JOURNALOFVIROLOGY, Oct. 1971, p. 579-588 Vol. 8,No.4 Copyright©1971 AmericanSocietyforMicrobiology PrintedinU.S.A.
Isolation
and Characterization
of
a
Herpesvirus
from Leukemic
Guinea Pigs
D. P. NAYAK
DepartmentofMedicalMicrobiologyandImmunology,Schoolof Medicine, Universityof California, LosAngeles, California 90024
Received for publication 7 June 1971
A guinea pigherpesvirus (GPHV) hasconsistently been isolated fromleukemic
lymphoblasts of strain-2 guinea pigs. GPHV is serologically related to the guinea
pigherpes-like virus isolatedby HsiungandKaplow. The virions of GPHVconsist
of an icosahedralcapsidcontaining a dense nucleoprotein core enclosed in a
double-layered membrane.The averagediametersofGPHVvirionandcapsidwere 166 and
101 nm,
respectively.
Studies onthemorphogenesis of GPHVrevealed that, as inotherherpesvirusinfections, onlythenakedcapsidswith or withoutthenucleoprotein
corewerefound in the infectedcellnuclei;itwasalso learned that thevirionacquired
its
envelope by budding
from thenuclear membrane oftheinfectedcells. However,GPHV-infected cell nuclei also contained dense fibrous rods, resembling nucleo-protein core outside the capsids, and tubules resembling viral core protein. The
capsids were often embedded in dense granular antigen. GPHVdeoxyribonucleic
acid (DNA) has a density of 1.716 g/ml in cesium chloride compared to herpes simplexvirus DNA (p = 1.728g/ml) and cellularDNA (p = 1.700g/ml).
Opler's
observation
(18)
ofC-type particles
in lymphoblasts of leukemicguinea pigs suggested
aviral
etiology
forguinea
pig
leukemia. Nadel etal. (14), and more recently Feldman and Gross
(6),
alsoobserved suchparticles
inleukemic,
butnotinnormal, guinea
pigs.
However,no onehassuccessfully
demonstrated that theseC-type
particles
are theleukemogenic
agent inguinea
pigs. We also studied the role of viral agentsin guinea
pig
leukemia andconsistently
found C-type particles budding from theendoplasmic
reticulum of leukemic
lymphoblasts.
However,we were unable to
produce
this agent in tissueculture in sufficient
quantity
to characterize itsbiological
andbiophysical properties.
During
this
investigation,
weconsistently
noted theactivation
ofasecond virusinleukemiclympho-blasts culturedinvitro. Thisagent hasthe
prop-erties
ofherpesvirus.
A similar virus bas beenisolated from normal and leukenic
strain-2
guinea
pigs by Hsiung
andKaplow
(9)
and morerecently
fromother strains ofguinea pigs
aswellby Bhatt et al.
(2).
This report describes somemorphological
and biochemical characteristicsof this
guinea
pig
herpesvirus (GPHV)
whichwasisolatedfromleukemicstrain-2
guinea pigs.
MATERIALS AND METHODS
Guineapigs. All the strain-2guinea pigs (both in-fected anduninfected) used in theseexperimentswere
kindly supplied by S. Opler of Stanford University.
Leukemia in these animals is maintained by
inoculat-ingtissues ofinfected animals (spleen, blood, tumors, etc.)into healthy animals.Noinfectious leukemogenic
agenthas yet beenisolated from theseleukemicguinea pigs. Infectedanimals develop leukemia within 2 to 3 weeks afterinoculation. Theanimalsin terminal phase
ofleukemiawereexsanguinated bycardiacpuncture.
Gross andhistopathology ofthediseasehave been
de-scribedbyOpler (19). HartleyandEnglish short-hair
strainsofguineapigswerepurchasedlocally.
Cell culture. Usually, circulating lymphoblasts or
lymphoblastsfromalocal tumor,mesentericlymphoid
tumors, and spleen of leukemic guinea pigs were
placedintissueculture. ModifiedEagle medium
con-taining inactivated fetal calf serum (10 to 20%),
tryptosephosphate broth
(10%,)
penicillin (200IU/ml), and streptomycin (200,Ag/ml) was used. The
medium was normally changed twice a week or as
needed. Guineapig embryocultures (GE) were pre-pared from 1-to 1.5-month-oldembryos byusingthe
usual procedureofcell culture.
Labeling ofinfected cells. Monolayers of cultures
were infected with GPHV. When the initial
cyto-pathic changes became evident (usually 6 to 8 days
after infection), the old medium was replaced with
medium containing 3H-thymidine (specificactivity, 25 Ci/mM; 20 uCi/ml) and dialyzed fetal calf serum (10%).Thefresh mediumwasincubated for48to 72
hruntilall of thecells rounded up,atwhich time the
medium and thecellswere harvested and processed
forvirus purification as well as for the isolation of intracellularDNA.
579
on November 11, 2019 by guest
http://jvi.asm.org/
NAYAK
Viruspurification. The culture medium was freed from cells and subcellular debris by two cycles of
centrifugationin an RC-2B Sorvallcentrifuge(10,000
rev/min for 10 minin an SS 34 rotor).Viruswas con-centrated twice at the interface of 70 and 20% sucrose
andfinally purified by density-gradient centrifugation
in apreformedlinear70 to20%sucrosegradient (15).
Aftercentrifugation,thedensity, radioactivity, and
in-fectivity ofeachfractionwereanalyzed,andpeak
frac-tionswereexamined bynegative stainingandthin
sec-tioningfor the presence of virusparticles.
Isolation and analysis of viral and cell DNA.
Deoxy-ribonucleic acid (DNA) frominfected cellsand
puri-fiedviruswasisolated andanaylzedin a cesium
chlo-ride gradient asdescribed by others (21, 25). Thecells
orfractionscontaining viruswerelysed with sodium
dodecyl sulfate (3%), dialyzed, and treated with
self-digested Pronase (200,ug/ml). DNA soisolated
wasmixedwith concentratedcesium chloridetomake
asolution withadensityof 1.700g/mlandcentrifuged
at35,000rev/min for72hr at 20 C inanSW 50 rotor.
Fractionswerecollectedandanalyzed for densityand
radioactivity.
Electronmicroscopy.Smallpiecesofselectedtissues,
culturedcells, or pelleted virus materialswere
imme-diately fixedin2%glutaraldehydeinphosphate buffer
(pH 7.2 to 7.4) for 30 to 60 min, washed in three changes of phosphate buffer for 1 min each, and treatedwith
1%
osmium tetroxideinphosphate buffer(pH 7.2 to 7.4) for 30 min. Thetissuewasthenrinsed
inRinger's solutionandblock-stained for 30min in
1% aqueousuranylacetatebroughttopH4.9to5.0
withNaOH.All of theforegoing solutionsweremade
slightlyhypertonic withsucrose.Dehydrationwas
ac-complished with the usual ethanol series. Thetissue
blockswereinfiltratedfirstwitha1:1 mixture of
ab-solute ethanol and embedding media and then
em-beddedinUNOXepoxy (Spurr).Fornegative
stain-ing, virus particleswerefixed on carbon-coatedgrids
andstained with1%phosphotungstic acid.
RESULTS
Isolation
of the GPHV. Spleencells, lymphoidtumors, local tumors, and
circulating
leukemiclymphoblasts
wereplaced in culture in modifiedEagle medium containing inactivated fetal calf
serum
(10
to20%)
and 10%tryptosephosphatebroth. The medium was changed
twice
aweek.In such cultures, most of the cells remained in
suspension, andover 80% weredead in72hras
determined by dye exclusion. Some fibroblasts
adhered tothe bottom ofthe dishes in about a
week. Bythesecond week, a few foci ofclumped
cells
appearedinculture.Finally,
all ofthecells
would round up and die around the fourth or
fifth week ofculture. It is at this time that the
herpes-like virus (GPHV) can be first seen in
the electron microscope. The results were the sameifafecderlayerof GEculturewasusedfor
these leukemic
lymphoblasts.
Morphology. The morphology of GPHV was
studied both bynegative staining of intact virions isolated from sucrose-density gradientsaswellas
by positive staining ofa thin section of
virions
and infected cells. It was clear that the tissue
culture-activatedGPHV wascompletelydifferent
from mature and
immature
C-typeparticles
ob-served in vivo in leukemic lymphoblasts by
Opler
(18), Nadel et al. (14), Feldman and Gross (6), andbyus (Fig. 1). TheseC-type viruseswith an
average diameterof 101 nm bud fromthe
endo-plasmic reticulum into the cytoendo-plasmiccysternae.
GPHVon the otherhandhas themorphological
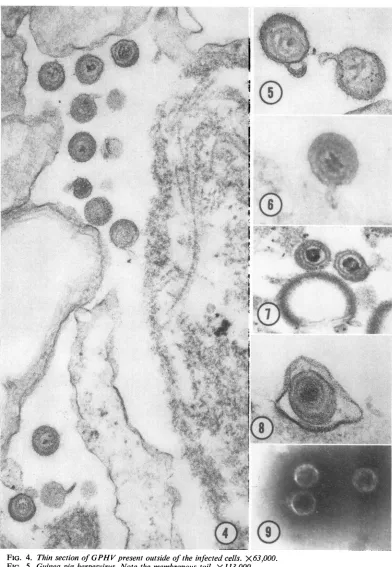
characteristics ofa herpesvirus (Fig. 4-9; Table
1).
Mature GPHV consists of an icosahedralcapsid containing
a densenucleoprotein
coresurrounded bya double-layered membrane. The
outermembraneoften extends with the
formation
ofasmall tail. Mature GPHVis spheroidal
and
has an averagediameter of 166 nm. The
icosa-hedral capsid is about 101 rum. There is a dense material between the capsid and the outer
en-velope (Fig.5and7). Negative staining of purified virions andcapsids also revealed thesame
struc-tural components. Large virion-like structures
containing
several(as
many aseight) capsids
(Fig. 12) and naked capsidswere oftenfound in
some
preparations
of purified virion isolatedfromsucrosegradients(Fig. 9).
Biological and serological properties. Once the
virus is activated it can be passed serially in
normal guineapig embryonic culture. Itusually
takes 7 to 14days to
produce visible
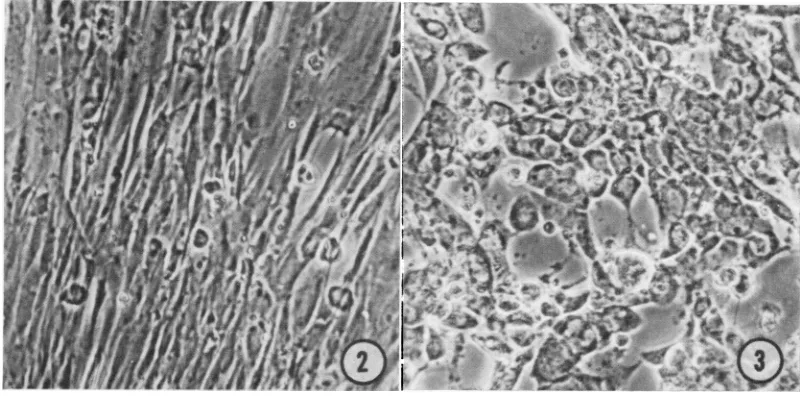
cytopathic effect (CPE; Fig. 2 and 3). CPE consists of in-fected cells which become round, refractile, and loose and which often clump together. In GEcells,
themaximum titeris around 104infectiousunitsasdetermined by endpoint dilution in tissue culture. This titer is50- to 100-foldovertheinput
virus.
Freezing and thawing once reduced theinfectivity at least 10-fold. The infectivity of GPHV is also sensitive to ether treatment. CPE
in GE cultures after infection was enhanced by incubation in reduced temperature (33C), in acidic culture
medium,
and in mediumdeficient
in
arginine.
All ofthese
conditionshave
beenfound to activate the EB virus inBurkitt's
lym-phoma
cells(7,
9).
GPHV also causes CPE inprimary rabbit kidney
cells
and Vero monkey kidney cells and isserologically
relatedto GPHLV(9)
as determined by neutralization test and by immunofluorescence technique. Virus neutraliza-tion test was carried out onguinea pig embryo
cultures as described by Bhatt et al. (2);
virus
titerwas reduced 1,000-fold
by
thehomologous antisera and the antiseraagainst
GPHLVpro-vided
by
G. D.Hsiung.
Morphogenesis.Morphological developmentof
subviralcomponents and
assembly
ofthesecom-580 J. VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
VOL.8,1971 HERPESVIRUS FROM LEUKEMIC GUINEA PIGS 581
M~~~~~~"
ke
Ai
i
b''~
X:
.A4'
*tshE
'4 C.,
%
t.
*.S.
^i",`
i*
B s ' ts '' ; - FXXt~~~~~~~~~~~~Zt
i*'''+4
Wk,
4i"yf:'m
d^
V*6
C ~ ~ X ,4! ' ;<
:WNPon:- l 0w > ttX_
[image:3.489.38.434.67.380.2]FIG. 1. ImmatureC-type
particles
inleukemiclymphoblasts budding from
endoplasmic
reticulum. X84,000.
FIG. 2. Uninfected guinea pig
embryo
culture. X125.FIG. 3. Guineapig
embryo
cultureinfected
with GPHV.Cytopathic
effectwasevident 7days
postinoculation.
X 125.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.489.35.435.415.613.2]@ee
%. tt=e z -s;S'st-afs
5iP w O $ ..,mtr
SZ
-*t .S
3|- ,f i
v,.e >;
Sll;> ;5-2 f , ¢;
/4
I
[image:4.489.64.456.45.612.2]it It
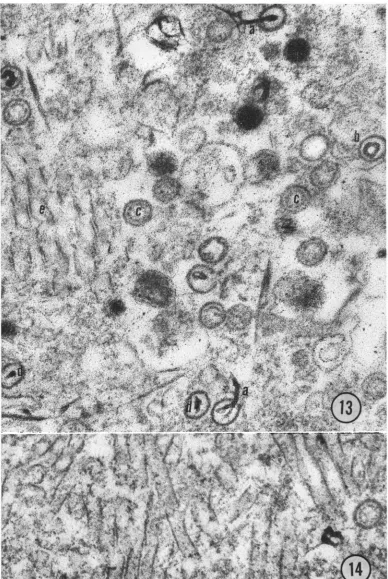
FIG. 4. ThinsectionofGPHV presentoutsideoftheinfectedcells. X 63,000. FIG. 5. Guinteapigherpesvirus. Notethemembranous tail. X 113,000. FIG. 6. Guineapigherpesvirus.Notethe segmentation of nucleoid. X 113,000.
FIG. 7. Guinea pigherpesvirus. Notethe eccentricpositionof thecapsidandpresenceofdensematerialbetween
thecapsid andenvelope. X84,000.
FIG. 8. Guinea pig herpesvirus enclosed in apinocyticvacuoleon the cellmembrane. X113,000.
FIG. 9. Negativelystainednaked capsids without any envelope isolatedfromsucrosegradient. X84,000.
582
4k
on November 11, 2019 by guest
http://jvi.asm.org/
HERPESVIRUS FROM LEUKEMIC GUINEA PIGS
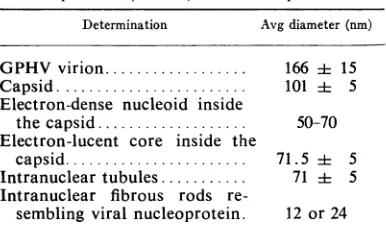
TABLE 1. Morphological characteristics ofguinea pig herpesvirus (GPHV) and its components
Determination Avg diameter(nm)
GPHV virion... 166 i 15
Capsid... 101 i 5
Electron-dense nucleoid inside
thecapsid ... 50-70 Electron-lucent core inside the
capsid... 71.5i 5
Intranuclear tubules... 71i 5
Intranuclear fibrous rods
re-sembling viral nucleoprotein. 12 or24
ponentsintomorphologicallymaturevirionswere
studied in GE cell monolayers grown over a
plastic sheet and infected with GPHV. When
infected foci became evident, the cellswere fixed
andembedded; specificfoci of cellswere selected
under the light microscope, sectioned either in
parallel or vertical plane to themonolayer, and
examined with the electron microscope. In this
way, cells inthe same foci atdifferent stagesof
infection could be observed.
Ingeneral, the morphological development of
GPHV followed closely that of other
herpes-viruses. The nuclear phase of GPHIV consisted
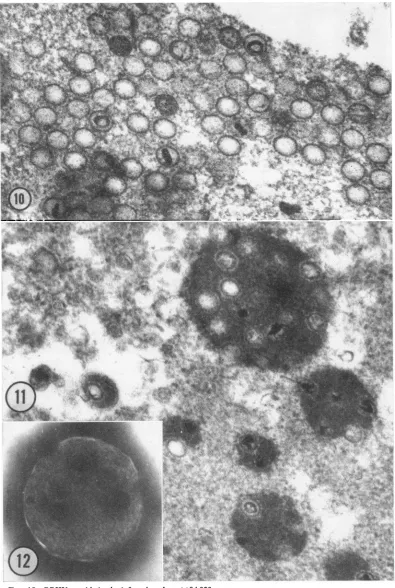
ofnaked icosahedral capsids,mostofwhichwere
empty (Fig. 10). The electron-dense material,
when found inside thecapsids, appearedin
differ-ent stages of condensation: ring form or dense
nucleoid (Fig. 10 and 13). Some capsids possibly
containedonlycoreproteinasapparent fromthe
lack of electrondensity (Fig. 13).Thesestructures
representing different stages in the development
of GPHVcapsidswere similar to those found in
other herpesvirus infection (3, 4). Also found
inside the nucleuswere dense,rod-like structures
as reported in some herpesvirus-infected cells
(4). The rods resembled DNA protein core and
wereoften found in close association withcapsids
(Fig. 13). Inside the nucleus, capsids wereoften
found embedded in a spherical electron-dense
material (Fig. 11) which was similar to
intra-nuclear granular antigens inherpesvirus-infected
cells (16, 23). Inlater stages ofinfection, these
intranuclear structurespossibly disintegrate into
smaller pieces containing one or more capsids and are released into the medium after lysis of
infected cells. This could explain the origin of
somelarge virus-like particles containing multiple
capsids found in the preparation of purified
virions(Fig. 12).This denseintranucleargranular
antigen is present also around single capsids, is
often eccentric in arrangement, and resembles
closely the dense material often found between
the capsid and the envelope (Fig. 4, 5, 7). A
similar dense material between the envelope and
the capsids in the herpesvirus associated with
frog renal adenocarcinoma has been reported
(24). Tubular structures which have the same
density and diameter as the empty coreproteins
found inside the capsids can also be seen in the
nucleus (Fig. 14). Such tubules have beenfound
in guinea pig cytomegalovirus infection (13).
Complete virionswere not found in the
nucleus;
however, cytoplasmic invagination containing
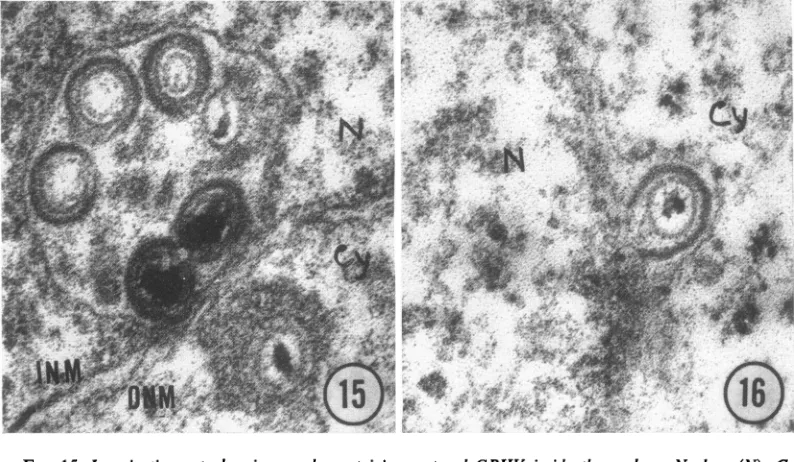
matured virions was observed inside thenucleus
(Fig. 15). The virions acquire their envelope
during budding from the nuclear membrane (Fig. 16) as has been reported by others (4).
Matured virions were found in closeassociation
with the plasma membrane and outside the cell
membrane(Fig. 8), but very few could be located
inthe cytoplasmic vacuoles.
Putification of viiions and characterization of
viral genome. The morphology and the
matura-tion of the virions suggest that this agent is a
herpes-type virus. Next. experiments were de-signed to purify the virus and characterize the
viral genome. Cells were infected and labeled with 3H-thymidine (3H-TdR) when foci
became
visible.After labeling for 72 hr, themediumwas harvested. The virus was concentrated on 2 M
sucroseandpurifiedonasucrose-densitygradient
(20 to 70%) as described above. A sample of eachfractionwascounted, and the fractions
con-taining
the peak ofradioactivity were examinedby negative staining for virus particles. The in-fectivity in thepeakfractions of the gradientwas
around 105-5 units/ml and coincided with the peak ofradioactivity. The recovery ofthe total
infectious virus in the sucrose gradient was only
10 to 15% of the totalinfectious units used in the
purification procedure. DNA was isolated from purified
GPi-V
andwasfoundtobeheavierthan GE cell DNA (p = 1.700 g/ml).GPkIV
DNA had a density of 1.716 g/ml in cesium chloride(Fig. 17).
When DNA was isolated from infected cells
after
labeling with 8H-TdR,
asabove,
the newly synthesized viralDNAalso had apeak of 1.716g/ml
(Fig. 18). 14C-TdR-labeled DNA ofHeLacells
infected with
herpessimplex
viruswasusedas a marker in the same
gradient.
HeLa cell DNAand HSV DNA hadadensity of 1.700 and1.728 g/ml, respectively, incesium chloride (20).
3H-labeled DNA isolated from purified GPHV and 14C-labeled DNA isolated from
GPHV-infected cells had the samedensity if run inthe
same cesium chloride gradient. Further proof
thatthe nucleicacidin GPHVvirionisDNA was
demonstratedby its deoxyribonucleasesensitivity, resistance to ribonuclease, and resistance to
alkali.
VOL.
81
1971 583on November 11, 2019 by guest
http://jvi.asm.org/
541
FIG|.1.y. Mutil capsids...embdde '"ingrnua anie in...th nuli .Als not Sinl casd suroude by.t,.
FI.1!Ngtvl stie viu-lk patces cotainingFmutil casd islae fro sucrose grains
X84.000. GH asd
nteifee
ulu.X400
584
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.489.65.460.36.624.2]41AW
4
.44
$
Ii .
,,'.A;,
/\'k 4b i.:
I.
*0.i;_.R/;t, .,' >t;i d
~~~~~fr4~~~I
FIG. 13. Sectionofaninfected nuclei. X84,000. (a) Fibrousrodresemblingviralnucleoprotein. (b)Ring form
of nucleoprotein inside the capsid. (c) Capsids containing protein core only. (d) Section of a densenucleoprotein
core. (e)Sectionsof tubules.
FIG. 14. Sectionofmassesof tubulesfound in infectednuclei. X84,000.
585
', ',.
vlp
2.'.I ..",
I
...
Ion November 11, 2019 by guest
http://jvi.asm.org/
[image:7.489.44.432.32.611.2]586
NAYAK J.VIROL.d
.T4rm..4.e
FIG. 15. Invagination cytoplasmic vacuole containing matured GPHV inside the nucleus. Nucleus (N), Cy
(cytoplasm), INM (inner nuclearmembrane), ONM(outer nuclearmembrane). X84,000.
FIG. 16. BuddingGPHVfromnuclearmembrane. X84,000.
300
DROP NUMBER
FIG. 17. Cesium chlorideequilibrium centrifugation of3H-labeledguinea pig herpesvirusDNA (a) along with I4C-labeled guinea pig embryocell DNA(b).
DISCUSSION
Characteristics and morphogenesis of GPHV.
GPHVreportedinthispaperisclosely related, if
notidentical,toGPHLVisolatedby Hsiung and
Kaplow (9) and Bhattetal. (2) and is, therefore,
different from guinea pig cytomegalovirus. The
morphological and biochemical data indicate
that GPHV has the characteristics of a
herpes-virus. It isanenvelopedviruswithaicosahedral
capsid which contains a nucleoprotein core. Its
=5.0
i- 1:-;E
T
o00
*200
*100
3 b c
814
100 200 300 0R0p NU M B E R
400
180
170
_
160
FIG. 18. Cesium chloride equilibriumcentrifugation
of3H-labeled guinea pig cell DNAinfectedwith guinea
pigherpesvirusandU4C-labeledHeLacellDNAinfected
with herpes simplex virus. (a) Intracellular herpes simplex DNA. (b) Intracellular guinea pig herpesvirus DNA. (c)HeLacell DNA.
nucleic acid is double-stranded DNA. The
as-sembly
ofcapsids
andnucleoprotein
takesplace
in the cell nucleus, and the outer envelope is acquired
during budding
from the nuclear mem-brane. The virion is released in the mediumby
reverse
pinocytosis
orby
lysis
ofcell. Thelarge
numbers of empty capsids in the nucleus, the large number of fibrous structures
resembling
DNA-containing protein core, and the
paucity
on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.489.64.461.71.302.2] [image:8.489.69.272.342.506.2]HERPESVIRUS FROM LEUKEMIC GUINEA PIGS
of
complete
virions either
in the cytoplasm oroutside the cell indicate a
possible
defect in the assemblyprocess.Relationship ofGPHVwith guinea pig leukemia.
Twodifferentviruses have, therefore, been found
in lymphoblasts of strain-2 leukemic guinea
pigs. (i) A C-type
virus particle
hasbeen observed in vivo, but usually not in vitro (Fig. 1).(ii)
GPHV, a herpes-type DNA
virus,
isisolated
fromguinea pig cells only in vitro culture andnot
invivo.
Itis
possible
thatguinea
pig leukemia is caused
by oneofthesetwovirusesor
by
theinteraction
of both viruses or
by
none. Wepsic et al.(27)
and Sarma etal.
(22)
havesuggested that certain forms ofguinea pig
leukemia may notpossibly
be
caused
by
aninfectiousvirus. Studiesby Opler
(18), Nadel et al.
(14),
and Feldman and Gross(6),on theotherhand,suggestthataC-type virus
is the cause of the
guinea pig
leukemia even thoughphysicochemical
characterization of thisvirionand its definitive role inguinea pigleukemia
remains to be done. Their strongest argument,
thatsuch C
particles
arepresentonly
in leukemic guineapigs,
has been weakenedby
the recentfinding of C-type
particles
resemblingGPLV-Opler in germinal centers of lymph nodes of
Hartley guinea pigs, a
strain
notsusceptible
toleukemia
(11).
However, the presence ofC-type
particles
in nonleukemicguinea pigs
does notruleoutits
etiological
roleinguinea
pig
leukemia.GPHV, ontheother
hand,
hasnowbeenisolated
from leukemic and
nonleukemic
strain-2guinea
pigs (2,
9).
Preliminary
datasuggestthatGPHV,
when inoculated
directly,
fail toinduceleukemia
in strain-2 or Hartley
guinea
pigs (unpublished
data).
Inability
to induceimmunity against
asimilar
herpesvirus isolated by
Bhatt et al.(2)
indicated the tolerance of
guinea pigs
to GPHVantigen,
suggesting
eithercongenital
orhereditary
transmission of GPHV. The characteristics of
GPHV are very similar to those of EB virus
(EBV) found inBurkitt'slymphoma.EBVis also activated in
lymphoblasts only
in tissue culture andnotfound inlymphoblastic
cells invivo(5).
Theconditions ofactivation forEBV and GPHVare very similar
(7,
8).
Thedensity
of GPHVDNA
is similar
tothatofEBVDNA(26)
whichis also
similar
tocytomegalovirus
DNA. Theinfection of EBV is also widespread and not
limited to persons with Burkitt's lymphoma.
Herpes-typeviruses havebeenfoundasthecausal
agent in
lymphoma
inmonkeys(12)
and Marek'sdiseaseinchickens (17). Possible role ofGPHVin inducing guinea pig leukemia has recently been
suggested (10).
Whether GPHV aloneorinasso-ciation withaC-typevirus has any
etiological
role inguinea pigleukemia remains to be established.ACKNOWLEDGMENTS
The author gratefully acknowledges the assistance of Zane Price for electromicroscopy and acknowledges his indebtedness to G. D.Hsiung of Yale University School of Medicine and Veterans Admimstration Hospital, West Haven, Conn., for her suggestions andalso for supplying the immune sera to GPHLV (9).
This work was supported by a SeniorDerhnam Fellowship
(Dl52),American Cancer Society,California Division, and also by California Institute of Cancer Research, and Cancer Research Coordinating Committee, University of California.
LITERATURE CITED
1. Abraham, A., and P. Tegtmeyer.1970. Morphological changes in productive andabortive infection by feline herpesvirus. J.Virol. 5:617-623.
2. Bhatt, P. N., D.H.Percy, J. L.Craft, and A. M. Jonas. 1971. Isolation and characterization of a herpes like (Hsiung-Kaplow) virus from guinea pigs. J. Infec. Dis. 123:178-189. 3. Cook, M. K., and J. F. Sears, 1970. Preparation of infectious
cell-freeherpes-type virus associated withMarek'sdisease. J. Virol. 5:258-261.
4. Cook, M. L., and J. G.Stevens. 1970.Replicationof varicella-Zoster virus inceU culture:anultrastructural study.J. Ultra-struct.Res.32:334-350.
5. Epstein,M. A.,and B. G. Achong. 1970. The EB virus, p. 231-248. In D. P.Burkitt andD.H.Wright (ed.), Burkitt's
lymphoma.E & S Livingstone,London.
6. Feldman, D. G., and L. Gross. 1970. Electron microscopic study of the guinea pig leukemia virus. CancerRes. 30:2702-2711.
7. Henle,W., and G.Henle. 1968. Effect of arginine-deficient media on the herpes-type virus associated with cultured
Burkitttumorcells.J. Virol.2:182-191.
8. Hinuma,Y.,M.Konn, J.Yamaguchi, D.J.Wudarski, J. R. Blakeslee, Jr.,and J. T.Grace, Jr. 1967.Immunofluorescence
and herpes-type virus particles in the P3HR-1 Burkitt
lymphomacell line.J. Virol. 1:1045-1051.
9. Hsiung, G. D., andL.S.Kaplow. 1969 Herpeslike virus iso-lated fromspontaneouslydegenerated tissue culture derived fromleukemia-susceptible guinea pigs.J.Virol. 3:355-357. 10. Hsiung, G.D.,and L. S.Kaplow. 1970.Theassociation of
herpes-likevirus andguinea pigleukemia.Bibl. Haematol. 36:578-583.
11. Ma,B.I.,D.C.Swartzendruber,and W. H.Murphy. 1969. Detection of virus-likeparticlesingerminalcentersof nor-mal guineapigs. Proc. Soc. Exp. Biol. Med.130:586-590.
12. Melendez, L.V.,R. D.Hunt,M. D.Daniel,B.J.Blake, and F. G. Garcia. 1971. Acute lymphocytic leukemia in owl monkey's inoculated with Herpesvirussaimiri.Science 171: 1161-1163.
13. Middlekamp,J.N.,G.Patrizi,and C.A. Reed. 1967.Light
and electron-microscopic studies oftheguinea pig cyto-megalovirus.J.Ultrastruct.Res.18:85-101.
14. Nadel,E.,W.Banfield,S.Burstein,and A. J.Tousimis. 1967. Virusparticles associated with strain-2guinea pig leukemia
(L2C/N-B).J. Nat.CancerInst.38:979-982.
15. Nayak,D.P.,and M. A. Baluda. 1967. Isolationandpartial
characterizationof nucleicacidofinfluenzavirus.J. Virol.
1:1217-1223.
16. Nazerian,K., andH.G.Purchase. 1970. Combined fluores-cent-antibodyand electronmicroscopy study of Marek's disease virus-infected cell culture. J. Virol.5:79-90. 17. Nazerian,K., andR. L.Witter. 1970. Cell-freetransm;ssion
and in vivoreplicationofMarek's disease virus. J.Virol. 5:388-397.
18. Opler, S. 1968. Newoncogenic virus producingacute
lym-phatic leukemiainguinea pigs. Bibl. Haematol.31:81-88. 19. Opler, S. 1969. Morphology ofcavianleukemia, p. 65-72.
VOL. 8,1971 587
on November 11, 2019 by guest
http://jvi.asm.org/
588
NAYAKNCI Monogr. No. 32. National Institutes of Health, Bethesda, Md.
20. Plummer, G., C. R. Goodheart, D. Henson, and C. P. Bowling. 1969. Acomparative study of the DNA density andbehavior in tissue cultures of fourteen different herpes viruses.Virology 39:134-137.
21. Roizman, B.,andP. R.Roane, Jr. 1964. The multiplication of herpessimplex virus. II.Therelationship between protein
synthesis and the duplication of viral DNA in infected HEp-2 cells. Virology 22:262-269.
22. Sarma, P. S., P. J. Ueberhorst, V. Zeve, H. T. Wepsic, J. Whang-peng, B. Zbar,and R. Huebner. 1970. L2C/N-B guineapig leukemia failure to demonstratetransmissable leukemogenic virus, p. 574-577.In Proc. 4th Int. Symp. Comp.Leukemia Res.,Cherry Hill, N.J. Karger, Basel. 23. Schwartz, J., andB.Roizman. 1969. Similarities and
differ-J. VIROL.
encesin thedevelopment of laboratorystrains and freshly isolatedstrainsofherpessimplex virus in HEp-2 cells: elec-tronmicroscopy.J.Virol.4:879-889.
24. Stackpole, C. W. 1969. Herpes-type virus of the frog renal adenocarcinoma I. Virus development intumortransplants maintained at low temperature. J. Virol. 4:75-93. 25. Stevens,J. G.1966. Selective inhibition of herpes virus DNA
synthesisatelevated temperature. Virology 29:570-578. 26. Wagner, E. K., B. Roizman, T. Savage, and P. G.Spears. 1970.
Characterization of DNA of herpesviruses associated with Lucke's adenocarcinoma of the frog and Burkitt lymphoma ofman.Virology 42:257-261.
27. Wepsic,H.T., B.Zbar, J. Whang-peng,T.Vorsos, andH.J. Rapp. 1970.Guinea pig leukemia L2C: characterization of chromosomes and attemptstodemonstrateantigenicity. J. Nat.Cancer Inst.45:99-105.