearing would explainthedistribution into discrete whole, half, and quarter molecules (6).RIspreadinthe absence ofureahadahigher
proportion ofwhole molecules thanRIspreadin the presence of urea (Fig. 10, 11). Since the RI spread in the presence of urea would have a
slightly greaier rigidity than that spread in the absence ofurea, it would havea slightly greater
susceptibilitytoshear. Syntheticpolymers (poly-isobutylenes) as small as 500,000 in molecular weight are susceptibleto shearing (5), but ithas
been reported that double-stranded RNA from cells infected with fr bacteriophage is not de-graded inablenderat 10,000rev/min (17).
The problem of unequivocally identifying the single-strandedcomponent of RI is verydifficult
to solve. Single-stranded R17 RNA has four characteristics which distinguish it from double-stranded RF: (i) aggregation in the absence of urea, (ii) loss during spreading, (iii) sensitivityto
ribonuclease, and (iv) lengtlh distribution around asingle mode ratherthan the tri-modal distribu-tionof RF.Noneof thesecharacteristicsisuseful
fortheidentification of the single-stranded
com-ponent of individualRI molecules. Inthepresent
study, only RI approximately 1 ,u in length was
used for structure analysis. The I-, component
was considered to be the double-stranded main chain,andthecomponentsofshorter length were considered to be the single-stranded branches.
VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
[image:15.490.67.413.56.441.2]FIG. 17. Replicative
inttermediate deniatured
with (a)dimethyl
sulfoxideor (b) byheatin1g
at 37 C for 3miii;
both samples were spread inithepreseniceofurea. X 46,800.144
on November 11, 2019 by guest
http://jvi.asm.org/
[image:16.490.70.461.36.632.2]REPLICATION OF PHAGE RNA
This approach is in keeping with theoretical and experimental analysis of RI (11-13). It enabled us to determine the length distribution of branches andcorrelate branch length with position on the main chain. There is admittedly an unavoidable
ambiguity in identifying a "starting end" of the
.12
.10
0
r .08
e 0
E .06
0 u 04
0 0 .02
F-F
F-H
.1 .2 .3 1 .5
main chain. However, this can be eliminated by taking a starting end such that the branch point would beofapproximately the samelength as the main chain. If one measured from the other end
of the main chain, then y would equal L - X where L isthe maximum length of the mainchain.
.6 7 .9 1.0 1.1 1.2
l (length in,a)
FIG.18. HistogramofRI denatured withDMSO; a total of860 molecules wereanalyzed.
.12
.1_
0
- .10
0 0
E
- 06 0~
c
O .04
U-.02
1.0 1.1 1.2
L(length
ink,;)
FIG.19. HistogramofRIdenaturedbyheating(3miii,97 C); a total of389 moleculeswere analyzed.
145
VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
[image:17.490.59.440.162.393.2] [image:17.490.53.446.422.640.2]GRANBOULAN AND FRANKLIN
But rearranging the analysis so that y X for all determinations does not detract from the
established geometrical relationship between the branch and the branchpoint.
The RI used inthisanalysishadapproximately
one single strand per double strand (11, 12).
Assuming aPoissondistribution ofsingle strands, approximately 37%ofthe population should be
unbranched whereas only 50% of the population had visible branches. This may be due to two factors: loss of branches by shearing during spreading and the presenceof very short branches which cannot be distinguished from the back-ground grain. The presence of a smaller propor-tion oflonger branches in RI spread in the pres-enceof urea may be indicative of losses by
shear-FIG.20. Example ofa mixtureofRFandR17RNAspreadinthepresenceofurea;inthisexamplethetwotypes ofmoleculesweresimplymixedtogetherandthenspread. X 46,800.
146 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:18.490.59.438.170.634.2]REPLICATION OF PHAGE RNA
ing, since the single-stranded branches should be
stifferin thepresenceofurea; verylongbranches might be denatured from the double strand in urea, since onlyafew hydrogen bondsmaybind
itto the main chain.
Denatured RI behaves like single-stranded RNA in that it aggregates if spread in the absence of urea. Similardistributions wereobtained after denaturation atlow temperature in the presence
ofDMSO or athigh temperature inthe absence of DMSO. The highest peak corresponded to the complete RNA molecule. No molecules longerthan R17 RNA werefound,but molecules
of allpossible shorterlengthswerefound. Neither
distribution couldbefitted to the expected
distri-bution ofequal numbersof molecules of all
pos-sible chain lengths from N = 0 to 3,341, plus twice this number of N = 3,342 (12, 13). Con-sidering thedifficultiesofaccurate measurements, however, the distribution of heat-denatured RI (Fig. 19) approaches the theoretical distribution. Except for the peaks at 0.30 to 0.35 ,uand 0.40 to 0.45 u, the
frequency
of0 < N < 3,342 wasapproximately 0.5
a.-d
the frequency for N =3,342 wasapproximately 1.0. Thus, this
distribu-tion wouldalsofavoran RIstructure containing onlyonebranchperdoublestrand.
No branched molecules were found when RF and R17 RNA were mixed, even ifthe mixture
was treated with phenol before examination by
electron
microscopy.
This makes itunlikely
thatRI arises as an artifact by association of single-stranded viral RNA and double-stranded RNA
during phenol extraction of infected cells.
Forfurther investigation ofstructures such as
RI, it is imperative that some ultrastructural
technique
bedeveloped
todistinguish
single-stranded anddouble-strandedRNA. Theanalysis ofthebranchesofRIcould then bemadewithoutany ambiquity.
ACKNOWLEDGMENTS
We acknowledgethe excellent technical assistance of Elizabeth Hinckley and Marianne Salditt, who prepared thenucleic acids used in this study, and of Alain Niveleau, who assisted in the electron
micro-scopic preparation.
Thisinvestigation was supported by Public Health
Service grant AI-07645 from the National Institute ofAllergyand Infectious Diseases andby grant GB 5365 from the National Science Foundation.
LITERATURE CITED
1. AMMAN, J., H. DELIUS,ANDP.H. HOFSCHNEIDER. 1964. Isolation and properties of an intact
phage-specific replicative form of RNA phage
M12. J. Mol. Biol. 10:557-561.
2. ARNoTr, S., F.HUTCHINSON, M. SPENCER,M. H. F. WILKINS, W. FULLER, AND R. LANGRIDGE.
1966. X-ray diffraction studies of double helical ribonucleic acid. Nature 211:227-232. 3. ARNorT,S., M. H. F. WILKINS, W. FULLER, AND
R. LANGRIDGE. 1967. Molecular and crystal
structuresof double-helical RNA. II.
Determi-nationand comparison of diffracted intensities for the a and d crystalline forms of reovirus RNA andtheirinterpretation intermsofgroups
of threeRNA molecules. J. Mol. Biol. 27:525-533.
4. ARNOTT,S., M. H. F. WILKINS,W. FULLER, AND R. LANGRIDGE. 1967. Molecular and crystal structures of double-helical RNA. III. An 11-fold molecular model and comparison of the agreement between the observed and calcu-lated three-dimensional diffraction data for 10- and 11-fold models. J. Mol. Biol. 27:535-548.
5. BESTUL, A. B., AND H. V. BELCHER. 1953. Deg-radation of polyisobutylenes on shearing in solution. J. Appl. Phys. 24:1011-1014. 6. DAVISON, P.F. 1959. Theeffect ofhydrodynamic
shear on the deoxyribonucleic acid from T2 and T4 bacteriophages. Proc. Natl. Acad. Sci. U.S. 45:1560-1568.
7. ERIKSON, R. L., AND E. ERIKSON. 1967. Structure and function ofbacteriophage R17replicative
intermediate ribonucleic acid. II. Properties of the parental labeled molecule. J. Virol. 1:523-528.
8. ERIKSON, R. L., M. L. FENWICK, AND R. M. FRANKLIN. 1964. Replication of bacteriophage RNA:studies onthe fate of parental RNA. J. Mol. Biol. 10:519-529.
9. ERIKSON, R. L., AND R. M. FRANKLIN. 1966. Symposium on replication of viral nucleic acids. I. Formation and properties of a repli-cative intermediate in the biosynthesis ofviral ribonucleic acid. Bacteriol. Rev. 30:267-278. 10. FRANCKE, B., AND P. H. HOFSCHNEIDER. 1966. Infectious nucleic acids of E. coli bacterio-phages. IX. Sedimentationconstantsand strand integrity of infectious M12 phage replicative-form RNA. Proc. Natl. Acad. Sci. U.S. 56: 1883-1890.
11. FRANKLIN, R. M. 1966. Purification and proper-ties of thereplicativeintermediate of the RNA bacteriophage R17. Proc. Natl. Acad. Sci. U.S. 55:1504-1511.
12. FRANKLIN, R. M. 1967. Replication of bacteri-ophage ribonucleic acid: somephysical proper-ties of single-stranded, double-stranded, and branched viral ribonucleic acid. J. Virol. 1: 64-75.
13. FRANKLIN, R. M. 1967. Replication of bacteri-ophage ribonucleic acid: some properties of native and denatured replicative intermediate. J.Virol 1:514-522.
14. GRANBOULAN, N., AND R. M. FRANKLIN. 1966. Electron microscopy of viral RNA, replicative
form, and replicative intermediate of bacteri-ophage R17. J. Mol. Biol. 22:173-177. 15. GRANBOULAN, N., AND K. SCHERRER. 1966.
147
VOL. 2, 1968
on November 11, 2019 by guest
http://jvi.asm.org/
GRANBOULAN AND FRANKLIN
Examen au microscope electronique de RNA monocatenaires. J. Microscop. 5:54a. 16. HOTHAM-IGLEWSKI, B., AND R. M. FRANKLIN.
1967. Replication of bacteriophage ribonucleic acid: alterationsin polyribosome patterns and association of double-stranded RNA with
polyribosomesin Escherichia coliinfected with bacteriophage R17. Proc. Natl. Acad. Sci. U.S. 58:743-749.
17. KAERNER, H. C., AND H. HOFFMANN-BERLING. 1964. DieBildungvonRNS-Doppelstrang zur Vermehrung eines RNS enthaltenden Bak-teriophagen. Z. Naturforsch. 19:593-604. 18. KATZ, L., AND S. PENMAN. 1966. The solvent
denaturation of double-stranded RNA from
poliovirus infected HeLa cells. Biochem. Biophys. Res. Commun. 23:557-560.
19. KLEINSCHMIDT, A. K. 1967. Nucleinsauren. Ar-chitektur und Wandlungsfahigkeit im mole-kularen Bild. Naturwissenschaften 54:417-428. 20. KLEINSCHMIDT, A. K., T. H. DUNNEBACKE, R. S. SPENDLOVE, F. L. SCHAFFER, AND R. F. WHIT-COMB. 1964.Electron microscopy ofRNAfrom reovirus and wound tumorvirus. J. Mol. Biol. 10:282-288.
21. KLEINSCHMIDT, A. K., D. LANG, D. JACHERTS, AND R. K. ZAHN. 1962. Darstellung und Langemessungen des gesamten desoxyribo-nucleinsaure-Inhaltes von T2-Bakteriophagen. Biochim. Biophys. Acta 61:857-864. 22. LANG, D., A. K. KLEINSCHMIDT, AND R. K.
ZAHN. 1964. Electronen mikroskopische Dars-tellung, Langenverteilung und Konfiguration einstrangiger Desoxyribonucleinsaure im Cyto-chrom-c-Film. Biophysik 2:73-78.
23. LANGRIDGE, R., AND P. J. GOMATOS. 1963. The structureof RNA. Science 141:694-698. 24. LODISH, H. F., AND N. D. ZINDER. 1966.
Semi-conservative replication of bacteriophage f2 RNA. J. Mol. Biol. 21:207-209.
25. LUZZATI, V., J. WITZ, AND A. MATHIS. 1967. The structure of nucleic acids in solution, as determined by x-ray scattering techniques: DNA, RNA, poly A, p. 41-55. In D. Shugar
[ed.], Genetic elements, properties and func-tions. Academic Press, Inc.,New York. 26. MILLS, D. R., N. R. PACE, AND S. SPIEGELMAN.
1966. The in vitrosynthesis ofanoninfectious complex containing biologically active viral RNA. Proc. Natl. Acad. Sci. U.S. 56:1778-1785.
27. PACE, N. R., D. H. L. BISHOP,ANDS. SPIEGELMAN.
1967. The kinetics of product appearance and template involvement in the iin vitro replication of viral RNA. Proc. Natl. Acad. Sci. U.S. 58:711-718.
28. SINHA,N.K., R. K. FUJIMURA,ANDP.KAESBERG. 1965.Ribonucleasedigestion of R17 viral RNA. J. Mol. Biol. 11:84-89.
29. TOMITA, K. I., AND A. RICH. 1964. X-ray dif-fraction investigations ofcomplementary RNA. Nature201:1160-1163.
148 J. VIROL.