Effect of Host Cell Wall Material
on
the
Adsorbability
of
Cofactor-requiring
T41
DENNIS T. BROWN2 AND THOMAS F. ANDERSON
InstituteforCancerResearch, Philadelphia, Pennsylvania 19111
Receivedforpublication 7March 1969
Theadsorbability of T4onhostcells wasdetermined asafunction of timeafter
their liberation from infectedcells. Freshlyliberated (nascent) particles are readily adsorbed but lose theiradsorbabilitywithahalf-time of about 2daysat5C,butonly about 20minat37 C.Theycanbe made adsorbableagain withana-amino acid co-factor likeL-tryptophan, and thisstateofadsorbabilitycanbe stabilizedbycellwall material from Escherichiacoli.Such stabilized particles lose theiradsorbabilityata ratesimilartothatatwhichnascentparticles lose theirs. Mostfreshlyliberated parti-clesareobservedbymeansof electronmicroscopytohave "debris" attachedtotheir
baseplatesandtohavemostof theirsix,long tail fibersfree,whereas "old"particles
that have lost theiradsorbability appear relatively "clean" with mostof their tail fiberswrappedaround their sheaths. Nascentparticleshave densities thatarelower
than those of old particles. The material responsiblefornascentadsorbabilityseems tobeafragmentofthe host's cellwall,fornascentadsorbabilityisdestroyed by
lyso-zyme.Furthermore, nascentT4particles liberated from hostcells withradioactively labeledwallscarrythe label indensity gradientsbut lose itastheyloseadsorbability. Inaddition, onlyasmallproportion of particlesliberated from infectedspheroplasts
arenascentlyadsorbable, whereas mostparticlesliberated from intact cellsare ad-sorbable.
Bacteriophage particles in stocks of certain
strains of T4 and T6 are incapableofadsorbing to host bacteria unless they have been activated
byanadsorption cofactor (2, 3, 4). L-Tryptophan
wasfoundtobe themosteffectivecofactor, being capable ofinducingnearlymaximaladsorbability at concentrations of 2 x 10-6 g/ml.The
activa-tion of T4bycofactor isreversible;after the co-factor is dilutedto nonactivating concentrations, 99 % of tryptophan-induced adsorbability is
lost in 3to 5min.
It has been reported (6, 12, 15, 16) that the effect of the cofactor is to interact with the T4 particle in such a way as to induce a
configura-tional change in its tail fiber apparatus. In the absence of cofactor, cofactor-requiring T4 particles are often observed by means of
elec-tron microscopy to have most of their six tail
fibers lying in close association with the sheath and collar regions of the phage structure. The
same T4 observed in the presence of activating concentrations of tryptophan have mostof their
IPresented inpartby Dennis T. Brown in partial fulfillment oftherequirements foraPh.D. degreeattheUniversityof
Pennsyl-vania.
2Present address: Department ofBiological Sciences,
Dart-mouthCollege, Hanover,N.H. 03775.
tail fibers inanextendedstate,free of thesheath and available for interaction with the host-cell
receptors. It is thought that these changes in tail fiber arrangement can account for the observed change in therateofsedimentationfrom
S2,,
=1,100 for inactive particles that are streamlined
with their tail fibers wrapped around the tailto
S20,
= 850 for tryptophan-activated particleswhose extended tail fibersexertafrictional drag as they move through the medium. Cummings et al. (7) suggested thatalterationsin thesize of the phage head may also contribute to changes inthe rates ofsedimentation.
Gamow and Kozloff (12) postulated that the function of tryptophan in activating cofactor-requiring phage is to combine with a folic
acid-like residue located in the baseplate of the T4
particle. This interaction results in a
configura-tional change in a postulated hinge structure located in the baseplate, thus changing the
orientation of the phage tail fibers.
Even though cofactor-requiring T4 particles were found to be incapable of forming plaques
when plated directly with Escherichia coli strain B on cofactor-free agar, it was noted (3) that
host bacteria that had previously been infected
withcofactor-requiringT4could formplaqueson
94
on November 11, 2019 by guest
http://jvi.asm.org/
CELL WALL MATERIAL
ANDWADSORBABILITY
OF T4bacteria growingon suchmedia. Since the
produc-tion ofa plaque involves many rounds of
infec-tion, reproducinfec-tion, and liberation of phage, it
was thought that the daughter particles must
somehow have been infectious. However, when the progeny phage produced in such plaques were
isolatedandtestedbyplating in the presence and
in the absence ofcofactor, they were found to requirecofactor for adsorption justas the parent
particlehad. Anderson (3) attributed thisrather
surprising resulttothe possibilitythatthe progeny
phage had obtained cofactor from the host cells growing on synthetic agar.
In 1952, Wollman and Stent (25) named this state of adsorbability possessed by the newly
released T4 "nascentactivity" andtermed phage
which possessed thisactivity "nascent" phageas
opposed to "quiescent" or cofactor-requiring
phage. Theydemonstrated that nascent
adsorba-bility was due to an active state which was lost
much more slowly on dilution than
tryptophan-inducedadsorbability andsuggestedthat nascent
activity might be due to a tryptophan-like
co-factor (perhaps tryptophan itself) binding to a
special nascent surface possessed by the
newly
released T4 particles. This nascent surface was
presumed tobind cofactor morefirmly than the
cofactorsitesonquiescent phage,thusaccounting
for the slow loss of nascent adsorbability. The
special surface was supposed to be lost more
slowly than the
tryptophan-like
cofactor andwas
presumed
not tobelost atall solong
asco-factorwasboundtoit.
In1956, Jerne(14) discoveredthatthe
adsorba-bility of
tryptophan-activated
T4 could bestabilized
by
"A8 serum." This was serum thathad beencollected fromahorseonly8days after
the animal had received an
injection
of T4.Although thisearlyserumhad notdeveloped an
appreciable
titer ofneutralizing
antibodies, itpossessedtheremarkablepropertyinthepresence
oftryptophan of
rendering
T4 adsorbable toitshost after the tryptophan had been removed.
Jerne's results
opened
up anotherpossibility:
nascentadsorbability might be dueto asubstance
acquired during its latent
period
that stabilizesthe
phage
in itsadsorbablestate.The
results
reportedinthisstudy
indicatethatthe latter
possibility
is correct in a sense.Nas-cently adsorbableT4 has alowerdensityin CsCl
than do quiescent
particles,
which suggests that thesubstance hasadensitythatis lower than thatofquiescent particles and a volumethat is
rela-tively large. When viewed
by
means ofelectronmicroscopy, quiescent
particles
appear "clean"with their
long
tail fiberswrapped
around theirsheaths, whereas
nascently
adsorbableparticles
are observed to have
particles
of "debris"at-tachedto their baseplateregionswith most of their
long tail fibers free. A number of observations
indicate that the material that is responsible for
nascent adsorbability is derived from fragments ofthe host'scellwall. (i) Nascentadsorbabilityis destroyed by lysozyme. (ii) The adsorbability of tryptophan-activated T4 can be partially stabilized by a cellwallextract from E.coli.(iii) T4 particles that have been freshly liberated from host cells whose cell walls have been labeled with 14C-labeled glucosamine containthis label but lose it as they spontaneously lose adsorbability. (iv) Whereas most of the particles liberated from
whole cellshave nascentadsorbability,very fewof
those liberated from spheroplasts whose cell walls have been largely destroyed by lysozyme
before lysis possess this trait.
MATERIALS AND METHODS
Bacteria and bacteriophage. E. coliB has been
de-scribed (2). The bacteriophage T4 used throughout this study was obtained by isolating a single clear (r-) plaque from Anderson's stock labeled T4 5/10/45 (2).
General methods. The methods used were mainly
those described by Adams (1). F medium contained per liter of distilled water: NH4Cl, 1 g; KH2PO4,
1.5 g; Na2HPO4, 3.5 g; sodium lactate, 10.0 ml of a 60% syrup (Sigma Chemical Co., St. Louis,Mo.);and MgSO4, 0.01 g (added separately). N broth contained perliter ofdistilled water:Bactotryptone, 10 g; yeast extract,5g;NaCl, 10 g; and glucose, 1 g. The pH was
adjustedto 7.0 to 7.2with 1 N NaOH. Agar was added totheseliquid media at final concentrations of 1.0 and 0.6%, respectively,for plates and overlay. To prepare
hightiter stocks, 3 XD medium was usedasdescribed by Fraser (10).
Assaysweremade on N-agartodetermine thetotal titerofasuspensionofcofactor-requiringT4.Assays
onF-agar wereusedto count theparticlesthat were
capableof adsorbingtobacteria in the absence of
co-factor. Thefraction(R)of thephagepopulation capa-ble ofadsorbingin theabsenceof cofactorisexpressed astheratio of the number (F) of plaques obtainedon
F-agar to the number (N) of plaques obtained on
N-agar.
Preparation of E. coli extracts. Crude E. coli
ex-tractswerepreparedfrom 64Fhard-agarplatesthat
had been seeded with E. coli Bandincubated over-night at 37C. The resulting bacterial lawn was har-vestedinto 160 mlofFmedium. The cellsuspension wasthenpelletedinaclinicalcentrifugeatabout5,000
X g for 15 min andresuspendedina10-ml volumeof
Fmedium. This very thicksuspensionwasthentreated threeorfour times inaFrenchpressurecell(American
Instrument Co., Inc., Silver Spring, Md.) at 25,000
psi,being carefultokeepthetemperaturesof both the cell and the treated material below4C. Theresulting
pastewasthentreated withadropletof chloroformto
kill any survivingbacteria. These chloroform-treated cellwallfragmentshad littleabilitytokillT4particles
added later. Theywerequicklyfrozen at -78Cand storedat -20 Cfor later
use-VOL. 4,1969
95
on November 11, 2019 by guest
http://jvi.asm.org/
Electronmicroscopy. Themethods used were gener-ally those of Anderson (5). Preparations were nega-tively stained with sodiumsilicotungstate atpH 7 to increase contrast andresolution. All preparations were studiedbyusingaSiemensElmiskopIelectron micro-scope.
RESULTS
Preparation of nascently adsorbable T4. Woll-man and Stent (25) demonstrated that their phage T4-38 exhibited a nascent state of in-fectivity byassayingthe yieldofphageproduced in a one-step growth experiment and comparing the titers obtained on F and N agar. We have
repeated this experiment with results that agree
with those of Wollman and Stent. During the first 25 min of the latent period before the in-fected cellsbegin to lyse, the assaysonFand N
agar are identical, for each infected bacterium
forms a plaque with equal efficiency on the two
media. After 25 min, however, when the cells
begin to lyse both titers increase, but the titers
on Fagarbegintofall below thoseonNby
ever-increasing amounts; thefreshly liberated particles
slowly lose theabilitytoformplaqueson Fagar
until after 2 hr only 1 particle in 1,000 forms a
plaque on F. If suchlysatesareallowedtostand
at room temperature for a day, the value of R (= Assayon Fagar/AssayonNagar) fallstoas
lowas5 X 10-6.
To study effectively the problem of nascent
adsorbability, itwasnecessary tohavereadily at
hand a stock suspension ofphage known to be
nascently active. Thiswasaccomplished by
cool-ing to0Casuspension ofinfected bacteria inF
medium after only 30 min of aeration.
Chloro-form was then added to induce premature
lysis.
A trace of deoxyribonuclease was also added to
decompose the liberated deoxyribonucleic acid
(DNA) and soreduce the viscosity. After
stand-ingat0Cfor 8to 10min,the titerson Fand N
gaveR values between 0.6 and 1.0. Itwasfound thatthenascent
adsorbability
of thephagein such lysates couldbe preserved indefinitely by adding glycerolto afinalconcentration of 5%and quick-freezing small samples in an acetone-dry-ice bath at -78 C. After storage at -40C for months, these lysates showed no measurablede-creasein R,although someloss in total titerwas observed. Therefore,these preparations provided
areadilyavailable sourceofnascentlyadsorbable
phagefor the studies that followed.
Factors affecting the rate of loss of nascent adsorbability. The initial observation that the nascent stateofadsorbability is lost more slowly
thantryptophan-induced adsorbability prompted
astudyofconditions whichmightbe expected to
change this observed rate. A number of factors
hadnoobservableeffect. The rate wasessentially constant between pH 6 and 9 and in NaCl concentrations between 4 M and 0 (distilled water);nordid thepresence orabsenceofoxygen
orofvisible light affect the rate.
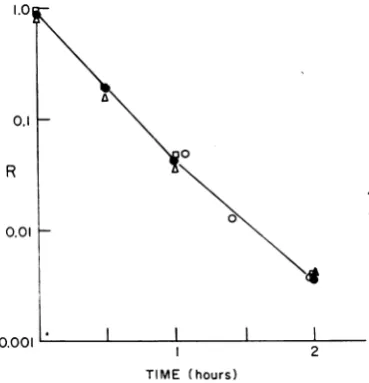
The experiment of Fig. 1 shows that tryptophan (and the other compounds and known cofactors such as phenylalanine and tyrosine in N) have no effect on the loss of nascent adsorbability; nor does the reactivation by tryptophan of a phage preparation which has lost part of its nascent adsorbability affect the rate of loss of adsorba-bility. The reactivated phage return to a state of
adsorbability equal to that of the control and
quickly lose their tryptophan-induced adsorba-bility when the tryptophan is removed. These
0.1
R
0.01
2 TIME (hours)
FIG. 1. Effect oftryptophan andNmediumon the
rate of loss of nascent adsorbability of T4. Samples (0.1 ml) ofafreshly thawednascentphagepreparation
wereaddedtoeachoffour tubescontaining:tube 1, 10 mlofFmedium (a); tube 2, 10mlofFmedium
con-taining 100 ,ug ofL tryptophan/ml (A); tube 3, 10
ml ofNbroth (O); and tube 4, 10 ml ofFmedium
(0). Allsamples wereincubatedat 37 C. At various
times, samples oftubes 1, 2, and 3 were dilutedand
plated with B on F and N. The three samples lost
nascent adsorbability at essentially identical rates,
in--dicating that neither tryptophan alone nor theother
components in N medium affect the rate of loss of
nascenttadsorbability.Thesampleintube 4wasallowed partiallytodeactivateforIhr, at which time tryptophan
wasaddedto makethefinalconcentration 100,ug/ml.
After3 min, thesamplewasdiluted to anonactivating
concentration of tryptophan (0.01 ,ug/ml). After an
additional 5 mintoallowthephage tolose its
trypto-phan-induced activation, the dilutedpreparation was
assayedonFandN.Hereagain,theRvaluesare
iden-ticaltothoseinthe other three tubes.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.491.271.456.248.440.2]WALL MATERIAL AND ADSORBABILITY
resultsdisagree with those of Woilman and Stent
(25) and willbe discussed later.
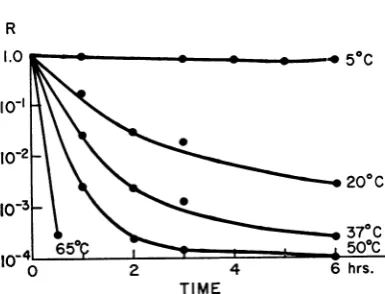
Temperature did affect the rate of loss of nascentadsorbability as shown in the experiment ofFig. 2 in which the ratio R is plotted against the time ofincubation at different temperatures. These results are in good agreement with the
data reported by Wollman and Stent (25) and
approachfirst-order kinetics for the initial period asthoughthereaction
T4(nascently absorbable)
-T4 (nonabsorbable)
were the principal one occurring in this system.
Plotting log
ki
against 1/T, onecanestimate theactivationenergy(Ea)for the reactiontobe11,000
cal.
The kinetic observations indicate a low
tem-perature
dependence
for the conversion of the majorityofthe nascent phageto cofactor-requir-ing phage. On the other hand, a lack ofhomo-geneity inthepopulationcouldgiveanapparent
value ofEa that islowerthanthatforanyof the
particles. Infact, at all the temperatures studied
therateof deactivationdecreasesasR
approaches
itsminimal value of5 X 10-6to5 X 10-A.Some,
but not all, of the particles that lose activation
very slowly prove to be mutants that have
re-duced requirements for cofactor
(4).
Itis important toemphasizethatnascentphage
deactivate much more slowly than
tryptophan-activated phage at each of the temperatures
studied.
Electron microscopy of nascently adsorbable
R
TIME
FIG. 2. Tlte deactivation of nascently adsorbable
T4r- at different temperatures. Samples (I ml) ofa
freshly thawednascentphage suspensionwereaddedto
9-ml volumes ofF medium at the indicated
tempera-turesandwereassayed from time to timeonFandN
agartodeterminethe R valueofthepopulation, which
is shownplotted against time.
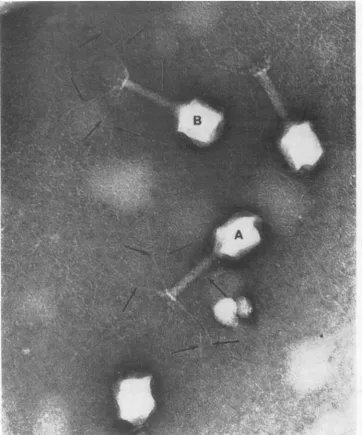
phage. The electron microscope was used to
determine whether nascently adsorbable T4
possess any grossmorphological differencesfrom
cofactor-requiring phage. In the following
ex-periment, nascentphagewereobtained by
allow-ing infected bacteria to lyse on a carbon-coated
electron microscope grid. E. coli B growing in
the log phase in F medium were infected with
tryptophan-activated T4r [multiplicity of
in-fection(MOI) =5].After5minhadbeen allowed
for adsorption, the infected bacteria were
pel-leted. The supernatant fluid containing
unad-sorbed phage plus thecofactorwasdiscarded and thecells wereresuspendedinfreshFmedium. The
resuspended cells were incubated for 20 min at
37C, at which time a small droplet of the
suspension wastransferred to aprepared copper
grid. The grid, held with forceps, wasincubated
foran additional 10 minat 37C, afterwhich it
was washed with F medium and then negatively
stained and placed inthe microscope todry.
Microscopic fields produced by this method
were found to contain many unlysed bacteria
aswell as bacterial debris; however, some areas
suchasthatshown inFig. 3 wererelativelyclean.
In this instance one can see the end of a
bac-terium which has broken open to release its
contained phage onto the grid surface. One is
struckby thefactthatmostof the phagehave all
six of their tail fibers in the extended state. On
two ofthe particles thesix tail fibersare pointed
out with arrows, but many of the other phage
mayalsohave allsixfibers extended. The release
ofallsix tail fibers isgenerallynot seeninpurified
cofactor-requiring phage evenin the presence of
highconcentrationsoftryptophan (6, 15).
Comparison of the adsorption ratesof nascently adsorbable T4 and oftryptophan-activated T4. If
the rate ofadsorption ofa population ofphage
is limited by therate at whichthetail fibers can be made available for interaction with the host receptors, the nascently active phage, having all
tail fibersextended, might be expectedto exhibit
a more rapid adsorption rate than
cofactor-acti-vatedphage.
The rate of adsorption of cofactor-requiring
phage activated by N broth, where its rate is a
maximum(24), wascompared to thatofnascently
adsorbable phage in F medium at 15 and 37C.
Contrary to expectation, the results (Fig. 4), indicate thatnascentT4 inFadsorbsto B
some-what more slowly than does cofactor-activated
T4inN medium.
Data from five such adsorption experiments
were averagedtogivetherate constantsrecorded
in Table 1.Adsorptionrates wereappreciablefor
bothforms ofT4atboth37 and15C. Thisresult
97
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.491.36.229.425.572.2]FIG. 3. Electronmicrogr-aph showinignascentphageproduced by
allowinig
bacteriainfectedwithcofactor-requir-ing T4 tolyseonanelectronmicroscope grid maintained at 37 C. Tailfibers
oni
twoparticlesarepoinited
out witharrows.Stainedwithsilico-tungstate.
differs from the observations of Wollman and Stent (25), who reported that nascent phage
adsorbedveryslowlywitharateonly0.015thatof
the same phage in broth at 15C. It should be
noted that the nascently adsorbable phage in F
medium appear to adsorb at only a slightly slower rate than do
cofactor-requiring
phage in broth. On the one hand, this difference in ratesmay be due to some effect other than available
cofactor; for instance, the different nutrients
may have different effects on the bacterial sur-faces. On the other hand, the lower apparent
adsorptionratefor thenascentphage may bedue
toitsprogressive lossof
adsorbability
duringtheexperiment,whereasthecofactor-requiringphage
in broth have sufficient availablecofactor to
on November 11, 2019 by guest
http://jvi.asm.org/
[image:5.491.81.446.69.505.2]T4
1.0o 0.8
0.6
0.4
0.2
0o
0
}-
W--'*I
37°C;////
}=-°
150C/
I
5 10
[image:6.491.35.226.45.193.2]TIME (MIN)
FIG. 4. Comparison of the rates ofadsorption of
nascent T4r- andof N broth-activated
cofactor-requir-ing T4r- at 15and 37 C. Cofactor-requiring T4r- was dilutedintoNbroth. Nascent T4r-fromafrozen
prep-aration wasdilutedinto Fmedium.At timezero,a
vol-umeofE. coliB wasaddedtoeach tube tomake their
finalconcentration 2 X 108 cells/ml and that of the phage S X 107 particles/ml. The reaction mixtures
wereincubated at either 37or15 C. At various times, samples of thereactionmixture werediluted1/100into
medium containing anti-T4 serum at a concentration
sufficient to inactivate99.9% oftheunadsorbedphage in 2min. Theinfectedbacteria were thenplatedonN agar. Solid line indicates T4 activated with nutrient
broth;broken line indicatesnascentlyactiveT4.
TABLE 1. Velocity constantsfor the adsorption of
nascent T4andcofactor-activatedT4 at 15and
37 Ca
Temp Cofactor-requiring Nascently-activephage phage in broth inFmedium
15C (2.3 7 0.5) X 10-10 (1.9 i 0.9) X 10-10
37C (12.5 4 0.3) X 10-10 (8.2 4 0.5) X 10-10
aValues areexpressed as milliliters perminute.
sure thatthey will all be adsorbable throughout the experiment. It was concluded from these
experiments that nascently adsorbable phage possess an ability to adsorb to bacteria that is
nearly equivalent to that of cofactor-activated phage.
Retention of nascent adsorbability by intra-cellular phage. The experiments described above
have been confined to free phage that had been released from the hostcellatthe end of thenormal
30-min latent period. The experiment of Fig. 5 wascarriedout todetermine whether thenascent
state of adsorbability can be retained by the
phage inside the host cell beyond the normal
latent period. If the nascent adsorbability were dueto some special nascent cofactor which was
incapableofdiffusingout of the cell, one might
expectthephagetoretain theirstate of
adsorba-bility throughout an extended latent period. If,
[image:6.491.239.434.66.171.2]TIME AFTER INFECTION(hrs.)
FIG. 5. Effect ofthe extended latent periodon the
nascent stateofadsorbability.E.coliB,rapidlygrowing in F medium, were infected with cofactor-activated T4r-, centrifuged, and resuspendedas previously de-scribed. After incubationat37Cfor26min,KCNwas
addedto themixture toafinalconcentration of6 X
10-' mtoprevent spontaneouslysis. The suspensionof infectedcells wasstoredat 37C. At various time
in-tervals, samples of this suspension were lysed with chloroformat0C andwereassayedonFandNagar.
The assay dilution tubes were then returned to 37 C andreplated from timetotimetofollow thecourse of thephageactivityafter thedelayedlysis. The topline shows theR valueof thephageat thetimeof induced
lysis(indicated by arrows).The descending lines
repre-sentthedecreasein Rwith timeat37 Cafter the
in-ducedlysis.
on the other hand, nascent adsorbability were due to a diffusable cofactor or to some time-labile configurational difference at the cofactor sites, one would expect the phage to lose its
nascent adsorbability and be released from the
cellinareduced stateofadsorbability.
The results of such an experiment are shown inFig. 5, in whichRisplotted against time after infection. It is shown that phage released from thehost cell upto 12 hrafter the normal latent
period possess nascent adsorbability and that this adsorbability is lost only after the phage is
released from the host cell. The fact that phage retained thenascent stateofadsorbabilitysolong as they remained inside the host cell was inter-preted to support the notion that nascent ad-sorbability is due to some special "nascent
co-factor" that cannot diffuse from the cell.
Nascent cofactor from bacterial cells. An
at-tempt to find thenascentcofactor in extracts of
uninfected E. coliBthathad beengrownin F is describedin Fig. 6. Evidently, something similar
tothenascentcofactorcanbe obtained from the
uninfected E. coli cell. The activating ability of
this nascent cofactor is considered to be rather
low, for6 hrwasrequiredtoreactivate thephage from R = 10-a toR = 10-3, whereas the same
phage had been released withR = 1.0atthe end
ofa normal 30-min latentperiod.
A situation similarto thiswas found by Jerne a
w
cs:
m
0
Io
z
0
Li.
99
on November 11, 2019 by guest
http://jvi.asm.org/
[image:6.491.36.228.349.432.2]1.0r
o0-1
10-2
,o--0 -° -=
-I\ o tc
1o a
10-3['
I ~~~~~~~~~~~~~b
10-4 ta
015l & I I
2 3 4 5 6
TIME (hours)
FIG. 6. Reactivation of quiescentT4r- by F-grown
E. coli extract. T4r- (R = 5 X 10-5) wasaddedto
1-ml samples of E. coli homogenatethat had been pre-paredasdescribed in MaterialsandMethods. The re-sulting mixture was incubated at 37C, and samples
weredilutedfrom timeto timeandplatedonFand N
with the result shown incurvea. Onlya slight
activa-tion isproduced bytheextractina6-hrperiod.Oneof
thedilution tubes takenfromthisreactionmixturewas
allowed tostandat37Candsampleswerereplatedat
various intervals onF and N. The inducedactivity is
lostatarate (curve b)that is much slower thanwould
have been expected for tryptophan-induced activity.
Curve c shows the results ofa similar experiment
in which activation by E. coli extract occurred in
thepresenceof50 jAg of L-tryptophan permland in
which samples were diluted 1/1,000at various times
intocofactor-free F medium. They wereassayedonF
and Nafterincubationfor5 min hadbeenallowedat
37 C to eliminate any tryptophan-inducedactivation. The1/1,000dilution tube taken at2hrwasincubated
furtherat37C andassayed fromtimetotimeonFand
Nagarstogivethe resultsplottedincurved. L-Trypto-phan greatly increases therateofactivationbythe E.
coli extractandtheactivatedphagelosesits
adsorba-bilityatthe slowratethat is characteristicof nascently adsorbable phage.
(14) in his studies ofa phage-specific antiserum
produced during the early period of immuniza-tion of a horse against T4. This antibody was found to activate T4 permanently, but at a rate
that was very slow unless the phage had been activated by tryptophan. We have found (Fig. 6, curve c) that tryptophan similarly enhances
therateof activation of T4byE.coliextractsand
that, when the tryptophan is diluted out, this
adsorbability is lost slowly at a rate equivalent tothat ofnormally producednascentphage (Fig. 6, curve d). It is also shown that the activated
phageseemto retain thisactivitysolongasthey remain in the E. coli paste but the maximal observed activation represents only about 5%
of the total phage population. This could well
reflect competition between the activation
reac-tion and the spontaneous loss ofrenascent ad-sorbability. No appreciable loss of phage titer was observed during this experiment, which is
possibly a result ofpretreatingthe extracts with
chloroform.
Attempts weremadeto increase theactivating properties ofthe E. coli extractbyuse of various purification procedures. The extract was
centri-fugedat5C for 10 min at about 6,000 X gina
clinical centrifuge. When the supernatant fluid and theresuspended pelletweretested for
activat-ingabilityin the presenceof50 ,gof tryptophan
perml,the supernatantfluidwasfoundtopossess no detectable activating material, whereas the
pellet was found to induce adsorbability with
about the sameefficiency asdid theoriginal
ma-terial.Attempts to remove theactivating material
from the whole paste by exhaustive dialysis at
5Cagainst F medium failed.
The above results indicate that the nascent
cofactor is associated with the larger
faster-sedimenting portions of the bacterial paste,
possibly fragments of thecell
envelope.
Since E. coli cell walls possess substrate sites
for theenzymelysozyme, thisenzymewastested
for its effect on the
activating
properties of theE. coli extract. The resuspended pellet ofa 10 min, 6,000 rev/min centrifugation of crude E.
coli extract was treated with lysozyme at 3 x
10-5g/mland allowedtoincubateat37Cfor 10
min. Cofactor-requiring T4r- were then treated
withthismaterial for 50 minat37C in the
pres-ence of 50 ,tg of tryptophan per ml. A control
containing pelleted material had undergone a
similar incubation procedure, but it was not
treatedwithlysozyme.
The results (Table 2) indicate that lysozyme
destroys the activating ability of this material.
Other enzymesweretested for theireffecton the
activating properties ofthispellet fraction, with
uniformly
negative results.Effect of enzymes on the deactivation rate of nascently adsorbable T4.The
experiments
already
described indicate that nascent adsorbability is
due to a special nascent cofactor. To further
characterizethe chemicalnature of thiscofactor,
the deactivation kinetics ofnascent phage were
studied in the presence of various enzymes. A suspension of freshly thawed nascent T4r- was treated with the enzyme to be studied and was
incubated at37C. Samples of this reaction
mix-ture and a controlwithout enzyme were diluted
at intervals andplated on F and N media. The enzymes studied included Pronase (5 x 10-4 g/ml), trypsin (2.5 X 10-3 g/ml), deoxyribo-nuclease (1.0 X 10-3g/ml), ribonuclease (1.0 x
on November 11, 2019 by guest
http://jvi.asm.org/
[image:7.491.61.250.48.225.2]CELL WALL MATERIAL AND ADSORBABILITY OF
10-3 g/ml), phospholipase C (8.0 x 10-4 g/ml),
and lysozyme (at 2.1 X 10-4 and 8.8 x 10-3
g/ml).
Lysozymealonewasfoundtoincrease therate ofdeactivation withaconcentrational dependence
that is shown in Fig. 7. Lysozyme's activity is
probably not due to its high net-positive charge
(isoelectric point = pH 11) causing it to react
nonspecifically with the phage particles, for trypsin with essentially thesameisoelectric point
atpH 10.8 (18) hadnoeffect. On the other hand,
the rate of deactivation is not proportional to lysozyme concentration and was much slower
than might have been expected for lysozyme act-ingon a readily available substrate. Itmay well
be that the lysozyme-sensitive site is associated with some other type of macromolecule on the
phage particle that limits its accessibility to the
TABLE 2. Effect of lysozyme on the activating
properties of E. coli extract
Material tested pRvdlued4
T4r- + 50S g of tryptophan per
(no extract)... T4r- + 50SAg of tryptophan per + extract...
T4r- + 50 jAg of tryptophan per + lysozyme-treated extract (3 105goflysozymeperml,50min 37C)...
aMeasured after dilution to
tryptophanconcentration.
1.0
0.1
R
0.01
0.001
b
I
I
jC I
20 40
[image:8.491.246.438.207.323.2]TIME
(min)
FIG. 7. Effect oflysozyme on the
nascent adsorbability. Freshly thawe nascentphage wasallowedto incuba
(a) andinthepresenceof2.1 X
10-perml(b)and 8.8X 10-3goflysozyn
various times, the threepreparations
F and N agar and the R value ofi
against incubationt time.
ml
*-- 5 X 10-5
- 1
enzyme. At any rate, it would appear that the nascent cofactor contains material that is
sus-ceptible to lysozyme.
Buoyant density of nascently adsorbableT4.To determinehow muchof the relatively low-density
cellwallmaterial is boundtonascentT4,wehave
compared its density in CsCl with that of T4
from the same preparation that had been
incu-batedat37 Cto acquire the needfora cofactor. The cofactor-requiring particles (R = 10-4)
band in peaks at p = 1.49 and p = 1.51 g/ml
(Fig. 8). In contrast, the nascent particles (R =
1.60 L
1.50
*T4r-R 10-4 oT4-R .8
--o."~!t 1
so ~:~o---f-Q
0.0.
.AI 0
6 12 18
FRACTION NUMBER
24
LO
.8
.6 0m260
.4
.2
30 36
ml FIG. 8. Densities in CsCl of nascently adsorbable
3 X
10j
T4r- and of cofactor-requiring T4r- from the sameml preparation. Nascent phage were prepared by the
X technique ofone-stepgrowth;30ml ofE.coliB,rapidly
at growing in3 XDmediumataconcentrationofSX 108
* X 10- cells/ml, were infected (MOI = 5) with cofactor-re-nonactivating quiringT4r-inthepresenceof 10-4gofadded trypto-phanperml.After5 minat37 C had beenallowed for adsorption, the infected bacteria werepelletedand
re-suspendedin 10 ml of fresh 3 XD. The infectedcells
wereincubatedwith aerationforan additional 26 min
and then were quickly cooled in an ice-salt bath. Chloroform was added to lyse the cells and a trace
of deoxyribonuclease was added to reduce the
vis-cosity of the mixture. After 10 min at 0 C had
been allowedtoobtain maximallysis, themixturewas
divided intotwo5-mlportions.One samplewasallowed
to remainat 0C; the otherwasincubatedat37 Cfor
5hr. The R valueofthe material storedat0Cwasthen foundtobe close to 1.0,whereas thatofthe37C
ma-terialwas10-4. CsCIwasdissolved in thelysates to a
concentration of0.715 g/ml. The samples were
cen-trifugedinlusteroid tubesat39,000to 40,000rev/min
for18 hrat3to4 C(SpincomodelL-4,SW50 rotor).
60 80 Fractions were collecteddropwise andused to
deter-mine OD260 (optical densityat 260nm) profiles,
via-bility profiles, andindices of refraction. If viability
? rate ofloss of assays were tobedone, theentirecollection, dilution,
d preparation of and plating procedure (toadditionof agar) wascarried
ite at 37 C alone out at5Cto reduce the lossofnascentadsorbability.
4 goflysozyme The secondsamplewasgivenalow-speed centrifugation neperml(c). At to remove some ofthefast-sedimentingdebris. It was
were assayed onz allowedto standfor24 hrat37Cto losenascent ad-each wasplotted sorbability. Figure10b shows the sedimentationprofile
ofa10-p.liter portion ofthissample.
101
on November 11, 2019 by guest
http://jvi.asm.org/
[image:8.491.39.234.252.568.2]0.8) evidently have lower densities, for they distribute themselves overa broadregion whose
densityrangesfrom 1.46to 1.50g/ml. Assays of the phage in various fractionsindicated that the R value is about the same at all densities; ap-parently the activatingnascent cofactor material comes inavarietyof sizes.
Electron microscopy of nascent T4. If the
nascentcofactor iscapable of shifting the density of phage particles as much as we observe, it should be possible to see the attached material byelectron microscopy. Freshlyprepared lysates
ofnascentphagewerefoundtocontaintoomuch
bacterial debris to give unequivocal results. Therefore, freshlyprepared lysates werepurified by several rounds of alternate high- and low-speed centrifugation at low temperature. Ap-proximately 40to 60% of the phage population
lost nascent adsorbability during this procedure.
Figure 9 shows phageparticles from several such preparations. Material can be seen attached to thebaseplate regions ofmanyparticles. This ma-terial isseentovaryinamountfrom rather large pieces on someparticles to small, hardly visible fragmentsonthe tail pins of others. This observa-tion agrees with the density-gradient studies in indicating that thematerial bound to the phage has a wide range of sizes. Thearrow in Fig. 9a
points out a piece of free material identical in
size andappearancetothat attachedtothe nearby phage particle. This fragment maybe a pieceof
material which had been liberated from the tail of a phage particle which has lost its nascent
adsorbability.
Binding of cell wallfragmentsto nascentphage.
Thatnascentadsorbabilityis relatedtothe
inter-action of phage with host cell wallswas further
demonstrated by the experiments presented in Fig. 10 and 11.
Since it isknown thatN-acetylglucosamine is aprimary constituent of theE.coli "rigid layer" (19, 20), E. coli B cells were grown in 14C N-acetyl glucosamine to make their rigid layers radioactive. Nascent phage produced from in-fection of these hot cells were sedimented on
sucrose gradients (Fig. 10) and banded on cell
density gradients (Fig. 11) before and after losingnascentadsorbability.
As isshown in Fig. 10a, phage havingnascent adsorbability are able to affect the sedimenta-tion profile of 'IC-labeled fragments of the
bac-terial wall. Cofactor-requiring phage on the
r)ther hand do not (Fig. 10b).
When nascent phage prepared from
"4C-labeled cellsare banded on CsCl (Fig. 11), the
phage having nascent adsorbability are seen to bandpolydispersely in the gradient (see also Fig. 8). The 14C isseentoband in regions
correspond-ing tothe infectivity peaks (Fig. Ila). It is also
found that the ratio of counts 'IC to infectivity
is higher for those phage taken from the less
dense regionofthegradient, indicatingthatphage
bandingintheseregionshavelarger fragments of
cell wall material boundto them. The
cofactor-requiring phage(R = 1 X 10-s) inFig. lib are
found tobandin a morerestricted region of the
gradient and have less of the label associated withthem than do the nascentphageinFig. 1 la. Adsorbability of phage produced from
sphero-plasts.The
preceding
studies have indicated thatnascent adsorbability is the result of the
inter-action ofT4 phage with host cell wall material
atsometimeduringthelatentperiod.Ifthis is so,
it should be possible to produce
tryptophan-requiring phage directly from spheroplasts,
cells whichhavehadmostof theirwalls removed
enzymatically (11).
E. coli B cellswere
infected
with T4r- in thepresence oftryptophan (MOI = 10). A sample
ofthecellswasconvertedtospheroplastsin
16%
sucrose by a modification of the method of
Guthrie and Sinsheimer (13). After incubation
for 30 min to allow development of daughter
phage particles, the spheroplasts were diluted
1:100 at 0 Cin Fmediumtorupturethem and
releasethe daughter particles. Only 16% of the
phage produced by spheroplasts were able to
form plaqueson Fmedium,whereas 82% ofthe
particles produced by control cells formedplaques
on F medium (Table 3). The simplest
explana-tion is that theremoval of the cell wall
prior
tothe release ofprogeny T4preventsmost
daughter
particles from becoming
nascently
adsorbable.Theequivalence in burst size in
experiment
andcontrol indicates that thepresenceof thecellwall
itself isprobably unimportant for phage
produc-tion.
DISCUSSION
Particles ofT4 are liberated fromhostcells in
a state of nascent adsorbability in which they
canreadily adsorbtohostcellswithout additional
cofactor (25). Once liberated into the medium, however, the particles spontaneously lose this nascentadsorbabilitywith ahalf-time ofonly 20 min at 37 C.Thus, particlesinolder stocksor in
preparationsthat havebeenpurifiedin the course
ofafewhourslose theiradsorbability. They
can-not adsorb to host cells unless they have been
reactivated by a cofactor like L-tryptophan.
Activation by tryptophan is reversible; it is so
rapid that half-times oftheforward and reverse
reactionsaremeasuredinseconds.
Ourresultsalso indicatethatnascent
adsorba-bilityis due to a substance we shall call"nascent
cofactor" that is attached to the phage particle.
on November 11, 2019 by guest
http://jvi.asm.org/
a
I.
b
.us1.C'h:t.X
FIG. 9. Electron micrographs ofnascentT4. Phagewereprepared by lysing infected bacteriaat28 minafter
inzfectionbychloroformtreatment.Thesenascently adsorbable particles were purified by several rounds ofhigh- and
low-speedcentrifugation carriedout at 4 to5C. The arrowspointoutmaterial attachedtothebaseplate regions of the particles. InFig. 9a, the arrowpointsout afragment equalin sizeandshapetotheoneattachedtothtephage; this fragment may havebeenreleasedbyaparticle which has lost adsorbability.Barsrepresent100 nm.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:10.491.44.450.67.570.2]Thenascent cofactor is evidently solarge that it
cannot diffuse from infected cells, for if their
lysis is delayed in KCN,daughter phage particles
canremain trappedinside for days without losing
their nascent adsorbability. Furthermore, the
N~
2
Q-U
35 40
15-b
L
0
5c
5 10 15 20 25 30 35 40
[image:11.491.160.442.60.379.2]FRACTION NUMBER
FIG. 10. Sedimentation profile ofnascent (a) and
cofactor-requiring phage (b) prepared fromcells which
have their "rigid layers" labeled with 14C. E. coli B
were growntoaconcentrationofS X 108cells/ml in
F medium andthenwerediluted into 5 mlofF medium
containing 0.05 mc of N-acetyl-'4C-D-glucosamine
(specificactivity, 10.4 mc/mM). The initial
concentra-tionofcells in the hot medium was 107/ml. After the
cells had been allowedto grow to S X 108/ml, they
were washed three times in coldmedium. These cells werenextinfected withcofactor-requiringT4atMOI=
10 in thepresenceof50 ,ug of tryptophanperml. The
infectedcells were centrifugedandresuspended in co-factor-free medium toafinalconcentration of2.5 X 101in2ml ofFmedium.Thismixturewasincubatedat
37 Cfor30 mininamicro-Fernbachflask.Atthe end
ofthe incubationperiod, themixture wascooledinan
icebath and treatedwith 0.2 mlof chloroform for 10
min. Twosampleswerecollected. One10-,lAitersample
containingnascentlyadsorbableT4waslayered quickly
ontoanice-cold5to30%sucrosegradient andspuI at
20,000rev/minfor10 min;equal-volumefractionswere collected, assayedfor infectivity, and countedfor14C content. The results ofthis experiment are slhown in
Fig. 1Oa.
15
-~lo
0
n
a.
-5 10
---10
L)
5'
0.
5 10 15 20 25 30 35 40
FRACTION NUMBER
FIG. 11. Densityin CsCl ofnascentT4 (a) andof
cofactor-requiring T4 (b) produced from cells which
have theirrigid layerslabeled with14C.Bacteriophage
werepreparedasdescribed inFig. 10. Onesamplewas
maintainedat0C toprevent loss ofadsorbability. A
secondwasplacedat37Cforaperiod of2 hr. CsCI
wasaddedtoboth whileat 0 Cto raise thedensity to
about 1.475g/cm3. Thesesampleswerecentrifugedin
theSW50-1rotorat35,000
revlmin
for18 hrat2C.Titer andcounts/minareshownplotted against fraction
numberfor nascentphage (a) and cofactor-requiring
phage (b).
mean buoyant density of nascently adsorbable T4 particles in CsCl is considerably lower than that ofparticles that have been allowed to lose
their nascent adsorbability. This indicates not
onlythatthe nascent cofactor islarge enoughto
affect the overalldensityofaT4particlebut also
that the nascent cofactor has a density that is
lower than that ofparticles that have been freed
of nascent cofactor. The fact that nascent
parti-cles display a considerable range of densities indicates that the nascent cofactor can have a widerangeofsizes. It is evident that this cofactor is not a discrete molecule but a substance or
aggregation of substances. The electron
micro-scope confirms this view: whereas particles in
on November 11, 2019 by guest
http://jvi.asm.org/
[image:11.491.57.249.135.414.2]CELL WALL MATERIAL
TABLE 3. Adsorbability of phage newly released from spheroplasts
Expt
~~Total
phage RvalueExpt produced
Controla 6 X 109 0.82
Experimentalb 4 X 109 0.16
aInfected cells suspended in F medium +
16%
sucrose.
6 Infected cells suspended in F medium + 16%
sucrose + 104 g oflysozyme/mi + 10-3M ethyl-enediaminetetraacetate.
"purified preparations" that have lost their
nas-cent adsorbability appear relatively "clean,"
nascently adsorbableT4particlesare seentohave
pieces of material of various sizes adhering to
their baseplate regions. This could well be the
nascent cofactor.
By means of electron microscopy, it is also
observedthat T4
particles
inpurifiedpreparations
often have their
long
tail fiberswrapped
aroundtheir tailsheaths, whereasnascentparticles have
all six long tail fibers freed from their sheaths,
withtheirtips freetocontactthesurfaces ofhost
cellsand initiatetheadsorption process (15, 22).
L-Tryptophan
also frees long tail fibers fromthesheathsofquiescentparticlestoactivatethem,but
oddly enough, even in a large excess of
trypto-phan, very few particles have all of their tail
fibers visibly free of their sheaths (6, 15). Thus,
it seems highly
probable
that thisfreeing ofthe tail fibers is thesignificant
morphologicalcon-figurationthatenablesT4particlestobe
adsorba-ble,nascentlyorotherwise.
Therateof
adsorption
ofnascentT4particlesis nearlyequal tothat oftryptophan-reactivated
particles. One
might
suppose that the presenceofwhat lookslikebacterial wall debrisassociated
with the
baseplates
of nascent particles wouldinterfere with the
complex
function of thisstruc-ture (22), but thismaterial
might
readily mergewith compatible substances endogenous to the
bacterial surface when the baseplate region is
brought intocontactwiththe hostcellwall. If so,
this material would
quickly
be prevented frominterfering
with the function of the baseplate, and the rate of infection would scarcely be affected.We have found that the cell wall fraction of
E. colican conferan active "renascent" state of
adsorbabilitytomany,butnottoall,T4particles
that havelosttheirnascent
adsorbability
onstand-ing.Two observationssuggest that thiswall
ma-terialis very similar ifnotidenticaltothenascent
cofactor. First, both the nascent
adsorbability
of the phage and the
activating ability
of theextracts are destroyed by lysozyme. Second, the renascent particles spontaneously lose their
adsorbabilityatmuch the same rate as the nascent
particles lose theirs.
Tryptophan greatly enhances the rate and ex-tent to which the cell wallfraction activates T4, as though tryptophan prepared the phage for reaction with the wall fraction by extending its
tail fibers. Thewallmaterial might thenlockthe
particleinthis adsorbable configuration inwhich
it canremain evenafterthe tryptophan has been removed. It seems probable that here tryptophan actsinmuch the same way asit does in promoting the permanentactivationof T4 by A8 serum(14). It would appear that cofactors like tryptophan reversibly activate T4 particles by extending their long tail fibers, whereas substances likethe
nas-centcofactorand A8 serum stabilize theparticles
inthis adsorbable configuration.
The susceptibility of nascent adsorbability of
T4 to egg-white lysozyme suggests that,6-1-4
bonds between N-acetyl-glucosamine residues
are essential to the activity of the nascent
co-factor. These bonds probably are not readily
accessible to the enzyme, however, for the rate
of lysozyme action is not
proportional
to itsconcentration. Nascent phage produced from
cells that had been prelabeled with N-acetyl-l"C
glucosamine are found to exhibit a variety of
densities thatis paralleled by the density profile
of this radioactive material. These phage also
showalargeramountof label associatedwiththe
less-denseparticlesthanwith thephageofgreater
density.
Sincetheradioactive labelintheseexperiments
isincorporated intotherigid layerofthecell wall
andsince theloss ofnascent adsorbability
coin-cideswithlossof label fromnewlyreleasedphage,
it may be concluded that the nascent cofactor
contains (in part at least) some of the host cell
wallmucopolymer layer.
In light ofthe evidence presented here, it is
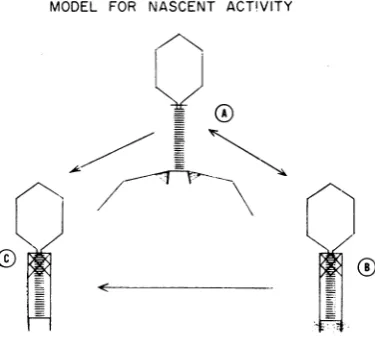
possible to present a reasonable model for the
nascent state ofadsorbability. Since the nascent
cofactor is an activity stabilizer rather than a
phage activator, one can assume that this
co-factorisbound tothe phage during thetime that
the tail fibersareintheextendedstate.
Electron microscopy indicates that the cell
wall material may be attached to the baseplate
and tail-pin region of the phage structure. The
baseplate of the phage may acquire cell envelope materialasit isformed within thecelland
some-time during thelatent periodeither prior to the
synthesis oftail fiberorbefore thetail fibers are
retracted. On the otherhand,thebaseplateofthe finishedparticle may associate witha portion of the cell wall of the infected bacteria. Since the
VOL. 4, 1969
105
on November 11, 2019 by guest
http://jvi.asm.org/
[image:12.491.45.239.69.166.2]cell membrane is interposed between the cyto-plasm and the cell wall proper, one must either
include the membrane in thefragment boundto
the phage or assume that the cell membrane has
been broken downatthis point,making possible
directaccess tothecellwall.
The location of the nascent cofactor on the
underside of the baseplate may account for its
abilityto preventthe retraction of the tail fibers.
Location ofthehinge (12, 17) which allowsthe tailfibers tochangeorientation isnotknownbut
couldwell be in the area where thebaseplate is
attached to the tail fiber. The cell wall material bound to theunderside ofthebaseplate may act insuch a way as to preventthishinge from work-ing, thus creating arigid tail fiber-extended
con-figuration. This might be accomplished by
link-ingthetail fiber to thetail pin. Theloss of
nas-centadsorbability would betheresultofremoval
ofthe cell debrisfrom the baseplate, freeingthe
hinge and allowing the tail fibers to bindto the
sheath. Theforcesremoving thenascentcofactor
from the phage may
simply
be due to kineticforces acting on the rigid tail fibers, eventually
freeing the hinge. The essential aspects of this
model are depicted
diagrammatically
in Fig. 12.ParticleA representsaT4particle in thenascent
state.Thenascentcofactoris showncross-linking the tail pins to the tail fibers, maintaining them in an extended state. T4 may lose nascent
ad-sorbabilityin two ways. (i) Thenascentcofactor
may be removed fromthebaseplate regionasin
particle C, allowing the retraction of the tail
fibers, or (ii) the tail fibers may free themselves
from the cofactor and retract, leaving the
co-factor attached to thebaseplate asin particle B.
MODEL FOR NASCENT ACT!VITY
0'
FIG. 12. Modelfornascentadsorbability. (See text
forexplanation.)
The reaction A -* B may be
reversible, allowing
the
phage
toreturntothenascent state ifthe tail fibersarereassociated with the cofactor afterdis-placement by
some phage activator. [This mayaccount for the return to the nascent state after
treatmentwithtryptophaninthe system of Woll-man and Stent
(25).]
Thereaction A -* Cwouldnotbereversible,asthecofactor is gone from the
baseplate. It would also be possible for particles
in the B state to convert to the C state, losing theirability to revertbackto the A state.
Thus,
anoverall reaction
might
beA
(adsorbable)
* B(nonadsorbable
butcapable
of
reverting)
->C(nonadsorbable)
The reaction A
'->
B -÷ C may accountfor theshape ofthecurves obtained from the studies of
therateofloss ofadsorbabilityofnascent
phage.
Thedeactivationcurve (Fig. 2) ofT4r- seemsto
possessaninitialregionoflinearity,followedbya decrease in deactivation rate prior to obtaining the lowest observed R value. The low R value observed for T4r-wasfound to be R = 5 X 10-6. This
change
inratemaybethe resultofkB
-cbeing
less thankAB-Thereactivation ofphage by the cell wall frac-tion of E. coliextractsis presumedto follow the modeljust described, with theexception that the tailfibersarefirstextended bytryptophan. If the
nascent cofactor is made up of sections of the cell wall consisting of protoplasmic membrane and complete multilayered wall, increase in the deactivationratebylysozyme may be by removal of some of the cell wall layers, leaving behind the membrane and other protein-containing
layers.Under thesecircumstances, the rate of loss
of nascent adsorbability may not reflect the
di-gestion by lysozyme, as there would still be
lysozyme-insensitive material to be removed. The spontaneous loss of nascentadsorbability by free phage could be dueto a simple dissocia-tion ofnascentcofactor from thephage. On the other hand, it is conceivable that spontaneous deactivation is duetothe action of the samelytic enzymes in T4 that hydrolyze cell wall material when T4 is adsorbed to host cells (19, 21, 23). In the latter case, to account for the intracellular retention of nascent activity, one would haveto
assume that these enzymes areeither blocked or cannotreach their substratesduring an extended intracellularsojourn of the virion.
Three possible models may be suggested to explain how phage obtain these cell wall frag-ments. (i) Phage particles maturate with their baseplates in association with the cell envelope. (ii) At some timeduring the latentperiod,phage attach by their baseplates to the cell envelope.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:13.491.60.248.447.616.2]CELL WALL MATERIAL AND ADSORBABILITY OF
(This attachment need not be permanent and at any given time only a few of the intracellular phage particles may actually be in contact with the wall structure.) (iii) Fragments of wall
ma-terial might be obtained during the physical
breakdown of the cell envelope (lysis).
Theplausibility of the third model is strength-ened by observations of T4-infected bacteria in
V.~~~~~~~~~~~
$,J
FIG. 13. Electron micrograph ofabacterium infectedwith cofactor-requiring T4 in the process oflysis. The
arrowpointsout amembrane-like structurewhich seems tosurroundthenewly released particles. Thismembranze seems tobeconitinuouswith the bacterial cell wall.
VOL. 4,1969
107
on November 11, 2019 by guest
http://jvi.asm.org/
[image:14.491.36.442.109.609.2]the process of lysis. By briefly exposing infected
bacteria to chloroform vapor on an electron
microscope grid, cells can be observed in the
process of releasing their contained phage. In
Fig. 13, thewall structure ofa bacterium seems
to be broken down insomeareaswhereprogeny
phageare seenspilling out, although outside the
normal confines of the host cell some of the
progenyparticlesseemtobe retained by a
mem-brane-like structure which is continuous withthe
bacterial envelope. (This structure may consist
of the lipid layers of the envelope.) It may be
that the phage obtain the fragments of the rigid layer while retained inside this baglike structure duringa temporary state of partial lysis. It thus seems possible that, much as certain animal
viruses acquire their envelopes from nuclear (8)
or cytoplasmic (9) membranes, the T4 phage
particle acquires its nascent cofactor from cell
wall material at the time of its release from the cell.
ACKNOWLEDGMENTS
Weacknowledge the excellent technical assistance of John M. Mackenzie, Jr. Manfred Bayer contributedmanyhelpful sugges-tions in thepreparation ofthismanuscript.
This investigation was supported by Public Health Service traininggrant5-TOIGM-00694-07totheUniversityof Pennsyl-vania from the National Institute of General Medical Sciences, by Public Health Service grants FR-05539 and CA-06927 to the Institute for Cancer Research, bygrantGB-4640fromthe Na-tional ScienceFoundation,andbyanaward toDartmouth Col-lege from the Brown-Hazen Fund administered by the Research Corporation ofNew York. Anappropriationfromthe Common-wealth of Pennsylvania totheInstituteforCancer Researchis
gratefully acknowledged.
LITERATURE CITED
1. Adams,M.H. 1959.Bacteriophages. Interscience Publishers, Inc., New York.
2. Anderson,T. F. 1945. The role oftryptophaninthe adsorp-tion oftwobacterial virusesontheir host E.coli.J.Cell. Comp.Physiol.25:17-26.
3.Anderson, T. F. 1948. Theactivation of thebacterial virus T4by L-tryptophan.J.Bacteriol. 55:637-649.
4. Anderson, T. F. 1948. Theinheritance ofrequirements for adsorptioncofactors in the bacterial virus T4.J. Bacteriol. 55:651-658.
5.Anderson, T. F.1962.Negative stainingand itsusein thestudy of virusesand theirserological reactions,p.251-262.InR. J. C. Harris, (ed.), The interpretation of ultrastructure, vol. 1. Symp. Int. Soc. Cell Biol. Academic Press, Inc., New York.
6. Cummings, D. J. 1964. Sedimentation and biological proper-ties of T phages ofEscherichia coli.Virology23:408-418. 7. Cummings, D. J., V. A. Chapman, and S. S. DeLong. 1969.
The sedimentation andconformational varianceamong T-evenbacteriophages.Virology 37:94-108.
8. Darlington, R. W., and L. H.Moss,III.1968. Herpes virus
envelopment.J.Virol.2:48-55.
9.Davis, B. D., R. Dulbecco, H. N. Eisen, H. S. Ginsberg, and W. B. Wood, Jr. 1967. Microbiology. Harper and Row Publishers, New York.
10. Fraser, D., and E. A. Jerrel. 1953. The amino acidcomposition of T3bacteriophage. J. Biol.Chem.205:291-295. 11. Fraser,P., H. R.Mahler, A. L. Shug, and C. A. J. Thomas.
1957. The infection of sub-cellular E.coli strainBwitha DNA preparation from T2bacteriophage.Proc. Nat. Acad. Sci. U.S.A.43:939-947.
12. Gamow,R.I.,andL.M.Kozloff. 1968. Chemically induced cofactor requirement for bacteriophage T4D. J. Virol. 2:480-487.
13. Guthrie, G. D., and R. L. Sinsheimer. 1963. Observationson
the infection of bacterialprotoplasts with theDNA of
bac-teriophage -X 174. Biochim. Biophys. Acta 72:290-297. 14. Jeme, N. K. 1956. Thepresenceinnormalserumofspecific
antibody against bacteriophage T4 and its increase during the earlieststagesof immunization. J. Immunol. 76:209-216.
15. Kellenberger, E., A. Bolle, E. Boy de la Tour, R. H.Epstein,
N.C. Franklin, N. K. Jerne, A. Reale-Scafate, J. Sechaud, I.Bendet, D. Goldstein, andM. A.Lauffer. 1965. Functions and properties relatedto the tailfibers ofbacteriophage T4. Virology 26:419-440.
16. Konner, L., and L. Kozloff. 1964. The reaction of indole and T2bacteriophage. Biochemistry 3:215-223.
17. Kozloff, L. M. 1968. The large DNA bacteriophages, p. 435-496.In H. Fraenkel-Conrat (ed.),Molecular basis of
vir-ology. ReinholdBookCorp.,NewYork.
18. Long, C. 1961.Biochemistry handbook. D.vanNostrandCo., Inc., Princeton, N.J.
19. Martin, H. H. 1966. The biochemistry of bacterial cellwalls, p.457-484. In Paul D.Boyer (ed.) Annual reviewof
bio-chemistry, vol. 35.AnnualReviews, Inc.,PaloAlto, Calif. 20. Salton, M.R.J.1964. The bacterial cell walls. American
Else-vierPublishing Co.,NewYork.
21. Sechaud, J., E. Kellenberger, and G. Streisinger. 1967. Perme-ability of cellsinfected with T4r andr+ phages. Virology 33:398-404.
22. Simon, L. D., and T. F. Anderson. 1967. The infection of Escherichiacoli byT2andT4bacteriophagesas seeninthe
electronmicroscope.II.Structure andfunction of the
base-plate. Virology 32:298-305.
23. Weidel, W., and J. Primosigh. 1957. Die gemimsame Wurzel derLysevonEscherichia colidurch Penicillin oder durch
Phagen.Z.Naturforsch.12b:421-427.
24. Wollman, E. L., and G. S. Stent. 1950. Studiesonthe activa-tionof T4bacteriophage bycofactor. I.Thedegreeof ac-tivity.Biochim. Biophys. Acta 6:292-306.
25. Wollman, E. L., and G. S. Stent. 1952. Studies on the
activation ofT4 bacteriophage by cofactor. IV. Nascent
activity.Biochim.Biophys.Acta9:538-550.