THE INFL1®lCE OF AGE ON '.mE PATHOGENESIS OF POXVIRUS DISEASES IN MICE AND STUDIES OF THE INTE~roN RESPONSE OF MICE 'It) VIRUSES
By
T. P. SUBRAHMANYAN
A thesis submitted for the degree of Doctor of Philosophy
in the
The results of the investigations described in this thesis are based upon work done by me without any help. The resul ts have been discussed i th Dr. C. A. Mime and many of the experiments prung from th se di oussions.
} P.
r
lXJ1 J M f)f;--.l' <'1 'l.
.
.
..
..
.
.
.
. ..
...
..
... .
To
~nderful is he who speaks of it.
Intelligent is he who learns it.
Blessed is he who, taught by a good teaoher, is able to understand it.
CONTENTS
Pag
A 1M 0 F TilE IlifVF.,,8 TI GA TI 0 N •••••••••••• ••• ,... 1
MATERIALS AND l~THODS
.
.
.
.
.
.
.
. .
.
.
.
.
~.
.
.
.
.
.
..
.
...
.
.
.
... . .
. .
.. •• 2••
. . .
.
.
.
.
.
.
.
.
. . .
.
.
. .
. .
...
.
.
.
.
.
..
.
.
.
...
.
.
.
.
.
.
.
.
. .
.
.. 2N eone. tal thym.ectonty' ". ... 2
Splenectomy ••••••••••••••••••••••••••••••••••••••••••• 2 ColI otion of urine from mice
.
.
.
..
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
2Footpad mea urements ••••••••••••••••••••• • • ••••••• 3 Viruses
..
.
.
.
.
.
. . .
.
.
.
. .
. .
.
. .
".
.
.
. .
.
.
.
..
.
...
.
.
.
,..
.
.
.
.
. . . .
.
.
3Virus titrations
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
. . .
.
..
.
. .
.
.
. .
4Poxvirus neutralization tests • • • • • • • • • • • • • • • • • • • • • • $ • • 5 Poxvirus haemagglutination inhibition~~ antibody titrations
...
.
.
.
..
..
.
".
.
.
.
.
.
.
.
. .
.
.
••
• • • • • • • • • • • • ••
5Haemolysin t1trations .•••••••••••••••••••••••••••••••• 6 Gel diffu ion tests
.
.
.
.
.
...
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
. .
•••••••• 6Fluoresoein conjugated antibody and immunofluorescent staining.. •••••••••••••••• ••••••••••••••••••••••••••• 6
Mouse emb~o fibroblast oultures •••••••••••••••••••••• 7 L cell cuI tures
.
.
. . . .
.
.
.
.
. . . .
.
...
. .
... .
. .
.
.
.
.
.
.
.
.
.
.
. . .
.
.. 7Chick embryo fibroblast oultures • • •
.
.
.
..
.
.
.
".
.
....
"..
8Peritoneal cell cultur s
. .
.
..
.
.
.
.
.
.
.
. .
.
.
.
...
.
.
.
.
. . .
.
.
..
.
.. 8~ethod of preparation of materials for interferon assay • ••••••••••.••• • ... dI • .. • • • • • • • • • • • • • • • • • • • .. .. • 10
STUDIES ON 'IRE IN:FLUENCE OF .AGE ON THE PATHOGENESIS
OF POXVI
nus
DISEASES •••••••••••••••••••••••••••••••••••••• 12General introduotion •••• o • • • • • • • • • • • • • • • • • • • • • • • • • • ~ • • 12
Introduction to the pr sent work •••••••••••••••••••••• 14
Susoeptibility of mice of d~fferent ages to poxviruses. 16
Lethal effects • • • • • • • • • • • • • • • • a 0 • • • • • • • • • • • • • • • • • 16
Infeotivity ••••••••••••••••••••••••••••••••••••••• 17
Pathogenesis ••••••••••••••••••• ~ ••••••••••••••••••••• 18 Growth of virus in young and adult mioe ••••••••••• 18 Immunofluoresoent and routine histologioal
studies ••••••••••••••••••••••••• a • • • • • • • • • • • • • • • • 20
Infeotion of liver by oiroulating virus ••••••••••• 22
HOst defence meohanisms • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 23 Immune response (a) Antibody respon e... 23
(b) Immune footsw lling and
resistance to ohallenge •••••• 24 Interfe on response ••••••• • • • • • • • • • • • • • a 0 • • • • • • • 24
Misoel1aneous faotors of possible importanoe in the
greater susceptibility of young mioe ••••••••••••••••• 25
Behaviour of peritoneal maorophages ••••••••••••••• 25
Ciroulating antigen ••••••••••••••••••••••••••• 27
Influence of intest1na1 organisms •••••••••••••••• 28
Attempts to depress the resista.noe of adul t mioe to
the lethal effeots of oowpox and eotre ' 1ia viruses $ . 30
Pag Effect of Thiotepa ••
.
.
. .
.
.
.
. .
.
.
.
.
. .
.
. .
.
.
.
~.
.
.
.
~.
.
33 Effect of splenectomy •••••••••••••••••••••••••••• 34Effect of cold environment ••
.
...
~.
.
.
.
.
.
.
..
.
.
.
.
.
35 Effeot of pregnancy .•••••••••••••••••••.•••••••••• 35Disous ion ••••••
.
..
.
.
.
.
.
.
. .
.
...
.
...
..
.
.
.
.
.
.
.
.
. . .
. .
.
.
.
. .
.
35 Interferon production ••••••••••••••••••••••••••••• 36 Immune re ponse. .
.
.
.
.
. . .
.
.
. .
.
...
.
. .
. .
.
.
.
.
.
.
.
.
.
....
.
.
38 Viraemia.
. .
.
.
.
.
.
.
.
.
... .
.
.
... .
.
.
.
.
.
.
.
.
. . .
.
.
.
.
.
.
.
..
.
.
.
42 Barriers.
.
.
.
.
. . . .
.
.
.
..
.
. . .
.
.
.
..
.
.
.
.
.
.
.
.
.
.
.
.
. .
... .
..
.
.
.
43Tissue susoeptibility •••••••••••••••••••••••••••• 45
Miscellaneous factors • • • • • • • • • • • • • • • • • • • • • • • • • • • a • 46
Neonatal thym otomy and other attempts to lower
the resistance of adult mioe to poxviruses ••• •••• 48
Cause of death •••••••••••••••••••••••••••••••• • •• 49
S Ul1lDlary ••••• .... It .. • • • • • • • • • • • • .. • • • • • • • .. .. • • • • • • • • • .. II • • • • 50
Age-dependent resistano 50
Neonatal thymectomy •••••••••••.•••••••••••••••••• 5
STO'DIES 0 THE IN RFERON RESPONSE OF MIe", 10 VIRUS J~
.
.
...
.
52G n ral introduotion • • • a • .. .. .. • • • .. • • .. .. • • • • • • .. • • • • • • • • • • • 52 Produo ion
.
.
. .
...
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
.
~.
..
.
. .
.
.
.
.
.
.
.
.
.
. .
.
53 Bl.osynthe is • • • • • • • • • • • • • • • • • • • • • • • • • ~ • • • • • • • • • 0 55 Effect of interferon and other agents onin rferon produotion •• ~ ••••••••••••••••••••• ~. •• 57 Properties of interf ron • •
.
.
.
..
.
.
.
.
.
. .
. .
.
.
..
.
...
. .
.
.
59Interferon and cytopathog nicity of viruses ••••••• 64
Role of interferon in virus diseas s • • • • • • • • • • • • • • 65 Introduotion to the pr sent exp riments ••••••••••••••• 67 Prelim1nary experiment •••••••••••••••••••••••••••••• 69
Comparison of the interferon response of
different strains of mice . a • • • • • • • • • • a . e • • •
Comparison of the plaque r duotion and CPE
• • • • • • 69
1nhibition methods for int rferon assay •••••••••• 70
Comparison of Sen[iki Forest and
enoephalomyo-carditis viruses •••••••••••••••••••••••••••••••••• 71
Compar1son of mouse embryo fibroblast and
L oel1s ••••••••••••••••••••••••••••••••••••••••••• 71
Attempt to 1norease the sens1tivity of the
plaque reduotion test in JlX)use embryo fibroblasts.. 71
R produo bility of the plaque reduction test •••••• 72 Therl'OO ta bili ty 0
r
moue interf ron • • • • • • • • •• •••• 72Importanoe of removing particles of inducing virus
from interferon preparations •••••••••••••••••••••• 73
Clearanoe of intravenously injeoted influenza
virus ••• • • • • • • • • • • • • • • e • • • • • • • • • • • • • • • • • • • • • • • • • 74
Absence of interferon-like inhibitors in blood
and tissue .* •••••••••••••••••••••••••••••••• 74 Absence of effeot of normal blood and issues on
interferon activity •••••••••••••••••••••••••••••• 74
Fate of ntravenouslyadmini tered int rreron ••••••• 75
Di appearanc of int rferon from th circulation.. 75
Pres nee of injeoted interferon in th tissues ••• 75
Page
Distribution of interferon 1n virus-infeoted mic ••••• 78
'/e t Nile virus •• ~ ... " .... • • • • • • • • • • • • • 79
Ectromelia virus (a) Footpad injection •••••••••••• 79
(b) Intravenous injection •••••••• 80
Interreron pr duction in ndce after the intravenous
injection of influenza virus ••••••••••••••••••••• ".,,~ 80 The height and duration of the interferon response. 81
The relationship between particle dose and
interferon re pon e •••••••••••••••••••••••• ~~.... 81 Interreron response fter
two
doses of virus • • • • • •of interferon as Production nd distributio
determined at 4~ hours •• • ••••••••••••••••••••••
lnt rferon olearance in mice tre ted with
82
83
influenza virus •••••••••••••••••••• " ••••••••••••• 84
tudies on the possible sources of interferon pres nt
in mice injeoted with influenza virus •••• " •••••••••••• 85
Blood •••••••••••••••••••••••••••••••• "." ••••
.
.
". .
85Sple n •••••••••••••••••••••••••••••••••••••••••••• 87
Liver ... .
.
.
.
.
.
.
.
.
.
" ,.
.
.
.
.
.
. . .
. .
.
.
. .
.
" ".
.
.
.
88 Histologioal study of the influenoe of interferon onthe cour e of ectromelia virus infeotion in mioe •••• ~. 90 Interferon produotion by peritoneal oells ••••••••••••• 93
Importanoe of the induoing virus ... 94
Induction of interferon production by Newc stle
disease viru •• ,,~ ••••• ~ ••• ~. • ••• ~ ••• • • • • 98
Sensitivity of maorophe.ges to the aotl.on of
interferon ••••••••••••••••••••••••••••• • •••••••• 99
D1seU6sion •• • •••••••• • ••• "
.
.
.
.
.
.,.
...
...,...,....
100the mouse ••••••••••••••••••••••••••••••••••••••••• 102 Produotion 0 f 1n terferon in vi \TO • • • • • • • • • • • • • • • • • 104
Rapid interferon r spon e to intravenously
1njeoted viruse •••••••••••••••••••••••••••••••••• 106
Souroes of the rapid interferon
.
.,.
.
.
. .
.
.
.
.
.
.
.
.
.
.
.
108 lAacrophages •••••••••••••••••••••••••••••••••• 108 Liver •••••••••••••••••••••••••••••••• • • • • • • • • I I I Spleen •••••••••••••••••••••••••••••••••••••••• 112Blood oe11s • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 113 Lllng •••••••••••••••••••••••••• ., ••••••••••••••• 114 Action of interferon 1n vi 0 against ectromelia
virus infection ••••••••••••••••••••••••••••••••••• 114 S~r~ •••••••••••••••••••••••• .,., ••••••••••••••••••••• 115
CONCLUDING REMARKS
• • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • • 118BIBLIOG~HY
••••••••••••••••••••••••••••••••••••••••••••••
121The course of a virus disease in an animal is determined by the
in r etion of the virus with many host factors which include the
specific l.nnnune response and a variety 0 f known and unknown
non-specific influences. In addition to the immune response, the age of
the host and interferon produotion by the host are likely to be
important. The present work describes a study of the influence of
age on poxvirus diseases, and experiments on the origin, fate, and
action of interferon.
The a~ of the investigation was thus twofold~ Firstly, it
sought to find out the reason for the greater susceptibili~ of
young mice to the lethal effects 0 f poxviruses. Secondly , it
attempted to determine the sites of interferon production in
virus-infected mice, the fate of interferon in the body and the effect of
interf ron on the oourse of ectromelia virus infection in mioe.
Since the investigation had two different though not mutually
exclusive objectives, this thesl.s is presented in two parts. The
first part deals with age-dep ndent resi tance of mice to the lethal
eff cts of poxviruses and the second with studies on the produotion,
Mice - Multicoloured, outbred mioe of th alter and ~iza H 11
In tl. tute train were u ed in m st experim nts Inbred mc 0
r
theC
57Bl nd eBA trains were us d in some :xp riments. Mice of th
C
S7Bl and
eRA
trains wer small r than Hall Institute mice of thearne age (T bl e 2).
Whil mice used in I ny experiment were reared under ordinary
(0) conditions without sp oia1 preoautions, mioe for some xperiments
wer rear d under low pathogen (LP) condition. The LP eolon~ of
utbred and nbred mice were start d with mioe del~ver d by Cae arean
eati n and nursed by rat or LP muse foster thers. The LP mice
were handled i.n special ro ms by a single p r on until used in the
xperiment •
In the experim nts on enterio baot ria (page 28) and on e
-na 1 thymectomy (pages 30 to 33 ), the mouse food oube contain d
0.09% chI r ~cetin and 0.05% achro~oin.
Neonatal t ymectoSf - This as done by M1.11er's
thod (1960).
Th flb enc the thymu we. verified at the end of the expert nt by utop y end maoro copic examine. ton.Spl
n
etomw
-
Thi
W 8 don by ext rioriz~ng the spleen thr ugha retrop riton a1 incision nd r moving it fter ligatin~ th pediel •
Th bsenoe f auxi11ia~ p1eens 8 check d at the end f th
xp r nt by autop y and ero epic examina ti n.
Colleotion of urine f m mioe - This s don by kneadin~ the
3
Footpad me ere me sured by
the techniqu de crlbed by Aokerman (1964) with dial g uge c lipers
(Schn 11 taster, Sy tam Kroplin, 'IYPe A .02 T B.C. K "plin G.m. b.h.,
Sohluchtern, Res en, Gar ny), which read to 0 05 mm.
Viruse - (a) Ectromelia virus - The two strains of ectromeli
virus used have been d soribed by Fenner (1949b). The Hempste d
mous train deriving by mouse pas sag only from the 0 riginal viru-lent strain i olat d by ~rchel (1930) was passed onoe in egg and u ad s a chorioallantoic membrane preparation. The Hampste d egg
train is much Ie s virul nt than the Hampstead mou e strain from
whioh
it
was d rived by serial pas age in eggs. It had been pa sedabout 60 times when used by ~enner (1949b) and an unknown number of
times S1nce then. It was used as a chorioallantoic mbrane pre
ara-tion.
(b) Cowp x and v ccinia Tirus s - The Brighton s train of cowpox · ru
nd the V-MH end 7N strains of vaccinia virus (Fenner, 1958) w re
u ed as chorioallanwic membrane pr parations.
(0) Influenza virus - Fully nrectious allantoic fluid cont ining the
PR 8 strain of J.nfluenza viru s us ed in mo st expe riment s, but
ohicken red oell el tea of the BEL strain stor d at 20 er used in
me experJ.ments. Th se eluates did not contain detectable amunts
of infeotious virus but h~d at Ie t 104 un1ts of baa gglutin n per (d) Sendai r - The Sendai t in of parainfluenza 1 virus obtain d
from Dr. D. O. White Ibourne, s used and stock virus W oontei ned
in allantoic fluid from infected eggs.
(e) N castle dis ase virus - A stra1n of Newoe. tle dise se -rus
obtained from Dr E. L Frenoh Melbourne, s used and stock vi us
s contain d in allantoic fluid from infected eggs.
(f) e t Nile virus - A suokling use brain preparation of the
Sara-fend train, p sed six times n mouse brains ince 1so1ation, as
u ad.
(g) Semlik1 Fore t virus - A strain of virus obtained from the
Rockefeller oundation Virus Laborator,y and passed 20 time in moue
brains and onoe in ohick embryo fibroblasts sinoe isolationg was used.
Stock virus w s contained in the m dium from infected chiok embryo
fibrobla ts. Infeoted adult mouse seMlln coll oted 24 hours aft r
intravenous inooulation of the iru was used in one experiment on
interferon produotion by .peritoneal 0 lis.
(h) Encephalolf.\V'Ccat'di tis virus- A strain of the virus obtained from
Dr. K. A. O. E1lem w s ueed. Stock virus was oontained in the dium
from inrected mouse embr,yo fibrobl ts.
Virus titrati ns - Poxvirusee ere titrated on the ohorio llantoic
membranes of ll-day old chick embryos by the teohnique of Watwood
tal. (1957) using 6 eggs per diluti n.
The
eggs were incubated at36° and p cks oounted on the second day in the case of 0
cainia virus I and on the third day in the case f eatro Ii r ..
e t i l e virus was titrated on ohiok embryo fibrobl ate by th
5
Infectiou and noninfectious prepa tiona 0 f infl u nza viru
ere titrated for hae gglutinin by testing s rial 2-fold dilution
for bility to agglutinate ch1cken red oells. The ndpoint w the
highest dilut on at hieh partial haema glutination oeeurr d; th
reoiprooal of the ndpoint dilution was taken as th number of
ha gglutinating units (HAU) pre ent in 0.25 ~ (test volume) of th
undilu d material. The numb r f influ nza 'Virus partiel s pr ent
in a gi en dose was caloulated on the basis of a particle : HAU ratio
of 107 : 1 (Faz lea de St.Groth and Webster. 1963) ..
Poxvirus n utr lization tests - Sar w re tested fbr the pres noe
of n utralizing antibody using the V-MH strain of vacoinia virus as
antigen. S ra were diluted 1 in 5, except when otherwise tated and
ml.xed with equal volume of a virus dilution adjusted to contain
bout 100 pf'u of the virus; nOrln8.1 mous serum was used f r the virus
oontrol. E oh mixture s incubated at 370 for 90 minutes and then
1nooulated 1nto 6 eggs. The neutral zing power of a serumw s expr sa d
s the per cent reduotion in oak hen compared ith the con Is.
Po xvi rUB ha magglutin tion inhibition (HAl) antibo~y titr, ti ns
-S ra from oowpox infected miee were heated at 56 for 30 minutes and
soreened f; r th pr 8 no of nonspeoific ha gglutinin8~ aoh arum
or the te t d luted in 2- 01 step in 0.25 ml vol Ulnes f
diluent (1% normal rabbit serum. in no mal ine) i ersp:x
hae gglutin ti n trays. To oh oup, 2 HAU 0 cowpox virus r the
for 20 minutes One drop (0.025 ml)
r
a 5% sp nsion of vacoini-u oeptlble fowl red cells as added 'bJ e ch cup and after inoubation
for 20 mdnut s at 370s th t ys were haken and re1noubated for 40
minut s before readi g. 'I'he titr W' s express d as the eciprocal of
the highest diluti n of the s rum which partially inhibits ha magglutin
a-tl n.
Haemoly in titr t1 ns - Sera for the test re diluted in 2-fold
tep in 0.25 ml volumes of calcium-magnesium aline, mixed ith 4
uni ts of guinea pip; serum complement (0.025 ml) end a 2% suspension of
sheep red c 11s (0.05 ml).
The
trays ere shaken ell, inoubated at370 and re d after EO minutes It !he titre was xpr s ad s the
oipro-oal of the highest dilution of the erum at which parti 1 haemolysis
oocurred
Gel diffusion test - The tests ere done on micros ope s11d s and
the diffusion medium was a mixture of equal parts of 3% agar, 3%
odium azide and 0.9% EDTA. Pr cipitin lin ere seen after 48 hours
at room tempe ature in a humid oh.alrh r. The gel lay rs w r Wi h d
and stained with Biebrich c rlet fbr better visualiz ti n.
Fluore 0 in conjugate~ antibody and immunoflu rescent at ining
-The globulin in a potent nti ocinia serum were precipitat d with
methanol, redia 0 ved and oonjugated i th fluore oein isothiocya te
(Syl'Vane. Chemical Company Orang New J rsey J U.S. ). Exe ss
fl uore 0 in was remQ-ved by di ly i against buffe, d salin (pH 7.2
at 40 The conjugate we. then ad orbed with rabbit I1ver nd c 10k
7
nd u ed after the ddition of one-tenth velum of Rhodamin
oonju-gated bovin serum albumin (Microbiologioal Associates, Bethesda,
ryland, U.S.-A.).
Ti sues for immunofluoresoent examination were snap-frozen in
liquid-nitrogen-cooled contain rs, and sectioned on a cryostat at 6
f·
~e sections were dried under a fan, fixed l.n aoeton and stained.
Fluor soent ob ervations wer made with a Zeiss microscope
quipped for fluoresc nt mioroscopy, using an Osram HB 200 m rcury
vapour lamp as the ultraviolet sourc •
Mouse embryo fibroblast cuI tures - Secondary cuI tures were made
from 15- to 16-day nnuse embryo in 5 cm petri di hes by seeding
6
2 x 10 cells in 5 m1 of growth medium, or in screw-capped tub s by
eeding 4 x 105 cells in 1 ~ of medium. The gro th medium was
Eagle's minimum asential medium containing 7% calf serum (heated at
56° for 30 minutes). The monolayers w re confluent and ready for use by 48 to 72 hours.
Xhe
petri dish cultur s used in the plaquea ays ere ov rlaid after inf ction wi th Earle's saline oontain·ng
0.5% laotalbumin hydrolysate, 0 1% Y ast xtract, 0.1% bovin pl sma
albumin fraotion V, 2% oalf s rum (heated at 56° for 30 minute),
0.0 d xtran and 0 75% agar.
L cell cuI ture - L oells of strain 929 obtain d from
Microbiologies. As oeiates, Bethesda, Maryland, UsS • • , wer cultured
5 om p tri dishes using the sam methods growth medium and overlay
Chiok embryo fibroblast oultures - Primary oultures of chiok
embryo fibroblasts ere grown from deoapitated II-day chiok emb~os n
5 om petri dishes. The growth medium as Hanks' ESS oontaining 0.8.%
laotalbumin hydrolysate and 8% oalf serum (heated at 560 for 30
minute). The agar overlay was the same as that used for mouse
emb~o fibroblasts. but did not oontain serum.
Peritoneal oell cultures - (a) Maorophage cultures s Peritoneal
washings were obtained from untreated (unstimulated) mice by the
technique described by Mims (1964b). The medium oonsisted of Hanks'
BSS th 20% calf serum (heated at 5So for 30 minutes) and 0.5%
laotalb~n hydrolysate. In some experiments, the oells were cultured
in "ring' chambers oonsisting of pyrex gla s rings sealed to
mioro-soope slides ith petroleum jelly (Cairns. 1960); the rings received
2 x 105 oell in 0.2 ml of medium. In other experiments, 2 to
6 x 106 oells were oultured 1n petri d1shes oonta1ning 5 m[ of medium,
or in sorew-capped bottles oontaining 8 to 10 ml of medium. After
1ncubation for several hours at 37 , when the maorophages had adhered
to glass, cells not stioking to glass were washed off (Mackaness, 1962).
For infeotion, virus as u ually added to the full volume 0 f the
medium, adsorptio in a small volume being carried out only hen
tated. At least 2 replicates ere used in eaoh test.
In fluore oent antibody studie , "ring!f cuI tures were incubated
wi h virus for 8 to 20 hours. The medium was then remowd and the
9
the cells were fixed in acetone and stained with f.luoresce~n-conjugated
antibody. At Ie st 400 cells ere examined for the pre ence of antigen
in each 0 the duplioate slides.
In the experiment to determine the e..m.ount of infeotious virus
produced by maorophages, cultures in petri dishes were used.
They
were 1nfected by 1ncubating with 106•3 pfu of attenuated ectromelia
virus in 0.1 m1 of medl.urn for 3 hours at 37°. The cul t-ures er then
ashed thrice, refed with 5 m1 of fresh medium, and incubated for a
further 24 hours at 37°. The whole oul tures wer then stored froz n
at _600 until titrated.
In the experiments on interferon production by macrophages, petri
di h and bottle cultures were used. Details of virus used and method
of infection re given in the section of interfer0n (Tables 26, 27). The
cultures were freeze-thawed before prooessing for 1nterferon a say.
(b) Stimulated peritoneal washings: Peritoneal exudates were induced
by the intraperi ton a1 injection of 1 to 2 ml 0 f sterile nutrient
broth (Dirao Laboratories , Detroit, Mi~higan, U. S.A.). Peri to neal
washings er oolleot d 20 to 24 hours later; on one occasion, a
dirf r ntial count showed 66% large mononucl ar cells, 23% small
mononuclear cells, 9.8% polymorphonuclear cells and 0.4% eosinophil '.
The c lIs er oul tored in petri dishes or in screw-capped bottles and
infected by adding viru to the medium wi thin 30 minutes of collection.
After appropri te incubation the cultures were freeze-tha ed, exoept
Method of preparation of materials for interferon assay
-aterials for interferon assay ere dialysed at 40 against glycine-HOI buffer (pH 2 0) overnight and returned to pH 7.2 by further dialysis against everal ohanges of Dulbecoo's PBS at 4°. The dialysed material was centr1fuged at lOO,OOOg for at least 60 minutes in a Spinco model L preparative ultracentrifuge, and the supernatant was assayed for
1nterferon activity. In some exper1ments on the sera of
influenza-treated mice, dialysis against acid buffer was omitted, since these sera were unlikely to contain inhibitory amounts of virus (this thesis, page 74).
Preliminary exper1ments showed that treatments involved in preparing material for interferon assay did not affect interferon
titreso A 10% suspension of brains of mice infected 11th West Nile virus was divided into 2 portions.
One
portion was processed as usual, while the other one was only centrifuged at lOO,OOOg for 90 minutesto remove the virus. Both.preparations gave identioal titr s of 800 units per ml. In another experiment, it me found that when stock
interferon was reprocessed, its titre remained unchanged.
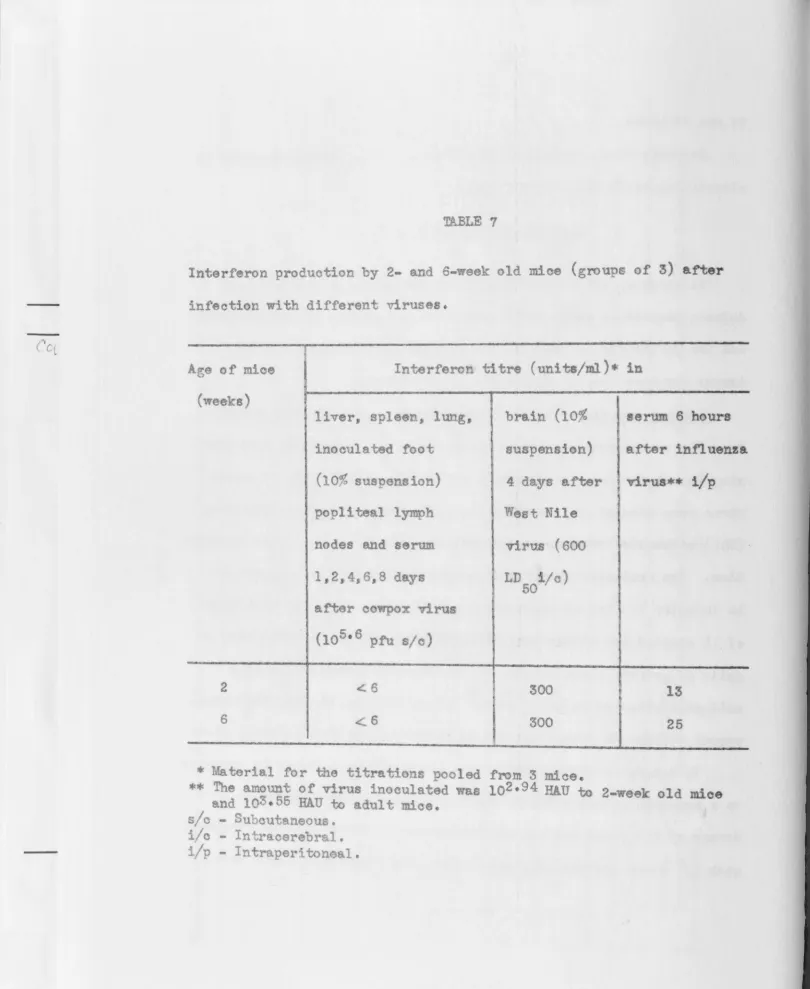
Standard interferon - A preparation of interferon was made from the bra1ns of adult mice infected ~ntraoerebrally with about 600 LD50 of West Nile virus and killed on the fourth day when the brains are
known to oontain large amounts of interferon (Finter, 1964d). A 10% suspension of the 1nfect d brains was processed for interferon
assay as describ d in the previous section (acid treatment and
80
70
c
0
10
60 EL-0
'+-Q.) :::::s
0-co c. 50
""'-0
c
0
...
.
-..0 ..c40 c
()--.!2
30' ! -. I
3 4 5
log10 reciproca
1 d
i1
ution [image:23.829.25.815.18.1008.2]~In~t~e~r~f~e~ro~n~~~~~~~ __ ~ - Interferon was assayed by th
fo1l ring stand rd procedur t
Confluent s condary oul tur s of mouse embry fibrobla t were
o
treated for 5 hours at 37 w1th 5 m[ of th appropriate dilution of
th D8 trial fb r a Bay in E gle' s medium. The fluid was then r moved
and the JOOnolayer inf eted with approxina t ly fl) pfu of Semllki
Forest virus in 0.1 ml of medium. After absorption for 75 minutes at
370 [I overlay medium was added. Two days later, the plates ere
stained with 1:20 000 neutral red for 2 to 3 hours at 37°, and plaques
counted. At lea t 5 plates wer used for each dilution and the average
pI que count oalculated in eaoh case. The percentage inhibition of
plaque counts in the treated plates, when oompar d to the average
plRque count in untreated control plates~ was oalculated for aoh
dilution. Results from a repres ntative test on standard interferon
illustrates the linear reI tionship observed between th p roentag
inhibi tion and the logarithm of he reciproca.l of the dilution over the
r ng been 30 and 80% inhibiti n (Fig_ 1). In each test a similar
graph s used to estimate the dilution at whioh th plaque count in
the treated plates would have been reduced to 50% of that in oontrol
plat • The reoiprooal of this dilution was d fined as the number of
un1ts of 1nterferon p sent in 5 ml of the starting naterial. Thus,
one unit of 1nterferon was defined as th amount contain d in 5 ~,
which reduoed the plaque count by 50%. For instanoe, if 5 ~ of a
1:100 dilution of an 1nterferon prepar tion reduced the plaque count
by 50%, the interferon titre of the preparation s taken to be 100
STUDIES ON AGE DEPENDENT RESISTANCE
GENERAL INTHDDUCTION
It has been recognized ibr
many
years that virus diseases agen rally more ev re in young than in adul t animal s. Andervont
(1927, 1929) reported that newly hatohed ohioks and 12-
to
14-day oldmioe were more susoeptible than adults
to
the lethal effects ofsub-outaneou ly inooulated neurovaccinia virus and herpes simplex virus
re pectiv lye Similar age-related effects have since been described
in the oase of several virus disease in a number of different speoies
of an~mal (Sigel, 1952). An important practioal applioation of
reater susoeptibility of young animals to virus dis as s is the u
of ohiok embryos and l.nfant
..
mo in virus ark •1 though young animals are gan rally more susceptible than adults
to virus disease, this is not invariably 0 (Sigel, 1952). For
instanc , the Lansing strain of poliomyelitis virus kills adul t mice
more re dily than young mice (Sabin, 1950; Da1ldorf, 1950); thi,
viru , however, is more lethal to baby cotton rat than to adu1 ts
(Pinto, 1948) 0 that the inver ion in age effect is not absolut •
dul t mioe are also more susoeptibl e than infanta to the 1 thal effects
of lymphocytic ohoriomeningi tis virus (1 i -mGY 1951) end thel.r gr ater
u 0 ptibility is thought
to
be due to their immune response beinguperior to that of infant mioe (Hotohin, 1962). Another 1nt resting
09. is that of the C nnecticut-5 train of Coxa one virus. Mioe
13
r i tant b een 5 nd 21 days and then moderat ly susceptibl unt11
8 or 9 we ks old (Pappenhei r e al., 1951); th newborn mic develop
Ie ions in y orgens, hil the old r mice in the s cond period of
leotive damag to the pancreas.
enhanoed usc ptibill.ty . how
The early de eriptive ork n g -dependent r sistance of animal s
to 'rlru di ea e soon foIl ed by ttempt to anal se the baS1C
elem nts constituting the U ge factor". Sabin and Olitsky (1937a~b;
1938 ,b,e) and S bin (1941) desar b d the development of barri rs to
the spr d of v iou1ar stom titis virus and equine encepha10my litis
virus through the body as th mice grew older. Subsequent work ith
a numb r f viru disee.ses ha made i t c1 ar that no single mechanism
aooount for e ry oa e of age-dependent re i ' tane to virus di a s~
For in tanc, hi1e the s r ad of v sieular tomatitis virus l.n adult
mic i limit d by barriers located at various plaoes in the body
(S bin and 01 tsky, 1937a,b; 1938a~b), re i tanoe of adult hamsters
and mice to mumps virus is thought to be due to their antibody respon
eing sup rior to that of susoeptible young animal (Over and
Kilham, 195 ; Ov r
an.
1954) and differences in re ist nee of mio ofdiff rent ag s to Coxsaokie Bl virus have be n hown to be paralleled
by differ no
1n
interferon roduction (He1neberg at al., 1984).Yet an ther po
who und that
ib1 meohani m ha been sugg at d by Johnson (1964)
rophages from resistant adult mice unlik tho from
usc tib1 young mice failed to spr ad herpes virus infection. That
y c rs ar involved in e-depend nt r istano to virus di e.. es
of re is nee doe not app ar at th same age for all viru dis ase •
Thi
view is alBo supported by the dditional obs rvation that theohang in u ceptibility to differ nt viru es t kee pI ce at dirf rent
rate , being gradual in oas like herpes simplex virus infeotion in
e (Johns n~ 1964) and rapid in other 11k yellow fever viru
infection in mice (Bugher, 1941). Therefore, several faotors acting
ingly or tog ther determine the ooourr nee of usoeptibility or
re i tano to virus diseases. Thee factor include th immune response
(Ov rman and Ki1ham 1953; Ove n, 1954), interferon produotion
(s ioki 196~J Helneberg at al., 1964), development of bar iers to
th pread of infeotion (Sabin and Olitsky, 1937a,b; 1938a,b,o;
Sabin, 1941; Johnson, 1964) and factor like body tempe tur (Enders
and Pe rson, 1941). The partioular combination op rating in any
tanc ould depend on the virus he host and the pa.ttern of their
interaotion
INTroDUCTIO TO 'ffiE PRESENT EXfERIMEN S
Study of ag -dependent resi t nee to virus di ease is likely to
be more profitable in the ease of virus disea es whose pathogenesis
i ell understood~ Sine the pathogenesis and histopathog n S1S of poxvirus inf otions in miee have b
(Fenner 1949a Roberts 1962, Mims
n p rtioularly
1964a), it a
ell studied
aonsid r d likely
th t useful ·nformation on the effect of age on virus diseases could
b obtained by a study of poxvirus infeotions in mice of diff rent
15
is known to ~ncre e th nore ing age. In an arly tudy 0 f e.g
-d P n-d n resi tance to poxviru s Green (1935) found th during a
rabbitpox pidemdo the great at mort lit.y occurr d in rabbits betwe n
the ~g s of 4 nd 9 weeks. Fenner (19490) found that 10- t l4-d Y
old ce w re killed more readily than adults by eotromelia virus. He
found the interval between virus in culation nd invasion of intern 1
organs to be horter in susceptible young than in resistant adult mdee,
and ooncluded that the young animals died due to the shorter time
ilable for antlb dy produotion and per ps les efficient antibody
r ponse. Oerskov nd Andersen (1938, quoted by Fenner, 19490) found
th t intrader.mally in oulated vaccini virus rapidly invaded internal
organs in 3-day old rabbits, but even small increases in age ere
coompanied by slower spread of the virus. Soh 11 (1960) r ported
th t adult mice of the C Bl strain resisted the leth 1 efr ots of
57
otromel ia irus better than adul t mie 0
r
three other strains lJ butth only vidence of superior resi t,ance in newborn C Bl mice as a
57
1 nger survi 1 tim th n in newborn mioe of a susceptible strain~
Mar nnikova nd Kaptsova (19 ) have reported that in racerebrally
inoculated variola virus killed mioe only if they are less than 12
days old~ The virus grew to higher titres in the brains of au oeptible
young mioe th in the brain f re istant adult mice, Virus B also
fo d ~n th lungs and kidney of young but not adult mice~
The pr sent work reports on the effect of age on the BUSO ptibility
of mice to the lethal ffeot of oowpox and ectromelia viru es. To
ee{
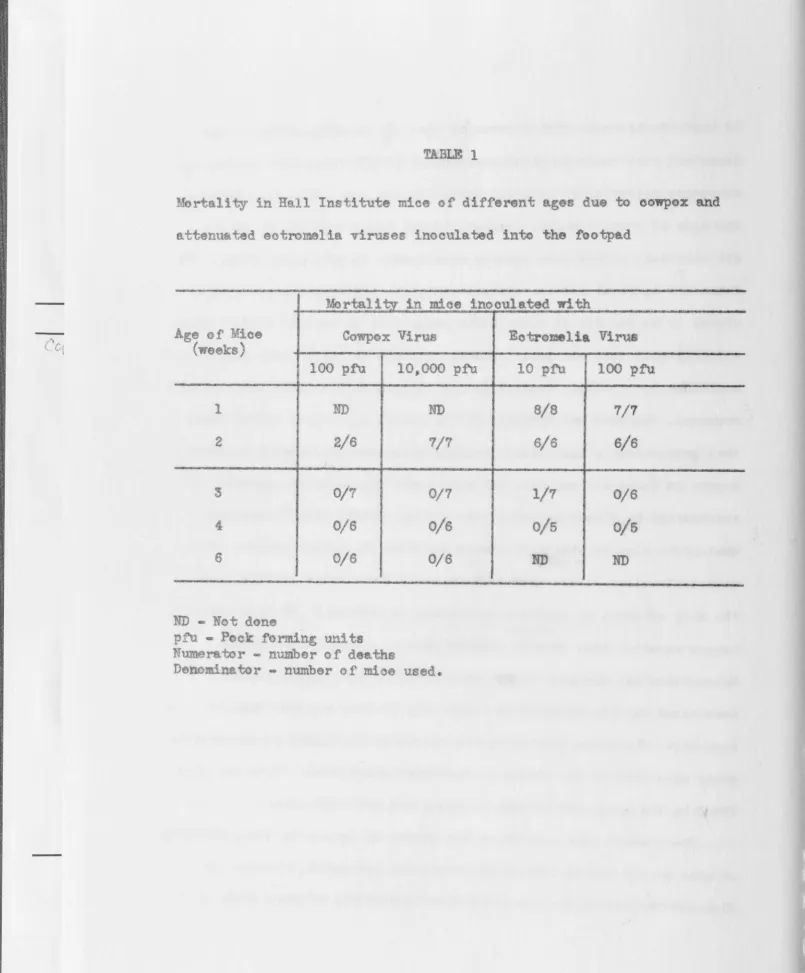
Mortality in Hall Institute mice of' different ages due to cowpox a.nd
attenuated eotromelia viruses inooulated into the footpad
-
-Mortal~t,y_ in Daoe inoculated with
Age of Mioe
(weeks) Cowpox Virus
100 pta 10,000 pf\1
1 ND ND
2 2/6 1/7
3 0/1 0/7
4 0/6 0/6
6 0/6 0/6
ND - Not don
pfu - Pock forming units
Numerator - number of deaths
Denominator - number of moe used.
-~-.
--Eotromelia Virus
10 ptu 100 pm
S/S 7/7
6/6 6/6
1/7 0/6
0/5 0/5
[image:30.840.20.825.18.990.2]16
oomparative study was made of the pathogenesis of the disease in young
and adult mice. Factors like the immune response, interferon produotion,
the ease of infection of maorophages and their abil1ty to spread
in-fection were studied in the two groups. Young mice were treated with
broad speotrum antibiotics to alter their intestinal flora and the
effeot of the trea~~nt on their susceptibility to poxviruses as
studied. Experiments were done to determl.ne whether large amounts of
viral nntigen were present in the oirculation of young mioe only.
Attempts were made to depress the resistanoe of adult mioe
to
cowpoxvirus by immunodepressive treatments including neonatal thymeotomy,
administration of the cytotoxio drug Thiotep and splenecto~.
Normal adult mice exposed to oold, and pregnant mioe were infected
l.th cowpox v~rus and observed to determine whether the resistanoe of
adult mioe was affected by oold or by pregnancy.
SUSCEPTIBILITY OF MICE OF DIFFERENT GES TO POXVIRUS S
Lethal effects - Hall Institute mioe of different ages ere
inoculated into the footpad with different doses of either cowpox
virus or the attenuated (Hampstead egg) strain of ectromelia virus.
The
mortali~r figures (Table 1) showed that these mice attained adu!tlevels of resistance to both viruses between 2 and 3 eeks of age.
Co ox virus was less virulent than the attenuated strain of e tromelia
virus. dult mice
cowpox virus even
re never killed by subcutaneously inoculated
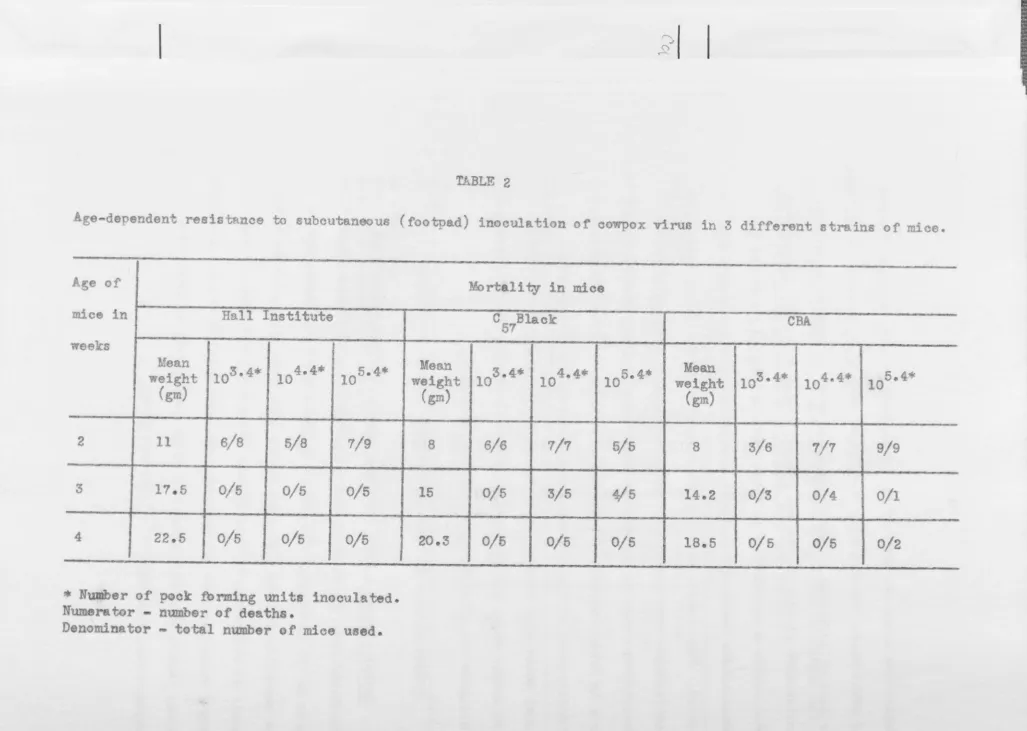
TABLE 2
ge~dependent resistance to subcutaneous (footpad) inoculation of oowpox virus in 3 different strains of mice.
sa: ....
.
_ _ :u:-. _ :ue.
Age of Mortal! ty in mice
mice in Hall Institute C
57Blaok CBA
weeks
Mean
103 • 4
*
4.4* 5.4* Mean 103 •4• 104 • 4*
105• 4*
Mean 103 • 4• 104 •4*
105 • 4•weight 10 10 weight weight
(gm) (gm) (gm)
2 11 6/8 5/8 7/9 8 6/6 7/7 5/5 8 3/6 7/7 9/9
:3 11.5 0/5 0/5 0/5 15 0/5 3/5 4/5 14.2 0/3 0/4 0/1
4 22.5 0/5 0/5 0/5 20.3 0/5 0/5 0/5 18.5 0/5 0/5 0/2
-
-
-
,. ...-r of pook fbr.mlng units inooulated. Numora tor - number of deaths.
[image:32.1050.18.1045.29.759.2]TABLE 3
eet
Suseeptibili ty of 2- and 6-week old mioe to cowpox virus inooulatedby different routes
Mioe lo~ number
or
pf'u* required to kill 50% of the mice ••subcutaneous intracerebral intranasal int ravenous
Young 2.6 1.4 2.3 ~4.9
Adult >7.3 308 6.9 ')' 5. 9
--- - "--
-• Pook forming units
[image:33.847.25.835.19.1007.2]In nother exp riment, 10-fold di1uti ns of cowpox virus ere
inoculated into the footpads of Hall Institute, C 7B1 and eBA m10e of
differ nt e.g d mortality rates d termined. Th. r suIts (Ti bl 2)
showed that H 11 Institut nd eBA mie 8.equired adult levels of re-is tanoe b tween 2 and 3 weeks 0 f ge, but C BI mioe did no t ttair:
57
adult level of r si tance until a aek lat r~ I all subsequent
xperim nt , 2-we k old Hall Insti tate mie (young mioeU) were oompared
wi h 6-week old mie ("adult mieen) of the same strain. Th experi nts
on g - 1 ted resi tance to poxvirus were don with cowpox viru ~
The rfect of drrd.ni tering cowpox virus by various route as
then xamined. S ri 1 dilutions of the virus were inoculated into
g up of young and adult Hall Institute mice using 6 mioe per group,
and the number of pock forming units required to kl.ll 50% of the mice
det rmined. Th r suIts (Table 3) how d that young mic re more
su
c ptibl what ver th route of inf otien.Infeot).
vi:t!Y -
The suboutan ou infecti vi ty 0 f oowpox virus inyoung and adult mice s compared by inoculating lO-fold diluti ns of
th viru into th ir footpa.ds. Primary fb tpad swellJ.ng was scor d
as clinioal evid noe of infection (Fenner, 1948) nd on the fourteenth
day urv1vors w re ch 11enged into the oppo ite foot with 106•7 pfu
of virul nt eotrom lia virus (Hrunp tead mouse strain)" Ocourr no of
immune eiling in th challenged foot (Fenner, 1948) was m asured at
9 hours and the mic observed for a furth r 21 day, for d ath du to
~
I
I
TABLE 4
Production of primary footpad swelling (infectivity), mortality, and of immunity (immune footpad
swelling and resistance to virulent eotromelia virus challenge) in 2- and 6-week old mice
followint; subcutaneous infection with cowpox virus
eClIJ.on
Pri inf'
r--
Cbal1en~e
of survivors (106•7 pfu of virulentrua)
loglO pfu·
I
Primary f'ootswellint; MortalityImmune
footswelling** Eesistance to challenge(survival)
Young Adult
Young
Adult Youn~ Adult Young Adult1.6
5/8
2/5 1/8 0/5 0/7 2/54/7
2/62.6 8/8
4/5
4/8
0/5 0/4 2/44/4
2/53.6 6/6 5/5 6/6 0/5 ••• 0/5
•
•• 1/54.6 5/5 5/5 4/5 0/5 1/1 2/5 1/1 4/5
5.6 2/2 5/5 2/2 0/5 ••• 3/5 • •• 3/5
"
-* Pock forming units
Measured 9 hours after challenge.
Numerator ... Number of reactors in the columns on footswell1ng, number of deaths in the colwms on
mortality and the number of survivors in the oolumns on survival.
Denomina.tor .,. Total number of mice used •
[image:35.1047.135.1037.20.820.2]7.3 (
nonl thaI to adult mice ev n when 10 pm were inoculated this thesis,
page 16 ) I but only 400 pfu killed half the young mice (T: ble 4). Both
groups er equally usceptible to infection as indicated by the
occurrence of primary footpad swelling (Table 4). None of the young
mice whioh surviv d 40 or 400 pfu of cowpox virus developed immune
footpad swelling when challenged but ome of the dults in comparabl
groups did; re istanee to the lethal effeots of the ohallenge was~
however, een in both the groups of mice (Table 4), even in those
surviving the smallest doses of oowpox virus.
Thus, as judged by the primary footpad swelling adult mioe are
a easily inf oted as young mioe by cowpox virus J but adults are mor
resi tant -00 the lethal effeots 0 f the virus. Survi ving young mice,
ho ever, fail d to d velop the immune footpad 5Welling r spon e although
they were able to resist challenge virus as re dily as adults.
PATHOGENESIS
In order to find a possible reason for the lethality of cowpox
virus in young but not in adult mio , studies wer made of virus
gro1th and histopathogenesis in young and adult mioe infect d with
oowpox virus II
Gro th 0 f virus :Ln young and adul t mi 0 - Cowpox virus w s
:Lnocula ted l.n to the footpads of young and adul t nuce, and footp d
sw lling, and virus titres in various tissues ere d te~ned daily.
i: ... ~. . 4 .I ",4.-}o=',,-''',,:Ij, ,;1 ~";,
'-n~'
h..
no:;;;~
:f , .
~~L':": . C'
,'~.'
\~' . .:
~~ , :
~.:~_: ~r
If·:. ,
~U· , t~i '.
l~ i .'
ll~ , ~
i
k~·:
i
n' "~ -'J ~.~'
ll:,', 'j
'a·
lt :;:
;, " ... ~
It' D~:\;'I:~: .,'
a{····c.
i~·:'~ .~ . . \'.;.' .;. ~~~~; ", ~
I,
,,'
..
'
f ~"'-i'" ~
l~~'"
",
l
~~U
~-l'" )
~ t·~:. ~,.:
:-It,·,!
" .. ""
'i.!.), ",
~r'.:'f
;\1-' .,
~
' :f', ~
f ':~~
. / t· ," ~f':'.:,':;
~, ...• ~;. ~; "
ll
l
~': .:~
i..,.'~' =r.",-, '
~i.' .
, f'~
'i' : ":",' ;'~.' ','
,
,.£'
H~:;';: 11' ,.,
,?-\.;.~:
k;:'! .
I
'
i'::",
:~ .. .-'~, " '<, H '
l
'~::,
.' .. '\;
, ~. ',- " f -;' •.• h ••. , ' , ' . ~: ;-~
;~'. " ' 1".' f'
",",
IJ·,:'
1
'
:;::::,
~ ~ '. f,
I -. ~:. "..
l ~':','~.
.I.' ,.'
~;'.i <
l
"-:::~ .,., .' ~ .. "", ..'"
.d·,
ii\.'
F~ "
tit
1\~:~
~'r~: ..
;1' ~,\ ,;
: f·'i.'
Ji; ,~
H,:-:
I
t' • i t . . ~l
i:'::
.
,
r, '.
~\'r,.>
~1;~"
."j'.
It'll, .. ~ .'
, ~n:.<i
. , ..
":S' }
~
~~k
~
..
,:r ~;,; , ~ ... ~: ' , I '
I
Ht~:'
~'; '~;
,~-:
.
>: " . )'f.'" .r. :: '
1\',', "
g' :~'; '; \ t ' .
!~:
h
i'.
' f'
,,· .
f
I:".::·'·
)t, •.: ..
~~'. :.
~' ,
~
..~.:
.f
'
t ..
~, .
,
,-;, :J;'~J:;l~~'("'~'-;' ••
~! 'l.""t"~i"~I·.i~~"J..:i'!
' ' ; ' ' p ;; • ; ; ; : ~ I' T • ; '-; to; r" '"1, • -;. ': • ~.l~ " ~ .. ' , 7 i ' \ . ;. . . . - , '
Footpad 6.2
Inoculum: 10 PfU
300..-/
j150
~
L
",0---,,"
~5
o
/0 " , '_ _ 0.""._0//
,
"
,
'
I
Footpad 2
4·
Inoculum:lO pfu
/
_. 0
-112I /'
/.... ..."
" """""~
:::J ~"
I*'''
,,0
01
b
Liver 2
-
E 8:::J '+-c. $26 0> o Q) .-E 4
+-'
~
...
.:; 2
4·
Inoculum: 10 Pfu
0/0/0_0
Spleen 5.3
Inoculum:10 Pfu
--
E :::J ~ $2 8' ..., 4~ .~ ... (/) 2.~ >0' I ! I - I I I I ! t - ! 10
o 2 3 .{ <; J. 0 2 3 4
r-after infect ion
- - - young mice - - - a d u l t mice
Fi~. 2. Results from 3 separate experiments showing the
development of footpad swelling and the growth of virus
in the inoculated footpad, liver and spleen, in young
and adul t mico inocula ted subcutaneous 1y wi th cowpox rus (virus t1ttes are expressed as the logarithm of
presented in Fig 2. Virus gro th was more rapid in the feet of
young mioe and was acoompani d by a gr ater d gree of footpad w ling.
P ak virus titr s in the feet of young mic ere higher than in adult
feet and after 4 d Y the titre fell in adult feet but not in the
t of young mioe. The liver and spleens of young mioe oontained
ubstantial amounts of virus J but very little was pres nt in these
organ in adult mice. In young mioe the infection also involved the
kin 0 f the general body surfaoe; a large area 0 f the skin taken from
the baok and abdom n of a siok young mouse oontained 106 pfu per gram,
and fooi of infeet10n in the skin were readily detected by
immuno-fluoresoent stain1ng in young mice but not in adults. In another
imilar experiment hole blood from young mioe was found to have a
virus titre of 104 pf'u per ml on the fourth day of infeotion and lesser
amounts on the sixth and ighth days· no vi rus as found in the blood
of adult mice at oorresponding times.
en 00 pox virus s inJected intravenously and blood virus
t tr deternun d a. t interval s, i t s found the. t virus was cleared
qually well by young and adult mice. Follow1ng an inJection of
106 pfu, 99% 0 f the injected virus was clear d wi thin 5 minutes by
young mce. This rate i oomparable to the olearanoe te of
pox-virus in adult mioe ( . , 1959a.). Therefore, th observed dir~ renoe
in vi r emie. b and adult mio was not due to differeno in
their ability to 01 r V1ru from th ciroulation.
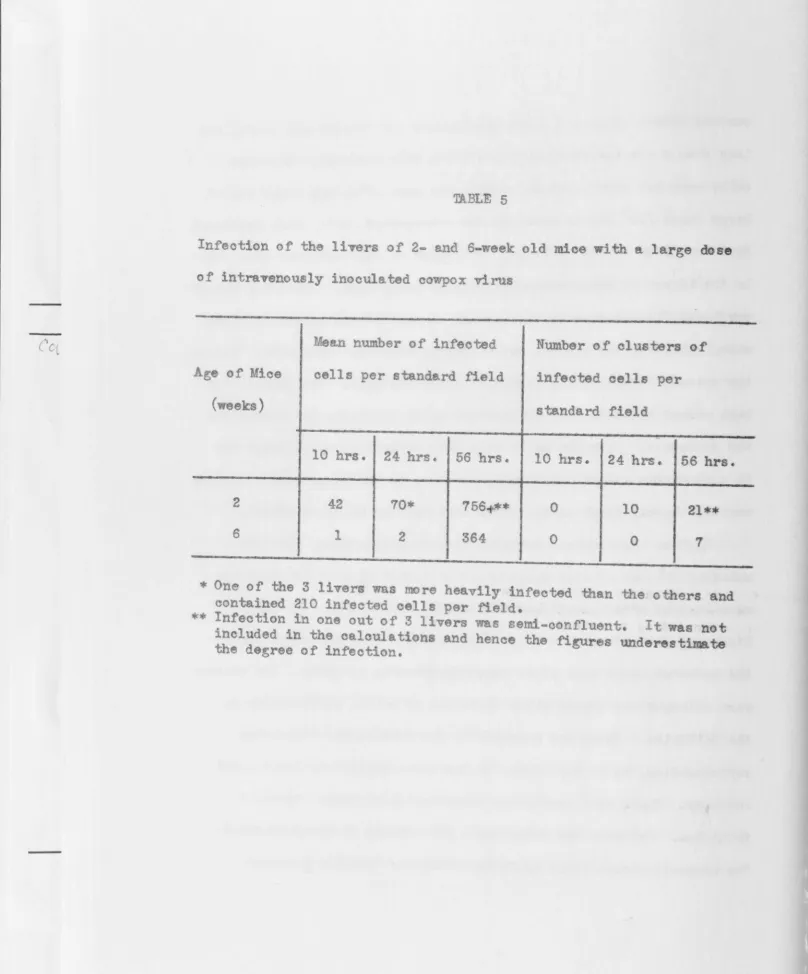
20
sub tanti 11y gre t r virus multiplication at the inoculation 6ite~
virae~ and s~gnifioant invasion of the internal organs~ The virus
titres in th liv r and spleen h ever, were muoh 1 ss than tho e
found by enner (1948) in the organs of adult mice dying of aoute lOOusepox.
Immunofluoresoent and routine histological studie - Experim nts
were carried out to determin whether the pattern of infeotion in
you g mice s differ nt from that in adults at th histologioal
level. Groups of young and adult mic were inooulated into the
foot-pa i h 105•9 pfu of cowpox virus and killed on the sixth day hen
the young mio er iok Eaoh animal was utopsied and speci ns for immunofluo so nt studi s collected. At utopsy, adult mioe
appeared no 1 exoept for footpad sw lling and enlargement of 100901
lymph nodes. The young mice had, in addition to the above changes,
numerous d~screte fooi of necrosis in the liver; the bone marro ,
01 arly vi ible in the tail and sternum, was a deeper red than in
normal mice of the same age. There were no skin lesions, except in
th inoculat d feet.
Immunofluore oent examination of seotions of liver, sp1een9
oardiac mu' 01 , brain lung and of blood smears from adults did not
reve 1 the pr sene f viru antige in a. 0 f th m. Th sick young
oe on the other hand, showed l~ted involv' ment of certain orga s.
In th liver th r were large, discr te fooi of rluore c nt hepatic
cen reI veins. Even so 1iv r involvement was limited nd fooa1, and
1 than a quarter of the hepatio cells were inf cted. lnr ct d
c lIs ere not seen in adult livers; and even after the inj otlon of
large dose (107 pfu or more) by the intravenou route, when scattered
fooi were seen in adult liv rs the number of foci seen was 1 ss than
in the liver of subcutaneously inf cted young mioe. There was almost
oonflu nt fluoresc nce on the serosal surface of the spleen of young
moe, involving the capsul and extending down the trabeculae. Within
th spleen, infection was oonfined to the red pulp. The skin of the
b ok showed discrete foci of infected oells involving the dermis and
the pide~s. Infeot d oells were also pres nt in the kidn ys and
in bone marro smear but rarely if ever, in the lung, brain, oardiac
musole thymu I lymph nodes, intestinal wall or in blood mears.
Bbutine hi tologioal sect~ons from formalin-fixedj
paraffin-mb dded tissues 0
r
sick young mice and normal mioe 0 f the s me agew re stained ith haematoxylin and eosin and examined. In the
liver of s10k young mice th re were neorotio foci corresponding to
the infec ad area en after immunofluor c nt taining. The spleens
re enlarged and sho ad gross depletion of oells, partioularly in
the follicle. Th re as necrosis in the oapsule and trabeoulae
oorrespond1ng as in the liv r, to th areas stained by fluorescent
antibody. There re hardly any morphologioally intaot lymphoid
follioles Pykno i nd karyorhexis were oommon in megakaryooytese
ss-.... 'H·' :~ni~!t~l .l...l;, . ,l~"l"-, '
a~,
,
r ;'
" ~L~':~:'i',;, .', ",: }-, .... ' t
~ ... " .'
<', ',,'
'.i:~~" .: !i~',' ~::
Ii,::' "
lV',
~i""
t
~,':,
,
~
~" ~,. ~ .. ' r,'
I
tt ~ l ~ ~,: ,;; ~ ~ :' ~
1~~~·'~
,tc, ,;
1
,;,':,-;
t::·,::; [image:41.847.25.833.22.996.2]~~,~~.{ t~·:·~·
r
!h,,:\ I~,: .• li, p:! .,:': ,. ii
,=
U/!
-I~' t'.·-',1, ·II 'i!t:';·jO'!
fi~' '.~ j
\';'~ ;',
I
l' .,. t;',:,.~ .,:, ,. ~'L"":w;:::;
~:~ .,::
1_ .• -:
it ,',
t
l ' 't
L:
, ".:
i t;':~:' ...
c-~
!1r:;';i~':" :
" ~ .. ~
'
1<,:
i' ( :
If.~::~
1
', ,
.\ -.\:'I'
t,'.'~':"~
. :. ~'
< . ; .
-if . .' '
It.":-:" ' h'r'
I
I~':;:::: fo, .... ~
J .t:·~,:
~~.,';;,:'~
I· ,- . .' ~~~: :'r~
.:-t
IT"
'}~'i>
;I\~
:
'. ~ ';-'.
l
'l~t;
, . t"
i'; ,'~ ':~
l
,·'t:',.: · . ~ .., t': .,:
t~::
·
'I ; : .'
1T\"", ..
K~~:~
tt;:
~l':Y
" I, I. '
ti";
-# •.
~~f~:::
l
'
,
,." ,
~'.,.:t~·. ,.
! ~ ,",. :'
r'
it'>
i ~;;. l~:-i,.
.~) ~"
~~-:~ ;
~,.~ ,~
'C:
H
Fj
I
ij' , It; . > _'f'
I
f, ,
·
,.
,~
... : '. ".:." .
~ i:~
>
' , '
-J
':>.'
It":t.' '-: :
eet
f
.~.' :~~'"
• it:.: j"'''' ":!tt~ .uni.l,n·:.:.u
TABLE 5
Infeotion ot the livers of 2- and 6-week old mice with a large dose
of intravenously inocula ted cowpox virus
Mean number of infected Number of clusters of
Age of Mice cells per standard field infeoted cells per
(weeks) standa.rd field
-10 hrs. 24 hrs. 56 hrs. 10 brs. 24 hrs. 56 bra.
2 42 70* 756-tW* 0 10 21
"ofr
6 1 2 364 0
0 1