JOURNAL OFVIROLOGY, Mar. 1990, p. 1217-1226 0022-538X/90/031217-10$02.00/0
Copyright©D1990, American Society forMicrobiology
Identification of Phorbol
Ester Response
Elements in
the Promoter
of
Epstein-Barr Virus
Putative Lytic Switch Gene BZLF1
ERIK FLEMINGTON AND SAMUEL H. SPECK*
Division of Tumor Virology, Dana-Farber Cancer Instituite, and Department ofPathology, Harvard
University
MedicalSchool,
44Binney
Street, Boston,
Massachusetts 02115Received 9October1989/Accepted 1 December 1989
Theproductofthe Epstein-BarrvirusBZLF1 geneencodes aprotein which is relatedtoc-fos,it has been showntobind specifically toaconsensus AP-1 site, and its expression in latently Epstein-Barrvirus-infected lymphocytes is sufficient to trigger the viral lytic cycle. We identified several elements within the BZLF1 promoter (Zp) which are responsive to the phorbol ester 12-O-tetradecanoylphorbol-13-acetate (TPA), an
inducerof the viral lytic cycle. These elements fallintotwoclassesbased onthefactors which bindtothese sequencesandtheirresulting functionalbehavior. Fourofthe elementsarehomologous (ZI elements)and share
homology to a protein-binding domain in the promoter region ofthe coordinately expressed BRLF1 gene. When cloned upstream of heterologous promoters, the ZI elements function as silencers which exhibit TPA-inducibleenhanceractivity. Adistinct TPA-responsive element(ZII)is locatedneartheTATA box and shareshomologywith theAP-l-binding sitein the c-junpromoter.Asynthetic oligonucleotide withasequence
corresponding tothe ZII element effectivelycompetes for binding of nuclear factors to the c-jun AP-1 site. Furthermore, wefound thatacomplex of c-jun and c-fos boundtotheZIIdomain.
Epstein-Barr virus (EBV) is a lymphotropic human
her-pesvirus which latently infects B lymphocytes, resulting in concomitant growth transformation of the infected cells. Duringvirallatency, onlyalimited number of viralgenes are
expressed, while an estimated 100 or more viral genes associated with the lytic life cycle are quiescent (5, 12). Indeed, only four viral promoters have been shown to be transcriptionally active during latency, and allareclustered nearthelatent origin of replication (oriP)(seereference 48). The sixvirally encoded nuclear antigens (EBNAs)presentin latently infected lymphocytesare encoded byexonsspread
over the left-hand 110 kilobases (kb) of the viral genome.
However, theirtranscription is drivenfromoneoftwoviral promoterslocatedattheleft-hand end of thegenome(10, 44,
46). The transcripts that encode the EBNAs all share com-mon 5' exons derived from the majorinternal repeat (IR1) region andarealternatively splicedtothevarious 3' coding domains (4, 8, 9, 40, 41, 44, 46, 47). Two otherviral genes
expressedduring latency encode membrane proteins andare underthecontrol of distinctpromoters (20, 30).
InsomeEBV-infectedcelllines, afraction of thecellscan be induced to enter the viral lytic cycle by treatment with various reagents, including phorbol esters, butyrate, iono-phore, andanti-immunoglobulin (6, 28, 34, 51, 55). Alterna-tively, thelyticcyclecanbe inducedby superinfectionwith the appropriate strain of EBV (42). The P3HR-1 strain of EBVis a particularly effective inducer oflatently infected cell lines. This strain of EBV has been shown to contain heterogeneous rearranged viralgenomes whichare respon-siblefor its enhanced ability to induce the lytic cycle (38). Heterogeneous rearranged viral DNA has beencloned, and a fragment of this defective genome containing the viral BamHI Z fragment rearranged to a portion of the viral BamHI W fragment can disrupt latency when transfected intolatentlyinfectedlymphocytes (14).Furtherstudies have demonstrated that expression of the BZLF1 genefrom this
*Correspondingauthor.
rearranged fragment is responsible for triggering the viral lytic cycle (13).
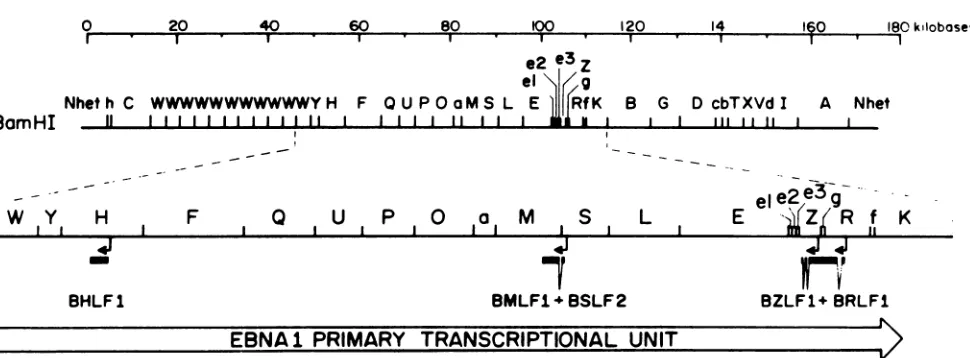
Shortly after induction of the viral lytic cycle, several regions of theviralgenomebecome transcriptionally active.
In additiontotheBZLF1gene,expression oftwoother viral
genes,BMLF1 and BRLF1,has beenidentified(Fig. 1) (11,
26, 33, 50, 54). All of these viral genes map within the
primary transcription unit ofone of the EBNA genes
ex-pressed during latency, EBNA1, and are transcribed from
the opposite strand (Fig. 1). Each of these viral genes
encodes a protein capable of trans activating other viral promoters(11, 26, 29, 32, 33, 50, 53). However,when these genes are transfected into latently infected lymphocytes under the control ofa strong heterologous promoter, only the product of the BZLF1 gene is able to trigger the viral lytic cycle (11). Furthermore, it has been shown that in the
presence ofaprotein synthesis inhibitor, cycloheximideor
anisomycin, only transcriptionof BZLF1 and BRLF1canbe induced (7, 49). The accumulated evidenceargues strongly
that the initial event in the lytic cascade is activation of BZLF1 transcription.
SeveraltranscriptscontainingBZLF1have been identified duringinduction of thelytic cycle,oneinitiatingfromZpand
twoothers initiatingfrom thepromoterfor the BRLF1gene
(7, 36). All three of these transcripts contain the complete BZLF1gene,which iscomposedof threeexons.TheBZLF1
protein (whichis also referred to asZEBRA [25] and EB1
[11]) is 245 amino acids long, and two regions of it are
partially homologous toc-fos (residues36to110 and 153 to 196) (19). Furthermore, it has been shown that the BZLF1 protein exhibits specific DNA-binding properties and can
recognize a site witha consensus AP-1 sequence (19).
Aregion of Zpfrom -226 to -118 has previously been reported tobe 12-O-tetradecanoylphorbol-13-acetate (TPA) responsivein HeLacells(53). Hereweshowthat thisregion ofZpisresponsivetoTPA in Blymphocytes. Moreover,we
havemappedthe bindingof cellular transcriptionfactorsto this region and also show that afragment ofZp containing thisregionfunctionsas asilencerwhich exhibitsanactivator
1217
Vol.64, No.3
on November 10, 2019 by guest
http://jvi.asm.org/
1218 FLEMINGTON AND SPECK
0 20 40 60 80
Nhet hC WWWWWWWWWWWY H F QUPOaMSL
II lIl1IlI II 1111f IIII111I1
H
IF
0
Q
IU
IP IO
100 120 14 160 180 kiloboses
I I*r -
-e2 e3 z el
E
(RfK
B G D cbTXVd I A NhetI I I III 11t I I
aM
S
L
E
el
e2g
nnn...11
f
K
BMLFI+BSLF2 BZLF1+BRLFI
EBNA
PRIMARY
TRANSCRIPTIONAL
UNIT
FIG. 1. Schematicexon mapof several viralgenesinvolvedin induction of thelytic cycleshown withrespecttothe BamHI endonuclease
mapof theB95.8strain of EBV.Thegenomelocationandtranscriptionalorientation of severalpromotersfor EBVearlygenesareindicated
(arrows) along with the correspondingexonstructureof thesegenes. Also shown is theprimary transcriptionalunit of theviralgenethat
encodesnuclearantigenEBNA-1. The lettersonthe EBVmaprefertoindividual BamHI restriction endonucleasefragments.
function inthepresenceofTPA when cloned upstream ofa
heterologous promoter. In addition, we identify specific
elements within this region of Zp and also fromthe down-stream region of Zp which areresponsive toTPA. Four of theseelements (ZI elements)arehomologousandmayform related protein-DNA complexes. These elements are
char-acterized by a conserved A+T-rich core sequence
juxta-posedtoaregion of somewhat variable homology.A distinct TPA-responsive element(ZII) is foundmoreproximaltothe
capsite. Thiselement containsanAP-1-like site which may
be functionally relatedtothe c-junAP-1 site. MATERIALS ANDMETHODS
Cell culture, transfections, and CAT assays. Cells were grownat37°C in RPMI 1640 medium (GIBCO Laboratories) containing10%newborn calfserumaspreviously described (47). Ramos cells (an EBV-negative Burkitt's lymphoma cell line)weretransfected by using DEAE-dextranaspreviously
described (37) but with the following modifications. The cells
were spundownat1,000 x g,washedoncewithRPMI 1640
mediumwithoutserum,and suspendedataconcentration of 3.3 x 107/mlinRPMI 1640withoutserum.Cells(2x 107;0.6 ml) were added to sterile tubes containing 4 ,ug of the relevant plasmid DNA, and DEAE-dextran was added to this to a final concentration of 200 ,g/ml (0.4 ml of 0.5 mg/ml). Cellswereincubated for 30minat37°C, followed by splittingofeach transfection intotwocultures containing 10 ml of RPMI 1640 with 10% newborn calf serum with or
without20ngofTPAperml.Exceptasnotedotherwise,the culture mediumposttransfection also contained 1.25 mg of
theophylline perml.
Cellswereharvestedat36 h posttransfection, spundown
at 1,000 x g, washed oncewith phosphate-buffered saline, and suspended in0.1 ml of0.25 MTris chloride (pH 7.5). The cell suspension was then lysed by three rounds of
freeze-thawing, and the debris was removed by
centrifuga-tion. The chloramphenicol acetyltransferase (CAT) activity of theextractwas determinedaspreviously described (23), and theextent of acetylation of chloramphenicolwas
quan-titatedby excision ofspotsfrom thin-layer chromatography plates, followed by liquid scintillation counting.
Plasmid constructions. The HindIII-BamHI fragment of
pSV2CAT containing the cat gene was cloned into the HindIII and XhoI sites of the Bluescript polylinker (BS-CAT). A BamHI-NaeI fragment containing the BZLF1 promoterwas then inserted upstream from thecatgene by
being cloned into the BamHI and HindIll sites of the Bluescript polylinker (Stratagene). The -221ZpCAT
con-structwas generated by digestionof thisplasmidwithSphI andBamHI,followedby bluntingwith T4 DNApolymerase and recircularization. Zpdeletionswere generated by using
exonuclease III and mung bean nuclease as previously described (27) andwere characterized by DNA sequencing (45). Site-directedmutagenesiswasperformedaspreviously described (22), (forthe mutationsintroduced, see Fig. 4A). The -41,-globin (-41BGCAT) plasmidwasdescribedby Fan and Maniatis (18). SVpCATwas generated by cloning the FokI (blunted)-HindIII fragment containing the simian virus 40 (SV40) promoter from pSV2CAT (23) into the EcoRV andHindIllsites of BS-CAT. Thefollowing double-stranded oligonucleotides, containing XbaI and BamHI sticky ends, werecloned in single and multiple copies into the BamHI and XbaI sites of -41BGCAT and SVpCAT: ZIB, CTAGCAGCTTATTTTAGACACTTC-3' and 5'-GATCGAAGTGTCTAAAATAAGCTG-3'; ZIC, 5'-CTAG CCTCCTCCTCTTTTAGAAACTAT-3' and 5'-GATCATA GTTTCTAAAAGAGGAGGAGG-3';ZII,5'-CTAGACGTC CCAAACCATGACATCACAGAGGAG-3' and 5'-GATCC TCCTCTGTGATGTCATGGTTTGGGACGT-3'. The(-221/ -129) SVpCATconstructwasgenerated by cloningtheSacl
(upstreamin theBluescript polylinkerfrom the bluntedSphI site in Zp)-EcoRI (created by site-directed mutagenesis of the
Z1ll
domain[21]) fragmentfrom the MIIImutantintothe SacI-EcoRI sites ofSVpCATand contains the regionfrom -221 to -129 base pairs (bp)upstream of the transcriptionstartsite. The(-129/-69)SVpCATconstructwasgenerated
by cloningthe NsiI(blunted)-EcoRI (created bysite-directed mutagenesis of the ZII domain) fragment into the SamI-EcoRI sites ofSVpCATand contains theregionfrom -129 to -70bp upstream of thetranscription startsite.
Preparation of nuclear extracts and DNase I footprinting. Nuclear extracts were prepared essentially as previously described (16). Briefly, 1 liter (approximately 1 g) of cells was harvested and washed with phosphate-buffered saline.
BamHI
,w
Y
II
4J
BHLF1
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.66.551.68.247.2]TPA RESPONSE ELEMENTS IN THE EBV BZLF1 PROMOTER 1219 TABLE 1. TPAinducibility of Zp and effect of theophylline treatment on various promoter-cat constructs in the
EBV-negative BL-cell lineRamosa
Mean + SE Zpactivitywith: Fold TPA inductionwith: Promoter
No treatment TPA Theophylline Theophylline andTPA Notheophylline Theophylline
-22lZp 0.03 +/- 0.005 0.04 +/- 0.004 1.0 +/- 0.13 25
SVp 0.11 +/- 0.03 0.12 +/- 0.01 1.3 +/- 0.07 1.0 +/- 0.11 1.1 0.7
JUNp 0.03 +/- 0.003 0.12 +/- 0.03 0.19 +/- 0.04 1.0 +/- 0.18 4.3 5.2
FOSp 0.01 +/- 0.005 0.28 +/- 0.07 0.07 +/- 0.01 1.0 +/- 0.22 28 14
aPlasmidswere transfected by usingDEAE-dextran,and cultures were incubated for 36 hposttransfectionwith and without 20 ng of TPA per ml and 1.2 mg
oftheophyllineper ml asindicated.The results areaveragesof three separateexperiments. In all cases,theophyllinetreatment increased theactivitywithout
significantlyaltering the TPA response. SVp, theenhancerlessSV40early promoter; JUNp(-132/+170CAT[1]),the promoter of the humanc-jun gene; FOSp
(FC4[15]),the humanc-fos promoter.
All subsequent steps were performed on ice or at 4°C. The cells were suspended in 5 volumes of ice-cold buffer A (10 mM HEPES
[N-2-hydroxyethylpiperazine-N'-2-ethanesul-fonic acid] [pH 7.9], 1.5 mM MgCl2, 10 mM KCl, 0.5 mM
dithiothreitol) and homogenized in a Dounce homogenizer, and the crudenuclear fraction was recovered by centrifuga-tion at 10,000 x gfor 10 min. The nuclei were suspended in buffer C (20 mM HEPES [pH 7.9], 25% [vol/vol] glycerol, 0.42 M KCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM
phenylmethylsulfonyl fluoride, 0.5 mM dithiothreitol) and homogenized again in a Dounce homogenizer. The extract was then stirred gently for 30 min at 4°C and clarified by
centrifugationat25,000x gfor30min. Thesupernatant was dialyzed against two changes of 50 volumes of buffer D (20 mMHEPES [pH 7.9], 20%[vol/vol]glycerol, 0.1 MKCl, 0.2 mMEDTA, 0.5 mMphenylmethylsulfonyl fluoride, 0.5 mM
dithiothreitol) for5h,followed bycentrifugationat25,000x gfor 20 min. Thesupernatant was recovered, and samples werestored at -70°C until readyfor use.
Footprinting was performedessentially as previously de-scribed (31). Binding reactions were performed in 10 mM Tris (pH 7.9)-0.5 mM EDTA-0.5 mM dithiothreitol-2.5% glycerol-2% polyvinylethanol with a final salt concentration of 40 mM KCl. Extracts were preincubated with 1 ,ug of
poly(dI-dc) in the reaction buffer for 10 min at 4°C. The labeledfragment was then added and incubated at22°C for 20 min before DNase I digestion. The samples were ex-tracted severaltimes withphenol-chloroform (1: 1), ethanol
precipitated, andrun ona6% acrylamide denaturing gel. For competition studies, the following oligonucleotides
wereused: RI,5'-AATTGAGGCTCGGTTATTTTGGTTCC T-3' and
5'-AATTAGGAACCAAAATAACCGAGCCTC-3'; RII, 5'-AATTGCCCCATGTGATGGTCAGGG-3' and
5'-AATTCCCTGACCATCACATGGGGC-3'. The ZIB and
ZIT
oligonucleotides used for competition studies are those described above for cloning into -41BGCAT andSVpCATplasmids.
RESULTS
TheBZLF1promoter (Zp)is TPAinducible in theabsence of other EBVantigens. Intissue culture, the virallyticcycle can be induced in various EBV-infected cell lines by treat-ment with several different reagents, including phorbol es-ters, butyrate, ionophore, and anti-immunoglobulin. The phorbolesterTPAis themostwidely used agent for produc-tion of virus from inducible cell lines and was chosen to
begin an analysis of the initial events involved in the lytic
cascade. For thesestudies,aregionof Zp from -221to +12 bp was clonedupstreamof the bacterial gene that encodes CAT (-221ZpCAT). -221ZpCAT was assayed foractivity
inaseries ofEBV-negativeandEBV-positiveB-cell lines in the absence or presence of20 ng ofTPA perml (data not shown). Although TPA-inducible activity was detected in EBV-positivecelllines, of the EBV-negative celllinestested (BL41, BJAB, and Ramos), only the BL cell line Ramos showed detectable activity (Table 1). These experiments
suggested that other EBVfactors are important for
activa-tion of Zp (21). However, to assess the response ofZp to TPAdirectly, it was necessary to studythis promoter in the absence ofother viral gene products through which TPA
inducibilitycould be mediated.
As will bereported elsewhere indetail(unpublished data),
treatmentof various cell lines with theophylline posttrans-fection caused a general enhancement in detectable CAT activity. It has been previously reported thattheophylline
treatmentinducesendogenousexpression ofthe differentia-tion-associated CR2 receptor (35). We initially tested this reagent to differentiate uninfected BL-cell lines to a state morecloselyapproximatingthatofEBV-infected B lympho-cytes,a statewhich may be necessary for efficientactivation of Zp. Theophyllinehas beenreportedtobea
phosphodies-teraseinhibitor andtoincreasecyclicAMP (cAMP)levels. As shownbelow, theBZLF1promotercontains an element
(ZII)which ishomologousto acyclicAMP responseelement
binding protein (CREB) site. Therefore, this reagent might activate BZLF1 expressionvia the
ZIT
domain.Theophyllinetreatmentincreased the TPA-induced
activ-ity of -221ZpCAT circa 10-fold in Ramos cells (Table 1).
However, this effect is not specific for the -221ZpCAT plasmid. Theophylline treatment of Ramos cells also pro-duced approximately a 10-foldincrease in CAT activity for
several other promoter-cat constructs. We also tested sev-eral other cell lines with a larger series of promoter-cat constructsandfound similar results (datanotshown).
Fur-thermore,asdiscussedbelow,wealsofoundthat theactivity
of the -221ZpCAT construct was not responsive to 8-bromo-cAMP or dibutyrl-cAMP, suggesting that elevated cAMP levels do not affect the activity of -221ZpCAT. Therefore,the dataindicate that the elevated levelsofCAT
activityintheophylline-treated cells is elicited viaageneral
mechanism.However,wedonotruleoutthepossibilitythat
theophyllinemayinsomewayactivateZp. We arecurrently testing whether theophylline exerts its effectby
increasing
the stabilityofcatmRNA.
Asshownfor severalexamplesin Table1,thebehavior of promoters in responseto TPAdidnotappeartobeaffected
by theophylline. Both the enhancerless
(without
the72-bp
repeats) SV40 early promoter and the human
P-globin
pro-moter(-41to +20) (datanot shown)exhibitednoapparent TPA inducibility eitherwith or withouttheophylline.
Simi-larly, the promoters for both c-jun andc-fos
exhibited TPA VOL.64,1990on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.612.58.556.95.165.2]1220 FLEMINGTON AND SPECK
inducibility which wasnotsignificantly altered by
theophyl-line. We therefore used theophylline to increase the sensi-tivity of this assay, thereby allowing detailed functional analysis ofZpinanEBV-negativecell line. Theexperiment whoseresults are showninTable 1alsodemonstratedthatin B lymphocytes the region ofZp from -221 to +12 bp is
responsive to TPA in the absence ofother viral antigens, demonstrating a direct link between cellular factors and
TPA-inducible Zp activity. This is consistent with the
pro-posed role ofthe BZLF1 gene in thelytic switch. Further-more, in the absence ofTPA, Zp exhibited little activity, consistent with its behaviorduringviral
latency.
Addition of upstream sequencesto-542bp didnotsignificantlyalter theTPA-inducible activity ofZp (data not shown). Therefore,
we focused our attention on characterization ofthe func-tional elements contained within the region from -221 to
+12bp.
Transcription factors from infectedand uninfectedB lym-phocytecelllines bindtoseveraldomains inZp.Asafirststep
in the identification and analysis of functional domains
within Zp, DNase I footprinting was used to assay for
protein-binding
domains. Nuclear extracts were prepared fromthe Ramos, Clone-13 (an EBV-positive Burkitt'slym-phoma
cellline), and X50-7 (atightlylatentlymphoblastoidcell
line)
celllines and usedtoassayforbinding
toZp (Fig. 2). Inthisanalysis, severalprotectedregionswereidentified(the
locationsofthefootprinted domains are shown inFig.
3A).
Each of these domains wasprotected with theextractfrom theEBV-negative cell line Ramosaswellas or better than witheither oftheEBV-positiveextracts.Thisindicates that protein factors binding to these regions are ofcellular
origin,
although this does not preclude thepossibility
that EBVfactors also bindto someofthe sameregions. Impor-tantly, it should be noted that neither TPA treatment northeophylline treatmentof Ramos cells altered the observed DNase I protection pattern(data notshown).
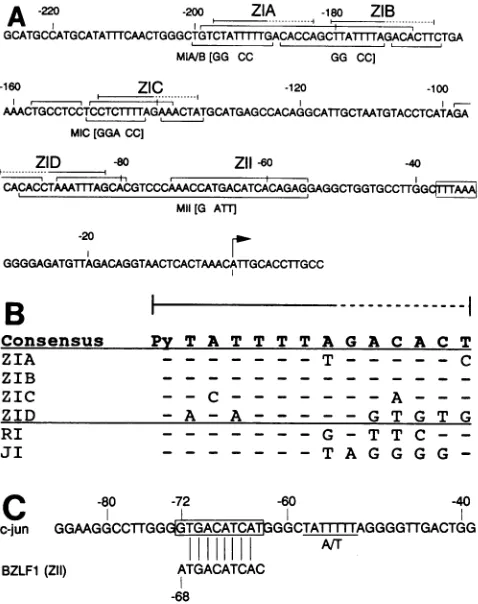
Multiple domains (ZI repeats) bind the same factor or
closely related factors.
Analysis
oftheprotein-binding
do-mainsequences(Fig. 3A) revealed thatfourof theseregions have significant homology to each other (ZI domains; an
alignment
of these domainsis shown inFig. 3B).Each repeatcontains anA+T-rich sequence
juxtaposed
to aregion
with a more complex sequence. The footprint for each repeat contains acharacteristic DNase I-sensitive site which mapsbetweentheleft-hand A+T-richsequenceand the
right-hand
sequence(see the minus strandforZIA, ZIB, and ZIC and theplusstrand for ZID inFig. 2), suggestingthateach repeat
consistsoftwobindingdomains.
To
investigate
thepossibility
that the same or closelyrelatedtranscription factors bindtothe ZIdomains,binding competition assays were performed by using synthetic
oli-gonucleotides.
Addition ofanunlabeledoligonucleotide
ho-mologoustothe ZIB domaineffectivelycompeted for
bind-ing
of nuclear factorstotheother ZI domains withinZp(Fig. 4A). Inthisexperiment,aninternalcontrolwasprovidedby the ZII domain. BindingtotheZIIdomainwasunaffectedby addition of the ZIB oligonucleotide. Moreover,oligonucle-otides homologous to the ZII domain or to an unrelated
binding
domain present in the BRLF1 promoter(RII) (Fig.4B)
didnotcompeteforbindingoffactorstothe ZIdomains(Fig. 4A).
DNase Ifootprinting analysisof the promoter for the EBV BRLF1 gene, which has also been shown to beexpressed very early in the onsetof the viral lytic cycle (7,
49),
revealedadomain,RI, which ispartially homologousto the ZI repeatsin Zp(Fig.3B). Aswasobserved with theZIBoligonucleotide,
an oligonucleotide homologous to RI alsoBZLF1 p
(-)
coc,
LO
ccu: X
II
ZIl ZID
T
ZICi
BZLF1p (+)
z a: x 0
ZIA ZIB T ZIC T
ZID
1
ZIl ZIB j
ZIA
I
FIG. 2. DNase Ifootprint analysisofZpfrom -221 to-35bp. Footprinting was performed withcrudenuclearextracts (80 p.gof totalprotein) fromtheindicatedcelllines (Ramos,anEBV-negative BL-cell line; Cl-13 (42), an EBV-positive BL-cell line; and X50-7 (43),alatentlyinfectedLCL-cellline). Protectedregionsare brack-eted, and ZI repeat sequences are indicated by vertical bars (the solid portion corresponds to the A+T-rich region within each repeat). N.E.,No extract. SeeFig.4for thecorrespondingfootprint sequence and type I repeatalignment.
competed for factors binding to the ZI domains, although slightly less efficiently (Fig. 4A). Conversely, the ZIB
oligo-nucleotide competed for binding of factorstothe RIdomain (Fig. 4B).
To rule out the possibility that the loss of binding of nuclearfactors tothe otherZIdomains by competition with the ZIB oligonucleotide was due merelyto loss of coopera-tive interactions between factors bound at the other ZI domains and those bound to the ZIB domain, competition assays were performed with Zp deletions (Fig. 4C). The -159Zp deletion, in which the ZIA and ZIB domains are deleted, had no effect on protection of the ZIC or ZID domain by Ramos nuclear extract, and the ZIB oligonucle-otide retained its ability to compete for factors that bindto both of these domains. Furthermore, deletion of the ZIC domain (-129Zp) had noeffectonbindingorcompetition at the ZID domain. We did not test whether competition for binding to ZIA is independent of the ZIB domain because of their close proximity. However, itappeared that binding of J.VIROL.
oVW 40 -*
,., 40
94p.
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.330.546.70.434.2]TPA RESPONSE ELEMENTS IN THE EBV BZLF1 PROMOTER 1221
A-220 -200 ZIA -180 ZIB
GCATGCCATGCATATi CAACTGGGCTGTCTATll GACACCAGcTTAT AGACA'CTTCTGA
MIAB[GG CC GG CC]
-160 ZIC -120 -100
, , ;- t,I
AAACTGCCTCCTCCTCTITUAGAAACTATGCATGAGCCACAGGCATTGCTMTGTACCTCATAGA MIC[GGACC]
ZID -80 ZIl-60 -40
,
I---CACACc-rAMmAGCACGTCCCAACCATGACATCACAGAGGAGGCTGGTGCTGuGffi
MIl [G ATT]-20 r0
GGGGAGATGTrAGACAGGTAACTCACTMACATTGCACCrTGCC
B
l
---Consensus Py T A T T T T A G A C A C T
ZIA - - - T- - - C
ZIB - - -
-ZIC - - C - - - A-
-ZID -A-A - - - G T G T G
RI - - - G- T T C
-JI - - - TA G G G G
-c
c-jun
BZLF1 (
-80 -72 -60 -40
GGMGGCCTTGGTGuGGCT TIA 1I AGGGGTTGACTGG
ANT
[ZII)
ATGACATCAC-68
FIG. 3. (A) Sequence of Zp and identification of footprint
do-mains. Overbrackets indicate sense (+) strand footprints, and
underbrackets indicate antisense (-) strand footprints. ZI repeat
sequences are indicatedby overbars (solid regions indicate A+T-rich domains). Specific base changes introduced for each site-directed mutation are shown below the corresponding sequence.
The arrowindicates the site oftranscription initiation. (B)
Align-mentofZI repeats. Theconsensus wasdeterminedonthe basis of
the four ZI repeats. Dashes indicate homology to theconsensus.
TheRIsequencespansfrom -160to-147 in the BRLF1promoter,
and the JIsequenceis indicated inpanelC. (C) Comparisonof the
c-junAP-1site with the BZLF1 ZII domain.
factorstothe ZI elements waslargely independent,and it is likely thatcommonfactors bindto thesedomains.
The ZI elementsareinvolved in the TPAinducibilityofZp. Toassay for the function of thebinding domains described above,aseriesof deletionmutantswereconstructed. These mutants were transfected into the Ramos cell line and cultured either withorwithout TPA(Fig. 5). Aplotof TPA
inducibility as afunction of the extent of deletionsuggests the presence ofmultiple TPA response elements (Fig. 6). Deletion of the ZIA and ZIB domains (-159Zp)decreased the TPA inducibilityfrom 20-fold to 9.1-fold. Further dele-tions which excised the ZIC and ZID domains resulted in only slightdecreases in TPA inducibility.
To assess the role of the ZI domains more precisely, site-directed mutagenesis of the A+T-rich sequence in the ZIAand ZIB domains (MIA/B)orinthe ZIC domain(MIC)
wasperformed (Fig. 3A).Both of these mutations resulted in a decrease in TPA inducibility (Fig. 5 and 6). The loss of TPA inducibility observed with MIA/B was approximately
equivalent to that of its counterpart deletion mutant, -159Zp,inwhich the ZIA and ZIB domainsaredeleted. In contrast, the loss of TPA inducibility observed with the MIC-mutantwasgreater than the loss observed bydeletion
of the sequence from -159 to -129 bp (which contains the ZICdomain) (Fig. 6), suggesting that the ZIC domain has a greater impact in the context of the upstream ZIA and ZIB domains. These results indicate that the A+T-rich se-quences are essentialfor the TPA inducibility conferredby this region of Zp.
To assaythe TPAinducibility of the ZI domains directly, single or multiple copies of the ZIB and ZIC domains were cloned upstream of the ,B-globin promoter (-41BGCAT) (Fig. 7). One copy of either the ZIB or ZIC oligonucleotide was sufficient to confer TPA inducibility on the ,-globin promoter. Cloning of three copies of the ZIB domain up-stream of the ,-globin promoter resulted in increased TPA
inducibilitycomparedwiththe-41BGCAT plasmid
contain-ing a scontain-ingle copy of the ZIB oligonucleotide. Notably, both the ZIB and ZIC oligonucleotides appeared to repress the activity of the
P-globin
promoterinthe absence of TPA. The mostdramatic repression was obtained with one copy of the ZIC oligonucleotide, which resulted in 53% reduction of promoteractivity.The repression of promoter activity conferred by the upstream ZI domains was more pronounced when a pro-moterfragment containing ZIA, ZIB, and ZIC was cloned upstream of the enhancerless SV40 promoter [Table 2; (-221/-129)SVp]. This construct, which leaves the ZIA, ZIB, and ZIC elements in their wild-type configuration, showed 6.4-fold TPA inducibility. However, the predomi-nant effect of adding this fragment upstream from the SV40 promoter wasrepression ofpromoteractivity inthe absence
of TPA, aneffect whichwas reversed with TPAtreatment. Thiseffectwaspartiallymimickedby cloningthreecopies of
the ZIB domain upstream of SVp (Table 2). In contrast,
cloning ofafragment ofZpcontaining the sequences from -129 to -69 bp (containing ZID but not ZII) upstream of
SVp exhibited no repressiveeffectand little TPA
inducibil-ity. Although this fragment contains the ZID domain, it also
containsapproximately 30bp ofupstream sequenceswhich
have not been rigorously characterized. Therefore, we do notknowwhether ZID is notresponsivetoTPA orwhether the apparent lackof TPAresponsiveness of this fragment is due to possible negative elements which might mask ZID TPA inducibility. This question requires further
investiga-tion.
TheZHIdomain is a TPAresponse element. The deletion
analysis shown in Figure 8indicates that the ZIT domain is alsoinvolved inthe TPA inducibilityof Zp. Comparisonof the TPAinducibilityof the -85Zpversus -65Zpconstructs
showed nearly a threefold loss ofTPA inducibility. More-over, when ZIl was mutated by site-directed mutagenesis
(Mll), the TPA inducibility of the promoter was reduced
from 20- to3.7-fold (Fig. 5and 6). We also tested the TPA
inducibility of the ZIl domain by cloningit upstream from the
P-globin
promoter(Fig. 7). Unlike the ZI elements, the ZIl domaindoesnot act as arepressorin the absenceof TPA in this system. Instead, the ZIl domain enhanced basal-levelexpression 4.0-fold, and addition of TPA resulted in a 2.7-fold increase inactivity.
The ZIT domain has a core octanucleotide sequence,
5'-TGACATCA-3', which contains apalindromic sequence found in both theconsensus heptanucleotide AP-1
recogni-tion sequence (2) and the octanucleotide CREBrecognition
sequence (39) (AP-1, TGA-TCA; CREB, TGA--TCA). The
palindromeintheAP-1-bindingsite isseparated by onlyone
nucleotide, whereas the CREB palindrome is separated by
two nucleotides, although exceptions do exist. The c-jun
promoter,forexample,containsanoctanucleotide sequence
VOL.64,1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.60.299.72.374.2]1222 FLEMINGTON AND SPECK
BZLF1 p(-)
N N 0 M N N CS
-.I
II
ZIA/B
ZIC
ZID
TI;IA
isuu..-.
zic
:S
ZIl
BZLF1
p(+)
I
*.
: |
t.
ZIB
*v
+a
*
.*.0 * ^I' w 4
**Itqp*eqp
ZIAI..
- w.*+
1
BRLF1
p(-)
B
=mCC CC Nv N
0 0 a
C:~ ~ z~ ( C
WWCDCOCOLiC
Z1+ + +Z Z
II
fl~~~~~~~!i
I~~~~I
a Iur.r,-.Rq
U,
Uw~~~~.SUe
1
w
w~~-
mmjMI!Rl3e:f
t
.*^~
-
*,,4
4.
s*i ! *-- w wo-.wRI
BZLF
p(-+t)
C
Li c
i.
CE5~~NEb9 a2Z
-!A
si,.mi
til
~~~~~C
Q3:
_
., - ,i Z Z N
w~~~~~~~
~~~~~~~~~~~~~tr
ZFIG. 4. (A)Competition for binding of transcription factorstoZIrepeats.An80-jig sampleof Ramos cell nuclearextractwaspreincubated
witheither 40or80ngof the indicatedoligonucleotides for 10 minatabout 4°C. A labeledsense(+)orantisense (-) Zpfragmentwasthen added, and the reactionwasallowedtoproceed for 20 minat22°C before digestion with DNaseI.(B) Competition for bindingtoBRLF1
promoter. A BRLF1promoterfragment extending from -190 to+4waslabeledon the antisense (-) strand. (C) Competition by theZIB
oligonucleotide for binding of nuclear factorstoZIdomainsusing Zp deletions. Labeled fragmentsweregenerated fromthecorresponding deletionmutants. Zpfragmentswerelabeledatthe HindlIl site between the catgeneand thepromoterfragment (sense strand).Promoter fragmentswerethen excised by digestion the Sacl site located in the Bluescript polylinker. Each fragment contains the sequencesextending
tothe indicatedbase in Zp plusanadditional 6 bp of the double-stranded polylinkersequence.An80-,ug sample of Ramos cell nuclearextract
andan80-ng sample of the ZIB oligonucleotidewereused.N.E., No extract;N.C., nocompetitor.
which is responsive to TPA and has been shown to be a
high-affinity AP-1-bindingsite (1). Moreover, thec-jun
pro-moterisnotresponsivetodibutyryl-cAMPin HeLacells (P. Angel, personal communication). The ZIl domain shares
many features with the c-jun AP-1 sites. (i) It contains an octanucleotide sequence with 8-of-8-bp homology to the c-jun AP-1 site (Fig. 3C). (ii) In our study, the BZLF1
promoter was not responsive to 8-bromo-cAMP or
dibu-tyryl-cAMPinthe Ramos cell line (datanotshown). (iii) The ZII domain is responsive to TPA. We therefore examined whetheracomplex ofc-jun andc-fos could bind tothe ZIl domain (Fig. 9). DNase I footprinting with extracts from baculovirus-infected insect cells (extracts were prepared either from cells infected with the wild-type virus or from
cellsinfected witharecombinant virusencoding theratc-fos
orc-jun gene) demonstrated binding of the c-jun-c-fos
com-plex totheZII domain.
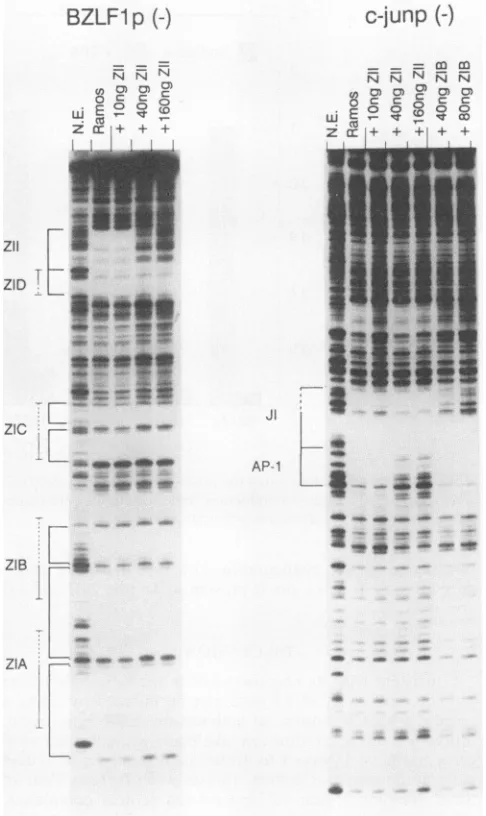
Consistent with the results obtained with HeLa cell
ex-tracts(1), thec-junAP-1sitewasprotected byextractsfrom theB-cell line Ramos(Fig. 8).Anoligonucleotide containing theZII domainwasableto competefor binding of
transcrip-tion factorstothec-jun AP-1 site(Fig. 8). Asacontrol, the ZlI oligonucleotide was used to compete for binding of factors to the ZIT site in the Zp promoter at the same
oligonucleotide concentrations. The ZII oligonucleotide competedfor factorbindingtoeither site withapproximately equal efficiency.On the basis of thesedata,weconclude that the ZII domainmay befunctionallyrelated tothe AP-1 site in thec-jun promoter.
Interestingly, although the ZIBoligonucleotidewhichwas usedas acontrol for the above-describedexperimentdidnot compete for factors binding to the c-jun AP-1 site, it did compete for factors bindingto a site adjacent toit (Fig. 8). Juxtaposedtothe AP-1 site isasequencehomologoustothe typeIdomain A+T-rich sequence(Fig. 3C). Unfortunately, this A+T-richsequenceinthec-junpromoteris recalcitrant to DNase I digestion. However, the sequence immediately
adjacent to the A+T-rich domain is sensitive to digestion and is protected in the presence of Ramos cell nuclear
extract (Fig. 8). The ability of the ZIB oligonucleotide to
compete effectively forbinding to this region suggests that the A+T-rich sequences are contained within this domain.
A
ZBl
ZID
T
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.57.561.73.408.2]TPA RESPONSE ELEMENTS IN THE EBV BZLF1 PROMOTER
1.4
* untreated * + TPA 1.2
0.8 a)
-0.6 (D
0.4
0.2
0
-221Zp -159Zp -129Zp -105Zp -86Zp -65Zp MIA/B MIC Mul
ZpCAT
Construct
FIG. 5. Bargraph illustratingtherelativeactivities of-221ZpCATandZpCAT site-directedanddeletionmutants(Fig.3A).CATassays were quantitated by liquid scintillation, and the activities are expressed relative to theactivityof -221ZpCAT (with TPA). The valuesgiven represent averages of three independent experiments.
Notably, a similar configuration of a type I domain juxta-posedto anAP-1-like site is present inZp(the ZID and ZIT domains).
DISCUSSION
Consistent with itsproposed role in the lytic switch, the promoter for the BZLF1 gene can be induced by TPA, a reagent which is known to activate the EBV lytic cycle. Moreover, this activation can take place without other viral gene products. DNase I footprint analysis of Zp identified multiple domains for cellular transcription factors. Four of these domains appear to bind related protein complexes, sincecomplex formation atthese sites can be competedfor successfully with an oligonucleotide containing only one of these domains (ZIB). In addition, several lines of evidence
indicate that these domains function as TPA response ele-ments. We further analyzed binding of factors to the ZI domains by gel retardation assays using oligonucleotides containing individual ZI domains (data not shown). The results of this study indicate that multiple transcription factors canbind toeach repeat. Further dissection of these
domains willberequired to assess the importance of specific
regions within the ZI repeats. However, the A+T-rich sequences appear to be important for their function, since
mutagenesis oftheA+T-rich core of ZIA-ZIBorZIClargely abrogates the TPA inducibilityattributableto those regions
of Zp fromfunctional analysis of Zp deletions.
Potentially,animportantfunction of the ZI repeats is their behavior as silencers which exhibit a TPA-inducible en-hancer function. This may be a mechanism which helps ensure quiescence of Zp and thereby EBV latency in the absence of the proper activation signals. TPA-responsive
elements with sequences similartotheZIelementsreported
here have not been previously described. However, an elementsimilartotheA+T-rich domainsdescribed here has beenreported in the enhancers oftwodifferent muscle genes (17, 24, 52) and in the promoter of theratmyosinlight-chain
25
20 _
15
CD p
S
10 _-.-5
0
-50
-200 -150 -100 -50
Nucleotides from cap site
FIG. 6. Scatterplotillustratingtheeffects of deletionsand site-directedmutationsonthe TPAinducibility of Zp. The solidboxes represent the TPA inducibilities of theindicated site-directed
mu-tants.
VOL.64, 1990 1223
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.131.487.76.304.2] [image:7.612.319.560.362.683.2]1224 FLEMINGTON AND SPECK
BZLF1
p
(-)
c-junp
(-)
*
e...e*
*
TPA -
+1
- +I
- +1
-+1
- +-41BGCAT (ZIB) BG (ZIB) BG
(ZIC),BG
(Zll),BG
1.2 x 1.9 x 2.5 x 3.2 x 2.7 x
FIG. 7. TPA inducibility ofZIand ZIl oligonucleotides cloned upstreamof the 3-globinpromoter. CAT assays were quantitated by liquid scintillation, and the fold TPA induction isindicated. These results were reproduced several times, and this reflectsa typical assay.
promoter (3). In the muscle creatine kinase gene, an up-stream enhancercontains a single copy of an element with the core sequence TTATTTTTA. This element has been
shown to bind two different factors, MEF-2 and MBF-1.
Synthesisof the former is rapidly induced upon
differentia-tion ofmyoblasts to myotubes, while the latter is found in
myoblasts and is down regulated during differentiation (24). Zp alsocontainsadistinct TPA response element located
withintheZlI domain. The ZlI domain contains an
octanu-cleotide core sequence (TGACATCA) homologous to the c-junAP-1-binding site (1). It was shown that c-jun and c-fos made frombaculovirus-infected insect cell extracts can bind to the ZlI domain. Binding competition studies demon-strated thatafactor(s) found in B cells bound with approx-imately equal affinity to the ZlI domain and the AP-1 site
foundin the c-jun promoter, suggestingmoredirectly that in Blymphocytesthesedomains are functionally related. How-ever, c-jun did not exhibit the ability to transactivate Zp when Ramos cells were cotransfected with ZpCAT and a c-jun expression vector (data not shown). In an analogous
experiment performed with a c-jun-cat construct (which
contains the c-jun AP-1 site), the c-jun promoter was not
transactivated by the c-jun-encoded protein (data not
shown). The latter is in contrast to results obtained with HeLa cells (1), which we were able to reproduce. It is
possible that the TPAinducibility ofthe ZlI domain in the Ramoscell line is due to a different member(s) of the AP-1
[image:8.612.316.559.73.481.2]familyand that c-junis not active in these cells or at least not TABLE 2. ciseffects of various Zpsequences on the
SV40early promotera
Mean+SEactivitywith: Fold
Plasmidinuto
No treatment TPA induction
SVp 1.0 +/- 0.07 0.9 +/- 0.11 0.9
3(ZIB)SVp 0.6 +/-0.05 1.2 +/- 0.09 1.9 (-221/-129)SVp 0.2 +/- 0.02 1.4 +/- 0.08 6.4 (-129/-69)SVp 1.0 +/- 0.05 1.1 +/- 0.04 1.1
a3(ZIB)SVp contains three copiesof the ZIBoligonucleotide upstreamof
theSV40 early promoter. (-221/-129)SVp contains promoter sequences from
-221 to-129,and(-129/-69)SVp contains the sequence from -129to -69
cloned upstreamof SVp. These resultsrepresent averagesof threeseparate
experiments.
_) __ C0
rE
z Er + + +
ZID
:iYe .i
_i
_4 _m
..:. \. iw
ZIA r
IJNNJ R CY)
a0) 0)gaC
z cc.+(,o ,r3+...+ +
iI
Jl1 -4.
-AP-1I
FIG. 8. Competition for binding oftranscription factors to the ZIldomain andthec-junAP-1domainwiththeZIIoligonucleotide. Experimentswereperformedasdescribedin thelegendtoFig.5by using the indicated amounts of the oligonucleotide. N.E., No extract.
active in conjunction with either the ZII or c-jun AP-1
domain.
Arecent report(53)investigating Zp activity in HeLa cells
mappedTPAinducibilitytotheregioncontainingZIA, ZIB, and ZIC. However, those investigators detected no TPA inducibilitymappingtotheregioncontainingZII. The reason for this difference is notclear, althoughit islikelyto reflect celltype-specificdifferences. Weconsidered the possibility thattheophyllinetreatmentactivated ZII to be responsiveto TPA. However, the effect ofdeletion of these domains in the
EBV-positive Burkitt's lymphoma cell line Cl-13 in the absence oftheophylline mimics the results presented here
(datanotshown).
We have shown that the TPA response elements in Zp appearto fall into two groups: the ZI repeats and the ZlI domain. Since different transcription factors interact with these distinct response elements, it is likely that TPA re-sponse is mediated viatwo different pathways. The nearly J. VIROL.
ZIB
go
f=:.
0
-.,on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.612.61.298.74.226.2] [image:8.612.63.300.611.680.2]TPA RESPONSE ELEMENTS IN THE EBV BZLF1 PROMOTER 1225
BZLF1
p
(-)
a
z Co2
t1)
0
+4
ZID
~a v
;.4 7,- -.
FIG. 9. Footprint analysis showing binding of c-jun and c-fosto
theZlI domain. A 2-,ugextractsamplewasused in each reaction, and for c-jun-c-fos binding, the reaction contained 1 ,ug of c-jun
extractplus 1 ,ug of c-fosextract.The controlextractwasfrom cells infected withwild-type baculovirus. N.E., Noextract.
complete loss of TPA inducibility and promoter activity which was observed when the ZlI domain was mutated
suggeststhatthis domain is anessentialpromoterelement. Incontrast, mutation of the upstream ZI domains (ZIAto ZIC) had a more moderate effect, indicating that these
elementsare notessential forZp activity. However, the ZI domainsareinvolvedintheTPAinducibility of Zp andmay
also functiontorepressZp in the absence of the appropriate
activation signals. As shown in the following report, the BZLF1 protein can transactivate Zp, and the combined
effects of BZLF1proteintransactivation and TPA induction via the upstream ZI domains (but not the ZIT domains)
appearto be synergistic (21). Furthermore, in thepresence
of BZLF1protein, afunctional ZlI domainisnotrequired.
ACKNOWLEDGMENTS
Weareparticularly gratefultoSadhnaAgarwal, Roman Herrera, and Tom Roberts forgenerousgifts of c-jun and c-fosextracts.We
thank Sadhna Agarwal and Joyce Fingeroth for helpful advice
during theprogressof this work andMary-Ann Sells,MikeKrangel, andCarolyn Doylefor criticalreadingof themanuscript.
This researchwassupported byPublic Health Service grant 5R01
CA43143 from the National Institutes ofHealth,aLeukemiaSociety
special fellowship to S.H.S., and a National Institutes of
Health-NationalResearch Serviceaward grant(1F32 CA08482-01)toE.F.
LITERATURE CITED
1. Angel, P.,K. Hattori, T.Smeal, and M. Karin. 1988. Thejun
proto-oncogeneispositively autoregulated byitsproduct, Jun/
AP-1.Cell 55:875-885.
2. Angel, P., M. Imagawa, R.Chiu, B.Stein, R. J. Imbra,H. J. Rahmsdorf, C. Jonat,P.Herrlich,and M. Karin. 1987. Phorbol
ester-induciblegenescontaina commoncis elementrecognized
byaTPA-modulatedtrans-actingfactor. Cell 49:729-739.
3. Arnold, H. H., E. Tannich, and B. M. Paterson. 1988. The
promoterof the chicken cardiacmyosin lightchain 2geneshows
cell-specific expression in transfected primary cultures of chicken muscle.Nucleic Acids Res.16:2411-2429.
4. Austin,P.J.,E.Flemington,C. N. Yandava, J. L. Strominger, andS. H. Speck. 1988. Complex transcription ofthe Epstein-BarrvirusBamHIfragmentHrightwardopenreadingframe 1 (BHRF1)inlatentlyandlytically infectedBlymphocytes.Proc. Natl.Acad. Sci. USA 85:3678-3682.
5. Baer, R.,A. T. Bankier,M. D. Biggin, P. L. Deininger,P. J. Farrell,T.J. Gibson,G.Hatfull,G.S.Hudson,S.C.Satchwell, C. Seguin, P. S. Tuffnell, and B. G. Barreli. 1984. DNA sequence andexpression ofthe B95-8 Epstein-Barr virus
ge-nome.Nature (London)310:207.
6. Bauer, G.,P. Hofler, and H. zur Hausen. 1982. Epstein-Barr virus inductionbya serumfactor:I.Induction andcooperation withadditional inducers. J. Virol. 121:184-194.
7. Biggin, M.,M.Bodescot, M.Perricaudet,and P. Farrell. 1987. Epstein-Barr virus gene expression in P3HR1-superinfected Rajicells. J. Virol.61:3120-3122.
8. Bodescot, M., 0.Brison,andM.Perricaudet. 1986. An Epstein-Barr virus transcription unit is at least 84 kilobases long. Nucleic AcidsRes. 14:2611-2620.
9. Bodescot, M.,B. Chambraud,P. Farrell, and M. Perricaudet. 1984. Spliced RNA from the IR1-U2 region of Epstein-Barr virus: presence of an open reading frame for a repetitive polypeptide.EMBOJ. 3:1913-1917.
10. Bodescot, M., M. Perricaudet, and P. J. Farrell. 1987. A promoter for the highly spliced EBNA family of RNAs of Epstein-Barrvirus. J. Virol.61:3424-3430.
11. Chevallier-Greco, A., E. Manet, P. Chavrier, C. Mosnier, J. Daillie,and A.Sergeant. 1986. BothEpstein-Barrvirus (EBV)-encoded trans-acting factors, EB1 and EB2, are required to
activatetranscriptionfroman EBVearlypromoter. EMBOJ. 5:3243-3249.
12. Cohen,L.K.,S. H.Speck,B. E.Roberts,andJ.L.Strominger. 1984.Identificationandmappingofpolypeptidesencodedbythe P3HR-1 strain ofEpstein-Barr virus. Proc. Natl. Acad. Sci. USA 81:4183-4187.
13. Countryman, J.,H.Jenson,R.Seibel,H. Wolf, andG. Miller. 1987.Polymorphic proteinsencodedwithin BZLF1 ofdefective and standard Epstein-Barr viruses disrupt latency. J. Virol. 61:3672-3679.
14. Countryman, J.,andG. Miller. 1985.Activationofexpression oflatent Epstein-Barr herpesvirus after gene transfer with a
small cloned subfragment ofheterogeneous viral DNA. Proc. Natl.Acad. Sci. USA 82:4085-4089.
15. Deschamps, J.,F.
Meilink,
andI. M. Verma.1985. Identifica-tionofa transcriptional enhancerelementupstream from the proto-oncogenefos. Science230:1174-1176.16. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcription initiation by RNA polymerase II in a
solubleextractfrom isolated mammalian nuclei. Nucleic Acids Res. 11:1475-1489.
17. Donoghue, M., H.Ernst, B.Wentworth,B. Nadal-Ginard,and N. Rosenthal.1988.Amuscle-specificenhancer islocatedatthe 3' end ofthe myosin light chain 1/3 gene locus. Genes Dev. 2:1779-1790.
18. Fan,C.-M.,and T.Maniatis. 1989. Twodifferent virus-inducible elements arerequiredfor humanP-interferon generegulation.
EMBO J. 8:101-110.
19. Farrell, P.J., D. T. Rowe, C. M.Rooney,and T. Kouzarides. 1989. Epstein-Barr virus BZLF1 trans-activator specifically
bindsto a consensusAP-1siteandis relatedtoc-fos. EMBO J. 8:127-132.
20. Fennewald,S., V. van Santen, and E. Kieff. 1984. Nucleotide sequence of an mRNA transcribed in latent
growth-transforming virus infection indicates that it may encode a
membraneprotein.J. Virol. 51:411-419.
21. Flemington, E., and S. H. Speck. 1990. Autoregulation of Epstein-Barrvirusputative lyticswitchgene BZLF1. J. Virol. 64:1227-1232.
22. Foss, K., and W. H. McClain. 1987. Rapid
site-specific
muta-genesisinplasmids. Gene59:285-290.
23. Gorman, C. M., L. F. Moffat, and B. H. Howard. 1982.
VOL.64, 1990
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.612.108.260.73.311.2]1226 FLEMINGTON AND SPECK
Recombinant genomes which express chloramphenicol acetyl-transferase in mammalian cells. Mol. Cell. Biol. 2:1044-1051. 24. Gossett, L. A., D. J. Kelvin, E. A. Sternberg, and E. N. Olson.
1989. A new myocyte-specific enhancer-binding factor that recognizes a conserved element associated with multiple mus-cle-specific genes. Mol. Cell Biol. 9:5022-5033.
25. Grogan, E., H. Jenson, J. Countryman, L. Heston, L. Grado-ville, and G. Miller. 1987. Transfection of a rearranged viral DNA fragment, WZhet, stably converts latent Epstein-Barr viral infection to productive infection in lymphoid cells. Proc. Natl. Acad. Sci. USA 84:1332-1336.
26. Hardwick, J. M., P. M. Lieberman, and S. D. Hayward. 1988. A newEpstein-Barr virus transactivator, R,induces expression of acytoplasmic early antigen. J. Virol. 62:2274-2284.
27. Henikoff, S. 1987.Unidirectional digestion with exonuclease III inDNA sequenceanalysis. Methods Enzymol. 155:156. 28. Kallin, B., J. Luka, and G. Klein. 1979. Immunochemical
characterization ofEpstein-Barrvirus-associated early and late antigens in n-butyrate-treated P3HR-1 cells. J. Virol. 32:710-716.
29. Kenney, S., J. Kamine, E. Holley-Guthrie, J.-C. Lin, E.-C. Mar, and J. Pagano. 1989. The Epstein-Barr virus (EBV) BZLF1 immediate-earlygene productdifferentiallyaffectslatent versus productive EBV promoters. J. Virol.63:1729-1736.
30. Laux, G., M. Perricaudet, and P. J. Farrell. 1988. A spliced Epstein-Barr virus gene expressed inimmortalizedlymphocytes iscreated by circularization of thelinearviral genome. EMBO J. 7:769-774.
31. Lee, W., P. Mitchell, and R. Tjian. 1987. Purified transcription factor AP-1 interacts with TPA-inducible enhancer elements. Cell49:741-752.
32. Lieberman, P. M., J. M. Hardwick, and S. D. Hayward. 1989. Responsivenessof the Epstein-Barr virus NotI repeat promoter tothe Ztransactivatoris mediated in acell-type-specificmanner by twoindependent signal regions. J. Virol. 63:3040-3050. 33. Lieberman, P. M., P. O'Hare, G. S. Hayward, and S. D.
Hayward. 1986. Promiscuous trans activation ofgene expres-sionby an Epstein-Barr virus-encoded early nuclear protein. J. Virol. 60:140-148.
34. Luka, J., B. Kallin, and G. Klein. 1979. Induction of the Epstein-Barr virus (EBV) cycle in latently infected cells by n-butyrate. Virol. 94:228-231.
35. Magrath, I., C. Freeman, and L. Novikovs. 1981. Induction of complement receptor expression in cell lines derived from human undifferentiated lymphomas. J. Immunol. 127:1034-1038.
36. Manet, E., H. Gruffat, M. Trescol-Biemont, N. Moreno, P. Chambard, J. Giot, and A. Sergeant. 1989. Epstein-Barrvirus bicistronic mRNAs generated byfacultative splicing code for twotranscriptional trans-activators. EMBO J. 8:1819-1826. 37. McCutchan, J. H., and J. S. Pagano. 1968. Enhancement of the
infectivityof simian virus 40 deoxyribonucleic acid with diethyl-aminoethyl-dextran. J. Natl. Cancer Inst. 41:351-357. 38. Miller, G., M. Rabson, and L. Heston. 1984. Epstein-Barr virus
with heterogeneous DNA disrupts latency. J. Virol. 50:174-182. 39. Montminy, M. R., K. A. Sevarino, J. A. Wagner, G. Mandel, and R. H. Goodman. 1986. Identification of a
cyclic-AMP-responsive elementwithin the rat somatostatin gene. Proc. Natl. Acad. Sci. USA 83:6682-6686.
40. Pearson, G. R., J. Luka, L. Petti, J.Sample, M. Birkenbach, D. Braun, and E. Kieff. 1987. Identification of an Epstein-Barr virus early gene encoding a second componentof the restricted earlyantigen complex. Virology 160:151-161.
41. Pfitzner, A. J., E. C. Tsai, J. L. Strominger, and S. H. Speck. 1987. Isolation and characterization of cDNA clones corre-sponding to transcripts from the BamHI H and F regions of the Epstein-Barr virus genome. J. Virol. 61:2902-2909.
42. Rabson, M., L.Heston, and G. Miller. 1983. Identification of a rare Epstein-Barr virus variant that enhances early antigen expression in Raji cells. Proc. Natl. Acad. Sci. USA 80: 2762-2766.
43. Rowe, D. T., P. J.Farrell,andG. Miller. 1987. Novel nuclear antigens recognized by human sera in lymphocytes latently infectedby Epstein-Barrvirus. Virology156:153-162. 44. Sample, J., M. Hummel,D.Braun,M.Birkenbach, andE.Kieff.
1986.Nucleotide sequences of mRNAsencoding Epstein-Barr virus nuclear proteins: a probable transcriptionalinitiationsite. Proc. Natl.Acad. Sci. USA83:5096-5100.
45. Sanger, F., A. R. Coulson, B. G. Barrell, A. J. H. Smith, and B. A. Roe. 1980.Cloninginsingle-stranded bacteriophageasan aid torapid DNAsequencing.J. Mol. Biol. 143:161-178. 46. Speck, S. H., A. Pfitzner, and J. L. Strominger. 1986. An
Epstein-Barrvirus transcriptfromalatently infected, growth-transformed B-cellline encodesahighlyrepetitive polypeptide. Proc.Natl.Acad. Sci. USA83:9298-9302.
47. Speck, S. H., and J. L. Strominger. 1985. Analysis of the transcript encodingthe latentEpstein-Barrvirus nuclearantigen I: apotentially polycistronicmessage generated bylong-range splicing of several exons. Proc. Natl. Acad. Sci. USA 82: 8305-8309.
48. Speck, S. H., and J. L. Strominger. 1989. Transcription of Epstein-Barrvirusinlatentlyinfected, growth-transformed lym-phocytes. Adv. ViralOncol. 8:133-148.
49. Takada, K., and Y. Ono. 1989. Synchronous and sequential activation oflatentlyinfected Epstein-Barr virus genomes. J. Virol. 63:445-449.
50. Takada, K., N. Shimizu, S. Sakuma, andY. Ono. 1986. trans
activationofthelatentEpstein-Barrvirus (EBV) genome after transfection ofthe EBV DNAfragment.J.Virol. 57:1016-1022. 51. Tovey, M., G.Lenoir, and J. Lours-Begon. 1978.Activationof latentEpstein-Barr virus by antibody to human IgM. Nature (London) 276:270-272.
52. Trask, R. V., A. W. Strauss, and J. J. Billadello. 1988. Devel-opmental regulationandtissue-specific expressionof the human musclecreatinekinase gene. J. Biol. Chem. 263:17142-17149. 53. Urier,G., M. Buisson, P. Chambard, andA.Sergeant.1989.The
Epstein-Barr virus early protein EB1 activates transcription fromdifferentresponsive elementsincludingAP-1bindingsites. EMBO J. 8:1447-1453.
54. Wong, K.-M., and A. J. Levine. 1986. Identification and map-ping of Epstein-Barr virus early antigens and demonstration ofa viral geneactivator thatfunctionsin trans. J.Virol. 60:149-156. 55. zur Hausen, H., F. O'Neil, and U. Freese. 1978. Persisting oncogenic herpesvirus induced bythetumourpromoter TPA. Nature(London)272:373-375.
J. VIROL.