The
Duck Hepatitis
B
Virus Pre-C Region
Encodes
a
Signal
Sequence Which
Is
Essential for Synthesis
and
Secretion
of
Processed Core Proteins but
Not
for Virus Formation
HANSJ. SCHLICHT,* JOCHEN SALFELD,t AND HEINZ SCHALLER
ZentrumfurMolekulare Biologie, University of Heidelberg, D-6900 Heidelberg, Federal Republic ofGermany Received 16 June 1987/Accepted 19 August 1987
Analysis of the serum of duck hepatitis B virus (DHBV)-infected ducks has revealed the presence of
C-terminally truncated viralcoreproteins(eantigens). These proteinsareglycosylated andthereforewerenot released from infectedcells by lysis but rather by active secretion, indicatingthat the DHBVcoreproteincan
besynthesized alternativelyas a cytoplasmicor asecretory protein. Transient expression of cloned wild-type DHBVDNA and ofaspecifically designed viralmutantinahuman hepatoma cell line (Hep-G2)showed that
theDHBVcore genepromoteris active in differentiated human liver cells and that synthesisand secretionof theprocessedcoreproteinsaredependent onthe expressionof the pre-C region,asmallopenreading frame
which precedes the core gene. In addition, these experiments showed that the mechanism ofcore protein
processingandsecretion isconserved between DHBV and the human hepatitis B virus and therefore might be important forthehepatitis B viruslife cyclein general. In spite of this, intrahepatic injection of the pre-C
mutantintouninfectedducks resulted in viremia without concomitant e-antigen synthesis, indicating that virus formation isindependent of pre-C expression.
Thehepatitis B virusesareenvelopedDNAviruses which
predominantly infect the liver (reviewed in reference 37). We have chosen duckhepatitis B virus (DHBV), which infects Pekinducks,to studythestrategies of hepatitisB virusgene
expression. Compared with the human hepatitis B virus (HBV), which only infects humans and chimpanzees, this animal model virus offers the advantage that systematic in vivo studiesonviralmutantscanbe performed. Inaddition,
since DHBV is the member of the hepatitis B virus family
mostdistantly relatedto HBV, acomparative study of both
viruses should give an insight into the principles ofgene
expression of all hepatitis B viruses.
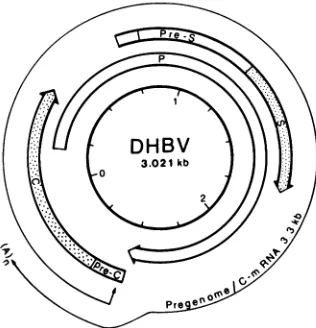
Sequence analysis (19, 33) showedthat the DHBVgenome comprises three open readingframes (Fig. 1). The pre-S/S
open reading frame encodes the viral surface proteins (27, 31). It completely overlapswith the P open reading frame, which is believedtoencode the viral polymerase (38). The C
open readingframe encodes the viralcore protein(33). The
strategy by which the core gene is expressed is ofspecial interest,since thisgeneappearstoencodemultipleproteins,
as is suggested by the fact that more than one core gene
product can be detected in the liver and serum of HBV-infectedpersons.
One of these proteins, HBcAg, has a strong tendencyto
aggregate to viral core particles (8, 22, 34)and is
predomi-nantly present in the liver. The other, HBeAg, has been characterized as aC-terminallytruncated core protein (36). Itdoesnotformaggregatesand is encounteredespeciallyin the serum. Recently, it was suggestedfrom in vitro studies thatsynthesisandsecretion of theHBeAg mightbe linkedto
theexpressionof thepre-C region (21, 24, 28, 29, 40),asmall openreadingframeprecedingthecoregenein allhepatitisB
viruses sequencedtodate. Inaddition, itwasspeculatedthat thepre-C sequence mightbenecessarynot onlyforHBeAg
*Correspondingauthor.
tPresent address: Dana-Farber CancerInstitute,Harvard
Medi-calSchool, Boston, MA 02115.
secretion but also for virus formation by targeting nascent cores to the membrane of the endoplasmic reticulum (24, 40),wherebuddingofnewly synthesizedvirusesis believed
tooccur (15).
Here we show that C-terminally truncated core proteins
similartothe HBeAgcanbe detected in theseraof
DHBV-infected ducks. Since these proteins areglycosylated, they were not released from lysed cells but ratherwere actively
secreted. We also present evidence that the biosynthetic pathway which leads to the synthesis of these proteins is conserved between HBV and DHBV, suggesting that the productionofprocessedcore proteins confersanadvantage to the hepatitis B viruses. In addition, we show that the
formation and the secretion of the truncated core gene
products are dependent on a functional pre-C region,
whereasvirusformation is not.
MATERIALSANDMETHODS
Animals. Pekin ducks (1-day-old ducklings) were
pur-chased from commercial suppliers and tested for DHBV
infection by dot-blot analysis as described previously (32).
Animalsweresacrificedattheageof 3weeks, and liverand serum sampleswere storedat -70°C.
Immunoprecipitationand Western blotanalysisofcoregene
products. For Western blot (immunoblot) analysis, 150 to
500 ,ul ofserum, upto 10 ml of tissue culture supernatant, 700 ,ul of cell lysate (see below), or 100 mg of liver tissue
homogenized in 1 ml of TNE (150 mM NaCl, 20 mM Tris
hydrochloride, 10 mM EDTA, pH 8.0) were subjected to
immunoprecipitation with 10 ,ul of anti-core immune serum
preadsorbed to 25 ,lI of protein A-Sepharose (Pharmacia Fine Chemicals) preswollen in phosphate-buffered saline
(PBS; 140 mM NaCl, 10 mM phosphate, pH 7.4). The sampleswereincubatedat 4°C with slight agitation for4 to 16 h. Unbound material wasremovedby three washeswith TNE, and the specificallybound proteinswere dissolved in 50 pul of sample buffer (3% sodium dodecyl sulfate, 2% 2-mercaptoethanol, 10%sucrose,0.1%bromophenolblue,5
3701
Copyright C) 1987,American Society for Microbiology
on November 10, 2019 by guest
http://jvi.asm.org/
3702 SCHLICHT ET AL.
FIG. 1. Schematic representation of the DHBV genome. The
arrows representthe three openreading frames asdefinedby start
and stopcodons. The depicted RNA ofmore than genome length
encodes the viral core protein and also serves as a pregenome for
viral replication.
mM EDTA, 200 mM Tris hydrochloride, pH 8.8). When samples were assayed directly, 10,ul of serum or 10
RI
ofliver homogenate was mixed with 40
RI
of sample buffer.After the samples were heated for 3 min in aboiling water
bath, they were applied to a 12.5% polyacrylamide gel.
Polyacrylamide gel electrophoresis and Western blotting
wereperformedessentially asdescribed previously (17, 39).
Aftertransfer, unoccupied binding sites were saturated by
incubatingthe filter foratleast 6 hinPBS-1% bovine serum
albumin (Miles Laboratories, Inc.). Core gene-encoded
pro-teins were detected by incubation of the filter with the
anti-core serum at a serum dilution of 1:2,000 in PBS-1%
bovine serumalbumin-0.01%sodium azide for 16 h atroom temperature. Excess antibody wasremoved by washing the
filter with three changes of PBS-0.1% Nonidet P-40 (total washing time, 30 min). Bound antibody was detected by
incubatingthe filter with '25I-labeled protein A (0.05
,uCi/ml;
Amersham Corp.) in PBS-1% bovine serumalbumin for3 h
at room temperature. Unbound protein A was removed by
washing with PBS-0.1% Nonidet P-40 as described above.
The air dried filter was exposed to X-ray film (XAR-5; Eastman Kodak Co.) at -70°C for 3 to 16 h with an
intensifying screen.
Endo F digestion. The serum-specific core proteins were
immunoprecipitated from duck serum(150 pdofserum, 350
[lI
ofPBS)asdescribedabove, washed, and solubilized in 50[lI
ofKNP buffer, pH 7.0 (5.2 ml of 0.5 M KH2PO4, 16.2 mlof 0.25 M Na2HPO4, 78.6 ml of H20) with 2% sodium
dodecyl sulfate and 2%2-mercaptoethanol by heating for 3
min in a boiling water bath. Then 700
RI
of KNP with 1% Triton X-100, 1% 2-mercaptoethanol, and 20 mM EDTA were added, the Sepharose was pelleted, and thesuperna-tant was transferred toa fresh tube. Fordigestion 0.5 U of
endo-p-N-acetylglucosaminidase
F (endo F; Boehringer, Mannheim GmbH, Federal Republic of Germany) wasadded, and the samples were incubated for 24 h at
37°C.
After digestion, sodium dodecylsulfate wasaddedtoafinal
concentration of 2% (75 of20% sodium dodecyl sulfate), andtheproteins wereprecipitated with trichloroacetic acid
(final concentration, 10%; 30 min on ice). The protein
precipitatewas pelletedand, aftercareful removal of
resid-ual acid,was dissolvedin 50
[lI
ofsample buffer.Generation of theC terminus-specific peptide antiserum. A peptidecomprisingthe last 13aminoacidsofthe DHBVcore
gene (sequence, S-S-S-S-H-H-R-S-P-S-P-R-K) was
synthe-sized by using an automated peptide synthesizer. For cou-pling, 7 mg of tetanus toxoid and 2 mg of
peptide
weredissolved in 1 ml of phosphate buffer (0.1 M, pH 6.8); with stirring, 0.5 ml of 21 mM
glutaraldehyde
wasaddeddropwiseover a period of 1 h at room temperature. After storage overnight at
4°C,
the mixture wasdialyzed againstPBS. For generation of an antiserum, twoNew Zealand Whiterabbitswere immunized with the coupled peptide. The first dose contained 300 pLg of coupled peptide in 2 ml of Freund
complete adjuvant. Booster injections (200
,ug
of coupledpeptide in 2 mlof Freund incompleteadjuvant)weregiven 3,
5, and 6 weeksafter thefirstinjection. After thethird booster
injection, blood was withdrawn and tested for an antibody titer against the peptide by enzyme-linked immunosorbent
assay asdescribedpreviously (25). The titerofbothsera was about 1:8,000. ForWesternblotting, the sera were usedat a dilution of 1:500.
Site-directed mutagenesis. The pre-C stop mutant was generated by site-directed mutagenesis with a DHBV 16 full-length genome cloned into the BamHI site of the vector
M13mp8 (3). For site-directed mutagenesis, a single
mis-match 18-mer oligonucleotide comprising nucleotides 2523 through 2540 of the DHBV minus strand (sequence, G-C-A-A-G-A-A-T-C-T-T-A-A-A-C-A-A-T) was synthesized by us-ing an automated oligonucleotide synthesizer. The melting temperature of this oligonucleotide was determined to be
58°C
for the wild type and63°C
for the mutant in 3 M tetramethylammonium hydroxide. Mutagenesis was carried out by a standard protocol (43). A phage clone, which strongly hybridized to this oligonucleotide after stringent washing(61°C,
3 M tetramethylammonium hydroxide), was selected by screening of the recombinantphage plaqueswiththe 5'-labeled mutagenizing oligonucleotide. This phage clone was plaque purified, and the T-to-A exchange
(nucle-otide 2531) was confirmed by sequence analysis.Preparation of DHBV circles for in vivo and in vitro transfection. A DHBV 16 full-length genome excised from plasmid 16t27 (32) was cloned into the
EcoRI
site of the vector pUC13. CsCl gradient-purified plasmid DNA (DHBV 16, wild type) or M13 replicative form (DHBV 16 pre-C mutant) were prepared by standard methods (see reference 43 for preparation of M13 replicative form). The DHBV genome was excised by cutting with the respective restric-tion enzyme and circularized by ligarestric-tion at a DNA concen-tration of 3 pmol/ml (4 U of T4 DNA ligase per ml, 16 h at15°C).
After ligation, proteins were removed by phenol extraction. The DNA was ethanol precipitated, dissolved in water, and dialyzed against 10 mM Tris hydrochloride-0.1 mM EDTA (pH 7.0).Transient expression of DHBV DNA in tissue culture. The human hepatoma cell line employed (Hep-G2) (1) was culti-vated in Dulbecco minimal essential medium (GIBCO Lab-oratories) containing 10% fetal calf serum (10 ml of medium per
25-cm2
flask). Cells were grown to 30% confluency in25-cm2
tissue culture flasks and transfected with 5 to 10,ug
of circularized DHBV DNA by using theCa3(P04)2
precipitate technique (13). After 4 to 6 h, a dimethyl sulfoxide shock was given (10% dimethyl sulfoxide in Dulbecco minimal essential medium-10% fetal calf serum, 20min
at37°C).
The dimethyl sulfoxide medium then was discarded, and 10 ml of fresh medium was added. After 48 h, the medium was removed, and the cells were lysed in 700,ul
of TNE-1% Nonidet P-40-0.5 mM phenylmethylsulfonyl fluoride. Insolublemate-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.107.265.76.240.2]rial was pelleted and discarded, and the clear lysate was
subjectedtoimmunoprecipitationand Western blotanalysis
as described above. Usually half of the DHBV proteins obtained from the cells or the medium of a 25-cm2 tissue culture flask was applied to a slot for gel electrophoresis and
subsequent Westernblotting.
Transfection ofducks with cloned DHBV DNA.
Four-day-oldDHBV-negativeducklings were transfected with 10,ug of
circularized DHBV DNA dissolved in 0.3 ml of PBS by
injecting the DNA solution into multiple sites of the liver.
After 3 weeks the animals were sacrificed, and liver and
serumsampleswere examined for DHBV DNA by dot-blot
analysiswith32P-labeled,cloned DHBV DNA. Usually50 to
80% oftheducks became DHBV positive after transfection.
Purification of virus by CsCl equilibrium centrifugation.
CsCl(971mg)wasdissolved in a mixture of 0.4 ml of serum and 3.2 ml of water. This solution was centrifuged in an
SW60rotor(Beckman Instruments, Inc.) for 19 h at 60,000 rpm at 20°C. Fractions of 200 ,ul were collected from the
bottom andassayed for DHBV DNA by dot-blot analysis.
ADH
BV-3E111411..
....I
SSO,
eozc,6
jaI
W 69k
_. 46
BDHBV-16
ll.~~~~~~~~~~~
l l l<
cD
,~'e~lOt
_DI
14
RESULTS
Western blot analysis of core gene products in serum and liver. To test whether proteins similar to the HBeAg are produced during DHBV infection, serum samples ofducks
endogenouslyinfected withDHBV were subjected to
immu-noprecipitation and Western blot analysis by using a rabbit
antiserum raisedagainst bacterially synthesized DHBVcore
antigen (DHBcAg; J. Salfeld, Ph.D. thesis, University of
Heidelberg, 1985). Two proteins with
molecuiar
sizes ofabout30 and 33kilodaltons(kDa) could be detected with this
antiserum only in DHBV-positive serum samples (Fig. 2,
lane 1). In somesera, two additional proteins with molecular
sizes of about 27 and 27.5 kDa were also present in small
quantities. Asis the case with HBV, theseproteins existin
a nonparticulate form, since the protein pattern remained
unchanged after preclearing ofthe serumby
ultracentrifuga-tion (data not shown). The soluble core proteins could be
easily detected in all serum samples from ducks infected
with wild-type DHBV (27 samples tested). In the liver,
multiplecoreproteins with molecular sizes ranging from32
to 46kDaweredetected(Fig.2,lane2).These proteinswere
located in particles, as could be shown by sedimentation
analysis (datanot shown). Adetailed analysis ofthe
struc-ture oftheliver-specificcore proteins will bethe subject of
aseparate report.
Serum Liver
6 kD
Amme~~~~~A
3i_
30
FIG. 2. Western blotanalysis ofcoregene-encoded proteins in
serumand liver.Samplesof 250,ulofserum(lane 1)and 1 ml of liver
homogenate (lane2)weresubjectedtoimmunoprecipitationwith the
anti-core serum as described in Materials and Methods, and the immunoprecipitates wereexamined forcoregene-encoded proteins
byWestern blotanalysis.
FIG. 3. Endo F digestion of the serum-specific core proteins. Serum samples from ducks (250 Ill) infected with either DHBV 3 or
DHBV 16 weresubjected toimmunoprecipitation with the anti-core
serum. The precipitated proteins were then analyzed by Western blotting either directly, after mock digestion, or after digestion with endoF. Inthe upper partthepotentialglycosylation sites within the
coregenes of therespective virus strains (amino acid positions4and 160 relative to the methionine encoded by the ATG at nucleotide 2647)areshown. MWM,Molecularweight marker.
The major core proteins in the serum are glycosylated.
Since the core
protein
ofDHBV 3, avirus strain which isendemic in Germany(33), contains two potential glycosyl-ation sites (Fig. 3A), we tested whether the secretedcore geneproducts detected in serum were glycosylated, asisthe casewith many secretoryproteins.Forthis purpose,
immu-noprecipitates obtained with the anti-core serum from
DHBV-positive sera weresubjected to digestionwith endo
F, an enzyme which cleaves both high-mannose and
com-plex-type, N-linked carbohydrate side chains (9). Afterendo
F treatment, twoproteins withmolecular sizes of about27
and 27.5 kDa could be detected (Fig. 3A, lane 3). Mock
digestion had no influence on the protein pattern (lane 1
versus lane 2). Thus, these proteins, which were present
onlyin smallamountsinuntreatedsera, appeartorepresent
theprimary(i.e., unglycosylated) secretory core gene
prod-ucts which become modified during
biosynthesis by
theattachment ofone or twocarbohydrate side chains.
To support this interpretation, we transfected
DHBV-negative ducklings with cloned DNA from DHBV 16, an
Americanisolate(19, 20)in
which,
dueto apoint mutation,the internal glycosylation site is
missing (Fig. 3B),
andthereforethe virusshouldproduce onlyamonoglycosylated
coregeneproduct.The results obtained withserafrom these
animalswith respecttothe secretedcoreproteinsareshown
in
Fig.
3B. Asexpected,
the 33-kDaprotein
wasmissing.
The coregeneproductswith molecular sizes of about 30 and 27 kDa werefound in about
equal
amounts,indicating
thattheN-terminal
glycosylation
site isnotusedasefficiently
astheinternalsite. After endo Ftreatment,
only
protein
bands atabout 27 kDa could beobserved(lane 3),
ashad been the case with the serumsamples
obtained from DHBV 3-infected ducks.Takentogether,these resultsdemonstrate that the 33- and 30-kDacore
proteins
representdoubly
andsingly
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.335.522.74.279.2] [image:3.612.127.228.551.669.2]3704 SCHLICHT ET AL.
~:1
2
3
T4
5
6
I'
F- C ,uG t'T1 (ofV~c-r
'-4.
__~~~~e
FIG. 4. The secreted core proteins are C-terminally truncated. Samples of1mlof liverhomogenateor250,ul ofserumofaDHBV-3 infected duck were subjected to immunoprecipitation (IP) with antisera raised against either the whole core protein (aC) or a C-terminalpeptide (aCT). Theimmunoprecipitates werethen ana-lyzedbyWesternblotting (WB) with eithertheanti-coreserumor theanti-C terminusserum.
atedforms ofa secreted 27-kDacore
protein,
respectively.In this paper the two glycosylated core gene products are termed gp33c and gp3Oc, whereas the unglycosylated, se-cretedforms arereferred to asp27c. By
analogy,
themajornonsecreted core proteins detected in aparticulate formin the liveraredesignated asp30c.
The major core gene products in serum are C-terminally
truncated. Since it has been shown for HBV that the secreted core gene product, the HBeAg, represents a
C-terminally truncated core protein, we raised an antiserum
againstachemically synthesized
peptide
comprising
thelast13 C-terminal amino acids of the DHBV core
protein
andexamined whether the same was true for the respective
DHBVproteins. This C
terminus-specific
antiserumprecip-itatedthemajorcoreproteinspresentin liver(Fig. 4,lane5)
and also recognized these proteins after Western blotting
(lanes 5 and 6). However, no reaction with the
serum-specificcore proteinscould beobserved(lanes 1, 3,and 4).
These dataindicate thatgp33c,gp3Oc,andp27c all represent
C-terminally
truncated core proteins. Thus, in analogy toHBV, these proteins will be referred to as DHBeAg. In
addition, thefact that undissociated cores could be
immu-noprecipitated fromliverhomogenatewith the anti-C
termi-nus serum shows that the C terminus ofthecapsid protein
must belocated at theoutside ofthecoreparticle.
Afunctional pre-C region is essential for thesynthesisand secretion of the processed core proteins. Since gp33c and
gp3Oc were sensitive to digestion with endo F, these two
proteins
have
been glycosylated during biosynthesis. It iswellestablishedthat allproteinswhich becomeglycosylated
mustbe translocatedthrough themembraneof the
endoplas-mic reticulum, a process mediated by a signal sequence
which,in most cases,is located at the very N terminus of an
exportedprotein (30).
In the case of all hepatitis B virus core proteins, a
candidate for such a signal sequence is the hydrophobic
peptide which is encoded by the pre-C region. To test
whether expression of the pre-C region was essential for
DHBeAg secretion, we introduced a stop codon into this
region by site-directed mutagenesis with a cloned DHBV 16
genome (Fig. 5A). Since sequence elements important for viralreplicationarelocated in the pre-C region, even a single
base change within this region might interfere with the
B..
Celis
i
1 2 3
d4 A
.,
e.
1.
AIed
rum
4 5
k 1- N.I
+. Er, of") F
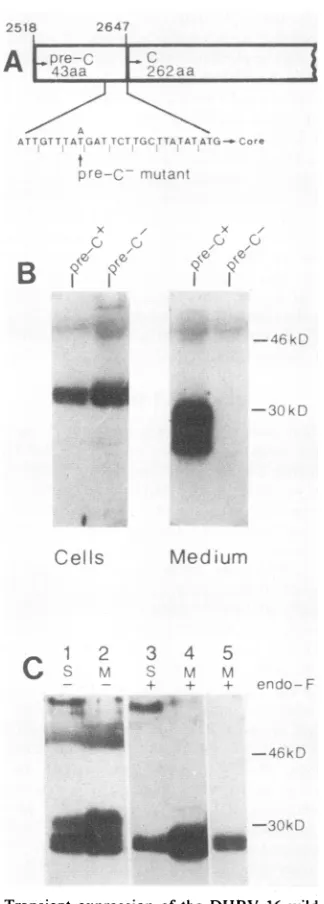
FIG. 5. Transient expression of the DHBV 16 wildtype and a
DHBV 16 pre-C mutant in a human cell line. (A) Schematic representation of the core gene 5' end. The eight pre-C codons preceding the core-ATG aregiven. The indicated pointmutation, whichresulted inan in-frame stopcodon within the pre-Cregion,
wasintroduced by site-directedmutagenesisofaDHBV16genome
cloned into M13mp8. (B) Comparison of the core gene products obtained after transient expression of the DHBV 16 wildtypeand the pre-C mutant in Hep-G2 cells. Cell lysates and tissue culture
supernatantswere subjectedtoimmunoprecipitation with the
anti-core serum asdescribed inMaterialsandMethods,andthe
respec-tive immunoprecipitates were analyzed by Western blotting. (C)
Western blotanalysisof the secreted coreproteinspresent in DHBV 16-positive duck serum(S), or in tissue culture medium(M) after transientexpression of DHBV 16wild-typeDNAinHep-G2cells. The samples weresubjected toimmunoprecipitationwith the anti-core serum, and the respective immunoprecipitateswereanalyzed by Westernblotting. Endo Fdigestion wasperformed where indi-cated. Lane 5 is a shorterexposure of lane 4.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.115.260.75.211.2] [image:4.612.362.524.88.540.2]DHBV PRE-C FUNCTION 3705
infectivity oftheclonedDNA. Therefore,wefirsttested the
influence ofthepre-C stop mutationon DHBeAg secretion
in tissue culture. For these experiments the human
hepa-toma cell line Hep-G2 was employed, a cell line which has
been shown recently tobe competent for HBV production
and HBeAg secretion (6, 35).
Theresult of thisexperiment is showninFig. SB. Whereas
the intracellular core proteins were qualitatively and
quan-titatively unchanged after transient expression ofthe
wild-typeand thepre-C mutant(lanes 1 and 2), secretionproved
to be completely dependent on a functional pre-C region,
since after transient expression of the pre-C mutant no
DHBeAg could be detected in the medium (lane 4 versus
lane 3). In neither case could the DHBeAg be detected
intracellularly. Thus, the pre-C region appears to encode a
signal peptide which is essential for the formation and
secretion of theprocessedcoreproteinsbut has noinfluence
on the synthesis oftheintracellular forms.
The high
efficiency
by which the DHBV C gene wasexpressed in the
Hep-G2
cells also enabled us to testwhether theprocessingstepswhich leadtothe synthesisand
secretion of modified core gene products are conserved
between human and duck cells. With the
exception
of aslightly
differentglycosylation,
theprocessed
core geneproducts which were secreted by this cell line
during
tran-sient
expression
of DHBV DNA(Fig.
5C, lane2)
areidentical with those which were detected in the serum of
infected ducks (lane 1). In
particular,
the C-terminalcleav-age appears to take place at the same sites within the core
protein, since after endo F
digestion
no difference betweenthep27c derived from duckserum
(lane 3)
and thep27c
fromthe tissue culture supernatant
(lanes
4 and5)
could beobserved.
A functional pre-C region is dispensable for virus forma-tion. To test the influence of the
pre-C
mutation on theinfectivity of the viral DNA, three
DHBV-negative
duck-lings were transfected with the mutated DHBV genome
by
intrahepatic
injection
of cloned DNA. After 3 weeks theanimals were sacrificed and tested for DHBV infection. As
shown
by
dot-blotanalysis
of liverhomogenate
and serum, twoofthe threeanimals had becomeDHBVpositive
atthistime (data not
shown).
Toprove that the DHBV DNAdetectedin the serumwas
located in viral
particles,
serumsamples
weresubjected
toCsCl
equilibrium
centrifugation,
and thegradient
fractionswere examined for DHBV DNA
by
dot-blotanalysis.
Thevirus-DNA
peak
wasdetected in thefractionscorresponding
to a
density
of 1.19 to 1.20g/cm3,
thedensity
of DHBVparticles (31;
Fig.
6A).Wethen examined
whether,
aftertransfection of thepre-C
mutant, viremia had
developed
in the absence ofDHBeAg
synthesis,
as should beexpected
from the tissue culturedata. Forthis purpose, serum and liver
samples
ofthe twoducks which became DHBV
positive
aftertransfection withthe
pre-C
mutant, as well ascontrolsamples
takenfromananimal transfected with
wild-type DNA,
were examined forcore gene
products
by
Westernblotting
(Fig. 6B).
Whereas the secretory coreproteins
could beeasily
detected in the serum of the control animal(duck
C),
noDHBeAg
was detectable in the sera of the two ducks which had beentransfected with the
pre-C
mutant(ducks
AandB),
indicat-ing thatthe infection was due to the mutant. No
DHBeAg
was visible even after a
10-fold-longer
exposure(data
notshown).
No differences could be observed withrespect
tothe
liver-specific
coreproteins
aftertransfection ofwild-type
andmutant DHBV DNA.
AFraction
Density
Duck A
Duck B
-.9
8
1i
, t t t? 3 1 :;:.
,
1 8g/cmi93
* v
l*
BuSerum
B Duck A B C A vBeCC
-4t.ikD
mm-FIG. 6. (A) Serum
samples
oftwoducks which becameDHBVpositive
aftertransfection with thepre-C
mutantweresUbjected
toCsCl
equilibrium
centrifugation. Samples
of thegradient
fractions then were tested for the presence of DHBV DNAby
dot-blotanalysis
with32P-labeled,
cloned DHBV DNA as aprobe.
Thedensities of the
gradient
fractions which contained thevirusDNAare
given.
(B) Examination of serumand liversamples
ofducks,eithertransfected with DHBV 16
wild-type
DNA(duck
C),orwith aDHBV 16mutantwhich carriedanin-framestopcodon withinthepre-C region
(ducks Aand B), for DHBV core geneproducts by
Westernblotting. Serum
samples
(150 ,ul)weresubjected
toimmu-noprecipitation
with the anti-coreserumbeforeanalysis.
The liversamples
(1mgof livertissue) wereassayed
directly.
To test whether the viral particles
produced
aftertrans-fection withthe
pre-C
mutantwereinfectious,
weinoculatedthree
DHBV-negative
ducklings
with serum obtainedfromthe two ducks which became DHBV
positive
aftertransfection withthe mutantDNA
(200
,ul
ofseruminjected
intrahepatically).
After3 weeks theanimals weresacrificed
and tested for the presence of DHBV DNA in liver and
serum
by
dot-blotanalysis
as well as for the presence ofeantigen
inthe serumby
Westernblotting.
Aswasevidencedby
the presenceofDHBV DNAinserumandliver,
all threeanimalshad becomeDHBV
positive
within
thisperiod,
butagain
noDHBeAg
could be detected in the serum(Fig.
7,
lanes
3, 4,
and5).
The amount ofviral DNA in liver and serum wascompared
betweensamples
taken from duckswhich became DHBV
positive
after transfection with thepre-C
mutant DNA(lanes
1 and2),
after infection withmutant virus
(lanes 3,
4,
and5),
and aftertransfection withwild-type
DNA(lanes
6 and7);
nosignificant
differencescould be
observed,
irrespective
ofthe fact thattransfectionor infectionwith themutantledtoviremiain the absence of
any
e-antigen
synthesis.
In this context it should bemen-tioned that
endogenously
DHBV-infected ducks or duckswhich were
experimentally
infected withserum takenfromendogenously
infected animals showed serum titers whichwere about 5to 10times
higher
thanthose shown inFig.
7,
whereas theamountsof viral DNA in the liverwere
compa-rable.
However,
since virus titers wereequally
low aftertransfectionwith the mutantorthe wild
type,
thereappearsto be no correlation between
e-antigen synthesis
and virustiter. That the virus titerwasalsolow afterinfectionwith the
VOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.324.545.71.284.2]3706 SCHLICHT ET AL.
A
[,z1
fc!c:to Tranrfst
M tjtinTFl Auitiu1t )Pv$
**,* * *
M
t
Mtci
t_
C-JS.)HP1\/- DN A
if,H:-'
DNDHV D tA;
r
n s rLJ m
DHF3BA. E S r
uru
rnFIG. 7. Detectionof DHBV-DNA in liverhomogenate and
se-rumby dot-blotanalysis (A) and of DHBeAg inserumbyWestern
blotting (B). Liver tissue (100 mg)washomogenizedin1mlofTNE
buffer, and 1 ,ul ofhomogenate wasspotted onto anitrocellulose filter. Serum samples (1 ,ul) were spotted directly onto the filter. ViralDNA wasdetected by hybridization with32P-labeled, cloned DHBV DNA. For Western blot analysis, 250 ,ul of serum was
subjectedtoimmunoprecipitation with the anti-core serum, andthe
immunoprecipitates thus obtained were analyzed as described in Materials and Methods. Lanes: 1 and 2, samples from ducks transfected with thepre-CmutantDNA;3 to5, samples fromducks
infected withmutantvirusbyintrahepatic injection of 200 p.1serum
taken from the ducksAand B inFig. 6;6and 7,samples from ducks transfected with wild-type DNA.
During transient expression of DHBV DNA in the human
hepatomacell lineHep-G2,acell linewhich is competent for
HBV production, large amounts of DHBV core gene
prod-ucts(gp33C, gp3Oc, p27c, andp30c) were produced. Thus it
appears that the cellular factors which regulate C-gene
expression act on the DHBV genome as on the HBV genome and that the species specificity of the hepatitis B viruses might not be due tospecies-specific regulatory proteins. We are currently testing this possibility by examining whether infectious DHBV particles are producedby the Hep-G2 cells after transfection with DHBV DNA.
The application of this system to the analysis of the expression of the DHBV core gene showed a variety of
interesting results.
(i) The DHBV core gene products can pass through two differentbiosynthetic pathways. In addition tothe structural coreproteins, p3oC, whichweredetected inlarge quantities in theliver, considerableamountsofC-terminally truncated coreproteins weredetected in the serumofDHBV-infected ducksaswellasin the medium of cells transfected withviral DNA. Inanalogy to the HBeAg, a similar protein known to be present in the serum of HBV-infectedindividuals, these proteinswere termedDHBeAg.
We have shown here that the bulk of the DHBVeantigens are glycosylated (the HBV eantigen contains no glycosyl-ation site). This is animportant finding, since it proves that thepresence of theseproteinsin the serum is not due to cell lysis but ratherto active secretion. Thus, as demonstrated here for the duckvirus, hepatitis B viruscoregeneproducts can enter twodistinctly different biosynthetic pathways (Fig. 8). One product, p30C, is synthesized in the cytoplasm and aggregates to form coreparticles, whicharethenexportedas acomponent of the virusparticles. In fact this protein might also be modified, asis indicatedby the pronounced hetero-geneity of the different core gene products which were detected in the infected liver. Anotherproduct,thepre-C/C
mutantvirusmaybedue to the low virus titeroftheserum
that was used for the infection. The question of whether
thereare anydifferences between the mutant andthe
wild-type virus with respect to pathogenicity is currently the
subject ofa systematic study.
Insummary, the data presented above prove that
expres-sion ofthepre-Cregionis notnecessaryforthegeneration of
infectious virus.Furthermore, the data show that the
inabil-ity to synthesize e antigens can be transmitted as a stable
trait which ischaracteristic for the generated DHBV mutant
andthat thereisnostrongselectivepressure withrespect to areversion to the wild type.
DISCUSSION
Inthework described here we present a system which for
thefirsttime allowed thecombined analysis of a hepatitis B
virusmutant in tissue cultureand in vivo. This was possible
because of the unexpected finding that the DHBV C-gene promoter is activein thehuman hepatoma cell line Hep-G2. In several reports it has been described that the HBV core gene is only poorly expressed in most cell lines after
transfection with viral DNA, whereas the surface proteins
are usually expressed at a high level (2, 5, 7). Thus it has
been proposed that transcription of the C gene is strictly
regulated and that the respective regulatory elements may
be, atleast in part, responsible for the species specificity of
thehepatitis B viruses.
I
C-GeneDGlycosylation
Core asse y 30C
Core assernbly
I
L ~ Ip27C Export
I :z:::::JIg9p30C
f ?
I I9~~P33c
Secretion
FIG. 8. Schematic representation of the two different biosyn-theticpathwayswhichare entered by thehepatitis B virus core gene productsas deducedfromthe experimental results presented in this report. The pre-C regionis shaded; the two different mRNAs are depicted as wavy lines with different thickness according to the different abundance of these mRNAs. Further details are given in
the text. ER, Endoplasmic reticulum.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.79.296.71.307.2] [image:6.612.337.548.445.658.2]protein, is translocated into the lumen of the endoplasmic reticulum, modified by glycosylation, truncated by C-terminal and, most likely, N-C-terminal proteolytic processing, and eventually secreted.
(ii) The pre-C region encodes a signal sequence which is essential for the synthesis and secretion of theprocessed core gene products. As mentioned above, aprocessed core pro-tein, the HBeAg, can be detected in serum during HBV infection. The process by which this protein reaches the serum remained unknown for a long time. Very recently, evidence was obtained indicating that partial deletion of the pre-C region abolished HBeAg secretion from mammalian cells (21, 24, 28, 29). However, many aspects, such as whether expression of thepre-C region was also essential for the synthesis of a correctly processed HBeAg, remained a matter of dispute.
In this study, we have constructed a pre-C mutant by introducing a stop codon into this region by a single base exchange. The expression of this mutant in tissue culture and in vivo showed that noDHBeAg was produced, either in a secreted form or in an intracellular form, when the pre-C region was interrupted by a stop codon. Consequently it appears that afunctional pre-C region is essential not only forthe secretion but also for the synthesis of the correctly processed e antigens. Since the synthesis of thecytoplasmic core proteins in tissueculture and in vivo was not affected by the pre-C mutation, most likely two different proteins are synthesized, a pre-C/C protein which is processed and secreted and a C protein which remains primarily intracel-lular and isonlyexported as acomponent of a virus particle. Thus, the signal sequence which is encoded by the pre-C region is only used optionally.
To date, only a few proteins have been described which are alternatively synthesized with or without a signal se-quence.Among these are the gag protein of Moloney murine leukemia virus (26), theplasminogenactivator-inhibitor (12), and the SUC2 gene product of Saccharomyces cerevisiae (4). In the last case it has been shown that the secretory and cytoplasmic proteins are encoded by the same structural gene but originate from two mRNAs with different 5' ends controlled by two differentially regulated promoters. It seems very likely that,inthe caseof the hepatitis B viruses, a similarmechanism leads to thedifferential synthesis of the pre-C/C and C proteins (Fig. 8). If so, the regulatory mech-anism which enables the synthesis of two different mRNAs from the core gene is conserved between human and duck liver cells, since expression of the DHBV core gene in the human hepatoma cell line Hep-G2 leads to theproduction of the same proteins that are synthesized in the infected duck
liver.
Major RNA species with heterogenous 5' ends which could be translated eitherinto the C- or thepre-C/C protein have beendescribed for the woodchuck and ground squirrel hepatitis B viruses (10, 23). However, no pre-C mRNAs have yet been detected for DHBV and HBV in RNA preparations obtained frominfected livertissue (3, 42). This failure is most likely due to the fact that, for unknown reasons, theabundance of this mRNA species is much lower in HBV- and DHBV-infected tissue than in the case of woodchuck and ground squirrel hepatitis B viruses. How-ever, since the pre-C mRNA is not packagedinto viral cores, as occurs with the C mRNA (11), even a low amount of pre-C mRNA could direct the synthesis of large amounts of e antigens.
(iii) The process of e-antigen formation appears to be conserved between the human and theduckhepatitis Bvirus.
Sofar, theprocessofe-antigen formation has been studied
only for HBV. This report describes for the first time the
structureand the biosynthesisof the processed core proteins
ofoneoftheanimalhepatitisBviruses and thus allowsus to deduce principles which underly the process of core protein secretion.
Ithas been shownforHBeAg that 34 or 36amino acidsare cleaved from the C terminus of HBcAg during e-antigen
biosynthesis (36). Despite the fact that DHBcAg is much larger than HBcAg (30.3 versus 21 kDa), the C-terminal
portion of DHBcAg whichismissinginDHBeAg appearsto
be of the same size, since the molecular size difference between DHBcAg (30.3 kDa)and unglycosylated DHBeAg
(27kDa)isabout3 to 4kDa. This calculation is based on the
assumption that most of the pre-C sequence has been
removed during DHBeAg biosynthesis, as is the case with most signal sequences (30).
When the DHBcAg amino acid sequence in the vicinity of
the putative cleavage site is inspected, a sequence motif is
found which is highly homologous to the known HBeAg
cleavage site (HBV, T-T-V-V-R-R-R; DHBV,
T-T-V-V-Y-G-R-R-R). This homology is especially striking since the
overallhomology between HBV and DHBV in this region is very low, i.e., less than 20% (33). Since a similar sequence is also present within the same C-gene region in woodchuck and ground squirrel hepatitis B viruses (H-T-V-I-R-R-R), it
is likely that this sequence motif represents a processing signal which is recognized in liver cells in a
species-independentmanner.Together with a conserved mechanism
for the generation of the C and the pre-C/C protein, this would explain why the DHBV core proteins, which are synthesized by the Hep-G2 cells, are identical with those that can be detected in infected ducks. However, the possi-bility that the cleavage might not take place at a single site is suggestedbythe presence of two primary secreted core gene products with slightly different molecular sizes (27 and 27.5 kDa, respectively).
Taken together, all of the dataindicate that themechanism which leads to the generation and the secretion of the truncated core proteins is conserved between HBV and DHBV, the most distantly related members of the hepatitis B virus family. This suggests that e-antigen formation con-fers an advantage to these viruses and may have played a role in their natural history.
(iv) Virus productionisindependent ofpre-C expression. In previous reports suggesting that the pre-C region was essen-tial for e-antigen secretion, it had also been speculated that the expression of thepre-C/C protein might beimportantfor the budding process by targeting nascent core particles to the endoplasmic reticulum membrane (24, 40).
We havechallenged this view by expressing aviralmutant with a defective pre-C region in vivo. Transfection of ducks with this mutant resulted in viremia without concomitant synthesis of DHBeAg, asensitive marker for pre-C expres-sion. Although we cannotrigorously exclude thatrevertants tothewild type were produced at a very low
frequency,
the fact that comparable virus titers were obtained after transfection with the mutant orthe wild-type DNAstrongly argues against afunctional role of the pre-C/C proteinin the process of virus formation. This view is also strongly sup-ported by the fact that the inability to synthesize eantigensis a stable trait, as could beshownby the transmission of the virus mutant.
The finding that production of infectious virus is not necessarily coupled with e-antigen synthesis is also of med-ical importance since, in HBV, the HBeAg is the most
on November 10, 2019 by guest
http://jvi.asm.org/
3708 SCHLICHT ET AL.
commonly used marker for determining the infectivity of
serumsamples (18, 41). InDHBV,outof27DHBV-positive
duck sera which were examined forDHBeAg by Western
blotting, none was DHBeAg negative. However, the fact
that this marker is not completely reliable has also been
suggestedforHBV,sincein afewcasesviral DNA hasbeen
detected in serawhich were negative forboth HBeAgand
anti-HBeAg (14, 16). It will be interestingto seewhetherat
leastsomeof thesecases aredue toaninfection withaHBV
pre-C mutant.
ACKNOWLEDGMENTS
We thankR.Pipkornforsynthesisof thepeptide,M.Nassaland
E. Lehmann for synthesis of the oligonucleotide, S. Selzer for providing Hep-G2 cells, P. Galle for help with the transfection experiments, C. Kuhn and E. Schlichtmann for artwork, and V. Boschforcritically readingthemanuscript.
This work was supported by the Deutsche Forschungsgemein-schaft (SFB 229), the Bundesministerium fur Forschung und
Technologie (BCT0381-5),and the Fond derChemischenIndustrie.
LITERATURE CITED
1. Aden,D.P.,A.Fogel,S.Plotkin,I.Damjanov,and B. Knowles.
1979.ControlledsynthesisofHBsAginadifferentiatedhuman
livercarcinoma-derived cell line.Nature(London)282:615-616.
2. Asselsberg, F. A. M., H. Will, P. Wingfield, and M. Hirschi.
1986.Arecombinant chinese hamsterovarycell linecontaining
a300-foldamplifiedtetramerof thehepatitisBgenometogether with a double selection marker expresses high levels of viral protein. J. Mol. Biol. 189:401-411.
3. Buscher, M., W. Reiser, H. Will,and H. Schaller. 1985.
Tran-scripts and the putative RNA pregenome of duck hepatitis-B
virus: implicationsforreversetranscription.Cell40:717-724. 4. Carlson, M.,and D.Botstein. 1982.Twodifferentiallyregulated
mRNAs with different5'ends encode secretedandintracellular formsofyeastinvertase. Cell28:145-154.
5. Cattaneo, R.,H.Will,G.Darai,E.Pfaff,and H.Schaller. 1983. Detection ofanelement of the SV40 late promoterin vectors used forexpressionstudies in COS cells. EMBOJ. 2:511-514.
6. Chang, C.,K.Jeng,C.Hu,S.Lo,T.Su,L.Ting, C.Chou,S.
Han,E. Pfaff,J. Salfeld, andH. Schaller. 1987. Production of
hepatitisBvirusinvitrobytransientexpressionof clonedHBV
DNA inahepatomacellline. EMBOJ.6:675-680.
7. Christman, J. K., M. Gerber, P. M. Price, C. Flordellis, J. Edelman, and G. Acs. 1982. Amplification of expression of hepatitis B surface antigen in 3T3 tells cotransfected with
dominant-actinggene andclonedviralDNA. Proc. Natl.Acad.
Sci. USA 79:1815-1819.
8. Cohen, B., and J. Richmond. 1982. Electron microscopy of hepatitisBcoreantigen synthesizedinE.coli. Nature(London) 296:677-688.
9. Elder, J. H., and S. Alexander. 1982. Endo-beta-N-acetylglu-cosaminidase F: endoglycosidasefromFlavobacterium
menin-gosepticumthat cleavesbothhigh-mannoseandcomplex glyco-proteins. Proc. Natl. Acad. Sci. USA 79:4540-4544.
10. Enders,G.H.,D.Ganem,and H. E. Varmus.1985.Mappingthe
major transcripts of ground squirrel hepatitis virus: the
pre-sumptivetemplateforreversetranscriptionisterminally
redun-dant. Cell42:297-302.
11. Enders, G. H., D. Ganem, and H. Varmus. 1987. 5'-Terminal
sequencesinfluencethesegregationofground squirrel hepatitis virus RNAs into polyribosomes and viral core particles. J. Virol. 61:35-41.
12. Genton, C.,E. Kruithof,and W. D. Schleuning. 1987. Phorbol
esterinducesthebiosynthesisofglycosylatedand
nonglycosyl-ated plasminogen activator inhibitor 2 in high excess over
urokinase-type plasminogen activator in human U-937
lym-phomacells. J. Cell Biol. 104:705-712.
13. Graham,F.L.,and A.J.vander Eb.1973. Anewtechnique for
the assay ofinfectivityofhumanadenovirus5DNA. Virology 52:456-467.
14. Harrison, T. J., V. Bal, E. G. Wheeler, T. J. Meacock,J. F. Harrison, and A. J. Zuckerman. 1985. HepatitisB virus DNA and e antigen in serum from blood donors in the United Kingdom positive for hepatitis B surface antigen. Br. Med. J. 290:663-664.
15. Kamimura, T., F. Yoshikawa, F. Ichida, and H. Sasaki. 1981. Electronmicroscopic studies of Dane particles in hepatocytes with special reference to intracellular development of Dane particles and their relation withHBcAg in serum. Hepatology
1:392-397.
16. Krogsgard, K., P. Kryger, J. Aldershville, P. Andersson, C. Brechot, and the Copenhagen hepatitis acute programme. 1985.
Hepatitis B virus DNA in serum from patients with acute
hepatitisB.Hepatology 5:10-13.
17. Laemmli, U. K.1970.Cleavage of structural proteins duringthe
assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
18. Liebermann, H. M., D. R. La Brecque, M. C. Kew, S. J. Hadziyannis, and D. A. Shafritz. 1983. Detection ofhepatitis B
virus DNAdirectly in humanserumbyasimplifiedmolecular
hybridization test: comparison to HBeAg/anti-HBe status in
HBsAg carriers. Hepatology 3:285-291.
19. Mandart, E., A. Kay, and F. Galibert. 1984. Nucleotide
se-quence ofacloned duckhepatitisB virusgenome: comparison withthe human and woodchuck hepatitis B virus sequences. J. Virol. 49:782-792.
20. Mason, W., C. Aldrich, J. Summers, and J. Taylor. 1982.
Asymmetric replicationof duckhepatitisBvirus in liver cells:free minus-strand DNA. Proc. Natl. Acad. Sci. USA 79:3997-4001. 21. McLachlan, A., D. R. Milich, A. K. Raney, M. G. Riggs, J. L.
Hughes, J. Sorge, and F. V. Chisari. 1987. Expression of
hepatitisB virus surface and coreantigens:influences of pre-S andprecore sequences. J. Virol. 61:683-692.
22. Miyanohara, A., T. Imamura, M. Araki, K. Sugawara, N.
Ohtomo, and K. Matsubara. 1986. Expression of hepatitis B
viruscoreantigengene inSaccharomyces cerevisiae: synthesis of twopolypeptides translated from different initiation codons. J. Virol. 59:176-180.
23. Moroy, T., J. Etiemble, C. Trepo, P. Tiollais, and M. A. Buendia. 1985. Transcription ofwoodchuck hepatitis virus in thechronically infected liver. EMBOJ. 4:1507-1514.
24. Ou, J.,0.Laub,and W.J.Rutter.1986.HepatitisBvirusgene
function: theprecoreregiontargetsthecoreantigentocellular membranes and causes the secretion of the e antigen. Proc.
Natl. Acad. Sci. USA83:1578-1582.
25. Pfaff, E., M. Mussgay, H. 0. Bohm, G. E. Schulz, and H. Schaller. 1982. Antibodiesagainstapreselected peptide
recog-nize and neutralize foot and mouth disease virus. EMBOJ. 1:869-874.
26. Pillemer,E.A., D. A. Kooistra,0.N.Witte, andI.L. Weissman. 1986. Monoclonalantibodytotheamino-terminalL sequence of
murine leukemia virus glycosylated gag polyproteins demon-stratestheirunusualorientation inthecellmembrane.J. Virol.
57:413-421.
27. Pugh, J. C., J.Sninsky, J.W.Summers,and E.Schaeffer. 1987. Characterization of a pre-S polypeptide on the surfaces of
infectious avianhepadnavirus particles.J.Virol.61:1384-1390.
28. Roossinck, M., S. Jameel, S. Loukin, and A. Siddiqui. 1986.
Expression of hepatitisBviralcoreregioninmammaliancells. Mol. Cell. Biol. 6:1393-1400.
29. Roossinck, M., and A. Siddiqui. 1987. In vivo phosphorylation and protein analysis of hepatitisBviruscoreantigen. J. Virol. 61:955-961.
30. Sabatini, D., G. Kreibich, T. Morimoto, and M. Adesnik. 1982. Mechanismsfortheincorporation ofproteins in membranes and
organelles.J. Cell. Biol. 92:1-22.
31. Schlicht, H. J., C. Kuhn, B. Guhr, R. J. Mattaliano, and H. Schaller. 1987.Biochemicalandimmunologicalcharacterization
ofthe duck hepatitis B virus envelope proteins. J. Virol. 61:
2280-2285.
32. Sprengel, R., C. Kuhn, C. Manso, and H. Will. 1984. Cloned duck hepatitis B virus DNA is infectious in Pekin ducks. J. Virol. 52:932-937.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
33. Sprengel, R., C. Kuhn, H. Will, and H. Schaller. 1985. Compar-ative sequence analysis of duck and human hepatitis B virus
genomes.J. Med. Virol. 15:323-333.
34. Stahl, S., P. Mackay, M. Agazin, S. Bruce,andK. Murray.1982. Hepatitis B viruscoreantigen: synthesis in E. coli and applica-tion indiagnosis. Proc. Natl. Acad. Sci. USA 79:1606-1610. 35. Sureau,C., J. Roup-Lemonne, J. Mullins, and M. Essex. 1986.
Production of hepatitis B virus by a differentiated human hepatoma cellline after transfection with cloned circular HBV DNA. Cell47:37-47.
36. Takahashi, K., A. Michada, G. Funatsu, M. Nomura, S. Usuda, S.Aoyagi, K.Tachibana, H. Miyamoto, M. Imai, T. Nakamura, Q. Miyakawa, and M. Mayumi. 1983.Immunochemicalstructure
ofhepatitis B e-antigen in theserum.J.Immunol.130:2903-2908. 37. Tiollais, P., C. Pourcel, and A. Dejean. 1985. The hepatitis B
virus. Nature (London) 317:489-495.
38. Toh,H., H. Hayashida, and T. Miyata. 1983. Sequence homol-ogybetweenretroviralreverse transcriptase and putative
poly-merases of hepatitis B virus and cauliflower mosaic virus.
Nature(London) 305:827-829.
39. Towbin, H., T. Staehelin, and J. Gordon. 1979.Electrophoretic transfer of proteins frompolyacrylamide gelstonitrocellulose sheets:procedure andsomeapplications. Proc. Natl. Acad. Sci. USA76:4350-4354.
40. Uy, A., V. Bruss, W. Gerlich, H. Kochel, and R. Thomssen. 1986. Precoresequencesofhepatitis B virus inducingeantigen and membrane association of the viral core protein. Virology 155:89-96.
41. Weller, I. V. D., M. J. F. Fowler, J. Monjardino, and H. C. Thomas.1982. Thedetection of HBV DNA inserumby molec-ularhybridization: amoresensitive method for the detection of complete HBV particles. J. Med. Virol. 9:273-280.
42. Will H., W. Reiser, T. Weimer, E. Pfaff, M. Buscher, R. Sprengel, R. Cattaneo, and H. Schaller. 1987. Replication strat-egyof humanhepatitis B virus. J. Virol. 61:904-911.
43. Zoller, M. J., and M. Smith. 1984. Oligonucleotide-directed
mutagenesis: asimple method using twooligonucleotide prim-ersand asingle-stranded DNA template.DNA3:479-488.