0022-538X/93/084497-07$02.00/0
Copyright © 1993, AmericanSociety forMicrobiology
Characterization of
a
Temperature-Sensitive
Mutant
of
the
UL15 Open Reading
Frame
of
Herpes
Simplex Virus
1
ALICE P. W. POON AND BERNARDROIZMAN*
The
Marjorie
B. Kovler ViralOncologyLaboratories, UniversityofChicago, 910 East58thStreet,
Chicago,
Illinois 60637Received11March1993/Accepted 23April 1993
The UL15 gene of herpes simplex virus 1 consists of two exons and is
highly
conserved among the herpesviruses sequencedtodate. Other than itshomologyto aphage protein involved in the packaging ofDNA, nothingis knownof itsfunction. This reportconcerns theisolationofatemperature-sensitive mutant witha mutationmapping in the UL15 open reading frame. Cells infected with the parent, mutant, and rescued viruses all make DNAatthenonpermissivetemperature. Directanalyses
oftheDNA andelectronmicroscopicstudies indicate thatalthough viral DNAismade, it isnotpackaged into capsids present in nuclei. These studiessuggest thatUL15 may be involvedinthepackaging of viral DNA.In this article,we report the isolation and phenotype in infected cells ofa temperature-sensitive (ts)mutant of the herpessimplex virus1(HSV-1)geneUL15. Relevanttothis report arethefollowing.
(i) Most isolates of HSV-1 passaged alimited number of times in cellsinculture exhibit a ts phenotype. This is the case for HSV-1 strain F [HSV-1(F)] passaged a limited number of times in cells in culture. Todeveloptsmutants,it is first necessary to select viruses which do multiply at
highertemperaturesto serve asgenetic wildtypes. Forthis purpose,we have in the past usedastrain of virus which is passaged serially in cell culture many times and which multiplies well at 39°C, the nonpermissive temperature. In thecourseofplaquepurificationsofvirusesfollowing
trans-fection of cells with intact DNA and fragments of a gene mutagenized at a specific site, we isolated numerous ts mutants notrelevanttothemutagenizedgene. One of these tsmutations turnedout tomapspecificallyin theUL15open readingframe. Thephenotypeofthismutant is described in this report.
(ii) The UL15 open reading frame is of interest from severalpointsof view.First,it isoneof the very few genes knowntoyield splicedRNAs(20).Itisparticularly
notewor-thy that exons 1 and 2 are separated bytwo open reading frames, UL16 andUL17, arranged antisenseto UL15. UL16 is not essential for viral replication in cells in culture (1). Attempts in thislaboratorytodeleteUL15orUL17were not successful (2a). However, a recentreportfrom this labora-toryshowed that acDNA copy of theUL15geneyielding a
single, unspliced RNA can replace UL15 exons 1 and 2 withoutaffectingthecapacityof the virustoreplicatein cells inculture (2).
(iii)Arecentpublication byDavison(6)drew attentionto the facts that the UL15 open reading frame is highly
con-served amongherpesviruses and that the separation of the two exons by antisense open reading frames is also con-served. Ahomologof theUL15openreadingframeexists in thechannel catfishherpesvirus.Davisonpointedoutthat the UL15 protein maybe related to protein 17 of the T4phage (6).Thisproteinis knowntobeinvolved inpackagingDNA (15, 16).
*Correspondingauthor.
We reportthat thets mutant atthenonpermissive temper-aturesynthesizes capsids and DNA butpackagingof DNA does nottakeplace.
MATERIALSANDMETHODS
Virus and cells.HSV-l(mP)wasthe namegivento astrain initially designated strain NT and isolated from a human infection in embryonated eggs(10).From the NTstrain,two viruses were isolated in cell culture. One, designated the microplaque strain [HSV-l(mP)], causes infected cells to aggregate in a fashion typical of most HSV-1 strains. The other, designated the macroplaque strain [HSV-1(MP)],
causescellstofuse intopolykaryocytes (7).Acharacteristic of HSV-l(mP) is that it grows as well at 39°C as at 34°C. Unlike the HSV-1(F) strain used in this laboratory as the prototype HSV-1 strain and passaged a limited number of times in cells in culture, HSV-l(mP) has been passaged at least 80 times in FL and HEp-2 cell cultures.
The sources and procedures for the propagation of Vero and rabbit skin cells have been described elsewhere (2, 14). Plasmid libraries. TheHSV-1(F) plasmid libraxy was de-scribed elsewhere (14). TheHindIllJfragment was cloned aspRB208 in pACYC184 (13a) and was derived from that plasmid. The derivation of the fragments shown in Fig. 1 was asfollows. DNA fragment EcoRI-2 was obtained by diges-tion ofHindIII-J. BamHI-BglII 4, SalI 6, andMluI 1 DNA fragments were obtained by digestion of EcoRI-2 with the appropriateenzymes. TheXbaIfragments of HSV-1 DNA (4)werethe kind gift of Mark Challberg.
Markerrescue. Rabbit skincells grown in 25-cm2 flasks were transfected with 0.6 to 0.8 ,ug of HSV-l(mP)ts66.4 DNAwith theappropriatefragment ofHSV-1(F)DNA. The cellsweregrownat37°C and were harvested when almost all cells exhibited cytopathic effects, usually 7to 8days post-transfection. Titers ofthe virusesobtained fromeach trans-fectionweredeterminedat 34 and39°C.
Efficiencyofplating at34 and39°C. Replicate cultures of Vero cells in
25-cm2
flasks were exposed in duplicate to various dilutions of thewild-type, mutant, orrescued virus andincubatedateither 34or39°C. After 3days,thecultures were stained and plaqueswere counted as previously de-scribed(7).Efficiencyofreplicationat34 and39°C.Replicatecultures 4497
on November 9, 2019 by guest
http://jvi.asm.org/
ofVero cells in 25-cm2 flaskswereexposed in duplicate to
1.0or0.1PFU ofwild-type,mutant,orrescuedviruspercell
andincubatedateither 34or39°C. At 18 hpostinfection, the
cells were harvested and the titers of the viruses were
determinedat 34°C inVero cells.
Extraction and analysis of HSV-1 DNA. In all of these studies, Vero cells grown in replicate 25-cm2 flasks were
exposedto 10 PFU ofwild-type, mutant, or rescued virus
percell andincubatedat34or39°C.Atspecifiedtimes, the
cellswereharvested, and DNAwasextracted andanalyzed,
asdescribedinResults.
Labeling andelectrophoretic separation of infected-cell
pro-teins. Replicate cultures ofVero cells in 25-cm2flaskswere
infected in duplicate with wild-type, mutant, or rescued
virus and incubated at either 34 or 39°C. The cells were
labeled byincubation with 1 ml ofmethionine-free medium supplemented with 50 ,Ci of (35S]methionine (1,000 Ci/ mmol; Amersham, Arlington Heights, Ill.) for 1 h at 13 h postinfection and then harvested, lysed, and electrophoret-ically separated on11% polyacrylamide gels and subjected toautoradiographyaspreviouslydescribed (12, 18).
RESULTS
Isolation of themutantHSV-1(mP)ts66.4.Theexperiments which led to the isolation ofthe mutant were as follows. Rabbit skin cell monolayers in 25-cm2 flasks were trans-fected with intact HSV-l(mP) DNA and DNA fragments containingtheopenreadingframeUL48,whichencodesthe protein designated the a trans-inducing factor (aTIF) by
Pellett etal. (13) orvirion protein 16 (VP16) by Spear and
Roizman (18). These fragments were modified by in situ
mutagenesis in order to replace selected cys codons. The
progeny of the transfectionwasplated atapproximately50 PFUperflask.Plaqueswereselected, purified,andplatedin replicate24-welldishesat34 and39°C.Isolates which failed to cause cytopathic effects at 39°C were selected, plaque
purifiedagain,andretested. The isolates whichreproducibly failedtomultiplyat39°Cwerestudied further.
Althoughalargenumber oftsmutantswereisolated,none
of the mutations mapped in the target gene. Some of the mutations mapped in genes whose phenotypes were well characterized in other studies. The mutation in themutant whichweselectedfor further studies and designated
HSV-1(mP)ts66.4 mapped in a gene whose function was not known.
Mapping of the mutation in HSV-1(mP)ts66.4. In these series ofexperiments,rabbit skin cellsweretransfectedwith intact mutant DNA and wild-type HSV-1 fragments. The
progenyoftransfectionwas plated at 34 and39°C. Several series ofexperimentsweredonetonarrowthe exactlocation of themutationresponsibleforthetsphenotype. Inthe first series, we tested individual fragments from an XbaI
frag-ment library, kindly donated by Mark Challberg, which
encompassthe entiregenome. Thefragmentwhich rescued the mutationwasXbaI-C(Fig. 1, line2;Table 1).Thenext series of transfections were done with DNA fragments
derived from HSV-1(F) DNAwith the region spanned by XbaI-C. Ofthe fragments tested,theoneswhichrescued the
mutationareshown inTable 1 and Fig. 1. Thetwosmallest fragments which individually rescued the mutation span
exon 2 ofUL15 (coding for the carboxyl terminus of the protein) and the UL18 openreading frame, and a fragment
whichencodes allofUL15alsorescued the mutation(Table 1). Several plaques produced at 39°C were harvested, and
thevirusesobtainedwereplaque purifiedand studiedfurther
a b
1
0
ULb-._
.I I..
3 A1,E1/l 2 A1
4
./
E SBgS ,,S~B Bg S
~
99
,, , pg
5 ,'|'Sol
----6.,'BBg %
SMS
Ia' e U c a
B SH
7 F
iM
8 UL
18
i
8-
L ULl9FIG. 1. HSV-1 DNA sequence arrangement and location of DNAfragmentswhich rescuedHSV-1(mP)ts66.4.Line 1, sequence arrangement of HSV-1 DNA (rectangles represent the inverted repeatsab, b'a', a'c', and ca, which flank the long and shortunique sequences,UL andUs); line 2, XbaI Cfragment; line 3, HindIII J fragment; line 4, EcoRI 2 fragment; line 5,BamHI-BgilI4fragment; line 6,SalI 6 fragment; line 7, MluI1fragment; line 8, map positions of the UL15, UL18, and UL19 genes relativetotheSall 6 fragment. The positions ofthe UL15, UL18, andUL19 transcriptional units were drawnaccordingtothestudy ofCosta etal.(5). B, BamHI; Bg, BglII; E, EcoRI; H, HindIII; M,MOuI;S,Sail;X,XbaI.
as representatives of rescued viruses. The two viruses studied extensivelywere designated
HSV-l(mP)Ra,
whichwas obtained by rescue with the Sall 6 fragment, and HSV-l(mP)Rb, which was obtained by rescue with the BamHI-BglII4fragment.
Plating andreplication efficiencies of thewild-type,mutant, and rescued viruses at the nonpermissive temperature. The results of the studies done with wild-type, mutant, and rescuedvirusesaresummarizedinTable 2. All of the assays weredonewithVerocells incubated ina34°C incubatoror submergedin a39°Cwaterbath. The resultsare
internally
[image:2.612.323.539.69.278.2]consistent: they show that the parent virus
replicates
and plates better at 39 than at 34°C, that the mutant virusTABLE 1. Resultsof markerrescueofHSV-1(mP)ts66.4
Fragmentor Virus titer(PFUI/mil) Ratio(39/
plasmidused 340C 390C 34°C)
None 3.5 x 107 <102 <2.9 x 10-6
XbaIC' 3.2 x 108 1.9 x 107 5.9x 10-2
3.4 x 10' 7.9 x 107 2.3 x 10-1 HindIII J 1.5 x 108 5.6 x 107 3.7 x 10-1
EcoRI 2 3.0 x 107 1.4x 106 4.7 x 10-2
SalI 6 7.7 x 107 3.6 x 107 4.7 x 10-1
BamHI-BglII4 2.5 x 107 7.4 x 106 3.0x 10-1
MluIla 2.4 x 108 1.8x 107 7.5 x 10-2
9.5 x 107 3.0 x 107 3.2 x 10-1
pRB4121b 4.7x 107 2.4 x 107 5.1 x 10-1
aDataforXbaI-CandMluI-1represent repeat
experiments.
bThisplasmidcontainsanintactcDNA copyof theUL15gene(2).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.312.554.578.699.2]TABLE 2. Plating efficiency and yield of parent, ts mutant, and rescued viruses at 34 and 39°C
Virus Platingefficiency(39/34°C)
Yield'
at1PFU/cell Ratio(39/Yield'
at0.1PFU/cell Ratio (39/Expt1 Expt2 34°C 390C 34'C) 340C 39°C )
HSV-1(mP) 0.97 0.98 3.7x 106 3.1 x 107 8.4 1.8 x 105 6.4 x 106 35.6
HSV- <6.7 x 10-7 <8.7 x 10-7 6.3 x 105 1.1 x 103 1.7 x 10-3 5.8 x 104 1.0 x 102 1.7 x 10-3 1(mP)ts66.4
HSV-1(mP)Ra 1.1 0.7 4.9x 106 2.2 x
107
4.5 2.7 x 105 4.3 x 106 15.9HSV-l(mP)Rb 0.77 0.81 6.0 x 106 2.1 x 107 3.5 3.9 x 105 5.6 x 106 14.4
aYield of virus at 18 h postinfection.
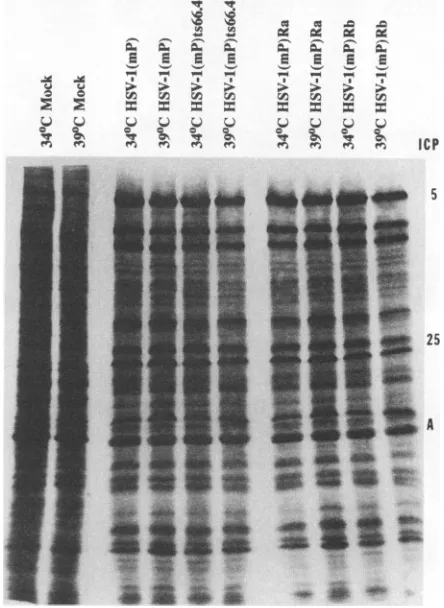
replicatesandplateswithmuch lowerefficiencyat 39 than at 34°C,and that therescued virusesresembletheparent virus. The synthesis of proteins by the parent, mutant, and rescued viruses at permissive and nonpermissive tempera-tures. Inthis series ofexperiments, replicate
25-cm2
cultures of Vero cells were infected in duplicate with the parent, mutant, orrescuedvirus(5 PFUpercell) and incubatedin a 34°C incubatororsubmerged ina39°C
waterbath.At 13h postinfection, the cells were labeled for 1 h with [315]me-thionine and then harvested, solubilized, and subjected to electrophoresisinpolyacrylamide gels.Theresultsshown in Fig. 2 indicate that electrophoretic profiles oflabeled pro-teinsfrom cells infected and maintained at the nonpermis-sive temperature could not be differentiated from those extracted fromcells infectedandmaintained atthe permis-sivetemperature.Electronmicroscopic studies. Thepurposeof this seriesof experiments wasto attempt todefine the phenotype of the virus in cells infected andmaintained atthe nonpermissive temperature.ReplicateVerocell cultureswereinfected with the parent, mutant, or
repaired
virus(5
PFUpercell)
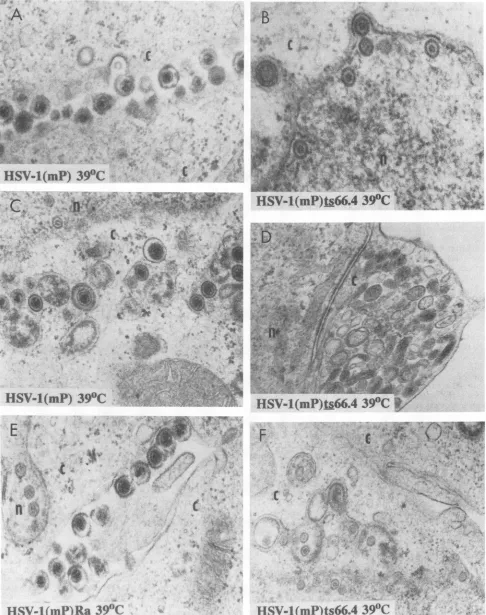
and maintained at39°C. The cellswerefixedat18 hpostinfection and thensectioned, stained,and examined in aSiemens102 electronmicroscope.Asshown inFig. 3,cellsinfected with HSV-l(mP) and maintained at 39°C contained numerous enveloped capsids containing DNA in the space between cells andin thecytoplasmicvesicles(Fig.3AandC, respec-tively). In contrast, the nuclei ofcells infected withHSV-1(mP)ts66.4
contained numerous capsids with scaffolding proteins(whichform athinringinside thecapsid)but lacking DNA. These capsids were scattered in the nucleus, and somewere inapposition tothe nuclear membrane(Fig. 3B and F). Emptycapsids within envelopes andvesicleswere alsoseenoccasionally (Fig.3B)butinfrequently.Astriking featureof cells infected with theHSV-l(mP)ts66.4virus was thepresenceofnumerouscytoplasmicvesicleslackingvirus particles (Fig. 3D). Some of these vesicles resembled frag-mented Golgirecently
reported to be present in infected Verocells (3). Electron micrographs of cells infected with HSV-l(mP)Raandmaintainedat39°Ccould not be differen-tiated fromthose ofcellsinfected with theparent virus(Fig. 3A and E). Similar results were obtained withHSV-1(mP)Rb-infected
cells. Weshould notethat at the permis-sive temperature(34°C),
the mutant virus could not be differentiated fromwild-type virus with respect to the pres-ence in infected cells of virionscontaining DNA (data not shown).Synthesisandpackaging ofviralDNA.The electron micro-scopic studies indicated that in cells infected with
HSV-1(mP)ts66.4
andmaintained atthe nonpermissive tempera-ture capsids containing the scaffolding proteinswere made andthatsomeof the emptycapsidswereenvelopedbut that noneof thecapsids contained DNA.To determine whetherthe defect at the nonpermissivetemperature was due to the absenceof viral DNA or a failure to package the DNA,three seriesofexperimentsweredone.
Theobjective of thefirsttwoseries of experimentswas to determinewhether cells infected with theHSV-l(mP)ts66.4 andmaintainedatthenonpermissivetemperaturesynthesize viral DNA. In the first series (Fig. 4), replicate Vero cell cultures were exposed to
10.
PFU of parent, mutant, or rescued virus per cell and maintained at 34 or 39°C. The cells were harvested at 15 h postinfection. Total DNA was'A rA
X
s~~~ >>E;>i >
X V
IcP 5
25
A
FIG. 2. Autoradiographicimagesof[35S]methionine-labeled pro-teins from replicate cultures of Vero cells infected with parent, mutant, and rescued viruses and from mock-infected cells main-tained at 34or 39°C. The cellswerelabeledfor 1 h at 13 hafter infection, harvested, solubilized, electrophoretically separated on an11% denaturing polyacrylamide gel,andsubjectedto autoradiog-raphy.Tohelp identifytheproteins, themajor capsid protein VP5 and the tegumentproteinVP16wereidentifiedbytheirinfected-cell protein (ICP) designations (5 and25,respectively)asdescribedby Honess and Roizman (11)andbyMorse etal. (12). A, actin.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.338.560.324.628.2].-A
,4
A.
.. ...
.,
c
.-;.f' ...
~
.-I' f.*i..
.tE
?Z
UQIL (mlf tOOC'l
.
-.
f.
..
*v s
)Gt~44~-t>HSV-l(mP)tL66.439WC
tA.~~
e
~~~~~..'r.
--
'
st
E*4
* c ~ ,;47
n.
C
HSV-lOnP)Ra
39WC
HSVal(mP)1s66A4
390C
.-~-:,.~ ~ ~ ~ ~ ~ ~ ~ ~
f~~*XitR
:41 ~
tA
HSI(mP-ts66.4 39(C;*
*~.4r}.^ ~ .t
s;,:..j _ . .~~~~~~~~~~~~ ..
HSV-1
(mP)ts66A4 39W>
S
FIG. 3. Electronmicrographs ofreplicate cultures of Vero cells infected withHSV-1(mP), HSV-1(mP)ts66.4, and HSV-1(mP)Ra and
maintainedat39°C. The cellswereharvested at18 hpostinfection.The diameter of the HSVcapsidis 105 nm.c,cytoplasm;n,nucleus.
I
'4
-1.
#i?
S5o
:I
I
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.60.546.75.690.2]x
E E E E
a
E
EE
C 0 0 C 05C CC
all le CP ePS t
CS
S Cx [image:5.612.118.259.74.356.2]*e e
FIG. 4. Autoradiographic images of 32P-labeled BamHI
frag-ments F, T, and X hybridized to electrophoretically separated BamHIdigestsof total DNAs extracted fromreplicate culturesof Vero cells infected with 1(mP), 1(mP)ts66.4, HSV-1(mP)Ra and HSV-1(mP)Rb and maintained at 34 or 39°C. The
infectedreplicatecell cultureswereharvestedat15 h afterinfection.
0%
U
y-X~~~
v . . c cE6
Ca- aL
a6 aw E6
4i:
a4
X,X,
Aextracted, digestedwithBamHI, subjectedto electrophore-sis in an agarose gel, transferred to a Zeta-probe blotting membrane,and probed with 32P-labeledBamHI F,BamHI T, and BamHI X fragments. Inthe second series of experi-ments(Fig. 5), replicate Vero cell cultures were exposed to 10 PFU of parent, mutant, or rescued virus per cell and maintained at 34 or 39°C. The cells were harvested at 2 or 15 h postinfection andsolubilized innonionic detergents, and the nuclei were separated fromthe cytoplasm by centrifuga-tion. The DNA extracted from each compartment was digested with BamHI, subjected to electrophoresis in an agarosegel,transferred to aZeta-probe blottingmembrane, andprobed with32P-labeled BamHI F and BamHI X frag-ments. The DNA detected at 2 h postinfection represents inputviralDNA,sincesignificantviral DNA synthesis does not begin until after 3 h postinfection (17). The results of both series ofexperiments indicate that cells infected with HSV-l(mP)ts66.4 produced significant amounts of viral DNA and that someof the mutant virus DNA wasdetected in the cytoplasm of cells maintained at the nonpermissive temperature. Moreover, analyses ofthe results of several experiments (not shown) suggest that the differences in amounts producedby parent, mutant, and rescued viruses seeninFig.4and 5representexperimental variabilityand do notreflect the mutantphenotypeofHSV-l(mP)ts66.4.
Theimpetusfor the third seriesofexperiments emerged from the results oftwo series of studies described above. Specifically, the electron microscopic studies described aboveand whose resultsareshowninFig.3indicatedthat at thenonpermissive temperatureHSV-l(mP)ts66.4DNAdoes notbecomeencapsidatedandenveloped.Intheexperiments whose results are illustrated in Fig. 5, we detected viral DNAinthecytoplasmofcells infectedatthenonpermissive temperature with HSV-l(mP)ts66.4. To determine whether the cytoplasmic viral DNA represents DNA contained in capsids, replicateVerocell cultureswereexposed to 10 PFU
as PErS S
0 0> > > >>r
PS \at s Y5
si
Ci
tLC; C; ; ;PP CL~ CJzCL~^
F
*m
2hours 15hours 2 hours
Nuclei
15hours
Cytoplasm
FIG. 5. Autoradiographic imagesof32P-labeledBamHI fragments F and Xhybridizedtoelectrophoretically separatedBamHIdigestsof nuclear andcytoplasmicDNAs extracted fromreplicatecultures of Vero cells infected with parent, mutant,orrescued virus and maintained at34or39°C. The infectedreplicatecellcultureswereharvestedat2and 15 hpostinfection.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.152.485.456.692.2]c A
aX
-sc c0
X X2 X S: XaLA SL
*OEEFk
S."4 a .'; " x
15hours 18 hours
FIG. 6. Autoradiographic images of 32P-labeled BamHI
frag-mentsFand Xhybridizedtoelectrophoretically separated BamHI digests of DNA extracted from DNase-treated cytoplasms.
Repli-cate cultures of Vero cellswere infected with1(mP), HSV-1(mP)ts66.4, HSV-1(mP)Ra, and HSV-1(mP)Rb, maintainedat39'C for 15 or 18h, andthen harvested inabuffercontaining Nonidet P-40.Thecytoplasmwasthenseparatedfromthenucleiby centrif-ugationanddigested withDNasepriortotheextractionof DNA.
of parent,mutant,orrepairedviruspercell and incubatedat 39'C.Thecellswereharvested15or18h afterinfection and solubilized in nonionicdetergents.Thecytoplasmseparated fromthe nucleiby centrifugation wasdigested with DNase
(50
jig/ml;
15 min).Thisprocedure, describedby Vlaznyet al. (19), causes the degradationof free DNA but does not affect the DNApackaged in capsids and exported into the cytoplasmafterenvelopment.Afterdigestion, the DNAwasextracted, digested with BamHI, electrophoretically
sepa-rated in an agarose gel, transferred to a Zeta-probe
mem-brane, andhybridizedwithBamHI-FandBamHI-X labeled with32p.Asshown inFig. 6,viral DNA resistant toDNase
was detected in cells infected with parent and rescued viruses butnotinthecytoplasm ofcellsinfectedwith thets mutant. We conclude that, consistent with the electron microscopicobservations, incells infected with themutant virusDNA ismadebutnotpackagedintocapsids. Theviral DNA detected in the cytoplasm of cellsmaintained at the nonpermissivetemperature (Fig. 5) mostlikelyrepresented leakageofviral DNAfromrelatively labile nuclei harvested at 15 hpostinfection andwasnotencapsidated DNA.
DISCUSSION
The salient features of the results of this study are as
follows.
(i) The prototype HSV-1 strain used in this laboratory is HSV-1(F). Like many isolates passagedalimited number of times in cells in culture, this isolate is ts with respect to
replication incells in culture at39°C. On theoccasionswhen we had to select ts mutants, this laboratory has used HSV-l(mP),alaboratory strain isolatednearly 40 years ago and passaged serially numerous times at 39°C. In the course of attempts to introduce site-specific mutations, we have isolated a number of ts mutants with mutations which mapped at avariety of sites within the genome. We suspect that these mutations do not represent nonhomologous
re-combination between the parent virus DNA and mutated DNAintroduced as a plasmid. Rather, it is ourimpression that HSV-l(mP) hasaccumulated a large number of muta-tions whicharemaintained in the stock. We should note that this is not a novelphenomenon. A number of mutants have beendiscovered in recent years in stocks of viruses which servedasparent stocks for generationof mutantviruses (8, 9).The relevantissues withrespect tothis report are that(a) thetsmutationcanberescued by DNAfragmentscontaining one specificsetof sequences and (b) the mutation maps in UL15, oneof the most conserved genesin therepertoire of herpesvirus genomes. Although there has been much spec-ulation on the function of the UL15 gene, no experimental data have beenreportedto date.
(ii) The results reported in this paper indicate that cells infected with the virus carrying a tsmutationin UL15make viral DNA at thenonpermissive temperature. However, the viral DNAis notpackaged, since infected cells maintained at thenonpermissive temperature do not containcapsids with DNA. Moreover, whereas the cytoplasm of cells infected with wild-type and rescued viruses contained DNase-pro-tected viral DNA, such DNA was not present in cells infectedwith the mutantvirus.
We conclude that UL15 protein as deduced from the phenotypeof
HSV-1(mP)ts66.4
is involvedinthe packaging of viral DNA. The isolation ofthismutantopens the way to detailed studies of itsfunction.ACKNOWLEDGMENTS
Wethank Shu-Fen Chou forassistance with theelectron
micros-copy.
Thesestudieswereaidedby Public Health Servicegrantsfrom the National Cancer Institute(CA47451)andthe National Institute for Allergy and Infectious Diseases (AI24009) and byan unrestricted grant from the Bristol-Myers Squibb Program in Infectious Dis-eases.
REFERENCES
1. Baines,J.D., and B. Roizman. 1991.The openreading frames UL3, UL4, UL10, and UL16 aredispensable for the replication of herpessimplexvirus 1 in cell culture. J. Virol. 65:938-944. 2. Baines, J. D., and B. Roizman. 1992. The cDNA ofUL15, a
highly conservedherpes simplexvirus 1 gene, effectively re-places thetwoexonsof thewild-type virus.J. Virol. 66:5621-5626.
2a.Baines,J. D., and B. Roizman.Unpublished data.
3. Campadelli, G.,R.Brandimarti,C. DiLazzaro,P. L.Ward,B. Roizman, and M. R.Tormsi.1993.Fragmentationanddispersal ofGolgi proteins and redistribution ofglycoproteins and gly-colipids processed through Golgi following infectionwithherpes simplexvirus 1. Proc. Natl. Acad. Sci.USA90:2798-2802. 4. Challberg,M. D.1986. Amethodforidentifyingthe viral genes
required for herpesvirus DNA replication. Proc. Natl. Acad. Sci. USA83:9094-9098.
5. Costa,R.H.,K.G.Draper, T.J.Kelly,and E. K.Wagner.1985.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.115.244.74.372.2]Anunusualsplicedherpessimplex virustype 1transcriptwith sequencehomology toEpstein-Barr virus DNA. J. Virol. 54: 317-328.
6. Davison, A. J. 1992. Channel catfish virus: a new type of herpesvirus. Virology186:9-14.
7. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968. Character-izationof herpes simplex virus strains differing in their effects onsocialbehavior ofinfectedcells.J.Gen. Virol.2:357-364. 8. Everett, R. D., and M. L. Fenwiclk 1990. ComparativeDNA
sequence analyses of the host shutoffgenesof different strains ofherpessimplex virus:type2strainHG52 encodesatruncated UL41product. J. Gen. Virol. 71:1387-1390.
9. Fisher, F. B., and V. G. Preston.1986. Isolationand character-ization of herpes simplex virus type 1 mutants which fail to induce dUTPase activity. Virology 148:190-197.
10. Hoggan, M. D., and B. Roizman. 1959. The isolation and properties ofavariant ofherpessimplex producing multinucle-ated giant cells inmonolayer cultures in the presenceof anti-body.Am.J. Hyg. 70:208-219.
11. Honess, R. W., and B. Roizman. 1973. Proteins specified by herpessimplex virus.XI.Identificationand relative molar rates ofsynthesis of structural and nonstructural herpes virus poly-peptides in the infected cell.J.Virol. 12:1347-1365.
12. Morse, L.S.,L.Pereira, B.Roizman,and P.A. Schaffer.1978. Anatomy ofherpessimplex virus (HSV) DNA. X.Mapping of viral genesbyanalysis of polypeptides and functions specified by HSV-1 xHSV-2recombinants. J.Virol. 26:389-410. 13. Pellett,P. E., J. L. C. McKnight, F. J. Jenkins, and B.Roizman.
1985.Nucleotidesequenceandpredicted amino acidsequence
of a protein encoded in a small herpes simplex virus DNA fragment capableoftrans-inducingagenes.Proc. Natl. Acad. Sci. USA82:5870-5874.
13a.Poffenberger, K,and B.Roizman.Unpublished studies. 14. Post, L. E., A. J.Conley, E. S. Mocarski, and B. Roizman.1980.
Cloningofreiterated andnonreiteratedherpes simplexvirus 1 sequences as BamHI fragments. Proc. Natl. Acad. Sci. USA 77:4201-4205.
15. Powell,D., J. Franklin, F. Arisaka, and G. Mosig.1990. Bacte-riophage T4DNApackaging genes 16 and 17. Nucleic Acids Res. 18:4005.
16. Rao,V.B., and L. W. Black. 1988. Cloning, overexpression, and purification of the terminase proteins gpl6 and gpl7 of bacteriophage T4. Construction of a defined in vitro DNA packaging system using purified terminase proteins. J. Mol. Biol.200:475-488.
17. Roizman, B., L. Aurelian, and P. R. Roane, Jr. 1963. The multiplication ofherpes simplex virus.I. Theprogramming of viral DNAduplicationinHEp-2 cells. Virology21:482-498. 18. Spear,P.G.,and B.Roizman. 1972.Proteins specified byherpes
simplex virus. V. Purification and structural proteins of the herpesvirion. J.Virol. 9:143-159.
19. Vlazny, D. A., A. Kwong, and N. Frenkel. 1982. Site specific cleavage/packaging of herpes simplex virusDNAandthe selec-tive maturation of nucleocapsids containing full length viral DNA. Proc.Natl.Acad. Sci.USA79:1423-1427.
20. Wagner, E. K 1985. Individual HSVtranscripts: characteriza-tion of specific genes, p. 45-104. In B. Roizman (ed.), The herpesviruses, vol. 3. Plenum Press,NewYork.