0022-538X/91/126637-08$02.00/0
Copyright © 1991,AmericanSocietyforMicrobiology
Splice Site Skipping in Polyomavirus
Late
Pre-mRNA
Processing
YING LUOAND GORDONG. CARMICHAEL*
Department ofMicrobiology, University of ConnecticutHealth Center,
Farmington,
Connecticut 06030Received 1 July 1991/Accepted 27 August 1991
Polyomaviruslate nuclearprimary transcripts contain tandemrepeatsof the late strand of the viralgenome,
as a result of inefficient transcription termination and polyadenylation. Pre-mRNA processing involves the
splicing ofshort noncoding late leaderexonstoeach other(removing genome-length introns)andthesplicing of the last leader to acoding bodyexon (such asforthemajor virion structural protein, VP1). Asa result,
cytoplasmicmRNAscontain 1to 12 tandem leaderexonsattheir5' ends thatarefollowed byasingle coding
exon. To understandmoreabout how polyomavirusexonsare spliced together, we studiedadouble-genome construct consisting oftwotandem but nonidentical polyomavirus late transcription units. The alternating leaderexonsaredistinguishable fromoneanother but retainidenticalflanking RNA-processing signals,asfor
thealternatingVP1exons.Wetransfected thisconstructandderivatives of it intomousecells anddetermined which leaderexons aresplicedtowhich others and whichVP1exons areutilized. Results showed that leader
exons arealmostneverskippedduring splicing andarespliced sequentiallytooneanother. On the other hand, VP1exons were often skipped, with the VP1 exonclosest to the polyadenylation site splicing tothe nearest upstream leaderexon. Splice site replacementexperiments showed that VP1 exon skipping isnot dueto a relativeweaknessofits3'splicesiteortoanysequenceupstreamof theVP13'splice site. Exon skipping is also not the result ofsequences within the VP1 exon. Rather, VP1 3' splice site skippingcan be eliminated by
replacing theinefficientlate polyadenylation signal withanefficientone, orbyinsertinga5'splicesite between the VPI3' splicesiteand thelatepolyadenylation site. Thus,sequencesthatcomposethedistal border of the VP1 exon caninfluence usageof theupstream3' splice site.
Polyomavirus late pre-mRNA molecules contain optional
splice sites and polyadenylation sites. Polyomavirus late
transcription termination andpolyadenylation are both
inef-ficient, leadingto theproduction ofaheterogeneous
collec-tionof nuclearprimarytranscripts, some of which are many
times the size ofthe viral genome (1-3, 37, 38). Figure 1 illustrates the current model for how one such giant pre-mRNA molecule is processed into a mature cytoplasmic messageforthemajor viral structural protein, VP1. At the 5' ends of lateprimary transcriptsisanoncoding exon, the late leader. Ingiant transcripts,this exon appearsmultiple times.
DuringRNA processing, leaders are spliced to each other,
skipping internal VP1 exons and removing genome-length
introns.Inaddition, for mVP1 molecules a body splice site is chosen to attach the final late leader exon to a terminal
coding exon. Thus, maturelate cytoplasmic messages
con-tain,attheir 5' ends, 1 to 12tandemcopies of the late leader
exon that arefollowed by a single coding exon (20, 21, 25,
36).The averagelate message contains threetofour tandem late leader units (36). Aninteresting featureof this system is that thesplice sites andpolyadenylation sites thatarechosen
are identicalto those thatareskipped.
Wehave beeninterested in understanding how exons are
spliced to each otherduring polyomavirus late pre-mRNA
processing. Tobeginto address thisissue, we have made a
series ofconstructs containing a duplicated viral genome, but with distinct late leader and VP1 exons. We have
focusedontheexpressionof the message for VP1, themajor capsidprotein,becauseitaccountsfor 80to85%of late viral message (21, 35). We show here that,following transfection ofour constructs into mouse cells, late leader exons were spliced sequentiallytoeach other inprimary transcriptsand
* Correspondingauthor.
were rarely skipped. VP1 exons, however, were often
skipped, with the 3'-terminal VP1 exon splicing to the nearest upstream leader. Splice site and polyadenylation signalreplacement experiments showedthat VP1 exon skip-ping isnotdue toarelatively weak 3' splice site that isoften
ignored. Rather,skippingof the VP13'splicesite isstrongly influencedbythe strength of the nearest downstream poly-adenylation site and by the presence of a downstream 5'
splice site. These results suggest that exon skipping in
polyomavirus isnormallyduetoinefficientrecognition ofthe
latepolyadenylation site andprovideinvivo supportfor the exondefinition modelrecently proposed byRobberson et al.
(32).
MATERIALSANDMETHODS
Materials. Restriction enzymes, T4 DNA ligase, DNA
polymerase I large fragment (Klenow enzyme), T4 DNA polymerase, and T4 polynucleotide kinase were from New EnglandBiolabs andwereusedassuggestedbythesupplier.
RNase T2 was from Bethesda Research Laboratories. [a-32P]dATP, [t-32P]UTP,[y-32P]ATP, and avian myeloblas-tosis virus reverse transcriptase were from Dupont, NEN
Research Products. Polyomavirus strain 59RA has been
describedpreviously (17, 18).This strain is very similarin its
nucleotide sequence to that published for strain A3 (14). Plasmidp43.25.67 iswild-type polyomavirus strain A2 with an XhoI linker at the PvuII site at nucleotide (nt) 5128, inserted at the BamHI site into pAT153 (39), and was generouslyprovided byA.Cowie andR. Kamen. All recom-binant plasmids used here were propagated in Escherichia
coliJM83.
Cell culture andtransfections. Mouse NIH 3T3 cells were obtained from the American Type Culture Collection. The techniquesfor their
propagation
and transfection havebeen 6637on November 10, 2019 by guest
http://jvi.asm.org/
leader-leader splice leader-body splice
L <V L VP1
5' VXselAAzoeZZA_
LL VPI
_ AAAZZn2M-AAAAAAAA
FIG. 1. Model for the processing of polyomavirus late giant primary transcripts and the production of multiple, tandem, non-translated late leader unitsonmaturemRNAmolecules. Because of inefficienttermination by RNA polymeraseII,late viral transcripts are heterogeneous in length, withmany representing multiple cir-cuits of the viral genome. The example shown is for atranscript created bytwo passes through the viral late region. During pre-mRNAprocessing,aleader-body splice joins the 3'-most noncoding late leader exon (L) to a message-coding body (here mVP1). In addition, leader-leader splicing removes a genome-length intron, yieldingafinal mVP1messagewithtwotandem leader unitsatits 5' end.
3'ss 5'ss
I L
SLM
1-13
1-13 5'ss
3'ss
AP1
described elsewhere (10). Briefly, all transfections were
performed byamodification of the calcium phosphate
trans-fection procedure of Chen and Okayama (11). Cells were
seeded ata concentration of 106 cells per 150-mm plate in
Dulbecco's modified Eagle medium supplemented with 10% bovine calfserumplusL-glutamine and penicillin and strep-tomycin at 37°C in 5% CO2 approximately 48 h prior to
transfection. Sixteen hours prior to transfection, the cells
wereserumstimulatedby the addition of fresh medium. The
total amount of DNA per transfection was 40 p.g. Before transfection,recombinantplasmidswerecutwithEcoRIand diluteligated with T4 DNA ligase as described previously (6).
Harvesting of cytoplasmic RNA. Cellswereharvested44to 48 h after initiation of transfection. Each plate was rinsed
with lx phosphate-buffered saline-minus (PBS-) (without
Mg2+ and Ca2+), and RNA was harvested as described elsewhere (10). Briefly, cells were scraped into PBS- and
pelleted by centrifugationat1,000 x gfor3 min. Thepellet wasresuspendedinPBS- andcentrifuged againat1,000 xg
for 3min.The cellpelletwasresuspendedin 3 ml ofNonidet P-40 lysis buffer and left on ice for 10 min. Nuclei were
removed by centrifugation at 1,000 x g for 3 min. To the supernatant(the cytoplasmicRNAfraction)wasadded 1.42
g of guanidine isothiocyanate powder. RNA was separated from DNA by centrifugation through a5.7 M cesium chlo-ridecushion for 20hat110,000x g(12).ThepelletedRNAs were drained well and resuspended in 300
RI
of 10 mMTris-Cl (pH7.0)-i mMEDTA(pH 7.0)-0.2%sodium
dode-cyl sulfate and ethanol precipitated. RNAs were
resus-pended in100 Il of distilled H20.
Construction of plasmids. All cloning was done by
stan-dardprocedures.Thestructuresof themajor plasmidsused inthis study areshown schematically in Fig. 2.
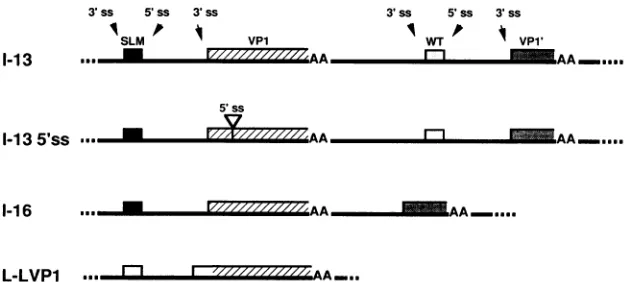
(i) Double-genome constructs. The construction of plas-mids pl-13 and pl-16isdescribed in detail elsewhere (27a).
Constructs 1-13 and 1-16 containduplicated viral genomes,
but withdistinguishablelate leader and VP1exons. Figure2 illustrates schematicallythe intron-exon structuresof these constructs. Plasmidpl-135'ss is thesameaspl-13,excepta
late leader 5'splicesitewasinserteddownstream of the VP1
3'ss 5'ss
WT
3'ss
I vp1' _M
man _ v zzzgzj</ "AA I^ |AAsmmn s
5'ss
son _ r //
//////AA,,,,,o....
1-16 son m Ui;///z7AA~§AA.
L-LVP1 ..
FIG. 2. Schematic diagram of the major constructs used in this study. Constructs were made as described in Materials and Methods. Before transfection, each was digested with EcoRI, which liberated the polyomavirus sequences from the plasmid backbone, and recircularized with T4 DNA ligase. 1-13 contains two copies of the viral late region, but the first copy has a substituted late leader exon sequence(SLM[6]), while the second has a truncated VP1-coding exon (VP1'). 1-135'ss is thesame as1-13,exceptthe late leader 5' splice site has beeninsertedinto the coding region of VP1, 80 nt downstream of the VP1 3' splice site.1-16is the same as1-13,except fordeletion ofsequencesincludingoneintact early coding region, replication origin, late promoter, and late leader exon. L-LVP1 (8) is a single-leader mVP1cDNAwhichalsocontains an upstream single-leader exon (flanked by its normal splice sites) and an intron of 165 nt. The important splice sitesin theseconstructs are indicated. There is a minor 3' splice site (not shown) between the late leader and the VP1 3' splice site for theproductionofmessages encoding VP3. AA, polyadenylation signal AAUAAA.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.56.283.75.353.2] [image:2.612.142.459.504.645.2]3' splice site. To do this, we inserted an 87-bp fragment into the EcoRV site (14 bp downstream of the VP1 3' splice site). This fragment contained 44 bp upstream of the mutant SLMY (6) 5' splice site and 15 bp downstream of the leader 5' splice site. A 14-bp linker was added at both ends, giving anXhoIsite upstream and a ClaI site downstream of the new 5' splice site. In this 81-bp fragment, only 21 bp derive from the wild-type polyomavirus genome, 6 bp upstream and 15 bp downstream of the leader 5' splice site.
(ii) Construct L-LVP1. Plasmid L-LVP1 was made by inserting a 165-bp fragment containing the late leader exon and flanking sequences (including the late promoter just upstream of the late leader 3' splice site) into a genome in which the VP1 intron had been precisely removed by oligo-nucleotide-directed mutagenesis. This construct was orig-inally made for other experiments in the laboratory and has been described in detail elsewhere (8).
PCR assays. Oligonucleotide primers were made by using a Milligen/Biosearch Cyclone DNA Synthesizer and the phosphoramidite chemistry.Oligonucleotide 275 (5'-TATCA CCGTACAGCCTTG) anneals across the VP1-late leader splice junction, with the terminal 3 bases annealing to the late leader and the rest annealing to the VP1-coding exon. Oligonucleotide 278 (5'-CCTGACATTTTCTATTTTAAG)
anneals to late cDNA molecules immediately upstream of the late leader exon. Oligonucleotide PE (5'-CTATGACT GTTGCCC) anneals to late mRNA just inside the VP1 exon. Oligonucleotide 279 (5'-TCCTAGATGAAAATGGAG) an-neals to the truncated VP1' exon but not to the wild-type VP1 exon.
The polymerase chain reaction (PCR) (34) was performed following reverse transcription of the RNA.Oligonucleotide 275, 279, or PE was added to the RNA in a buffer recom-mended by Perkin-Elmer Cetus. AMV reversetranscriptase (22 U) was added and allowed to incubate for 1 h at 45°C. TaqI DNA polymerase and oligonucleotide 278 were then added, and PCR continued for 30 cycles. Each cycle had three sections. The first was a 2-min incubation at 92°C. After a 30-s ramp time, the incubation was continued for 1
min at 45°C. Finally, after a 0.5-min ramp time to 72°C, incubation wascontinued for 1.5min. A 10-min incubation at 72°C followed the final cycle. Reaction products were re-solved on 6% polyacrylamide-urea gels, and bands were detected byautoradiography. It is important to note thatthe PCRamplification of RNA performed in ouranalyses accu-rately and reliably reflected the RNA amounts in our sam-ples. Suchverification has previously been presentedby our laboratory (20).
RESULTS
Constructs and experimental design. We constructed a
series of plasmids that contain duplicatedviral genomes, but withdistinguishable late leader and VP1 exons. Each of the constructs used contains at least one intact early coding
region and DNA replication origin, so each produces early
viral proteins and replicates in transfected mouse cells. Figure 2 illustrates schematically the intron-exon structures of the constructs used in the work described here. The primary construct is 1-13: it has two distinguishable leader exons and two distinguishable VP1 exons. The wild-type
(WT) and substituted (SLM) late leaders are of different sequence and differ by 6 nucleotides in length. We have shownpreviously that a virus with the SLM leader used here grows like wild type and is notdefectivein latepre-mRNA
processing (4, 6). TheVP1' exon contains a deletion within
the
wild-type
VP1 exon and is 81 nucleotideslong.
Thesplice
sitesbordering
thewild-type
and SLM leader exonsare
identical,
as arethe sequencesflanking
theVP1andVP1' exons.Thisconstruct wasdesigned
such thatafter transfec-tion into mousecells,
primary
transcripts
withalternating
wild-type and SLM leader exons and
alternating
VP1 andVP1' exons are
produced.
Since late promoteractivity
requires
only
a very smallregion
just
upstream ofthe late leaderexon(10,
33a),
latetranscription
from1-13caninitiate upstream of either the SLM orwild-type
leader exon.Construct I-135'ssis thesame as I-13 exceptthata5'
splice
site
(derived
fromthe lateleader)
has been inserted into the VP1 exon, 80 nucleotides downstream ofthe VP1 3'splice
site. Construct1-16is relatedto1-13 butcarriesadeletion of
the second
(wild-type)
late leaderregion.
L-LVP1 is agenome
containing
alateleaderunitupstream ofaprespliced
leader-VP1 cDNA.This constructmimicswild
type
butwith the VP1 3'splice
sitereplaced
witha leader3'splice
site.For all
experiments,
polyomavirus
single-
ordouble-ge-nome inserts from these constructs were excised from the parent vector backbones and recircularized
by
diluteliga-tion. Transfectionofthese recombinant viral genomes into
permissive
mouseNIH 3T3 cells mimics the infection proc-ess: late viral geneexpression
proceeds
in a manner thatproduces
giant
primary
transcripts.
Leaderexons are
spliced
sequentially
tooneanother ingiant
primarytranscripts.
Because mature mVP1 molecules can containupto12 tandemleader unitsattheir5' ends(20),
weasked how leader exons are chosen in the late
pre-mRNA
processing
pathway.
Can leader exons beskipped during
leader-to-leader
splicing
in the same way that VP1 exons appear to beskipped,
or aremultiple
leaders ingiant
transcripts
spliced
sequentially
to one another? This was examinedby
a PCR assay which has been shownby
ustoaccurately
andreliably
reflect the initialamount ofRNA in oursamples
(34).
Briefly,
anoligonucleotide
complementary
to a
leader-to-body
splice
junction (275)
was annealed tocytoplasmic
RNA isolated from transfected cells andex-tended
by
reversetranscription.
Thena32P-labeled
oligonu-cleotide
(278)
which binds to the 5' end of the reversetranscript
wasadded,
and the PCRwasused toamplify
theproducts.
The labeledoligonucleotide
annealsimmediately
upstream of the late leader exon. The sizes of the bands
observed after
gel
electrophoresis
reveal the distribution ofleaders at the 5' ends ofthe messages
(20).
Figure
3 shows theresultsofsuchaPCRexperiment
using
oligonucleotide
275,
which binds to thejunction
of the leader-to-VP1splice
as wellas tothejunction
of the leader-to-VP1'splice.
Inthisexperiment,
messagescontaining
oneandtwotandem leader unitswereobserved for wildtypeand constructsI-13and1-16.
Wild-type
viral RNAwasused hereto
provide
size markers forone and two tandemwild-type
leader units
(each
unit is 57nt).
1-16only
encodes SLMleaders,
which are 6 nucleotides shorter thanwild-type
leaders and which can be used as additional size markers.Thisaccountedfor thealteredleader ladderobserved forthis construct and allowed its use for size markers. Because
therearetwolatepromotersin
1-13,
weexpected
toseebothwild-type
and SLM leadersignals
attheone-leader level for thisconstruct. This was observed. At the two-leaderlevel,
however,
themajor
band seen was intermediate in sizecompared
with twowild-type
leaders or two SLMleaders,
indicating
that most messages contained onewild-type
leader exon
spliced
to one SLM leader exon. Ifwild-type
leaderswere
spliced
together
(skipping
SLMleaders)
in1-13 RNAprocessing,
or if SLM leaders werespliced
together
on November 10, 2019 by guest
http://jvi.asm.org/
275
VP1orVP1'
278
0
Pt
A 2 __.",--
1-..--leader structures
1 zi.
SLM
B
1-13 K .AAA A
1-16 A.
FIG. 3. PCR assay to determine whether leader exons splice
sequentiallytoeach other. Theoligonucleotides used for PCR (275 and 278) are shown at the top. The assay was performed as described in thetext, usingcytoplasmic RNA isolated from mock-transfected cells and cells mock-transfected with religated DNAs froma
wild-type viralgenome(WT),1-13,or1-16.Reaction productswere resolved on a 6% polyacrylamide-7 M urea gel and detected by autoradiography.Quantitationwasperformed withaBetagen
Beta-scope 603 Blot Analyzer. The expected positions of leader
struc-tures are shown on the left. Black boxes represent SLM leader units, which are 6 nt shorter than wild-type (WT) leader exons, shown as white boxes. At the bottom is an illustration ofagiant primary transcript, with sequential leader splicing indicated.
(skipping wild-type leaders),then bands wouldappearatthe
positions of molecules with two wild-type or two SLM
leaders. Such material accounted for less than 2% of the signal as determined with a Betagen Betascope Blot
Ana-lyzer, leadingustoconclude that the 1-13 leaderexonsmust
besplicing inasequentialfashion. Results atthethree-and
four-leader positions (data not shown) are also consistent with sequential leader splicing, but those results are more complex, asa result of the multiplepromoters that exist in
our constructs. Thus, although VP1 exons are frequently
skipped during leader-to-leader splicing, late leader exons are spliced sequentially to one another and are rarely skipped.
VP1exons arespliced tothenearest upstreamleaderexon.
Although leader-to-leader splicing does not involve leader exon skipping, the possibility remained that leader exons could be skippedduring leader-to-bodyexon splicing. Since we know the SLM leader isupstreamof the VP1 exon and
[image:4.612.321.531.73.363.2]the wild-type leader precedes the VP1' exon in primary transcripts from construct 1-13, then by measuring which leader is splicedto which exon, we can deduce whether a
FIG. 4. (A) PCR analysis of VP1 and VP1' exon splicing in constructs1-13and 1-16. Theanalysiswasasdescribed in thelegend
toFig. 3, exceptoligonucleotides 278 and279were used for VP1' spliced products, and oligonucleotides278 and PE were used for VP1 splicedproducts. Oligonucleotide 279only binds tothe VP1'
exon,allowingustoanalyzethesplicingof thisexonindependently
ofthesplicingofthewild-type (WT)VP1exon,whilePEbindsonly
totheVP1exon.Theinterpretationof leaderstructuresisshownto
the right. (B) Schematic representation of some of the spliced products inferredfrom the datashown in panelA. Construct 1-13 containstwolatepromoters, eachimmediatelyupstream ofaleader
exon. Only products that derive from the upstreampromoter are
shown.Since the 1-13genomeiscircularly permuted,the constructs derivingfrom the second promoterareanalogous.
VP1 or a VP1' exon is only spliced to the leader exon
immediately upstream of it. The PCR technique allowed such an analysis andwas done forboth the VP1 and VP1' exons. Atypicalresult is shown inFig. 4A. Wild type(WT)
and1-16wereused for sizemarkers,aseachcontainsonlya
single type ofleader exon (SLM). At the one-leaderlevel, almostall thesignalobserved for1-13wasattheSLM leader
position when using a VP1-specific probe and at the wild-type position when using a VP1'-specific probe. This indi-catesthat in thisconstruct, most VP1 orVP1' splicingisto the immediately upstream leader exon. Again, the leaders werenotskipped during pre-mRNA splicing, while the VP1 and VP1' exons were both skipped. Since the VP1' exon retainsonly81 ntderivingfrom thewild-typeVP1 exon [14
ntadjacenttothe VP1 3'splice siteand 67ntjustupstream of the poly(A) site], exon skipping is not likely to be the result ofsequences within the late viral coding region that inhibit theuseof theVP1 3'splice site.Wecannotexclude,
however, that thesesequencesmightsomehow influence the useof the VP1 3'splicesite.Figure4Billustratessomeof the
splicing eventsdemonstratedby thesedata.
leader
structures
E==
-=
2 78
C') co
0 v
--
-WT =
SLM _
= WI
_ AIM
gp
-x
L) Cl) co
0 v-
v-.7 -1 -1
--II.-
-
I--I
SL WT .. SLM
-7 Em"M AA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.72.269.74.372.2]Exons are not skipped because the VP1 3' splice site is weaker than the late leader 3' splice site. Figure 4A also shows that leader splicing in1-16can occur toeither the VP1 or the VP1' exon, indicating that exon skipping does not have to involve splicing only toa downstream leader exon. This construct contains only an SLM leader, which is upstream of a VP1 exon. The distal VP1' exon has no leader immediately upstream of it. As seen in Fig. 4A,however, the VP1' exon can still be spliced efficiently to the SLM leader exon, following skipping of the upstreamVP1 exon, indicat-ing that VP1 exon skippindicat-ing does not result from the presence of a downstream leader exon and is therefore not likely to be the result of especially strong leader splice sites that out-compete the VP1 3' splice site.
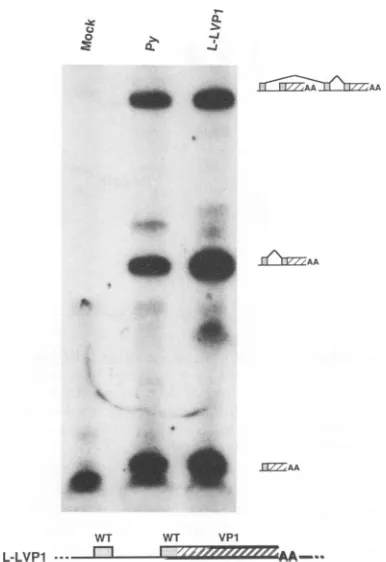
A possible explanation for VP1 exon skipping in polyoma-virus would be that the VP1 exon has an inherently ineffi-cient 3' splice site which is poorlyrecognized by the splicing machinery of the cell. To determine whether this model is correct, we analyzed construct L-LVP1. In L-LVP1, the VP1 3' splice site has been replaced with the late leader 3' splice site. Thus, a 3' splice site that is frequently skipped has been replaced with one that is rarely skipped. In addi-tion, this construct lacks almost all viral sequences that normally exist upstream of the VP1 exon. Cytoplasmic RNAs from L-LVP1 and wild-type polyomavirus were sub-jected to PCR analysis, and theresults are shown in Fig. 5. Bands at the one-, two-, and three-leader positions are shown. Clearly, L-LVP1 generated late messages with mul-tiple leader structures, so the modified VP1 exon of this construct must be skipped. Theimportantresult here is that the ratio of mRNAs with two leaders to those with three leaders for L-LVP1 is the same as the ratio ofmRNAs with one leader to those with two leaders for wild type. Quanti-tation of these results with a Betagen Betascope Blot Ana-lyzer revealed that in this experiment the frequency of exon skipping for wild type was about 40%, while the frequency for L-LVP1 was about 25%. In other experiments, the frequency of splice site skipping in wild type and L-LVP1 has been virtually identical. Weconclude from these exper-iments thatVP1 skipping does not occur because the VP1 3' splice site is weaker than the late leader 3' splice site, or because sequencesupstream of the VP1 3' splice site inhibit its use.
Late polyadenylation signal strength affects exon skipping.
Since VP1 exon skipping appears not to result from
rela-tively strong or weak splice sites or from intron or exon
sequences flanking the VP1 3' splice site, we asked whether
skipping might be influenced by the late polyadenylation
signal.We thereforeanalyzed a viral variant, ins5, provided
by N. Acheson. This mutantcontains two tandem polyade-nylation sites in the late transcription unit: one is the wild-type late polyadenylation site, and the other, just up-stream, is adifferent, moreefficient rabbit 3-globin polyade-nylation site (23). Wild-type or mutant viral genomes were
transfected into mouse cells as in earlier experiments, and the resultant cytoplasmic RNA was subjected to a leader
PCR assay. The results are shown in Fig. 6. As expected,
wildtypeproduced alargeproportion of VP1 messages with
multiple leaders. In contrast, however, mutant ins5 pro-ducedonly afew latemVP1 molecules which had more than
a singleleader, demonstrating that an efficient
polyadenyla-tion signal can suppress exon skipping. Quantitation with a Betagen Betascope Blot Analyzer revealed that ins5
pro-ducesabout95% single-leadermolecules in this assay. These results for ins5 are in good agreement with those reported
0 0. ..4
:^ AhI2~AA
.6
M 4
.
4b
L-LVP1
JflE2AA
AA
WT WT VP1
m I A
FIG. 5. PCRassay to determinewhether VP1 exon skipping is dueto arelativelyweak VP1 3' splice site or to a relatively strong leader 3' splice site. Cells were transfected with religated DNAs fromwild-typeconstruct pPyBS (Py)orplasmidL-LVP1.
Cytoplas-mic RNA was isolated44 hlater and subjected toPCRanalysis as
describedin thelegend toFig. 3.The positions expected formVP1 molecules with one, two, or three tandem leader units areindicated, and splicing events that generate the structures observed for L-LVP1 are shown to the right of the bands. Band intensities were determined withaBetagen Betascope BlotAnalyzer. The mocklane here shows an artifactual bandjust below the position of
single-leader mVP1 molecules. WT, wild type.
earlierby Lanoix and Acheson (23) forthe polyadenylation efficiency of this mutant.
A 5' splice site within the VP1 exon can prevent exon
skipping. The suppression of exon skipping by an efficient polyadenylation signalisdifficult tointerpret. Efficient
poly-adenylation is usually associated with efficient transcription
termination, as has been reported even for mutant ins5(23). Thus, the results shown in Fig. 6 might be the trivialresult of limited splicing choices in short transcripts. However, VP1 exon skipping could also be suppressed by another type of mutation. Construct 1-13 5'ss is identical to 1-13, except a
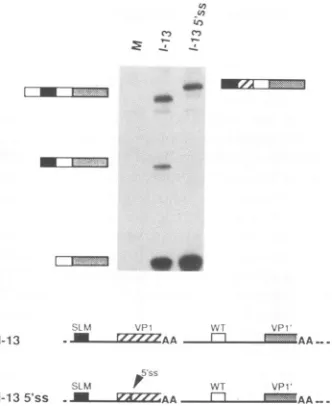
small DNAfragment comprising the late leader 5' splice site has been inserted downstream of the VP1 3' splice site. PCR analysis witholigonucleotide279 (specificforthe VP1'exon)
was used to examine exon skippingfrequency in this mutant, and theresults are shown inFig. 7. Strikingly, the VP1exon
in this construct is now almostnever skipped, asevidenced by the virtual absence of a band representing two tandem leaders spliced to the VP1' exon. Rather, the SLM leader is spliced efficiently to the VP1 3' splice site, and the inserted 5' splice site is used to splice to the downstream wild-type leader exon (top band in the I-13 5'ss lane). As would be expected, the VP1' exon is still efficiently skipped (data not
shown).
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.347.540.77.358.2]3 leaders
2leaders
1 leader
FIG. 6. Effect of poly(A) site strengthon VP1 exon skipping. Cells were mock transfected or transfected with religated DNAs from pPyBS (wildtype)orins5. CytoplasmicRNAwasisolated 44
h later and subjectedtoPCRanalysisasdescribed in the legendto
Fig. 3.The positionsexpected for mVP1 molecules withone,two,
orthreetandem leaderunitsareindicated.
Takentogether withtheabove data,weconclude thatVP1
exon skipping isalmostcertainlynotduetoaweak3' splice site orto inhibitory flanking sequences. Rather, use of the
VP1 3'splice siteappearstobe directlycoupledtotheuseof adownstreamRNAprocessing signal,eithera polyadenyla-tion signal or a 5' splice site.
DISCUSSION
Polyomavirus late pre-mRNAprocessing is unusual. Late viral nuclear primary transcripts containtandem repeats of
(0
LI)
C _ 1~ am
1-13 - - AAA m___AA
.--SL M WT VP<.
1-135'ss . AA rl ~AA--A
FIG. 7. Effect of a downstream 5' splice site on VP1 exon
skipping.Cells were mocktransfected or transfected withreligated
DNAs from I-13or I-135'ss. Cytoplasmic RNA wasisolated 44 h
laterandsubjectedto PCRanalysisasdescribed in thelegendtoFig.
3,using oligonucleotides278 and279,the latter of which is specific
for the VP1'exon.Interpretationsof theproductsare shownbeside
thelanes. WT,wildtype.
the late strandof the viral genome.
During
processing,
shortnoncoding 5' late leader exons are
spliced
to eachother,
removinggenome-length introns which containunused poly-adenylationsitesand 3'splice
sites. We have been interested in understanding how thisprocessing
isaccomplished
and how splice sites andpolyadenylation
sites areskipped.
Inthisstudy,weshowed thatleaderexons are
spliced
sequen-tially to one another in
giant
primary
transcripts and,
like most internal exons in known pre-mRNAmolecules,
arerarely skipped during splicing (Fig. 3 and 4). On the other
hand,
VP1-coding
exons areoftenskipped.
Hereweshowed that whenaVP1 3'splice
site isselectedforuse,it isspliced
to thenearest upstreamlate leader unit(Fig. 4).
HowareVP13'splicesites
skipped?
Aslate leaderexons appear to bespliced
sequentially
to one another ingiant
primarytranscripts, it is reasonableto assumethat the 5' and 3' splice sitesflanking
the leader areefficiently
selected in vivo. This ledus toconsider that the VP1 3'splice
sitemight
be recognized less
efficiently
(compared with the leader 3'splicesite), leadingtofrequentexon
skipping.
This possibil-ity was ruled out,however,
when we couldeffectively
replacethewild-typeVP1 3'splice
site withthe late leader 3'splice siteand still observe VP1 exon
skipping
atthe samefrequency as for wild type
(Fig.
5).Similarly,
when thedownstream leader exon was
deleted,
as inconstruct1-16,
VP1exons were still
skipped, by
leadersplicing
to adistant downstream VP1 3'splice
site(Fig. 4).
These results indi-cated that intron sequences upstream of the VP1 3'splice
site and VP1exonsequences
likely play
noimportant
role in theobservedexonskipping.
Since we were unable to affect VP1 exon
skipping by
exchanging splice sites,
wecompared
skipping
in several constructsthatdifferedintheirlatepolyadenylation
signals.
Mutant ins5 was constructed and
analyzed
previously
by
Lanoix and Acheson(23) andcontainstwo
polyadenylation
signals
in the lateregion,
onebeing
thewild-type
polyoma-virus signal and the other
being
from the rabbitP-globin
gene.In agreementwith the results
reported
by
Lanoixand Acheson (23), wefound that mostlate messages from ins5 have asinglelate leaderunit at their 5' ends(Fig. 6). Thus,
VP1 exons are
rarely
skipped
inthismutant.Normally,
polyomavirus
latetranscription
termination is inefficient (38). It ispossible
that thefragments
inserted toprovide
additionalpolyadenylation signals
in mutant ins5 containtranscription
terminationsignals
aswell.Whilemoreexperiments
areneededtorigorously
rule outthispossibil-ity, the sequences added in this construct are
short,
andanefficient
polyadenylation
signal
isnotsufficient for transcrip-tion termination.Transcription
terminationrequires
bothpolyadenylation
andother,
downstream sequences(13, 23,
27, 40). Levittetal.(26) demonstratedthat,
whenit isplaced
withinan
intron,
anefficientsynthetic
polyadenylation
signal
closelyrelated totherabbit,3-globin
polyadenylation signal
used
in
ins5 still does not preventtranscription reading
through until the end of the intron several hundred basesaway.
The suppression ofexon
skipping
seen withmutant ins5couldbe mimickedby
placing
a5'splice
site downstream ofthe VP13' splice site andbefore the inefficient late
polyade-nylation site (Fig. 7). Thisagain
demonstratedclearly
thatexon
skipping
could beaffectedby
changing
thesignals
that compose thedistalborderofthisexon. Inother worknowin progress, we areplacinga5'splice
site atvariouspositions
within the VP1 exon to examine the effect ofexon size on
splice site skippingin this system.
Howcould
polyomavirus
lateexonskipping
resultfromanon November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.134.222.78.243.2] [image:6.612.97.263.457.660.2]inefficient late polyadenylation site? Our working model, consistent with the results presented here, is that there must be coordinate recognition and use of the 3' splice site and polyadenylation signal that demarcate the borders of the terminal coding exon of pre-mVP1 molecules. In this model, recognition of a polyadenylation site for use is normally associated with the use of the upstream VP1 3' splice site-neither site is used unless both are used. In giant polyomavirus late primary transcripts, the polyadenylation site is inefficiently recognized; when this polyadenylation signal is not used, the upstream VP1 3' splice site is also
ignored. This model is also consistent with work reported elsewhere (27a), in which we examined 3'-end formation in a number of polyomavirus constructs including the double-genome constructs used in the study described here. In that work we showed that, when the wild-type late polyadenyla-tion signal was duplicated, the signal closest to the VP1 3' splice site was normally chosen, even though transcription proceeded past the downstream site, allowing the efficient accumulation of messages with multiple leaders. However, insertion of a 3' splice site just upstream of the second polyadenylation site allowed the use of both polyadenylation signals. Also, deletion of a late polyadenylation signal in a double-genome construct did not interfere with use of the upstream VP1 3' splice site, showing that VP1 3' splice site choice is independent of polyadenylation site choice. In other words, VP1 3' splice site usage, while normally cou-pled to polyadenylation site use, can still occur if the VP1 exon is placed in a different context.
Our findings lend in vivo support for the exon definition model of splice site selection proposed recently by Robber-son et al. (32). In that model, both boundaries of an exon must be recognized, ordefined, before RNAprocessing can occur at either one. Thus, forinternal exons, a 3' splice site and a downstream 5' splice site must both berecognized; for terminal exons, apolyadenylation signal could replace a 5'
splice site. For a terminal exon, if thepolyadenylation signal was not recognized, then the terminal exon would not be
properly defined, and the VP1 3' splice site could not be
used. Although one might envision regulation of VP1 exon skipping as occurring at either end of thisterminal exon, i.e., through an inefficient VP1 3' splice site or through an inefficient late polyadenylation site, the data presented here strongly suggest that the exon skipping observed in polyo-mavirus is due to the inefficiency of the latepolyadenylation
site.
The modelpresented for the coordinate choice of a splice site and apolyadenylation site isconsistent with a number of published observations on alternative splice site and poly-adenylation site choice in naturally occurring complex tran-scription units. In such systems, there isgrowing evidence that polyadenylation and splicing are not independent pro-cesses, i.e., regulation at either end of a terminal exon can affect exon skipping or polyadenylation site choice. In one example of cell-specific regulation, studies on the switch between secreted and membrane forms of immunoglobulin
M ,u heavy chain have suggested that this switch isregulated at the level ofpolyadenylation site choice (15, 19, 22, 28, 33). In the immunoglobulin M system, choice of alternative polyadenylation signals leads to altered splicing pathways, and there may be competition between splicing and poly-adenylation (30, 31). Incontrast, the expression ofcalcitonin and the calcitonin gene-related peptide, although controlled in acell-specific manner from a complex transcription unit containing two polyadenylation sites, appears to be regu-lated at the step of splice siterecognition, which in turn leads
totheuseofthenearestdownstreampolyadenylationsignal (7, 16, 24).
Other results froma number ofrecent studies have been
interpreted as demonstrating that splicing dominates over polyadenylation. Ifanefficientpolyadenylationsite is
placed
within an intron, then itcan be skipped; if the same siteis
placed within an exon, or if surrounding splice sites are
mutated, then it is used (5, 9, 26). Niwa et al. (29) have
suggestedthatplacementofapolyadenylation site withinan
introninactivates polyadenylation notbecause the polyade-nylation machinery cannot compete with the splicing
ma-chinery, but because it cannotinteract with it, reflecting an
underlying polarity of pre-mRNA processing signals. Our
results are consistent with this view. ACKNOWLEDGMENTS
We thank N. Achesonforproviding mutant ins5 and N. Barrett, E. Carmichael, R. Hyde-DeRuyscher, and A. Roome for helpful
comments onthismanuscript.
This work was supported by grantCA 45382 from the National CancerInstitute.
REFERENCES
1. Acheson, N. 1976. Transcription during productive infection with polyomavirusandSV40. Cell 8:1-12.
2. Acheson, N. 1984. Kinetics andefficiencyofpolyadenylation of late polyomavirus nuclear RNA: generation of oligomeric poly-adenylated RNAs and theirprocessing intomRNA. Mol. Cell. Biol. 4:722-729.
3. Acheson, N. H. 1978. Polyoma giant RNAs contain tandem repeats ofthe nucleotide sequence of the entire viral genome. Proc. Natl. Acad. Sci. USA 75:4754-4758.
4. Adami, G., and G. G. Carmichael. 1987. Thelength butnotthe sequenceof thepolyoma virus late leaderexonisimportant for both late RNA splicing and stability. Nucleic Acids Res. 15: 2593-2610.
5. Adami, G., and J. R. Nevins. 1988. Splice site selection domi-nates over poly(A)site choice in RNA production fromcomplex adenovirus transcription units. EMBO J. 7:2107-2116. 6. Adami, G. R., and G. G. Carmichael. 1986. Polyomavirus late
leader region serves an essential spacerfunction necessary for viability and late gene expression. J. Virol. 58:417-425. 7. Amara, S. G., R. M. Evans, and M. G. Rosenfeld. 1984.
Calcitonin/calcitonin gene-related peptide transcription unit: tissue-specific expression involves selective use ofalternative polyadenylation sites. Mol. Cell. Biol. 4:2151-2160.
8. Barrett, N. L., G. G. Carmichael, andY. Luo. 1991. Splice site requirement for theefficientaccumulationofpolyoma virus late mRNAs. Nucleic Acids Res. 19:3011-3017.
9. Brady, H. A., and W. S. M. Wold. 1988. Competition between splicing and polyadenylation reactions determines which ade-novirus region E3 mRNAs are synthesized. Mol. Cell. Biol. 8:3291-3297.
10. Cahill, K. B., andG. G. Carmichael. 1989. Deletion analysis of the polyomaviruslate promoter: evidenceforbothpositive and negative elements in the absence of early proteins. J. Virol. 63:3634-3642.
11. Chen, C., and H. Okayama. 1987. High efficiency transforma-tion of mammalian cells by plasmid DNA. Mol. Cell. Biol. 7:2745-2752.
12. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W.J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 18:5294-5299.
13. Connelly, S., and J. L. Manley. 1988. A functional mRNA polyadenylation signal is requiredfortranscription termination by RNA polymeraseII. Genes Dev. 2:440-452.
14. Deininger, P. L., A. Esty, P. LaPorte, H. Hsu, and T. Fried-mann. 1980. The nucleotide sequence and restriction enzyme sites of the polyoma genome. Nucleic Acids Res. 8:855-860. 15. Early, P., J. Rogers, M. Davis, K. Calame, M. Bond, R. Wall,
on November 10, 2019 by guest
http://jvi.asm.org/
and L. Hood. 1980.Two mRNAs can beproduced from asingle immunoglobulin gene by alternative RNA processing pathways. Cell20:313-319.
16. Emeson, R. B., F. Hedjran, J. M. Yeakley, J. W. Guise, and M.G.Rosenfeld. 1989. Alternativeproduction of calcitonin and CGRP mRNA is regulated at the calcitonin-specific splice acceptor. Nature(London)341:76-80.
17. Feunteun, J., L. Sompayrac, M. Fluck, and T. L. Benjamin. 1976. Localization of gene functions in polyoma virus DNA. Proc. Natl. Acad. Sci. USA68:283-288.
18. Freund, R., G. Mandel, G. G. Carmichael, J. P. Barncastle, C. J. Dawe, and T. L. Benjamin. 1987. Polyomavirus tumor induction in mice: influences of viral coding and noncoding sequences on tumorprofiles.J. Virol.61:2232-2239.
19. Galli,G., J. W. Guise, M. A.McDevitt, P. W. Tucker, and J. R. Nevins.1987.Relative positionand strengthsofpoly(A)sitesas well as transcription termination are critical to membrane ver-sus secreted >.-chain expression during B-cell development. GenesDev. 1:471-481.
20. Hyde-DeRuyscher, R. P., and G. G. Carmichael. 1990. Poly-omavirus late pre-mRNA processing: DNA-replication-associ-ated changes in leader exon multiplicity suggest a role for leader-to-leader splicing in the early-late switch. J. Virol. 64: 5823-5832.
21. Kamen, R., J. Favaloro, and J.Parker. 1980. Topography of the three late mRNAs of polyoma virus which encode the virion proteins. J. Virol. 33:637-651.
22. Kemp, D. J., G. Morahan, A. F. Cowan, and A. W. Harris. 1983. Production of RNA for secreted immunoglobulin chains does notrequire transcriptiontermination 5' to the M exons. Nature (London)301:84-86.
23. Lanoix, J., and N. Acheson. 1988. Arabbit ,-globin polyadenyl-ation signal directs efficient termination of transcription of polyomavirusDNA. EMBO J. 7:2515-2522.
24. Leff, S. E., R. M. Evans, and M. G. Rosenfeld. 1987. Splice commitment dictatesneuron-specific alternativeRNA process-ingincalcitonin/CGRPgeneexpression. Cell 48:517-524. 25. Legon, S., A. J. Flavell, A. Cowie, and R. Kamen. 1979.
Amplification in the leader sequence of late polyoma virus mRNAs. Cell 16:373-388.
26. Levitt, N., D. Briggs, A. Gil, and N. J. Proudfoot. 1989. Definition ofan efficient synthetic poly(A) site. Genes Dev. 3:1019-1025.
27. Logan, J., E.Falck-Pedersen,J. E. Darnell, and T. Shenk. 1987. Apoly(A) addition siteand a downstream termination region are required for efficient cessation oftranscription by RNA poly-merase II inthe mouse Pmaj-globin gene. Proc. Natl. Acad. Sci. USA84:8306-8310.
27a.Luo, Y., and G. Carmichael. 1991. Splice site choice in a complextranscription unitcontaining multipleinefficient
poly-adenylation signals. Mol. Cell. Biol. 11:5291-5300.
28. Mather, E. L., K. J. Nelson, J. Haimovich, and R. P. Perry. 1984.Mode ofregulationofimmunoglobulinmuand delta chain expressionvariesduring B-lymphocytematuration. Cell 36:329-338.
29. Niwa, M., S. D. Rose, and S.M.Berget. 1990. In vitro polyade-nylation is stimulated by the presence ofanupstream intron. GenesDev.4:1552-1559.
30. Peterson, M. L., E. R. Gimmi, and R. P. Perry. 1991. The developmentallyregulated shift frommembranetosecreted ,um RNA production is accompanied by an increase in cleavage-polyadenylation efficiencybutnomeasurablechangeinsplicing efficiency. Mol. Cell.Biol. 11:2324-2327.
31. Peterson, M. L., and R. P. Perry. 1986.Regulation ofproduction ofALm and >L mRNArequires linkage ofthe poly(A) addition sites and isdependentonthelength ofthe
.Us.Um
intron. Proc. Nati. Acad. Sci. USA83:8883-8887.32. Robberson, B. L., G. J. Cote, and S. M. Berget. 1990. Exon definition may facilitate splice site selection in RNAs with multiple exons. Mol. Cell. Biol. 10:84-94.
33. Rogers, J., N. Fasel,and R. Wall.1986. A novel RNA in which the 5' end is generated by cleavage at the poly(A) site of immunoglobulin heavy-chain secreted mRNA.Mol. Cell. Biol. 6:4749-4752.
33a.Roome, A., and G. Carmichael.Unpublisheddata.
34. Saiki,R. K., S. Scharf, F. Faloona, K. B. Mullis, G. T.Horn, H. A. Erlich, and N. Arnheim.1985.Enzymaticamplificationof P-globin genomic sequences and restriction site analysis for diagnosisof sicklecellanemia. Science 230:1350-1354. 35. Tooze, J.,ed.1980. Molecularbiologyoftumorviruses, 2nd ed.
DNA tumor viruses. Cold Spring Harbor Laboratory, Cold SpringHarbor, N.Y.
36. Treisman, R. 1980. Characterization ofpolyoma late mRNA leader sequences by molecular cloning and DNA sequence analysis. NucleicAcids Res. 8:4867-4888.
37. Treisman, R., and R. Kamen.1981. Structure ofpolyomavirus late nuclear RNA. J. Mol. Biol. 148:273-301.
38. Tseng, R. W., and N. H. Acheson. 1986. Use ofa novel S1 nuclease RNA-mapping technique to measure efficiency of transcription termination on polyomavirus DNA. Mol. Cell. Biol. 6:1624-1632.
39. Tyndall, C., G. LaMantia, C. Thacker, and R. Kamen. 1981. A region ofthe polyoma virus genome between the replication origin andlateprotein codingsequences isrequired in cis for bothearlygeneexpression and viral DNAreplication. Nucleic AcidsRes. 9:6231-6250.
40. Whitelaw, E., and N. J. Proudfoot. 1986.a-Thalassaemia caused byapoly(A) site mutation revealsthattranscriptional termina-tion is linked to 3'endprocessing inthe human 4globingene. EMBO J. 5:2915-2922.