0022-538X/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Intracellular Expression of RNA Transcripts Complementary to the
Human Immunodeficiency Virus Type 1 gag Gene Inhibits
Viral Replication in Human CD4
1
Lymphocytes
GABOR VERES, SONIA ESCAICH,† JENNY BAKER, CARMEN BARSKE,‡ CRETON KALFOGLOU,
HEINI ILVES, HIDETO KANESHIMA,
ANDERNST BO
¨ HNLEIN*
Progenesys Program, Systemix Inc., Palo Alto, California 94304
Received 22 May 1996/Accepted 4 September 1996
Intracellular expression of antisense transcripts was evaluated for its potential to interfere with human
immunodeficiency virus type 1 (HIV-1) replication. Retroviral vectors encoding HIV-1
C
-gag
complementary
sequences downstream of a selectable gene (neo, puromycin gene, or Lyt2 gene) were stable and yielded high
titers. Human CEMSS T cells were transduced with amphotropic retroviral vectors to express RNA
comple-mentary to the
C
-gag
sequence of HIV-1. Replication of laboratory-adapted HIV-1 strains was inhibited by more
than 1 order of magnitude (log
10) in these transduced cells even at high inoculation doses (4
3
10
4
50% tissue
culture infective doses). Antisense-mediated anti-HIV efficacy was further demonstrated by survival of CD4
1cells in these cultures relative to controls. The level of anti-HIV-1 activity of the
C
-gag
antisense sequence
correlated with the length of the antisense transcript. Maximal anti-HIV efficacy was observed with
comple-mentary sequence more than 1,000 nucleotides long, whereas transcripts less than 400 nucleotides long failed
to inhibit HIV-1 replication. Expression of
C
-gag
antisense RNA also reduced HIV-1 JR-CSF replication 10-fold in
primary CD4
1lymphocytes. These results obtained with a T-cell line and primary peripheral blood
lympho-cytes indicate the potential of long antisense RNAs as an efficient anti-HIV-1 therapeutic agent for gene therapy.
Gene therapy approaches (intracellular immunization) were
proposed in 1988 (3) as an alternative treatment modality for
human immunodeficiency virus type 1 (HIV-1) infection.
Sev-eral therapeutic genes, including transdominant mutant
pro-teins (2, 4, 33, 34), RNA decoys (13, 32), ribozymes (21, 37),
and antisense sequences (7, 11, 28, 30), have been evaluated
preclinically (9). Multiple therapeutic genes (a transdominant
Rev gene and RNA decoys) have been evaluated previously
for their potential to inhibit HIV-1 replication in vitro (4, 5,
8). The results demonstrate that constitutive expression of
RevM10, a transdominant mutant of the regulatory HIV
pro-tein Rev (14), consistently blocks HIV-1 replication (4).
Sim-ilar results have been reported by others (2, 15), and in 1995,
several patients received genetically modified peripheral blood
lymphocytes (PBLs) carrying a transdominant Rev gene (20,
36).
Protein-based anti-HIV strategies might lead to induction of
an immune response directed against the gene-modified cells,
and recent results with transduced T lymphocytes provide
ex-perimental evidence supporting this hypothesis (25). Hence,
we focused our efforts to develop alternative therapeutic genes
to RevM10 on RNA-based approaches, which could be used
in appropriate vectors for clinical applications. Regulation of
gene expression by antisense RNA occurs naturally in both
pro-karyotes (19) and eupro-karyotes (31), and we explored whether
constitutive expression of antisense transcripts could inhibit
HIV-1 replication. Previously, intracellularly expressed
anti-sense RNAs targeted at tat, rev, and vpu (24), the 5
9
leader-gag
region (29), reverse transcriptase (16), primer-binding site
(11), and the packaging signal (
C
) sequences of HIV-1 have
been shown to inhibit viral replication in vitro for extended
periods. The level of inhibition observed in these studies was
variable, depending on the size, the target sequence, the
sec-ondary structure, and the expression level of the antisense
RNA. On the basis of these results, we decided to reevaluate a
previously published (7) long antisense RNA complementary
to
C
and the gag gene and to express it as part of the murine
leukemia virus (MLV) long terminal repeat (LTR)
promoter-directed retroviral transcript downstream of a selectable gene.
The CD4
1T-cell line CEMSS was transduced with retroviral
constructs expressing different lengths of the complementary
sequences, selected, and subsequently infected with HIV-1.
The results of our experiments indicate that intracellular
ex-pression of sequences complementary to the HIV-1 gag gene
inhibits HIV-1 replication of laboratory-adapted strains
effi-ciently over a range of inoculation doses (4
3
10
2to 4
3
10
450% tissue culture infective doses [TCID
50]). This effect is
based on specific reduction of the full-length, 9.0-kb HIV
mRNA which can anneal to the complementary recombinant
antisense transcript. Retroviral vectors expressing shorter
an-tisense sequences were less effective, resulting in a complete
loss of HIV-1 inhibition with transcripts shorter than 400
nu-cleotides (nt). Antisense-mediated inhibition of HIV-1
repli-cation was also demonstrated in primary T cells.
MATERIALS AND METHODS
Retroviral vector construction.The initial antisense and sense vectors were constructed by insertion of the 1,425-bp SacI-BglII fragment derived from the HIV-1 HXB2 isolate into the XhoI site of the pLP and pLN retroviral vector backbone (see Fig. 1). The neomycin-phosphotransferase gene (neo) of pLN (18) was replaced with the puromycin resistance gene fragment to generate the pLP vector. Deletion vectors were constructed by cloning the 475-bp SacI-PvuII subfragment corresponding to theCand 59end of the gag sequence and the 952-bp 39gag subfragment into the XhoI site of the pLN vector. The 39serial deletion fragments were generated by PCR using a fixed 59primer sequence. The 39ends of the fragments were defined by primers placed at 200-bp increments. The PCR fragments were 1,225, 1,025, 825, 625, 425, and 225 bp long and were
* Corresponding author. Mailing address: Systemix Inc., 3155 Porter
Dr., Palo Alto, CA 94304. Phone: (415) 813-5011. Fax: (415) 813-5101.
Electronic mail address: [email protected].
† Present address: Rho
ˆne-Poulenc Rorer, 94403 Vitry-sur-Seine,
France.
‡ Present address: Sandoz Pharma Ltd., Basel, Switzerland.
8792
on November 9, 2019 by guest
http://jvi.asm.org/
inserted as blunt-end fragments in the filled-in XhoI site. The 865- and 650-bp 59 deletion fragments were generated by PCR and inserted into the pLN vector as described above. Vectors for primary cell HIV infection experiments were gen-erated by exchanging the neo gene with the truncated mouse CD8 (Lyt2) cell surface marker (33). The 1,200-bp ClaI-BglII gag antisense fragment without the Csequence was inserted into the XhoI site of the pLLyt2 vector.
Retroviral vector packaging.Retroviral plasmids were transfected into the BOSC23 (22) packaging cell line by the calcium phosphate transfection method. Supernatant from the ecotropic packaging cell lines was used to transduce the amphotropic PA317 cells (17) by spinoculation (12). Stable cell populations were selected for drug resistance (puromycin, 10mg/ml; G418, 800mg/ml) or selected by magnetic bead (Dynal) enrichment for the Lyt2-expressing vectors by using biotinylated anti-mouse CD8 antibody. Retroviral titers were determined with NIH 3T3 cells by endpoint titration for the puromycin and neomycin resistance-carrying vectors and by fluorescence-activated cell sorter (FACS) analysis for the Lyt2 vectors (26).
Transduction of the human T-cell line CEMSS.The human CEMSS T-cell line was maintained in RPMI 1640 medium supplemented with 10% fetal calf serum. CEMSS cells were transduced with PA317-derived amphotropic super-natants by spinoculation (12) and selected with puromycin (2.5mg/ml) or G418 (800mg/ml). Stable, drug-resistant CEMSS populations or clones were analyzed by Northern (RNA) blotting to determine the steady-state RNA expression levels.
HIV infection assays.Transduced CEMSS cells (populations or clones) (23 105/ml) were inoculated with various doses (400 to 105TCID
50/ml) of HIV-1
(HXB3 or SF2) at room temperature for 2 h. The TCID50of the cell-free HIV-1
stock per ml was determined by endpoint titration as previously described (10). After a washing to remove the virus inoculum, cells (23105/ml) were seeded in
RPMI 1640 plus 10% fetal calf serum and incubated at 378C. Every 3 or 4 days, cell counts were determined (Coulter Counter) and 1 ml of culture supernatant was collected, cleared by centrifugation, and cryopreserved and stored at2708C. The cells were then diluted in fresh medium at 23105/ml. HIV-1 replication was
measured by determination of the p24 antigen (Ag) concentration in culture supernatants with an enzyme-linked immunosorbent assay (ELISA) kit (Dupont-NEN). HIV-1-inoculated cultures were maintained for 2 weeks or until the control cultures died.
CEMSS infectivity assay.To compare infectibility of the CEMSS cultures, pLN-C-gag/AS and pLN-C-gag/S vector-transduced cells were infected with an HIV-1 isolate carrying the hygromycin resistance gene in place of the nef open reading frame (8). Plating the cells in methylcellulose (MC) and using hygromy-cin selection allowed us to determine the infectibility of the sense and antisense cell populations (number of Hygrcells). Cells (500 and 5,000/ml) were seeded in
duplicate in MC with RPMI 1640 supplemented with 10% fetal calf serum to determine seeding efficiency. Three days later, hygromycin (Boehringer Mann-heim) was added at a final concentration of 400mg/ml to half of the cultures to determine HIV infection rates (number of Hygrcolonies). The MC colonies were
scored after 6 days without selection and after 12 days with hygromycin selection. Detection of proviral HIV-1 in infected cells.For PCR analysis, cells were lysed in a buffer containing 50 mM KCl, 10 mM Tris (pH 8.3), 2.5 MgCl2, 1%
Tween 20, 1% Nonidet P-40, and 100 mg of proteinase K per ml at 568C for 2 h. Rev-specific primers (B1, [59TCGATTAGTGAACGGATCCTT 39] and B2.1 [59 CTCCTGACTCCAATATTGCAG 39]) were used to detect HIV-1. Cell lysates were mixed with the PCR buffer (Boehringer Mannheim), 200 mM deoxynucleoside triphosphates (Pharmacia), 100 pmol of Rev primers, and 1 U of Taq polymerase (Boehringer Mannheim). After denaturation (5 min at 948C), the samples were subjected to 40 cycles of amplification consisting of 30 s at 948C, 30 at 628C, and 1 min at 728C and 10 min of elongation at 728C in a thermocycler (Perkin-Elmer 9600).
Detection of intracellular p24 Ag expression by FACS analysis.Transduced CEMSS cells expressing antisense and sense HIV-1 sequences were inoculated with HIV-1 (105TCID
50/106cells per ml for 2 h). At days 3 and 5, the cells were
fixed in 1.5% paraformaldehyde for 30 min on ice and then washed twice in phosphate-buffered saline (PBS). The fixed cells were stained with a fluorescein isothiocyanate-conjugated anti-p24 monoclonal antibody (Coulter KC57 in PBS plus 0.5 mg of digitonin per ml) for 40 min on ice. The cells were washed in PBS plus 5% Tween 20 and finally resuspended in 1% paraformaldehyde for FACS analysis.
RNA isolation and Northern blot analyses.Total cellular RNA was extracted with RNAzol (Cinna/Biotecx), fractionated on 1.2% agarose–2.2 M formalde-hyde gels, and transferred to Hybond N membranes (Amersham). Both prehy-bridization and hyprehy-bridization reactions were carried out in Rapid-hyb buffer (Amersham). Oligonucleotides (100 ng) were radiolabeled to a specific activity of 33108cpm/mg with terminal transferase (Boehringer Mannheim), and the
membranes were hybridized with a radiolabeled oligonucleotide probe (53106
cpm/ml) for 2 h and washed with 13SSC (0.15 M NaCl plus 0.015 M sodium citrate)–0.1% sodium dodecyl sulfate at 658C.
Transduction and HIV-1 infection of human PBLs.PBLs were isolated from healthy donor buffy coats by gradient centrifugation. Enriched CD41cells were obtained by labeling bulk PBLs with biotinylated anti-CD81and anti-CD191 antibodies and subjecting them to depletion with streptavidin-conjugated mag-netic beads (Dynabeads M-280; Dynal AS, Oslo, Norway). The enriched CD41 PBLs were stimulated with phytohemagglutinin (5mg/ml) ong-irradiated allo-geneic feeder cells for 72 h in Iscove’s modified Dulbecco modified Eagle me-dium. PBLs (23106) were transduced by spinoculation in the presence of
Polybrene (8mg/ml). After 48 h, the cells were analyzed for CD41and Lyt21 expression by flow cytometry using anti-CD4 fluorescein isothiocyanate-conju-gated and anti-CD8 phycoerythrin-conjuisothiocyanate-conju-gated monoclonal antibodies. Lyt21 -expressing PBLs were again enriched by magnetic bead selection. After the first enrichment, the PBLs were expanded, and the CD41Lyt21cells were isolated by FACS (Vantage; Beckton-Dickinson). After the second enrichment,.90% of the cell population was CD4 and Lyt2 positive. Primary CD41T cells (53104)
were inoculated with 600 TCID50of HIV-1 JR-CSF (6) per ml in quadruplicate
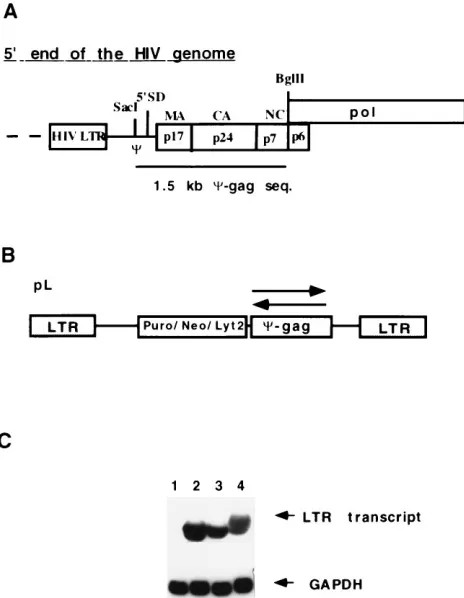
[image:2.612.62.294.70.369.2]FIG. 1. Structure and expression of retroviral vector carrying the antisense C-gag fragment. (A) Detailed map of the HIV-1 genome corresponding to the C-gag sequence. 59SD, 59splice donor site of the HIV-1 RNA; MA, CA, and NC, coding regions for the matrix, capsid, and nucleocapsid proteins, respec-tively, in the gag gene. The restriction enzymes used to excise the 1.5-kbC-gag frag-ment are indicated (SacI and BglII). seq., sequence. (B) General structure of the pL-based retroviral vectors encoding the antisense sequences. Puro, Neo, and Lyt2 are the selectable markers used with in the vectors (pLP, pLN, and pLLyt2, respectively). Arrows indicate the orientation of the inserted HIV-1 sequences. (C) Northern blot analyses of the antisense RNA expression in transduced CEMSS cell pools. The full-length LTR transcript carrying the antisense se-quence was detected with a gag-specific probe, and the lower panel was probed with a glyceraldehyde-3-phosphate dehydrogenase (GAPDH)-specific oligonucle-otide to confirm the amount of RNA loaded on the gel. Lane 1, CEMSS wild type; lane 2, pLP-C-gag vector; lane 3, pLN-C-gag vector; lane 4, pLLyt2-gag vector.
TABLE 1. Summary of the clonal challenge experiment
with the pLP-
C
-gag/AS constructs
aConstruct p24 (pg/106cells)
% CD41 cells
HIV DNA PCR resultb
pLP-
C
-gag/S
4
3
10
6–10
3
10
610–21
111
pLP-
C
-gag/AS
0–25
94–99
111
aTen CEMSS cell clones of each construct were infected with 400 TCID 50of
HIV-1 HXB3 per ml as described in Materials and Methods.
bHIV-1-specific DNA PCR was performed using Rev-specific primers.111, strong positive reaction.
V
OL. 70, 1996
INHIBITION OF HIV-1 REPLICATION WITH ANTISENSE RNA
8793
on November 9, 2019 by guest
http://jvi.asm.org/
4 days after the last restimulation of the cells. Half of the culture supernatant was exchanged daily for 9 days, supernatants were stored at2708C, and p24 Ag was determined by ELISA. Viable cells were counted by trypan blue exclusion 7 days after inoculation.
RESULTS
Constitutive expression of HIV-1
C
-gag
antisense sequences
in CEMSS cells.
The anti-HIV-1 efficacy of
C
-gag
complemen-tary transcripts (Fig. 1A) was evaluated in the CD4
1human
T-lymphoid CEMSS cell line. Retroviral vectors carrying the
sense (S) or antisense (AS)
C
-gag transcripts have been
de-scribed in detail in Materials and Methods. Initially, we used
the pLP vector with the puromycin gene as a selectable marker
(pLP-
C
-gag/S and pLP-
C
-gag/AS), but subsequently we
re-peated the experiments with the pLN vector with the neomycin
marker (pLN-
C
-gag/S and pLN-
C
-gag/AS vectors) to show
that the observed anti-HIV-1 effect is not specific to one
par-ticular vector construct. CEMSS cells were transduced with
PA317-derived amphotropic pLP-
C
-gag/S, pLP-
C
-gag/AS,
pLN-C
-gag/S, and pLN-
C
-gag/AS retroviral vectors (Fig. 1B) and
selected on puromycin (2.5
m
g/ml) or G418 (800
m
g/ml), and
stable CEMSS cell populations were established. The
ampho-tropic titers were comparable for the pLN and pLP vectors
(2
3
10
6to 5
3
10
6CFU/ml). The sense and antisense
C
-gag
sequences were part of the full-length viral RNA expressed
from the retroviral MLV LTR promoter. Comparable
steady-state RNA expression levels were detected by Northern blot
analyses with all vector constructs, regardless of the selectable
marker used (Fig. 1C). Vectors encoding the sense gag
mes-sage did not express detectable levels of Gag protein.
Inhibition of HIV-1 replication in CEMSS cell clones.
CEMSS cells were transduced with antisense or sense vectors,
and individual cell clones were isolated by limiting dilution. To
exclude clonal variation and differences unrelated to antisense
RNA expression, 10 CEMSS clones of pLP-
C
-gag/AS and 10
pLP-
C
-gag/S control clones were infected with HIV-1 HXB3.
All clones were at least 95% CD4
1as determined by flow
cytometry prior to HIV-1 infection (data not shown). Initially,
the CEMSS clones were inoculated with a low infectious dose
of HIV-1 HXB3 (4
3
10
2TCID
50
/2
3
10
5
cells per ml). All
CEMSS clones transduced with the antisense construct
dis-played remarkable resistance to HIV-1 replication, producing
between 0 and 25 pg of p24 per ml per 10
6cells at the end of
the experiment (day 22 postinoculation). In sharp contrast,
cultures of the 10 sense control clones produced between 4
3
10
6and 10
3
10
6pg of p24 per ml per 10
6cells at day 15, and
few live cells were present at this time. Expression of the
HIV-1 receptor molecule CD4 was unaltered after the first
round of infection in the 10 pLP-
C
-gag/AS CEMSS antisense
clones (Table 1). These cells were reinoculated with the same
viral load, and after an additional 17 days, the cells produced
p24 Ag levels which were slightly higher than in the first
infec-tion experiment but 3 orders of magnitude lower than those of
control cells, indicating that
C
-gag antisense expression
inhib-its HIV replication efficiently and persistently (data not shown).
DNA PCR amplification of all HIV-infected CEMSS
C
-gag/
AS clones confirmed the presence of HIV proviral genomes
(Table 1). Next, we repeated the HIV infection experiment
with a 100-fold-higher inoculation dose (4
3
10
4TCID
50
/2
3
10
5cells per ml) with 5 sense and 10 antisense CEMSS clones.
The CEMSS clones transduced with the sense construct
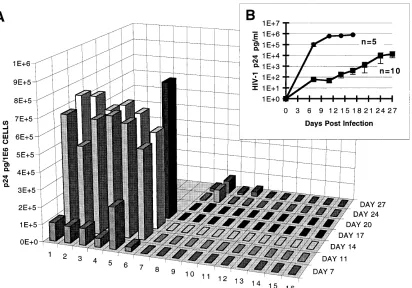
rap-FIG. 2. HIV-1 challenge of CEMSS clones expressing sense or antisenseC-gag RNA. (A) CEMSS cell clones (23105cells per ml) were inoculated with 43104
TCID50of HIV-1 HXB3 strain per ml. 1, CEMSS untransduced control; 2 to 6, CEMSS transduced with the sense vector; 7 to 16, CEMSS clones expressing theC-gag
fragment in antisense orientation. (B) Summary result of the clonal challenge experiment with CEMSS clones carrying the pLN-C-gag/S (F) and pLN-C-gag/AS (■) vectors.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.104.516.70.358.2]idly produced HIV-1 p24 Ag in the culture supernatants, which
were indistinguishable from those of the parental CEMSS cells
(Fig. 2). Seventeen days postinoculation, virtually all cells had
been eliminated by the cytopathic HIV-1 HXB3 isolate, and
the cultures were terminated. Only 4 of the 10 antisense clones
secreted detectable but markedly lower p24 Ag levels in this
time frame (Fig. 2A, lanes 7 to 10). The six other CEMSS
C
-gag/AS clones secreted only background p24 levels even at
this high inoculation dose (Fig. 2A, lanes 11 to 16). In
sum-mary, the average p24 Ag production of all CEMSS
C
-gag/AS
clones was more than 3 orders of magnitude lower than that of
the CEMSS sense control cells at day 15 postinoculation (Fig.
2B). Again, the majority of the CEMSS
C
-gag/AS cells were
viable and expressed the CD4 surface marker (89 to 99%).
C
-gag
antisense transcripts are effective in transduced
CEMSS cell populations.
Next, we inoculated drug-resistant
CEMSS cell populations, which better represent the average of
the transgene expression. CEMSS cell populations expressing
the
C
-gag transcripts in the sense or antisense orientation were
challenged with increasing doses of the HIV-1 HXB3 virus to
assess the potency of this strategy. HIV-1 replication kinetics
were accelerated with increasing inoculation doses (Fig. 3). In
the CEMSS sense control cells, the peak p24 secretion in the
culture supernatants was reached 11 days postinoculation with
the doses of 4
3
10
3and 4
3
10
4TCID
50
/ml, whereas no
plateau was reached at day 15 for the dose of 4
3
10
2TCID
50/ml (Fig. 3). HIV-1 spread was much slower in the
CEMSS antisense cultures at all inoculation doses (Fig. 3), and
the p24 levels in the antisense culture supernatants at day 11
were reduced by 2.9, 2.7, and 1.3 log
10with the doses of 4
3
10
2, 4
3
10
3, and 4
3
10
4TCID
50
/ml, respectively. The level of
inhibition was lower in the selected populations than in the
individual clones. This result probably reflects the range of
antisense RNA expression levels in the cell populations
rela-tive to that of the homogeneous clones, which can lead to
different replication kinetics. Similar results were observed
when CEMSS cell pools were infected with the less replicative
SF
2HIV-1 strain (data not shown).
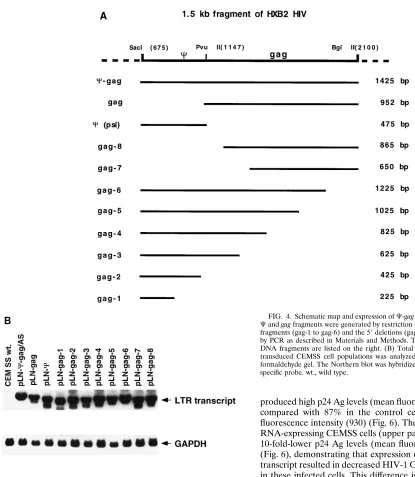
Minimal transcript length is required for optimal anti-HIV
efficacy.
To determine which part of the 1,425-nucleotide
C
-gag
antisense transcript was required for the antiviral activity, the
original SacI-BglII restriction fragment was dissected into a
475-bp
C
and a 952-bp gag sequence (Fig. 4A). In addition, 3
9
and 5
9
deletion mutants were generated by PCR amplification
and inserted in the pLN vector as described in Materials and
Methods (Fig. 4A). The steady-state levels of expression of the
different-length antisense RNAs in transduced CEMSS
popu-lations were comparable (Fig. 4B). CEMSS cells expressing
these different antisense fragments were inoculated with
HIV-1 HXB3 (4
3
10
2TCID
50
/ml). Deletion of HIV-1
C
from the
5
9
end of the antisense transcript had no measurable impact,
since the anti-HIV efficacy of the remaining 952-bp gag
frag-ment was identical to that of the full-length sequence (Fig.
5A). Expression of the antisense transcript to the
C
sequence
alone did not block HIV-1 replication (Fig. 5A) and was less
efficient than a comparable construct expressing similar-length
transcripts complementary to the gag sequence (gag-2, Fig.
5B). Analyses of all 3
9
deletions mutants of the antisense
C
-gag
sequence indicated an inverse correlation between the length
of the antisense fragments (gag-1 to gag-6) and the level of
HIV-1 p24 secretion in the culture supernatants of these cells
(Fig. 5B). Inhibition of HIV-1 replication appeared to be
max-imal when the
C
-gag antisense transcript was at least 952 nt
long. Seventeen days after infection, the highest percentage of
CD4
1cells was observed in cultures expressing the longest
C
-gag antisense sequence (97% of the gag-6 CEMSS cells
versus 2.7% of the gag-1 CEMSS cells). Deletions at the 5
9
end
of the antisense transcript were also constructed (Fig. 4A,
gag-7 and gag-8). Again, we observed decreased anti-HIV
ef-ficacy with shorter antisense transcripts, indicating that the size
of the antisense transcript rather than a specific subfragment is
responsible for the antiviral effect (Fig. 5B).
[image:4.612.69.283.70.513.2]Mode of action of the antisense
C
-gag
RNA.
To exclude
different infectivities of the sense- and
antisense-RNA-express-ing CEMSS populations, we directly measured the ability of
HIV-1 to infect these cells. First, a quantitative assay system
was developed to score survival of CEMSS cells in MC. For
this assay, we used an HIV-1 isolate encoding the hygromycin
resistance gene in place of the nef open reading frame
(HIV-Prohygro) (8). As a multiply spliced transcript, the HIV RNA
FIG. 3. Infection of transduced CEMSS cell populations with increasing HIV-1 doses. Mean p24 production in culture supernatants of CEMSS popula-tions challenged with increasing doses of HIV-1 HXB3 is shown. Results are shown for cells expressing sense (dashed lines) and antisense (solid lines) RNA. The experiments were done in duplicate.
V
OL. 70, 1996
INHIBITION OF HIV-1 REPLICATION WITH ANTISENSE RNA
8795
on November 9, 2019 by guest
http://jvi.asm.org/
encoding hygromycin resistance cannot anneal to the antisense
transcript, and following infection, HIV-Prohygro-infected
CEMSS cells give rise to drug-resistant colonies in MC. Using
this experimental system, we did not observe any differences in
HIV-1 infection between the CEMSS sense and antisense
pop-ulations (Table 2), indicating that HIV-1 can infect the two
cultures equally well. This result also demonstrates that the
antisense transcript acts at the posttranscriptional level and
cannot prevent integration of HIV-1 proviral DNA. To directly
test the impact of the antisense RNA on HIV-1 structural
protein expression, we measured intracellular HIV-1 p24 Ag
levels in infected CEMSS populations by flow cytometry.
CEMSS cell pools expressing the full-length (1,425-bp)
C
-gag
fragment in the sense or antisense orientation were inoculated
with 10
5TCID
50
of HIV-1 HXB3 per 10
6cells (multiplicity of
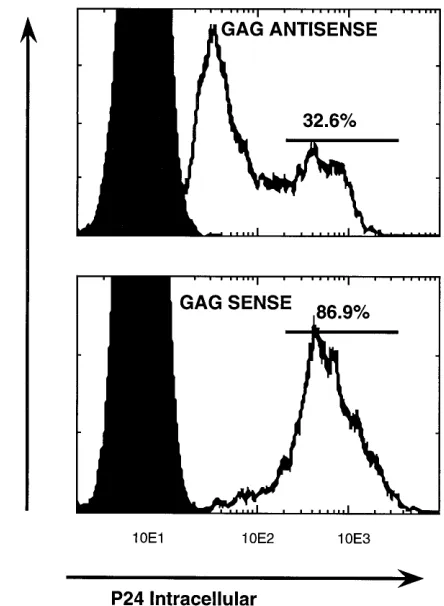
infection [MOI], 0.1). FACS analysis at day 5 demonstrated
that only 33% of the
C
-gag antisense RNA-expressing cells
produced high p24 Ag levels (mean fluorescence intensity, 594)
compared with 87% in the control cells at a higher mean
fluorescence intensity (930) (Fig. 6). The majority of antisense
RNA-expressing CEMSS cells (upper panel, left peak) express
10-fold-lower p24 Ag levels (mean fluorescence intensity, 50)
(Fig. 6), demonstrating that expression of the
C
-gag antisense
transcript resulted in decreased HIV-1 Gag protein production
in these infected cells. This difference is not explained by the
HIV-1 infection rates of these cultures, since we have shown
that the two populations can be infected equally well with
HIV-Prohygro (Table 2). In parallel, we prepared RNA from
these cultures and analyzed HIV-1 transcripts by Northern
blotting. All three HIV-1 RNA species in the total RNA of
the CEMSS cells carrying the antisense
C
-gag vector were
decreased (Fig. 7). Furthermore, quantitative analysis of the
Northern blot using a PhosphorImager (Molecular Dynamics)
showed that the ratios of the full-length (9.0-kb), singly spliced
(4.5-kb), and multiply spliced (2.5-kb) HIV-1 transcripts were
different in the HIV-1-infected sense and antisense
RNA-ex-pressing CEMSS cell populations (Fig. 8). HIV-1-infected
[image:5.612.56.471.63.540.2]C
-gag/S-transduced CEMSS cells expressed the full-length,
singly spliced, and multiply spliced transcripts at a ratio of
30:20:50, respectively. This ratio was changed to 15:20:65 in the
CEMSS cells expressing the complementary
C
-gag fragment,
resulting in a 50% reduction of the full-length HIV-1 transcript
which can anneal to the antisense RNA. This specific reduction
FIG. 4. Schematic map and expression ofC-gag deletion constructs. (A) The Cand gag fragments were generated by restriction digest. The serial 39deletion fragments (gag-1 to gag-6) and the 59deletions (gag-7 to gag-8) were generated by PCR as described in Materials and Methods. The lengths of the generated DNA fragments are listed on the right. (B) Total cellular RNA (10mg) from transduced CEMSS cell populations was analyzed on a 1.2% agarose–2.2 M formaldehyde gel. The Northern blot was hybridized with a neo- and GAPDH-specific probe. wt., wild type.
on November 9, 2019 by guest
http://jvi.asm.org/
of the full-length HIV-1 transcript also correlates with a
de-crease of the recombinant neo antisense transcript in the
HIV-1-infected cell population (Fig. 7), indicating that annealing of
the antisense transcript to the target HIV-1 RNA leads to the
specific degradation of both RNA species (1).
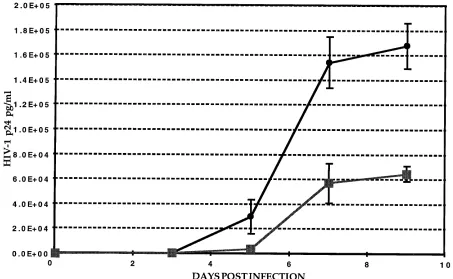
Inhibition of HIV-1 replication in human PBLs.
In addition
to efficacy studies with CEMSS cells, we also evaluated the
C
-gag antisense transcript in PBLs. Stably transduced,
acti-vated, and FACS-sorted human peripheral CD4
1and Lyt2
1T
lymphocytes expressing either the gag antisense or a control
sequence were submitted to HIV-1 challenge with the JR-CSF
clinical isolate. More than 90% of the sorted PBLs expressed
the CD4-Lyt2 surface markers. Triplicate cultures were
inoc-ulated with HIV-1, and virus replication was monitored by
measuring the culture p24 Ag levels every second day. As
shown in Fig. 9, p24 Ag production in the supernatants
in-creased over time, indicating the spread of the infection
through the culture. Although the cell population expressing
the antisense RNA produced p24 Ag, virus replication in this
culture was decreased by 87, 67, and 63.5% at days 5, 7, and 9,
respectively. These results indicate that gag antisense RNA
efficiently inhibits replication of the HIV-1 JR-CSF strain in
primary CD4
1T cells.
DISCUSSION
In this study, retroviral vectors expressing antisense RNA
against the HIV-1
C
-gag region were tested for their antiviral
efficacy in the human CEMSS T-cell line and in CD4
1PBLs.
On the basis of previous studies (8, 23), the antisense transcript
was expressed from the MLV LTR, which is a strong promoter
in the relevant human cell lineages. Previously, antiviral
activ-FIG. 5. Anti-HIV-1 efficacy of different deletion mutants. (A) HIV-1 chal-lenge (HXB3, 43102TCID
50/ml) of CEMSS cells expressing the
complemen-taryCand gag subfragment. (B) Anti-HIV-1 efficacy ofC-gag deletion con-structs. CEMSS cells transduced with the deletion constructs were infected with 43102TCID
50of HIV-1 HXB3 strain per ml.
FIG. 6. Intracellular p24 expression in HIV-1-infected CEMSS cells. CEMSS cells (106
/ml) expressing theC-gag fragment in the sense or antisense orientation were inoculated with 105
TCID50of HIV-1 HXB3 strain per ml (MOI, 0.1).
[image:6.612.332.554.70.373.2]Intracellular p24 expression was analyzed by FACS at day 5 postinfection. The negative-control results (noninfected cells) (solid curves), results for HIV-1-infected cells (open curves), and the gate settings (bars) are indicated. The values shown on the x axis are the mean fluorescence intensity for the p24-specific signal.
TABLE 2. Comparison of infection of CEMSS cells expressing
C
-Gag sense and antisense transcripts with
control HIV-1 infection
aCell population
No selection Selection
No. of colonies
Plating efficiency (%) SE
No. of
colonies SE % Hyg
r
C
-gag/AS
58.2
11.6
4.7
22.4
2.6
3.8
C
-gag/S
48.8
9.76
3.4
17.0
1.1
3.4
aCEMSS cells were infected with HIV-1 virus carrying the hygromycin resis-tance gene. After HIV-1 infection, 500 and 5,000 cells were seeded in 1 ml of MC supplemented with RPMI 1640 and 10% fetal calf serum in duplicate. Half of the cultures were selected on hygromycin. MC colonies on the dishes seeded with 500 cells were scored after 6 days to determine seeding efficiency, and colonies on the dishes with 5,000 cells were scored after 12 days to determine hygromycin resistance. The control, untransduced CEMSS cells have;10% seeding effi-ciency without selection, and no colonies were observed with hygromycin selec-tion.
V
OL. 70, 1996
INHIBITION OF HIV-1 REPLICATION WITH ANTISENSE RNA
8797
on November 9, 2019 by guest
http://jvi.asm.org/
ity of the 1,425-nt
C
-gag antisense sequence expressed from an
internal herpes simplex virus thymidine kinase promoter in
MT4 cells was reported (7). Our experiments support the
de-scribed anti-HIV efficacy of the antisense
C
-gag sequence, and
in addition to the previous report, our data for CEMSS cells
demonstrate antisense RNA-mediated HIV-1 inhibition over a
wide range of inoculation doses (4
3
10
2to 1
3
10
5TCID
50
/
ml). Suppression of HIV-1 replication correlated with
mainte-nance of CD4
1surface marker expression and cell viability.
These differences between the sense and antisense
RNA-ex-pressing CEMSS populations were not caused by differences in
HIV infectivity, as measured by a single-cell infection assay
system (Table 2).
In addition to cell lines, we also demonstrated HIV-1
sup-pression in PBLs transduced with retroviral vectors expressing
gag antisense transcripts. We have used the truncated mouse
CD8 molecule (Lyt2) as a selectable cell surface marker to
rapidly isolate transduced PBLs without any drug selection.
Over 90% of the selected PBLs expressed the antisense
tran-scripts, and we observed 70% reduction in HIV-1 replication.
Although suppression of HIV-1 replication was lower than in
the CEMSS cell line, this difference may be explained by the
residual nontransduced cell fraction (10%) and the different
replication kinetics of the HIV-1 in PBLs.
In a previous report (7), it was suggested that sequences
complementary to
C
were responsible for the observed
anti-HIV effect. Our study does not support this hypothesis.
Dele-tion analyses of the 1,425-nt
C
-gag fragment clearly
demon-strated that the packaging sequence (
C
) alone has no antiviral
effect. The 952-nt anti-gag sequence (Fig. 4B) which had the
C
sequence specifically deleted was as efficient in HIV
suppres-sion as the 1,425-nt
C
-gag antisense sequence originally
de-scribed (Fig. 5A). However, our deletion analysis suggests that
the length of the antisense transcript is important and inversely
correlated with the observed anti-HIV efficacy even at a low
(4
3
10
2TCID
50
/ml) viral load. There appear to be no specific
sequence requirements, as indicated by the gag-3 (625-nt) and
gag-7 (650-nt) deletion mutants. The levels of inhibition of
HIV-1 replication in CEMSS cells transduced with these
vec-tors were comparable (Fig. 5B), although the gag-2 constructs
lack any overlapping sequences (Fig. 4A), suggesting that the
ability to anneal to the target sequence may be the rate-limiting
step (35). Furthermore, our data suggest that increasing the
length of the antisense transcript has no negative impact on
retroviral stability, titer, and steady-state expression levels. In
addition to improved suppression of HIV-1 replication, longer
transcripts have the theoretical advantage that they can anneal
to a variety of HIV quasispecies including multiple point
mu-FIG. 7. Northern blot analyses of CEMSS cells inoculated with 105TCID 50
[image:7.612.113.239.67.259.2]of HIV-1 HXB3 strain per ml (MOI, 0.1). Total cellular RNA was isolated from CEMSS cells at day 5 postinfection, when the intracellular p24 antigen was determined. The Northern blot was probed with TAR-, neo-, and GAPDH-specific oligonucleotides. Lane 1, uninfected CEMSS wild-type cells; lane 2, uninfected CEMSS pool expressing theC-gag antisense fragment; lane 3, HIV-1-infected CEMSS pool expressing theC-gag antisense fragment; lane 4, HIV-1-infected CEMSS pool expressing theC-gag sense fragment.
FIG. 8. Quantitative analyses of the steady-state RNA expression levels in CEMSS cells expressing theC-gag sequence in the sense or antisense orientation. Levels of HIV-1-specific full-length (9.0-kb), singly spliced (4.5-kb), and multiply spliced (2.5-kb) RNAs in HIV-1-infected CEMSS cells were quantitated (Fig. 7, lanes 3 and 4) by using a Molecular Dynamics PhosphorImager. Results obtained from CEMSS cells expressing theC-gag sense (upper curve) and antisense (lower curve) RNAs are indicated.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:7.612.142.490.489.693.2]tations throughout the target transcript. As a consequence, the
occurrence of HIV-1 escape mutants should be less likely than
with other RNA-based strategies including short antisense
transcripts (16, 24), oligonucleotides (1), and ribozymes (21,
37).
We demonstrated anti-HIV efficacy of the antisense strategy
in transduced CEMSS cells at a high MOI (0.1; 10
5TCID
50
of
HIV-1 per ml). At such high inoculation doses, the majority of
the antisense RNA-expressing cells (67.4%) express detectable
levels (fivefold above background) of intracellular p24 Ag (Fig.
6), and it will be interesting to see whether this will result in
immunogenicity in patients. This experiment allowed us to
analyze the steady-state levels of the HIV-1 and recombinant
antisense transcripts. Our data indicate a specific decrease of
the full-length HIV-1 target RNA and, concomitantly, of the
recombinant antisense transcripts. This result implies
inhibi-tion of HIV-1 gene expression through RNA-RNA duplex
formation and subsequent degradation of the double-stranded
transcripts. The result of the HIV-Prohygro experiment
dem-onstrates that the
C
-gag antisense RNA cannot prevent
inte-gration of the incoming provirus, suggesting an exclusively
posttranscriptional mode of action. In contrast to ribozymes,
this antisense RNA-based strategy requires at least a 1:1 ratio
of antisense and HIV-1 target RNAs, but our experiments
demonstrate the feasibility of this approach. With the
retrovi-ral vectors described in this study, sufficiently high steady-state
levels of expression of the antisense transcripts can be readily
achieved in human T-cell lines and PBLs. Compared with
protein-based gene therapy strategies, RNA-based approaches
are less likely to induce immune responses to the genetically
modified cells. Our data provide the scientific rationale for the
clinical evaluation of the described strategy in transduced
pe-ripheral blood mononuclear cells or hematopoietic stem cells.
ACKNOWLEDGMENTS
The Progenesys program at Systemix is a research and development
collaboration jointly sponsored by Sandoz and Systemix Inc.
We thank M. Bonyhadi and U. Junker for constructive comments on
the manuscript. The following reagents were obtained through the
AIDS Research and Reference Reagent Program, Division of AIDS,
NIAID, NIH: CEMSS cells (catalog no. 776) from P. Nara and
HIV-1SF2
(ARV2) (catalog no. 275) from J. Levy.
REFERENCES
1. Anadozo, M. A., M. A. Wainberg, A. D. Friesen, and J. A. Wright. 1995. Sequence-specific inhibition of gene expression by a novel antisense oligode-oxynucleotide phosphorothiotate directed against a nonregulatory region of the human immunodeficiency virus type 1 genome. J. Virol. 69:1794–1801. 2. Bahner, I., C. Zhou, X. J. Yu, J. C. Guatelli, and D. B. Kohn. 1993.
Com-parison of trans-dominant inhibitory mutant human immunodeficiency virus type 1 genes expressed by retroviral vectors in human T lymphocytes. J. Vi-rol. 67:3199–3207.
3. Baltimore, D. 1988. Intracellular immunization. Nature (London) 335:395– 396.
4. Bevec, D., M. Dobrovnik, J. Hauber, and E. Bo¨hnlein.1992. Inhibition of human immunodeficiency virus type 1 replication in human T cells by ret-roviral-mediated gene transfer of a dominant-negative rev trans-activator. Proc. Natl. Acad. Sci. USA 89:9870–9874.
5. Bevec, D., B. Volc-Platzer, K. Zimmermann, M. Dobrovnik, J. Hauber, G. Veres, and E. Bo¨hnlein.1994. Constitutive expression of chimeric Neo-Rev response element transcripts supresses HIV-1 replication in human CD41T lymphocytes. Hum. Gene Ther. 5:193–201.
6. Cann, A. J. 1990. Stable and safe HIV provirus clones. Nucleic Acids Res. 18:6153–6154.
7. Coli, H., B. Fan, R. L. Joshi, A. Ramezani, X. Li, and S. Joshi. 1994. Inhibition of HIV-1 multiplication in a human CD41lymphocytic cell line expressing antisense and sense RNA molecules containing HIV-1 packaging signal and rev response element(s). Antisense Res. Dev. 4:19–29. 8. Escaich, S., C. Kalfoglou, I. Plavec, S. Kaushal, J. D. Mosca, and E. Bo
¨hn-lein.1995. RevM10-mediated inhibition of HIV-1 replication in chronically infected T cells. Hum. Gene Ther. 6:625–634.
[image:8.612.80.533.70.349.2]9. Gilboa, E., and C. Smith. 1994. Gene therapy for infectious diseases: the AIDS model. Trends Genet. 10:139–144.
FIG. 9. Inhibition of HIV-1 replication in transduced PBLs. Transduced and FACS-analyzed PBLs (CD41, Lyt21) were infected with the clinical HIV-1 isolate JR-CSF. Cells (53104) were inoculated in triplicate. PBLs were transduced with the pLLyt2-gag/AS (■) and pLLyt2-gag/S (
F) vectors.
V
OL. 70, 1996
INHIBITION OF HIV-1 REPLICATION WITH ANTISENSE RNA
8799
on November 9, 2019 by guest
http://jvi.asm.org/
10. Jacks, T., M. D. Power, F. R. Masiarz, P. A. Luciw, P. J. Barr, and H. E. Varmus.1988. Characterisation of ribosomal frameshifting in HIV gag/pol expression. Nature (London) 331:280–283.
11. Joshi, S., A. van Brunschot, S. Asad, I. van der Elst, S. E. Read, and A. Bernstein.1991. Inhibition of human immunodeficiency virus type 1 multi-plication by antisense and sense RNA expression. J. Virol. 65:5524–5530. 12. Kotani, H., P. B. Newton, S. Zhang, Y. L. Chiang, E. Otto, L. Weaver, R. M.
Blaese, F. W. Anderson, and G. J. McGarrity.1994. Improved methods of retroviral vector transduction and production for gene therapy. Hum. Gene Ther. 5:19–28.
13. Lee, T., H. F. Gallardo, E. Gilboa, and C. Smith. 1994. Inhibition of human immunodeficiency virus type 1 in human T cells by a potent Rev-response element decoy consisting of the 13-nucleotide minimal Rev-binding domain. J. Virol. 68:8254–8264.
14. Malim, M. H., S. Bo¨hnlein, J. Hauber, and B. R. Cullen.1989. Functional dissection of the HIV-1 Rev trans-activator derivation of a trans-dominant repressor of Rev function. Cell 58:205–214.
15. Malim, M. H., W. W. Freimuth, J. Liu, T. J. Boyle, H. K. Lyerly, B. R. Cullen, and G. J. Nabel.1992. Stable expression of transdominant rev protein in human T cells inhibits human immunodeficiency virus replication. J. Exp. Med. 176:1197–1201.
16. Meyer, J., S. Nick, T. Stamminger, F. Grummt, G. Jahn, and H. J. Lipps. 1993. Inhibition of HIV-1 replication by high-copy-number vector expressing antisense RNA for reverse transcriptase. Gene 129:263–268.
17. Miller, A. D., and C. Buttimore. 1986. Redesign of retrovirus packaging cell lines to avoid recombination leading to helper virus production. Mol. Cell. Biol. 6:2895–2902.
18. Miller, D. A., and G. J. Rosman. 1989. Improved retroviral vectors for gene transfer and expression. BioTechniques 7:980–988.
19. Mizumo, T., M. I. Chou, and M. Inouye. 1984. A unique mechanism regu-lating gene expression: translational inhibition by a complementary RNA transcript. Proc. Natl. Acad. Sci. USA 81:4123–4130.
20. Nabel, G. J., B. A. Fox, L. Post, C. B. Thompson, and C. Woffendin. 1995. A molecular genetic intervention for AIDS—effects of a transdominant nega-tive form of Rev. Hum. Gene Ther. 5:79–92.
21. Ojwang, J. O., A. Hampel, D. J. Lonney, F. Wong-Staal, and J. Rappaport. 1992. Inhibition of human immunodeficiency virus type 1 expression by a hairpin ribozyme. Proc. Natl. Acad. Sci. USA 89:10802–10806.
22. Pear, W., G. P. Nolan, M. Scott, and D. Baltimore. 1993. Production of high-titer helper-free retroviruses by transient transfection. Proc. Natl. Acad. Sci. USA 90:8392–8396.
23. Plavec, I., M. Agarwal, K. E. Ho, M. Pineda, S. Forestell, J. Auten, H. Matsuzaki, S. Escaich, and E. Bo¨hnlein.Optimization of retroviral vectors encoding the trans-dominant RevM10 protein for gene therapy of AIDS. Submitted for publication.
24. Rhodes, A., and W. James. 1990. Inhibition of human immunodeficiency virus replication in cell culture by endogenously synthesized antisense RNA. J. Gen. Virol. 71:1965–1974.
25. Riddel, S. R., M. Elliott, D. A. Lewinsohn, M. J. Gilbert, L. Wilson, S. A. Manley, S. D. Lupton, R. W. Overell, T. C. Reynolds, L. Corey, and P. D. Greenberg.1996. T-cell mediated rejection of gene-modified HIV-specific cytotoxic T lymphocytes in HIV-infected patients. Nature Med. 2:216–223. 26. Rigg, R. J., J. S. Dando, S. Escaich, I. Plavec, and E. Bo¨hnlein.1995. Detection of intracellular HIV-1 Rev protein by flow cytometry. J. Immunol. Methods 188:187–195.
27. Sczakiel, G., M. Homann, and K. Rittner. 1993. Computer-aided search for effective antisense RNA target sequences of the human immunodeficiency virus type 1. Antisense Res. Dev. 3:45–52.
28. Sczakiel, G., M. Oppenlander, K. Rittner, and M. Pawlita. 1992. Tat- and Rev-directed antisense RNA expression inhibits and abolishes replication of human immunodeficiency virus type 1: a temporal analysis. J. Virol. 66:5576– 5581.
29. Sczakiel, G., M. Pawlita, and A. Kleinheinz. 1990. Specific inhibition of human immunodeficiency virus type 1 replication by RNA transcribed in sense and antisense orientation from the 59-leader/gag region. Biochem. Biophys. Res. Commun. 169:643–651.
30. Sczakiel, G., and M. Pawlita. 1991. Inhibition of human immunodeficiency virus type 1 replication in human T cells stably expressing antisense RNA. J. Virol. 65:468–472.
31. Stout, T., and C. T. Caskey. 1987. Antisense RNA inhibition of endogenous genes. Methods Enzymol. 151:519–530.
32. Sullenger, B. A., H. F. Gallardo, G. E. Ungers, and E. Gilboa. 1990. Over-expression of TAR sequences renders cells resistant to human immunode-ficiency virus replication. Cell 63:601–608.
33. Tagawa, M., H. Nakauchi, L. A. Herzenberg, and G. P. Nolan. 1986. Formal proof that different-size Lyt2 polypeptides arise from differential splicing and post-transcriptional regulation. Proc. Natl. Acad. Sci. USA 83:3422–3426. 34. Trono, D., M. B. Feinberg, and D. Baltimore. 1989. HIV-1 gag mutants can
dominantly interfere with the replication of the wild-type virus. Cell 59:113– 120.
35. Tung, F. Y. T. 1994. Suppression of simian immunodeficiency virus replica-tion in primary peripheral mononuclear cells by antisense RNA. J. Med. Virol. 42:255–258.
36. Woffendin, C., U. Ranga, Z.-Y. Yang, L. Xu, and G. J. Nabel. 1996. Expres-sion of a protective gene prolongs survival of T cells in human immunode-ficiency virus infected patients. Proc. Natl. Acad. Sci. USA 93:2889–2894. 37. Zhou, C., I. Bahner, G. P. Larson, J. A. Zaia, J. J. Rossi, and D. B. Kohn.
1994. Inhibition of HIV-1 in human T lymphocytes by retrovirally transduced anti-tat and rev hammerhead ribozymes. Gene 149:33–39.