JOURNAL OFVIROLOGY, Jan. 1993, p. 142-149 0022-538X/93/010142-08$02.00/0
Copyright C 1993, AmericanSociety forMicrobiology
Differentiated Liver Cell
Specificity
of the
Second
Enhancer of Hepatitis
B
Virus
CHIOU-HWA YUHANDLING-PAI TING*
Graduate InstituteofMicrobiology andImmunology, National Yang-Ming Medical College, Shih-Pai, Taipei 11221, Taiwan, Republic of China
Received 31 August 1992/Accepted 13 October 1992
Hepatitis B virus is a hepatotropic virus. Its replication and gene expression are mainly restricted to hepatocytes in theinfective process.The viral geneexpression thusprovidesa uniquesystem with whichto study the control of tissue-specific gene expression. We have previously reported the identification and characterization of the second enhancer (enhancer II) of hepatitis B virus. In this report, we further demonstrate that the minimal functional constituents of the second enhancer,boxaandbox
13,
displaylivercell and differentiation state specificity.Moreover, boxaexhibits thesameliver cellanddifferentiation state
specificitywhenfunctioningas anupstreamregulatorforthe basalcorepromoter.Gel shiftexperimentsreveal
auniqueboxa-binding protein, proteina,which is presentonlyindifferentiated livercells,where enhancer II isfunctional. Theconverse istrueforanother box a-binding protein, protein f, which is presentonly in
poorlydifferentiated liver cells andnonlivercells. Thesimplest hypothesis thatexplains these results is that protein a activates and/or protein fsuppresses the enhancer and upstream regulator functions. Although
C/EBP isacandidate foratranscription factorthatinteractswith boxaorbox
13,
noneofthebindingfactors identifiedinthegelshiftassays,including proteinaandprotein f,islikelytobeC/EBP becausetheydiffer from C/EBPin heatlabilityandsequencepreference.Eukaryoticgeneexpressionis inlargepartregulatedatthe transcription level. Such regulation is governed by the constellation of trans-acting cellular factors that bind to specific cis-acting elements and act in either a positive or
negative manner (25). Cell-type-specific gene activation, a
primarydeterminant of cellulardifferentiation, representsa more complex type of interplay, as both constitutive and
tissue-restricted trans-actingregulatoryfactorsareinvolved
(24). Thedifferential sequence-specific recognition ofthese cis-acting elements in promoters and/or enhancers bytheir cognatefactors provides amechanistic basis for the tissue-and differentiation-specific regulation of gene expression. Thestudy ofmodel viralgeneswhichdisplaydistinct tissue
tropismcanprovidevaluableinsightinto theintricate effects of cell-type-specific transcription regulation on
differentia-tion.
HepatitisBvirus(HBV)isahepatotropicvirus.Although HBV DNAhas been found innonhepatictissues ininfected patientsand intransgenicmice,HBVreplicationand matu-ration and, as a result, the pathologicalprocess aremainly
restricted to hepatocytes (2, 5, 11, 12, 35). There are four promoters in HBV: two surface promoters, one core pro-moter, and one X promoter. These promoters produce
messagesthatencode structuralcomponentsofthe virusand
thepolymerasethat isessentialforviralgenomereplication
(4, 15, 23, 28, 29, 39, 41, 44). Moreover, the pregenomic RNAproduced by thecorepromoterserves as atemplatefor
replication via reverse transcription (31, 33). So far, two
enhancershavebeenfoundintheHBVgenome. Enhancer I
is locatedbetween the open reading frames of the surface
and Xgenesandpartially overlapstheXpromoter(15, 32). EnhancerII iswithinthe Xopenreadingframe (40, 42).The
latter has a unique bipartite structure: two minimal func-tional constituents, a 23-bp sequence called box a and a
*Correspondingauthor.
12-bpsequencecalledbox 1B,arebothrequiredforenhancer II function (43). Interestingly, either boxat or box ,Bin an
upstream position can regulate the activity of the nearby
corepromoter (41).
Using DNA transfection to bypass viral entryinto cells,
we and others have demonstrated that the expression of
HBVgenesexhibits liver cell and differentiationstate
spec-ificity, recapitulating what we observed in the infective
processin vivo(6, 8, 17, 30,34, 36, 39).Ourpreviousstudies show thatonlyinthe humanhepatomacell linesHepG2 and HuH-7, which have the feature ofwell-differentiated liver cells,does enhancer II have strongenhancing activityonthe
simian virus 40 (SV40) earlypromoter. In contrast, this is not seen in the poorly differentiated HA22T/VGH cells or
thenonliver HeLa cells(42).Theseresults alsoapplytothe upstreamregulatoryeffectonthebasalcorepromoter(BCP) (41).Thesedifferentiatedactions,therefore,maycontribute
at least in part to the observedhepatotropism ofHBV. In thisstudy, we furtherinvestigated the liver cell and
differ-entiationstate specificityof the minimal essentialelements, boxaand box 1,ofenhancerII. We also usedgelshiftand
footprinting assays to study the interactions of different trans-acting factors withthese sequence motifs. We found
thatHA22T/VGHand HeLacellsaremissingoneof thetwo
box a-binding factors, protein a, which is abundant in
HepG2 andHuH-7 cells. The firsttwo celllines, however, containanadditionalbindingprotein, proteinf.Thepossible involvement of thesefactors in theestablishment of differ-entiation specificity isdiscussed below.
MATERIALS ANDMETHODS
Plasmid constructions. The HBV sequence used in this study is of the adw subtype. The numbering ofthe HBV
sequence begins at the unique EcoRI site. All reporter plasmidsused in transfectionexperimentscontaina
head-to-tail trimeric tandem repeat, referred toas A3, of a237-bp 142
Vol.67,No. 1
on November 9, 2019 by guest
http://jvi.asm.org/
BclI-BamHI fragment from the SV40 polyadenylation signal. A3 is placed 5' of the promoter sequence of interest and has been shown to stoptranscription readthrough from spurious upstream initiation (42). Plasmid pBCP-CAT contains the
chloramphenicol acetyltransferase (CAT) reporter gene driven by the BCP, corresponding to nucleotides nt 1744 to 1851 of the HBV genome (41). Plasmids pa/BCP-CAT and
p,B/BCP-CAT were constructed by insertion of annealed oligonucleotide sequences corresponding to the box a(5'GA
TCCATCGATCAAGGTCTTACATAAGAGGACTCTT3'
and 5'AAGAGTCCTCT-TATGTAAGACCTTGATCGAT
G3'; underlined nucleotides indicate nt 1645 and 1669,
respectively, of the HBVsequence) and box1B(5'GCTACT
TCAAAGACTGTGjGTACCCATG3' and 5'GGTACCACA
GTCITTGAAGTAGCTGCA3'; underlined nucleotides in-dicate nt 1705 and 1721 ofHBV) oligonucleotides, respec-tively, in theposition immediately upstream of the BCP of pA3BCP-CAT. Plasmid pSVpCAT contains the CAT re-porter gene driven by the 188-bp SV40 early promoter. Plasmid
pSVpCAT/a1
was constructed by the insertion of box a followed by box 1 downstream of the CAT gene of pA3SVpCAT.Cell lines, transfections, and CAT assays. Three human hepatoma cell lines, the well-differentiated lines HuH-7 (26) and HepG2 (1) and the poorly differentiated line HA22T/ VGH(7),aswell as the human cervical carcinoma cell line HeLa, were cultured as previously described (41). Cells weretransfected withplasmids containingthe CAT gene by the calcium phosphate precipitation method. CAT assays were performedby the method of Gorman etal. (14). The amountsofprotein samplesin eachexperimentvariedbyno more than 5%. The CAT activity of each sample was normalized to the CAT activity of pSV2CAT, which was takenas100%.If CATactivitywashigh,the celllysatewas
serially dilutedbefore the CAT assay(41-43).
Preparation of nuclear extracts and heparin-agarose frac-tionation ofextracts. NuclearextractsfromHuH-7, HepG2, HA22T/VGH, and HeLacells wereprepared aspreviously
described (9). Extracts were stored in small aliquots at
-70°Cafterbeingquicklyfrozen underliquid nitrogen.The nuclear extract was fractionated at 4°C essentially as de-scribedbyLichtsteineretal.(21),withaslightmodification.
Briefly, 25 mg of crude nuclear extract in 10 ml of nuclear
dialysis buffer was loaded onto a 20-ml heparin-agarose
column (heparinUltrogel-A4R; 4to6 mgofheparinperml;
1852
IBF,Garenne,France). The proteins bound to heparin were collected by step elution with nuclear dialysis buffer contain-ing 0, 0.1, 0.2, 0.3, 0.4, 0.5, 0.6, or 1.0 M NaCl. For experiments in which nuclear extracts were subjected to a thermolability test, the 0.4 and 0.3 M NaCl fractions were heated(50°C for 5
min,
70°C for 5 min, or 90°C for 15min) and thencentrifuged at 12,000 xg for 10 minat 4°C. The supernatants were collected for the DNase Ifootprintingand gel shift analyses.DNase Ifootprinting and gel shift
analyses.
Theend-labeled probes were generated via fill-in with [a-32P]dATP (3,000 Ci/mmol; Amersham, Amersham, Buckinghamshire,En-gland) at either terminus of a 450-bp BamHI-HindIII
frag-mentcontaining the HBV sequence from nt 1402 to 1851. Plasmidscarrying this insert were cut with one enzyme, end labeled, cut with a second enzyme, and then subjected to agarose gel electrophoresis. The labeled fragments were eluted from the agarose gel with a Spin-X filtering unit
(Costar, Cambridge, Mass.).The DNase Ifootprintingassay was performed as previously described (9). The crude or fractionated nuclear extracts were incubated with labeled DNAfragment for 30 min at 4°C. After addition of MgCl2
and CaCl2, the proper amount of DNase I was added to digest for 90 s at room temperature. The reactions were
stopped by deproteination. The extracted DNA was ana-lyzedon4%
polyacrylamide-8
M ureagels.Annealed double-stranded oligonucleotides (200 ng)
cor-respondingtothe box aorbox ,B sequencewereendlabeled with 80 U ofpolynucleotidekinase(Toyobo, Osaka, Japan)
and 80 ,uCiof
[_y-32P]ATP
(5,000Ci/mmol; Amersham). The free[-y-32P]ATP
was removed from the labeledoligonucle-otides with an Ultrafree-C3 filter unit (UFC3 LGC NB;
Millipore, Bedford, Mass.). The gel shift assay was per-formedaspreviouslydescribed (43). Briefly, proteins (10to 20
p,g)
were incubated ina20-pl bindingbuffercontaining1 ,ug ofpoly(dI-dC).
poly(dI-dC) (Pharmacia,Inc.)
and 6 x 104 cpm of labeled DNA. The reaction mixtures were incubated at30°C for30 min, and protein-DNAcomplexes wereresolved on 4% polyacrylamidegels. Forcompetition experiments, 5-, 25-, 125-, and 625-fold molar excesses of unlabeled double-stranded oligonucleotide werepreincu-bated with nuclearextracts onice for 5 min before addition of probe. The oligonucleotides for double-stranded C/EBP
consensus sequenceswere
5'GATCGAACATATTGCGCA
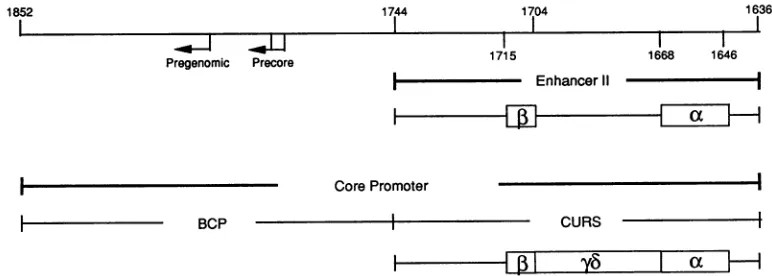
1744 1704 1636
Pregenomic Precore
I-BCP
1715 1668 1646I
- Enhancer11
-Core Promoter
CURS
[image:2.612.119.505.534.672.2]I
Li
IYz
I
az
FIG. 1. Locationsof enhancer II andcorepromoter.Aschematicrepresentationof thestructureof HBV fromnt1636to1852isshown. Thearrowsindicatethetranscriptionstartsitesforthe precoreandpregenomicRNAswithinthe BCP.EnhancerIIanditsminimal essential sequenceelements,boxa(nt1646to1668)and box P(nt1704to1715),aredemarcated atthe top. Thecorepromoter iscomposed of the BCP and itsupstreamregulatorysequence(CURS),anditsstructureis shownatthe bottom.Deletionanalysisrevealsthepresenceofatleast threefunctional constituents: box a, boxyb (nt1672to 1703),and box
P3.
Thefirsttwo arethemostpotent(41).on November 9, 2019 by guest
http://jvi.asm.org/
144 YUH AND TING
TABLE 1. Differentiated hepatoma cellspecificityof enhancer function onthe SV40earlypromoter
CAT activity (%)a
Cell line Induction(fold)'
pSVpCAT pSVpCAT/a,B
HepG2 1.60 72.0 45
HuH-7 1.72 25.8 15
HA22T/VGH 1.11 1.67 1.5
HeLa 1.90 2.28 1.2
aRelative CAT activity was calculated as a percentage of that of pSV2CAT. The valuesareaveragesof fourindependent experiments. pSVp-CAT contains the pSVp-CAT gene driven by the SV40earlypromoter.
pSVp-CAT/a1x has boxa followed bybox 1 downstream of the CATgene of pSVpCAT. Theoligonucleotides used for box a and box 13 aredescribedin
MaterialsandMethods.
bFold inductionwascalculated relative toCATactivity expressed from
constructslacking box aand box1 elements. The values are averagesof four independent experiments.
ATACAT'l'CCCAAGT3' and5'ACTTGGGAAATGTATT GCGCAATATGTTC3'.
RESULTS
Differentiated hepatoma cell
specificity
of enhanceractivity and upstreamactivator. Wehavepreviously(42) shownthat enhancer II (Fig. 1) displays differentiated liver cell speci-ficityinthat itisactivein such differentiated hepatoma cells as HepG2 and HuH-7 cells butnot inpoorly differentiatedhepatomacells suchas HA22T/VGHcells ornonliver cells such asHeLacells. Itwas subsequentlydemonstratedthat
enhancer II of HBV hasauniquebipartitestructure: a23-bp
sequence, box ao, and a 12-bp sequence, box
1,
are both sufficient and necessary for enhancer IIactivity.Neither box a norbox C alone stimulates theactivity of the SV40 early promoter,regardlessofwhether it is located in the upstream or downstream position. We were interested in whether these minimal functional constituents exhibit the same celllineage anddifferentiationstatespecificity. Toaddressthis,
boxaand box 1 werecloneddownstreamof the SV40early promoterandtested for their enhancer activities in four cell lines. It turnsoutthat although there isnegligiblestimulation in HA22T/VGH and HeLacells, there are 45- and 15-fold enhancements in HepG2 and HuH-7 cells, respectively
(Table 1). The results indicate that these minimal essential
[image:3.612.60.299.96.170.2]elements, i.e., boxa inconjunction with box
1,
retain the same differentiatedliver cellspecificityasdoes enhancer II in its entirety.TABLE 2. Differentiatedhepatoma cellspecificityof upstream activatingfunctiononthe BCP
Induction
Cell line CATactivity
(%)a
(fold)b
pBCP-CAT pa/BCP-CAT p,Q/BCP-CAT Box a Box13
HepG2 0.038 4.56 0.296 120 7.8
HuH-7 0.080 3.28 0.512 41 6.4
HA22T/VGH 0.150 0.615 2.40 4.1 16
HeLa 0.110 0.198 0.726 1.8 6.6
a Calculation of CATactivity is describedin Table 1,footnotea. pBCP-CAT contains the pBCP-CATgenedriven by the BCP. pa/BCP-CATand p,B/BCP-CAT havebox a andbox13,respectively, immediatelyupstream oftheBCP ofpBCP-CAT. The oligonucleotides used forbox aandbox13aredescribed
inMaterialsand Methods.
bCalculatedasdescribed in Table 1,footnoteb.
We havefound that either boxa orbox1, especiallybox aL, in the upstream position has stimulatory effects on the activity of the nearby BCP. We placed box ax or box 13 upstreamofthe BCP and assayed their stimulatory activities infour cell lines. Boxal activates the BCPstrongly inHepG2
and HuH-7 cells (120- and 41-fold, respectively) but acti-vatesit onlyslightly in the HA22TIVGH and HeLa cells (4.1-and1.8-fold, respectively). In contrast, box 13 has moderate activities in all cell lines tested (Table 2). This clearly
establishes that box a has differentiated hepatoma cell specificity.
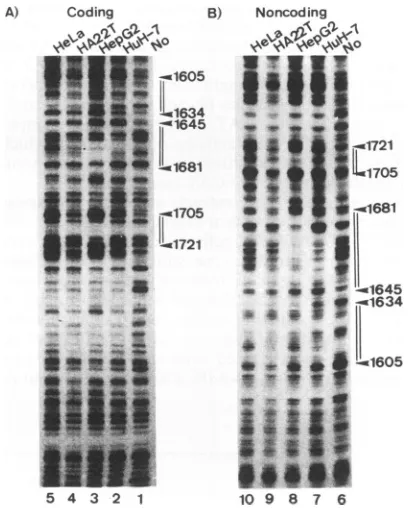
DNase Ifootprinting analysis of the box (K-box
13
region with crude nuclearextractsof four different cell lines. One of the simplest hypothesestoexplain the cell lineageand differen-tiation statespecificityof boxaand box13 is either that an essential trans-acting factor(s) that implements the activa-tionis presentin differentiated liver cells or that anegative factor(s) that silences the expression is present in poorly differentiated liver cellsor nonliver cells.The two possibil-itiesare,of course, notmutuallyexclusive.To addressthis hypothesis, DNase I footprinting analyses to detectbinding of box a and box 13 were performed. As shown in Fig. 2,protection of both box a and box 13 is seen with nuclear
A) Coding B) Noncoding
*5IIga-1605
li
641st
61634
-~1645
1< i ~~~~~~~-1721
U~1681
IL
1705It' II
6-w1705
* *51
1681
_
RI .
d Z
*
4i <
.1645f-l1634
'ji6!jii
5 4 3 2 1 109 8 7 6
FIG. 2. DNase I footprinting analysis of the core promoter region of HBV with crude nuclear extracts of HuH-7, HepG2, HA22T/VGH, and HeLacells. Thefootprintinganalysis was
per-formedby usingaBamHI-HindIII fragmentcontaining nt 1402to 1851 ofHBV.Either thecodingornoncoding strand was end labeled with the Klenow fragmentofEscherichia coli DNA polymerase. TheprotectedregionsofHepG2areindicatedbydouble lines onthe right,withnumbersreferringtonucleotidepositionswithintheHBV genome and arrowheads indicating the DNase I-hypersensitive sites. (A) Footprinting analysis of the coding strand. The
end-labeledprobewasincubated with noprotein(lane 1) or with 120 ,ug ofprotein from crude nuclear extracts of the cell lines indicated above lanes 2 to 5. (B) Footprinting analysis of the noncoding
strand.Theend-labeledprobe was incubated with noprotein (lane6)
orwith 120pLgofproteinfrom crude nuclear extracts of the cell lines indicatedabove lanes 7to10.
J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.337.541.322.576.2] [image:3.612.59.301.592.671.2]Huh-7 HepG2 22T HeLa
No 0.3 0.4 03 0.4 0.3 0.4 0.3 0.4 (M)
isI
J
1 2 3 4 5 6 7 8 9
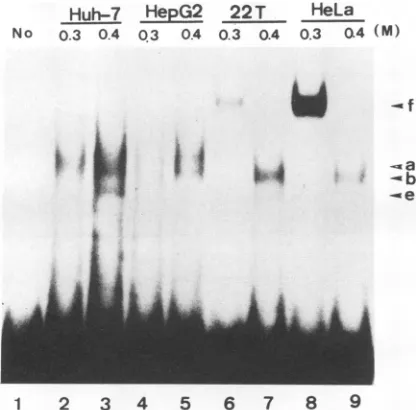
FIG. 3. Gel shift analysis of box a-bindingproteins in four cell lines.Double-strandedoligonucleotidescorrespondingtothe boxa
sequence were end labeled with polynucleotide kinase and
incu-bated with proteins of fractionated 0.3 and 0.4 M NaCl nuclear
extractsofHuH-7, HepG2,HA22T/VGH (22T), andHeLacellsat
30°Cfor30 min. Afterincubation,theDNA-protein complexeswere resolvedon anative 4%polyacrylamide gel.Lanes: 1,noprotein;2
to 9, 10 p.g of the indicated salt eluents of the heparin-agarose column from theindicatedcelllines.Fourshiftedbands,a,b,e,and f,areindicated.
extracts from all four cell lines. The observation that the patterns found in all cell lines are not completely identical suggeststhatthebinding proteins for either boxaorbox arelikelytobedifferent.
Analysisof the boxa-andbox
13-binding
proteinspresentinfourcell linesbygel shiftassay. In earlier studies,we have
demonstrated the existence of gel shifting activities from fractionated nuclear extracts ofHepG2 cells. Two shifted
bands,aandb,aredetected when the boxaoligonucleotide
isused astheprobe,whilebandscand dareformedwhen the box oligonucleotide is used. These DNA-binding proteins, being candidate transcriptionfactors that mediate
theenhancer function, displaydistinctsequencepreferences andheatlabilities (43). It isnotclear, however, whether the
same DNA-binding activities are present in these different cell lines. Toassess this, a gel shift assaywas performed.
The labeled box a and box oligonucleotides were
incu-bated with different fractions of nuclearextractseluted from theheparin-agarose column.As showninFig.3,whenboxa
isusedasthe probe, ashiftingcomplex that migratestothe
samepositionasband bispresentinthe 0.4 M NaCl fraction of all four cell lines. Three apparently different shifting activities are observed. A band shift identical toband a is presentonlyinthe0.4MNaCl fractionofbothHepG2and HuH-7 nuclear extracts. A shifting activity that produces bandfispresentinstead in the 0.3 M NaCl fractionof both theHA22T/VGH andHeLa nuclearextracts.Binding activ-ity f is muchmoreabundantintheHeLanuclearextractthan intheHA22T/VGHnuclearextract. Anothershifting activ-ity yielding band e ispresent in the0.4MNaCl fraction of the HuH-7 nuclearextract but not in that ofHepG2.These resultsare summarizedinTable 3.
Twobox shiftingbandsdesignatedcand daredetected by incubationwith the 0.3MNaClfractions of the nuclear extractsfromHuH-7, HepG2,andHA22T/VGHcells,while onlybandcis observed with the 0.3 MNaCl fractionfrom HeLa cells(Fig.4andTable4).Noother gelshiftactivityin
responsetoeither boxaorbox is detected fromanyother
fraction of nuclearextractsof thesefour cell lines.
Theobservation that shifting complexes of identical
mo-bilities aredetected with fractionated nuclearextractsfrom different cell lines suggests that theseDNA-bindingproteins
maybethesame.Tofurther addressthis,theheatsensitivity
and thesequencepreferenceof thesebindingproteinswere
examined. For heat sensitivity, the fractionated nuclear extracts were first treated ata giventemperature and then tested for their intactnessby thegel shiftassay. As for the
sequencepreference experiments, oligonucleotides of simi-lar butnotidenticalsequencesweretested for their abilities todisrupt theformation of thepreviouslyidentified shifting complexes. Inview of theweakhomology exhibited by box
a
(CAAGGTC`TlACATAAGAGGACTC`I`T,
fromnt1645to 1669) and box (CTACTTCAAAGACTGTG, fromnt 1705 to 1721) to the C/EBP consensus sequence (ATTGCGCA [image:4.612.75.283.78.283.2]AT) (abbreviated C/EBPpp) (3, 10, 13, 19), all three oligo-nucleotideswerechosen for thecompetition study.
TABLE 3. Boxa-bindingproteins of four celllines
NaClconcnand Relative Temp
sensitivity'
Sequence specificitycellline position
37°C
50°C
70°C
90WC0.4M
HuH-7 a ++ ++ - - Box a>> box - C/EBPpp
b ++ ++ - - Box a>>box
P
- C/EBPppe ++ ++ ++ ++ NDb
HepG2 a ++ ++ - - Box a>> box B - C/EBPpp
b +++ ++ - - ND
HA22TNVGH b ++ ++ - - Box a>> boxl - C/EBPpp
HeLa b ++ ++ - - Box a>> boxP- C/EBPpp
0.3 M
HA22TNVGH f ++ + - - Box a>boxP - C/EBPpp
HeLa f ++ + - - Boxa>box 3- C/EBPpp
aTemperaturesensitivitywasshownby the relative intensitiesof
shifting
bands.Thenumber ofplussignsindicates theabundance ofbindingproteins,whileaminussignindicates the absence ofbinding proteins.
bND,notdetermined.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:4.612.58.567.556.697.2]146 YUH AND TING
Huh-7 HepG2 22T HeLa
No 0.3 04 0.3 0.4 0.3 0.4 0.3 0.4 (M)
A A
Li
0.4M 0.4M 0.4NM 0,4M
Hu,-7 HepG2 22T HeLa
° 50 70 - 707 - 507090 - 50 70 90
i. -C
-ad
I
-b
_~~~~~~~~~~~~~"^ 11
1 2 3 4 5 6 7 8 9 10 11 12 13
0.3M 0.3M
-
0T
HeLa- 50r70 god - 50 70 90
1 2 3 4 5 6 7 8 9
FIG. 4. Gel shiftanalysisof box 3-binding proteinsinfour cell lines.Double-strandedoligonucleotides correspondingtothe box 3
sequencewerelabeledasprobes.Otherexpenmental detailsare as
described inthelegendtoFig.3.Lanes: 1,noprotein;2to9,10,ug of the indicated salt eluents of theheparin-agarosecolumn from the indicated cell lines. Twoshifted bands,cand d,areindicated.
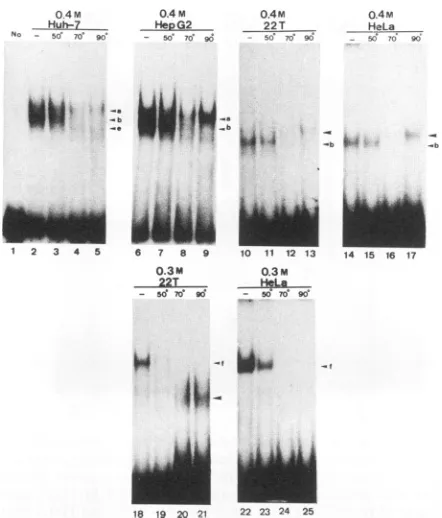
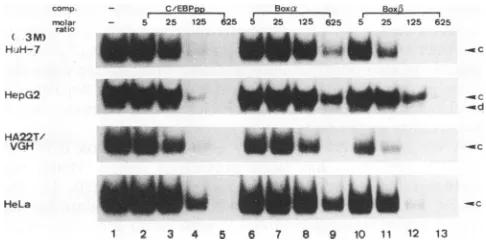
As shown in Fig. 5, the box a-binding proteins that are
presentinbandsaand baremoderately heat resistant, since
bothcanwithstand heattreatment at50 butnot at70°Cfor 5 min.A novel band thatmigratestothesamepositionasband aemergesonlyafter heattreatmentinall four cell lines. This band ismostlikelyaspuriousheat-resistantbinding protein. With respecttothesequence preferencefor the box a-bind-ing proteins, unlabeled box a, box 1, and C/EBPpp were
added at 5-, 25-, 125-, and 625-fold molar excesses as
competitors inthegelshiftassay. Complexb canbe
inhib-ited only bythe box asequence but is slightly affectedby
box,BandC/EBPppata625-foldmolarexcessintheHuH-7,
HA22TNVGH,
and HeLa nuclear extracts (Fig. 6). This is also thecase fortheHepG2 cells (datanotshown; a0.5 M NaCl fraction containing protein a only is used inFig. 6).Takentogether,thesamegelshiftposition,heatlability,and relative sequencepreference stronglysuggestthat the bind-ing proteins incomplexesb (referredto asproteins b) from
all four cell lines are identical. Similarly, proteins a in
FIG. 5. Heat sensitivities of boxa-bindingproteins in four cell lines.End-labeled boxawasincubated with 10 pg ofproteinof the indicated salt eluents from the indicated cell lines without heat
treatment (lanes 2,6, 10, 14, 18,and22)orwith heattreatmentat
50°Cfor5min (lanes 3, 7, 11, 15, 19,and23),70°Cfor 5min (lanes
4, 8, 12, 16, 20,and24),or90°Cfor 15min (lanes 5, 9, 13, 17, 21,and 25).Lane 1,control(no protein).Otherexperimental detailsareas described in thelegendtoFig. 3. Theshiftedbands,a,b,e,andf,
areindicated with arrowheads.Another shifted band thatappeared during 90°C treatmentis indicated with largerarrowheadswithout letters.
complexesafrom theHepG2andHuH-7cellsareprobably
identical and exhibit the same sequencepreferenceas
pro-teinb. Members of the thirdgroupof boxa-binding proteins, proteinsf incomplexes f,whicharepresentonlyinHA22T/ VGH and HeLa cell extract, areprobablyidenticalaswell because both aredestroyed whenheated at50°Cfor 5 min and exhibit the sequence preference box a > box C/EBPpp (Fig. 7).
As forthe box 1-binding proteinsin the shiftedcomplexes
candd, proteinscandproteindareboth heatlabile,asthey
[image:5.612.328.549.71.330.2]aredestroyed by heattreatment at50°Cfor 5 min (Fig. 8). TABLE 4. Box 3-bindingproteinsof four celllines
Celllinea Relative Temp
sensitivityb
Sequencespecificityposition 37°C 500C 70°C 900C
HuH-7 c ++ - - - C/EBPpp -box 1 > box a
d ++ + - - NDC
HepG2 c +++ - - - C/EBPpp -box c> box a
d ++ + - - C/EBPpp -boxc >boxa
HA22T/VGH c +++- - - C/EBPpp -box c > box a
d ++ + - - ND
HeLa c +++ + - - C/EBPpp -box c >box a
aData arefor the0.3 M
NaCl
fractionsof nuclear extracts of theindicatedcell
lines. bSee Table3, footnotea.cND,notdetermined.
J. VIROL.
,
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.75.291.76.288.2] [image:5.612.60.566.598.694.2]comp. - CEBPE, Box.k Box altao - 5 25 125 625 5 25 125 625 5 25 125 625
(0.4M)
HuH-7 a
HepG2
*3
ibWw
0.3M Huh-7
No - 50 70' 90
o.3M 0.3M oQ3M
HeG92 22T _HeLa
- 50 70 g0 - 50 70 90 - 50 70' 90
i d -Wx
iI
j#4
HA22T/-
HG2T_ i i * **ll~~~~~~~~~~-bb
Jb
HeLa _1" " -b
1 2 3 4 5 6 7 8 9 10 11 12 13
FIG. 6. Relative efficiencies of competition by box a, box1, and C/EBP consensus sequences for box a-binding proteins a and b. End-labeled boxawasincubated with 10pgof protein of the 0.4 M NaCl fraction ofHuH-7,HA22T/VGH, and HeLa cells as well as the 0.5MNaCl fraction of HepG2 cells. Lanes: 1, no competitor; 2 to5,unlabeled C/EBPpp; 6 to 9, box a; 10 to 13, box
P3.
The molar excessof eachcompetitor in the gel shift assay is indicated above each lane. Twoshifted bands, a and b, are indicated on the right.Moreover, both displaythe sequence preference C/EBP
-box1 >box a(Fig. 9).These results support theconclusion that the proteins c and d from the different cell lines are probably identical.
DISCUSSION
Specializedhighereukaryoticcellsdiffer from each other by virtue of the proteins that they make. Most of the control of geneexpressionthatunderlies thesedifferences operates at the level of transcription. The interaction of various
trans-acting factors with their cognate specific cis-acting elements is the major point at which such regulation is exerted. Theunique hepatotropismdisplayed by HBV in the infective cycle suggests a stringent requirement for the
liver-specific factors for viral gene expression. It is hoped that aninsightinto suchtranscriptionalcontrol in HBV as a model system will shed some light on how other
liver-specificgenes areregulated.
We have previously reportedthe identification and char-acterization of the second enhancer(enhancer II)of HBV. Enhancer II activates such homologous promoters as SPI, SPII,and XP and suchheterologouspromoters as the SV40
-12 3 45 6768 9
FIG. 8. Heatsensitivities of box 1-bindingproteins in four cell lines. End-labeled box13 was incubated with 10 ,ug of protein of the 0.3 M NaCl fractions from the indicated cell lines without heat treatment(lanes 2, 6, 10,and14) or with heat treatment at 50°C for 5 min(lanes 3, 7, 11, and15), 70°C for 5min(lanes 4, 8, 12, and 16), or 90°C for 15 min (lanes 5, 9, 13, and 17). Lane 1, control (no protein). Otherexperimental detailsare asdescribed inthelegendto Fig. 3. The shifted bands, c and d, are indicated witharrowheads.
early promoter in a position- and orientation-independent
manner(42).Furtheranalysisreveals thatenhancer II has a unique bipartite structure in that two interacting minimal essential elements,box aand box1, arebothsufficientand necessaryforthe enhancerfunction. Interestingly,box a or box D alone stimulates the nearby downstream BCP (41).
Box a stands out as a strong regulator, while box D has a moderate level ofactivity.
Bothenhancer II and the coreupstream regulatorexhibit
differentiatedlivercellspecificity.Whentested in
transient-transfection experiments, both are functional in well-differ-entiatedliver cellssuch asHepG2and HuH-7 cells but not in
poorly differentiatedlivercellssuch as
HA22T/VGH
cells ornonliver cells such as HeLa cells. In this study, we have
demonstrated that box a in conjunctionwith box ,B retains notonlythe enhancer IIfunctionbut alsowell-differentiated
liver cell specificity. Furthermore, box a, as an upstream
regulatorbyitself, alsodisplays well-differentiated livercell
specificity. Box
13,
in contrast, has moderate upstream regulator activity in both liver and nonliver cells.Theobservation that the two minimal essentialelements of the second enhancer, box aandbox
1,
displaydifferen-comp - C/EBP _ 8ooxa 80x,.
molar - 5 25 125 625 5 25 125 625 5 25 125 625
ratio
(3M)
H.FH-7 - W bi - c
comp. molar ratio (0-.3M)
HA22T/ VGH
- CrEBPpp- BoxrBox 1_,
- 25 125 625 25 125 625 25 125 625
.-,&
t-.4
4I
...
HeLa 2 w s
-1 2 3 4 5 6 7 8 9 10
FIG. 7. Relativeefficienciesofcompetition byboxa,box13,and C/EBP consensus sequences for box a-binding protein f. End-labeled box a was incubated with 10pgofthe0.3M NaClfractions ofHA22T/VGHand HeLa cells intheabsenceofcompetitor(lane 1) and in the presence of unlabeled C/EBPpp (lanes 2 to 4), box a (lanes5 to7),and box1 (lanes8to10)competitorsattheindicated molar excesses inagel shift assay.One shiftedband,f,is indicated ontheright.
HepG2
HA22T/
f ViGH
mw_6
1k
UOWNw
ad
HeLa -C i
1 2 3 4 5 6 7 8 9 10 11 12 13
FIG. 9. Relativeefficiencies ofcompetitionbyboxa, box
13,
and C/EBP consensus sequences for box 13-binding proteins c and d. End-labeled box 13 wasincubated with 10 ,ug of the 0.3 MNaCl fractions from the indicated cell lines intheabsenceofcompetitor (lane 1)and inthe presence ofunlabeledC/EBPpp(lanes2to5),box a (lanes 6 to 9), and box 1 (lanes 10 to 13) competitors at the indicated molarexcessesinagelshift assay.Twoshiftedbands,candd,areindicatedontheright.
...Idllpwoo
-a I I
I
A-
A..
.'k
-c
on November 9, 2019 by guest
http://jvi.asm.org/
[image:6.612.317.554.75.197.2] [image:6.612.56.295.76.228.2] [image:6.612.316.559.527.649.2] [image:6.612.58.296.574.651.2]148 YUH AND TING
tiated liver cell specificity is intriguing. The simplest model to explain the enhancer function in conjunction with its tissue specificity is that a certainactivator(s) is present and functional in thedifferentiated liver cells only to effectuate theenhancerfunction. Thisfactor(s) may beeithermissing
orrendered functionless becauseofmodification or antago-nismby another suppressor(s) in poorlydifferentiated liver cells or nonliver cells. This may well be the case for the upstream regulator function of box a, which exhibits the sametissue specificity as enhancer II. Footprinting and gel shiftanalyses were thus performed toexamine and compare thebox a- and box,3-bindingactivities in the fourdifferent celllines. The footprinting experiments reveal that protec-tion over this region is observed with the nuclear extracts fromall four cell lines, indicating that both sequencemotifs are bound with nuclear factors despite their different func-tionalstatuses. These nuclearfactors aremostlikely not all identical, as the footprint patterns are notallidentical. These differences are borne out in the gel shift analysis. First, protein c appears to be presentin all fourcelllines tested. Its ubiquitous presence makes it the most likely candidate for the transcription factor that mediates the upstream activa-tion by box 13 observed in all four cell lines. Second, box a-binding protein a is present in well-differentiatedliver cells such as HepG2 and HuH-7 cells but not in poorly differen-tiated liver cells such as HA22T/VGHcells ornonliver cells such as HeLa cells. The converse is true, however, for a different factor, protein f. This correlates well with the stimulatory activities of box a aseither the upstream acti-vatoror the enhancer in these four celllines.
Boxa not only canfunction as an upstreamregulator for the BCP but also can serve as an indispensable part of the second enhancer. The presence of dual functions in a mere 23-bpsequenceelement and the same celland differentiation state specificities exhibited by both suggests that the same factor(s) binds to this sequence motif andparticipates inboth
functions. In fact, detailed mutational analysis of box a reveals the same sequence requirement for both enhancer and upstream regulatorfunctions, furtherlending support to this notion (unpublished data). It is likely that protein a, whose presence parallels the functional status of both the upstream regulator and enhancer II, effectuates both stimu-lations in cooperation with other boxa- and box 1-binding
proteins. Of equal likelihood is thatprotein b is thepositive regulator that can lock the enhancer and the upstream regulator in a constantly "on" state except whenprotein f turnsoff suchactivation in poorlydifferentiated liver cellsor nonliver cells. These twosituations are not mutually exclu-sive. It is also possible that protein a and protein f are the sameprotein, albeit with different modifications.Anattempt tocharacterize these proteinsbiochemically in greaterdetail is currently under way.
A weak sequence homology of box a and box 13 to the
C/EBP consensus has been previously noted. Being the regulator for a variety of liver-specific genes (10, 13, 16, 18-20, 27, 38), C/EBP seems to be a good candidate for the transcription factor that interacts with either sequence motif (22). This is, however, not likely to be the case. First and foremost, C/EBP is present at a very low level in HepG2 cells (13), where we observe the greatest effect of both the enhancer and the upstream activator. Second, on the basis of sequence preference and heat sensitivity, proteins a, b, c, and d are distinct from C/EBP. In this study, we tested the fifthbinding protein, protein f, and showed that it cannot be
C/EBP either. However,we cannotcompletelyrule out the
possibility that any of these proteins is a member of the extendedC/EBP family (3, 37).
In conclusion, we have demonstrated that the minimal essentialelements ofenhancerII,boxao and box1,andbox aaloneas an upstreamregulator displaywell-differentiated liver cell specificity. Gel shifting experiments show that proteinaand proteinfarecandidatestomediate the stimu-latory or suppressive effects. Box a- and box 1-binding proteins appearnot tobe C/EBP. We hope thatthestudyof theinteraction between these factorsand theirbindingsites canhelp usunderstand how otherlineage-specific genesare controlled in different systems. Moreover, theperturbation of thisregulatory hierarchymay underliederangementsseen inpathologicalprocessessuchasoncogenic transformation.
ACKNOWLEDGMENTS
WethankShiuh-Wen Luoh foracritical review of andsuggestions forimprovingthemanuscript.
This research was supported by research grants NSC-82-0419-B010-022MB andNSC-81-0419-BO10-519 from the National Science Council of theRepublicofChina.
REFERENCES
1. Aden, D. P., A. Fogel, S. Plotkin, I. Damjanov, and B. B. Knowles. 1979. Controlled synthesis of HBsAgin a differenti-atedhuman liver carcinoma-derived cellline.Nature (London) 282:615-617.
2. Araki, K., J. Miyazaki, 0. Hino, N. Tomita, 0. Chisaka, K. Matsubara, and K. Yamamura. 1989. Expression and replica-tion ofhepatitis B virus genome intransgenicmice. Proc.Natl. Acad. Sci. USA 86:207-211.
3. Cao, Z., R. M. Umek, and S. L. McKnight. 1991. Regulated expressionof threeC/EBP isoformsduringadipose conversion of 3T3-L1 cells. Genes Dev. 5:1538-1552.
4. Cattaneo, R., H. Will, N. Hernandez, and H. Schaller. 1983. Signals regulating hepatitis B surface antigen transcription. Nature(London)305:336-338.
5. Cattaneo, R., H.Will,and H. Schaller. 1984. HepatitisBvirus transcriptionin the infectedliver. EMBOJ.3:2191-2196. 6. Chang, C., K. S.Jeng, C. Hu, S.J. Lo,T. S. Su,L. P.Ting,
C. K. Chou, S. H. Han, E.Pfaff, J. Salfeld, and H. Schaller. 1987. Production of hepatitis B virus in vitro by transient expression of clonedHBV DNA inahepatoma cell line. EMBO J. 6:675-680.
7. Chang,C., Y. Lin, T.-W. O-Lee, C.-K. Chou,T.-S. Lee,T.-J. Liu, F.-K.P'eng, T.-Y.Chen, and C.-P. Hu. 1983. Inductionof plasma protein secretion in a newly established human hepa-tomacell line. Mol. Cell. Biol. 3:1133-1137.
8. Chang, H. K., and L. P. Ting. 1989.Thesurface gene promoter ofthehumanhepatitisBvirusdisplays apreference for differ-entiatedhepatocytes.Virology170:176-183.
9. Chang, H.-K., B.-Y.Wang, C.-H. Yuh, C.-L. Wei, and L.-P. Ting. 1989. A liver-specific nuclear factor interacts with the promoter region of the large surface protein gene of human hepatitisBvirus. Mol. Cell. Biol. 9:5189-5197.
10. Christy, R. J., V. W.Yang, J. M. Ntambi, D. E.Geiman,W. H. Landschulz, A. D. Friedman, Y. Nakabeppu, T. J. Kelly, and M. D. Lane. 1989. Differentiation-induced gene expression in 3T3-L1preadipocytes: CCAAT/enhancerbinding protein inter-actswith and activates the promoters oftwoadipocyte-specific genes. Genes Dev. 3:1323-1335.
11. Elfassi,E., J. L.Romet-Lemonne, M. Essex, M. F.McLane,and W. Haseltine. 1984. Evidence of extrachromosomal forms of hepatitis B viral DNA in a bone marrow culture obtained from
a patient recently infected with hepatitis Bvirus. Proc. Natl. Acad. Sci. USA81:3526-3528.
12. Farza,H., M. Hadchouel, J.Scotto, P. Tiollais, C. Babinet, and C. Pourcel. 1988.Replicationand geneexpression of hepatitisB virus in a transgenic mouse that contains the complete viral
genome.J. Virol. 62:4144-4152.
13. Friedman, A. D., W. H.Landschulz,and S. L.McKnight. 1989. J. VIROL.
on November 9, 2019 by guest
http://jvi.asm.org/
CCAAT/enhancer binding protein activates the promoter of serum albumin gene in cultured hepatoma cells. Genes Dev. 3:1314-1322.
14. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982. Recombinant genomes which express chloramphenicol actyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 15. Guo, W., K. D.Bell, and J.-H. Ou. 1991. Characterization of the
hepatitis B virus EnhI enhancer and X promoter complex. J. Virol.65:6686-6692.
16. Herrera, R., H. S. Ro, G. S. Robinson, K G. Xanthopoulos, and B. M. Spiegelman. 1989. A direct role for C/EBP and the AP-I-binding site in gene expression linked to adipocyte differ-entiation. Mol. Cell. Biol.9:5331-5339.
17. Honigwachs, J.,0.Faktor,R. Dikstein, Y. Shaul, and0. Laub. 1989. Liver-specific expression of hepatitis B virus is deter-minedby the combined action of thecore gene promoter and the enhancer.J. Virol. 63:919-924.
18. Kaestner, K H., R. J. Christy, and M. D. Lane. 1990. Mouse insulin-responsiveglucosetransporter gene:characterization of thegene and transactivation by the CCAAT/enhancer binding protein. Proc. Natl. Acad. Sci. USA 87:251-255.
19. Landschulz,W.H., P. F. Johnson, E. Y. Adashi, B. J.Graves, andS. L. McKnight. 1988. Isolation of arecombinantcopyof thegeneencodingC/EBP. Genes Dev. 2:786-800.
20. Li, X., J. H. Huang, H. Y. Rienhoff, Jr., and W. S.-L. Liao. 1990. TwoadjacentC/EBP-bindingsequencesthat participatein the cell-specific expression of the mouse serum amyloid A3 gene.Mol. Cell. Biol. 10:6624-6631.
21. Lichtsteiner,S.,J.Wurin,and U.Schibler. 1987.The interplay ofDNA-binding proteinsonthepromoterofthe mousealbumin gene. Cell 51:963-973.
22. L6pez-Cabrera, M., J. Letovsky, K Q. Hu, and A. Siddiqui. 1990.Multipleliver-specific factors bindtothehepatitisBvirus core/pregenomic promoter: trans-activation and repression by CCAAT/enhancerbinding protein. Proc. Natl. Acad. Sci. USA 87:5069-5073.
23. Malpiece, Y., M. L.Michel, G. Carioni, M. Revel, P.Tiollais, andJ. Weissenbach. 1983. ThegeneSpromoterofhepatitisB virusconfersconstitutivegeneexpression. Nucleic AcidsRes. 11:4645-4654.
24. Maniatis, T., S.Goodbourn,andJ. A. Fischer. 1987.Regulation of inducible and tissue-specific gene expression. Science 236: 1237-1245.
25. Mitchell, P. J., and R.Tjian. 1989. Transcription regulation in mammalian cells by sequence-specific DNAbinding proteins. Science245:371-378.
26. Nakabayashi, H., K.Taketa, M. Miyano, T.Yamane, and J. Sato. 1982.Growth ofhumanhepatoma cell lines with differen-tiated functions in chemically defined medium. Cancer Res. 42:3858-3863.
27. Poli, V., and R. Cortese. 1989. Interleukin 6induces a liver-specific nuclear protein that binds to the promoter of
acute-phasegenes.Proc. Natl.Acad. Sci. USA 86:8202-8206. 28. Rail,L.B.,D. N.Standring,0. Laub,and W.J.Rutter.1983.
Transcription of hepatitis B virus by RNA polymerase II. Mol. Cell. Biol. 3:1766-1773.
29. Raney, A. K., D. R.Mulch,and A.McLachlan. 1991.Complex regulation of transcription from the hepatitis B virus major surfaceantigenpromoterin humanhepatomacell lines. J.Virol. 65:4805-4811.
30. Roossinck, M. J., S. Jameel, S. H.Loukin,and A. Siddiqui.1986. Expressionof hepatitisBviralcore region inmammalian cells. Mol. Cell. Biol. 6:1393-1400.
31. Seeger, C., D. Ganem, and H. E. Varmus. 1986. Biochemical andgenetic evidencefor the hepatitisBvirusreplication strat-egy.Science 232:477-484.
32. Shaul, Y., W. J. Rutter, and 0. Laub. 1985. Ahumanhepatitis Bviral enhancerelement. EMBOJ. 4:427-430.
33. Summers,J., and W. S. Mason. 1982.Replication of thegenome of ahepatitis B-like virus byreverse transcriptionofanRNA intermediate. Cell 29:403-415.
34. Sureau,C., J.L.Romet-Lemonne, J.I. Mullins,and M. Essex. 1986. ProductionofhepatitisBvirus byadifferentiatedhuman hepatoma cellline after transfectionwithcloned circularHBV DNA.Cell 47:37-47.
35. Tiollais, P.,C. Pourcel, and A. Dejean. 1985. The hepatitisB virus. Nature(London) 317:489-495.
36. Tsurimoto, T., A. Fujiyama, and K. Matsubara. 1987. Stable expression and replication of hepatitis B virus genome in an
integratedstateinahumanhepatomacellline transfected with thecloned viralDNA. Proc.Natl. Acad. Sci. USA84:444 448. 37. Williams, S. C., C. A. Cantwell, and P. F. Johnson. 1991. A familyofC/EBP-relatedproteins capable of forming covalently linked leucine zipper dimersin vitro.GenesDev.5:1553-1567. 38. Wilson, D.R., T. S.-C.Juan,M. D.Wilde,G. H.Fey,andG.J. Darlington. 1990. A58-base-pair region of the human C3gene confers synergistic inducibility by interleukin-1 and interleu-kin-6.Mol. Cell. Biol. 10:6181-6191.
39. Yaginuma, D., Y.Shirakata,M.Kobayashi, and K. Koike. 1987. HepatitisBvirus(HBV)particlesareproducedinacellculture systemby transientexpression of transfectedHBV DNA. Proc. Natl. Acad. Sci. USA84:2678-2682.
40. Yee,J.K. 1989. Aliverspecific enhancerinthecorepromoter regionof human hepatitisB virus. Science 246:658-661. 41. Yuh,C.-H.,Y.-L.Chang,andL.-P.Ting.1992.Transcriptional
regulation of precore and pregenomic RNAs of hepatitis B virus.J.Virol. 66:4073-4084.
42. Yuh,C.-H.,and L.-P.Ting.1990. The genomeofthehepatitisB virus containsasecondenhancer:cooperation oftwoelements within this enhancer is required for its function. J. Virol. 64:4281-4287.
43. Yuh,C.-H.,andL.-P.Ting.1991. C/EBP-likeproteins binding
to thefunctional box-a and box-, ofthe second enhancerof hepatitisBvirus. Mol.Cell Biol. 11:5044-5052.
44. Zhou, D. X., and T. S. Yen. 1991. The hepatitis B virus S promotercomprisesaCCAATmotif andtwoinitiationregions. J. Biol. Chem.266:23416-23421.