D022-538X/78/0027-0459$02.00/0
Copyrighti 1978 AmericanSocietyforMicrobiology Printed in U.S.A.
Seven
Complementation
Groups
of
Respiratory Syncytial
Virus
Temperature-Sensitive
Mutants
H. B. GIMENEZt AND C. R. PRINGLE*
Medical Research CouncilVirologyUnit, InstituteofVirology, Glasgow Gll 5JR, Scotland
Receivedforpublication13April1978
Fifteen temperature-sensitive mutants of the RSN-2 strain of respiratory syn-cytial virus have beenclassified into sixcomplementation groups, two of which appeared tobehomologous withtwoof the threecomplementationgroups of the A2strain describedbyWrightetal. (P.F.Wright,M. A.Gharpure,D.S.Hodes,
and R. M. Chanock,Arch. Gesamte Virusforsch.41:238-247). Thus seven
com-plementationgroups ofrespiratorysyncytial virus, designated A, B,
C,
D, E, F,and G, have been defined. The frequency and type ofmutant isolated varied
accordingtostrain;groupCwasuniquetothe A2strain,and groupsD, E, F,and
Gwereuniquetothe RSN-2strain. Thehighest
complementation
indexeswereobtained
by
preincubatio
foation atres ye mperaturefor40to 50h inthecaseof A2 strain mutants o fr 1-2 strainmutants.Genetic recombinationwas notdetected.
Temperature-sensitive (ts) mutants have been isolated fromtwostrains ofhuman
respi-ratorysyncytial(RS)virus. Seven mutantswere
isolated from the A2 strain by Gharpure et al. (5) and classifiedintothree groups (designated
A, B, and C) by genetic complementationtests
(11). Five of these seven mutants belonged to
complementation group A. Another36 mutants
were isolatedfromthe RSN-2 strain of RS virus
byFaulkneretal. (4), andaminimumof three
complementation groupswasrecognizedonthe
basis ofgenetic and physiological studies. We now reportfurther genetic characterization of the tsmutantsof theRSN-2 strain and identi-fication of thehomologiesofthe
complementa-tion groupsof the A2 andRSN-2strains.
Opti-mal conditions forcomplementationwerequite
distinct in the two strains. Definition of these conditionswas anessentialpreliminaryto dem-onstration of interstrain complementation. We suggest thatasimilar approachmay be
appro-priatewith other viruses which do notgrowto
hightiter and wherecomplementationhas been
difficulttodemonstrate.
Fifteen of the RSN-2 ts mutants have been
classifiedinto six
complementation
groups, twoof whichappearedto behomologouswith
com-plementation groupsA and B of the A2strain.
Thus sevencomplementation groupsare
recog-nized now in RS virus: groups A, B, and C, representedbyA2strainmutants,andgroups A, B,D, E, F, andG,byRSN-2strainmutants.
Genetic recombinationbetween
complement-t Presencomplement-t address: Faculcomplement-tad de CienciasExactasy Natu-rales,Universidad de BuenosAires, Buenos Aires,Argentina.
ing ts mutants was not detected in agreement with the previousfindingsofWright et al. (11) and Faulkner et al. (4). It is likely, therefore, that the RS virus genome is an unsegmented molecule andthat there may be at least seven
primarygeneproducts.Theidentificationof
be-tweensixandeightvirus-specified polypeptides
inpartiallypurifiedRS virus and in extracts of
RS virus-infected cells reinforces this conclusion (3, 7, 13). However, because five of the seven groups are represented by single mutants, the
occurrence of intracistronic complementation cannotbe excluded.
MATERIALS AND METHODS
Cells. The BS-C-1 and HeLa cell lines were ob-tainedfrom J. F. Williams,Institute of Virology, and
propagatedinrollerbottle culturesin Eagleminimum essentialmedium with twice the normal concentration of amino acids (Glasgow modification) and supple-mented with 5% fetal calf serum (virus and myco-plasma screened,Gibco-Biocult, Glasgow).
Virus. The A2 strain wild type and mutants tsl (A2), ts2 (A2), andts7 (A2) wereprovided by Linda
Richardson, National Institutes ofHealth,Bethesda, Md.The origin and characteristics of the RSN-2 wild type andts mutants have beendescribed previously (4). In this paper, to avoid confusion, the strain of originofindividual mutantswill be indicatedin paren-thesesfollowingthe mutant number.
Complementationtests. Complementation tests werecarried outby the method described byFaulkner etal. (4),with theadditional modificationsdescribed in thetext.Thecomplementation index was calculated asthe ratio:yieldfrom themixed infection (tsx+tsy) at39°C/sumoftheyields of the single infections (tsx) +(tsy)at39°C,where theyieldsareassayedat31°C, 459
on November 10, 2019 by guest
http://jvi.asm.org/
460 GIMENEZ AND PRINGLE
and tsx andtsy are anypair ofmutants. The multi-plicity of each mutant was the same in mixed infection andself-infection; i.e.,the totalmultiplicitieswere not
adjusted. Previousexperiments showed that this did not bias theresults.
Preparationof mutant stocks. RS virus does not grow tohigh titer,therebylimitingthemultiplicityof infection which can be achieved. The following
method of stock preparation wasfound to give the highestvirus titers. Each mutant was inoculated into a 30-ml (1-ounce) bottlecontaining 10' BS-C-1 cells
and incubated until cytopathiceffect was extensive. The monolayerwasscrapedinto suspension, and the
whole culture wasinoculated immediately into a 2-liter (80-ounce) roller bottle containing 108 BS-C-1 cells. The culture wasincubated for 1 h at 31°C to allow absorption of the inoculum. Then 40 ml of incubation medium wasadded, and the culture was
incubatedat310Cfor 5days.The incubation medium was discarded andreplaced by30ml offresh incuba-tionmedium. Incubationwascontinued at310Cfor a further48 to 72huntilcytopathiceffect wasalmost
complete.The attached cells wererapidly scrapedinto the medium, which was immediately dispensedinto vials and frozen inliquidnitrogen. Approximately10' PFU wereobtained from oneroller bottle culture.
Infectivityassay. Infectivities were measured on
BS-C-1cellmonolayers.Dilutions ofvirus-containing fluidswereinoculatedin0.2-mlvolumesandincubated for 30 min at 310C forabsorption. Then the
mono-layerswereoverlaid with 5 to7mlofEaglemedium
containing 0.9% agar and 2% fetal calf serum. The
infectedmonolayerswereincubated for 5 to 7 days at
39°C (therestrictive temperature) or7to 10daysat 31°C(thepermissivetemperature) inC02-gassedand humidified incubators. The monolayers were fixed with Formolsaline and stained with Giemsa. At both temperatures, RS virus plaques appeared as darkly staining heapsofcells,asdescribedpreviously(4, 12). The nonsyncytial plaque morphology of mutant ts2
(A2), which according to Gharpure et al. (5)
distin-guishesthismutantfrom the other A2 strainmutants, was notobserved in BS-C-1cellsbecauseallstrains of human and bovine RS virus so far examinedproduce
the sameuniform focalplaqueonBS-C-1 monolayers
under agaroverlay (3).
Growthcurvedetermination.Replicatecultures of2 x10 HeLa cells in 30-ml(1-ounce) screw-capped
bottleswereinfected withapproximately1PFU of the wild type of either the A2 or the RSN-2 strain per cell. The infected cultures were incubated at390Cfor 30 min toabsorb the inoculum. The inoculumwasthen removed by two washes of coldmedium, and finally 5 ml ofwarm incubation mediumwas added and the bottles were placed in a water bath at 390C. Two bottles were removed at intervals up to 44 h after infection and frozenat-70°C. The infectivities were measured onBS-C-1monolayersat310Cin thenormal
manner.
RESULTS
Growth curve of wild-type virus at
re-strictivetemperature.Figure1showsthatthe
growth of wild-typevirusof the A2 and RSN-2 Y
I
I
-J
x 04
-LI
z
0 0
-J
I-0
C-)
P-4
--4
-u m
0
I
I-I
HOURS
FIG. 1. Growth of wild-typevirus (ts+) ofthe A2 andRSN-2strains in HeLa cellsat39°C. Monolayers ofHeLa cells in 1-ounce (ca. 30-ml) bottles were
infectedwith 1 PFUofts+(A2)orts+(RSN-2)percell asdescribed in the text and incubated in a water bath at39°C. Replicatecultures were harvestedby freeze-thawingatvarious timesafter infection, and totalinfectivity wasassayed on BS-C-1 cell
mono-layers at 31°C. (a- ) ts+ (RSN-2); (---) ts+
(A2).
strains in HeLa cells at 390C was essentially similar. The yieldof theRSN-2 strain virus in
HeLacellswaslower, althoughthe cultures had
been infectedatthesamenominalmultiplicity.
The incremental yields, however, were
compa-rable. This result isprobablyareflection of the
pasthistoryof thetwostrains andthe factthat
the inoculum virus titers were determined on
BS-C-1 cell
monolayers.
The RSN-2 strain hadbeengrownexclusivelyonBS-C-1 cells and
ex-hibited some adaptation to these cells (4),
whereas the A2 strain had beengrown predom-inantlyin HeLa cells.
Effect of theperiodof incubationat31°C
oncomplementationyields.Factorsaffecting
theefficiency ofthe standardcomplementation test wereexaminedtoimprove consistencyand reducevariability between experiments. Itwas
confirmed thatcomplementation indexes were
higher andmorereproducible when HeLacells
wereusedashost for the mixed infection rather thanBS-C-1 cells (4). Anotherimportantfactor
was the length of the period of incubation at
permissive temperature
(310C)
before removalof the inoculum and incubation at restrictive
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.501.271.463.59.322.2]SEVEN RS VIRUS COMPLEMENTATION GROUPS
temperature (390C). Table 1illustratesthat the
complementation index in a mixedinfectionof
two A2strainmutants wasincreased from 100
to 813 by increasing the incubation period at 31°C from 1 to7 h.Other experiments showed
that increasing the310C incubation period
be-yond 7 h did not further enhance the comple-mentation index.
Effect ofthe period of incubation at
re-srictivetemperature oncomplementation.
Theeffectofincreasingperiodsofincubationat
390C on the complementation index is
illus-tratedinTable2.HeLa cells wereinfectedwith
two complementing mutants from the RSN-2
strain, usingtwodifferent periods ofpreliminary
incubation at310C, namely, 1.5h and the
opti-mal7h. Thecomplementationindexes increased up to 88 h after absorption and subsequently
declined.Atall times thelongerincubation
pe-riod at
310C
gave higher complementationin-dexeswiththese RSN-2strain mutants, as
ob-servedpreviouslywith A2 strain mutants (Table
1).
Optimum incubation periodsat390C.
Ini-tiaily,
theoptimumperiod of incubationat390Chad been determined for mutants of the A2
strainandfoundtobe between 40 and 50 h. This
isillustratedinFig.2aformixed infection by tsl
(A2)and ts7(A2). However,severalexperiments
showed thatcomplementationbetween mutants of the RSN-2 strainwas poorunder these
con-ditions. The experiment with mutants ts17
(RSN-2)and ts19 (RSN-2)illustrated inFig. 2b
shows that muchlongerincubationwasrequired
to obtain satisfactory complementation with
RSN-2 strain mutants. The optimal
period
ofincubationat
390C
wasbetween 80and90h, i.e.,TABLE
1. Increasein
compkmentation
index(C1)with timeof absorptionat31°C
Absorp- Incuba-Mutants Host cell h?n pe. toniodat at390CteCI
31°C (h) (h)
tsl (A2) + HeLa 1 47 100
ts7 (A2) 2.5 45.5 160
4 44 240
7 41 813
TABLE 2. Complementationindexesinmixed
infectionofHeLa cells with mutantstsl 7(RSN-2)
andtsl9(RSN-2)
Absorption Complementationindexat time of incubation timneat310C (h)at390C:
(h) 63 72 88 96 112 135
1.5 19 75 78 76 6 10
7 64 90 210 145 13 38
2000
x
21600
61200
z 800
e 400
I
0
C-40 60 80 100
HOURS
FIG. 2. Complementation index after various
pe-riods of incubation at restrictive temperature. (a)
Complementationindexes in amixed infection with two A2 strain mutants, tsl (A2) and ts7 (A2). (b)
Complementationindexes in a mixed infection with two RSN-2 strain mutants, tsl7 (RSN-2) and tsl9
(RSN-2). (c) Complementation indexes in an
inter-strain mixedinfection,tsl(A2)andtsl9 (RSN-2).
twice as long as in the case of the A2 strain
mutants.Underthese conditionsof infection the observedcomplementationindexeswerehigher
thanpreviously reportedfor anypairof RS virus
ts mutants. The data illustrated in Fig. 2a, b,
andc wereobtained inasingleexperiment,and therefore the indexes are directly comparable.
Figure 2c illustrates the complementation
in-dexes obtained in a mixed infection with one
mutant from each strain, tsl (A2) and
tsl9
(RSN-2).
It isapparentthat interstraincomple-mentation experiments between the A2 strain
and theRSN-2 strainrequire careful design and evaluation.
ThedatainTable3 show that itwasnecessary to measure complementation indexes in inter-strain
experiments
over a range of incubationtimesspanningtheoptimaofthe two strains. In
this
experiment,
twots mutants oftheA2strain, representingcomplementation groupsAandB,werecrossed with fourmutually complementing
461
VOL. 27, 1978
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.501.250.444.63.369.2] [image:3.501.45.240.466.557.2] [image:3.501.46.240.527.660.2]TABLE 3. Interstraincomplementation; effect of varying the period of incubation at 39°C
Complementation indexes in mixed infection at time of incubation (h) at39°C":
Mutant 37 43 50 61 75
t.s ts2 ts1 ts2 tsl ts2 tsl ts2 tsd ts2
(A2) (A2) (A2) (A2) (A2) (A2) (A2) (A2) (A2) (A2)
tsl(RSN-2) 0.8 <1 96 <1 4 50 2.7 1 5.5 <1
tsl7(RSN-2) <1 0.4 73 <1 2 1.3 <1 1.1 1.1 <1
tsl9(RSN-2) 5.4 0.2 535 <1 7 78 ND <1 ND <1
ts20(RSN-2) <1 <1 67 <1 2.6 86 0.5 <1 3.7 <1
a
Optimum
values are indicated in italics. ND, Not done.mutants[tsl(RSN-2),ts17(RSN-2),ts19 (RSN-2), andts2O (RSN-2)]oftheRSN-2strain.
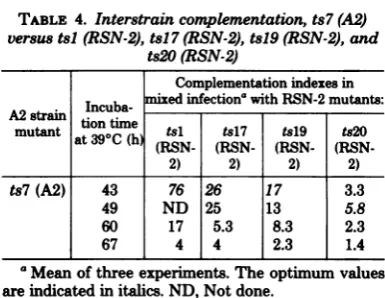
Mu-tant tsl (A2) complemented all fourRSN-2 ts mutantsoptimallyat43h,whereasmutant ts2
(A2) complemented mutants tsl (RSN-2), tsl9
(RSN-2),andts2O(RSN-2)at50 honly. Mutant ts2 (A2)didnotcomplementmutantts17
(RSN-2) at any time, and both mutants have been
assigned to the same complementation group
(group B) onthe basis of these and additional
data. Mutants tsl (RSN-2), tsl9 (RSN-2), and
ts2O (RSN-2) complemented both tsl (A2) and ts2(A2) ateither 43or50h,and therefore did not belong to complementation group A or B.
Since these three RSN-2 tsmutants werealso
complemented by ts7 (A2), representing group
C, they are assigned to new complementation
groups, whichhave been designated D, E, and
F,respectively (seeTable4).
The seven complementation groups. Of the remaining 32 RSN-2 strain mutants, only
one [mutant ts6 (RSN-2)] complemented each
ofthe six mutantsrepresentinggroupsA, B, C,
D, E, and F. Mutant ts6 (RSN-2), therefore, representedtheseventhcomplementationgroup
(Table 5). Table 5 also shows thatmutantts26
tecombinants was very low, they could have (A2),tsl (RSN-2),tsl7 (RSN-2), tsl9 (RSN-2), ts2O(RSN-2),andts6(RSN-2),but not tsl (A2).
Mutantts26
(RSN-2), therefore,
wasconsideredtobelongtocomplementationgroupA.Another nineRSN-2 tsmutants complementedthe
mu-tantsrepresentinggroupsA, C, D, E, F,andG,
but not tsl7 (RSN-2) (Table 6) oreach other
(datanotshown). Therefore,thesemutantshave beenplacedincomplementationgroup B. The remaining21RSN-2 straintsmutantsfailed to complement,orcomplemented sporadically,and
couldnotbe classifiedintoanycomplementation
group.
Absence of genetic recombination. No wild-typerecombinants have been found previ-ously in the progeny ofmixed infections of ts mutantsofeithertheA2strain(11)orthe
[image:4.501.266.459.204.353.2]RSN-2 strain (4). Nevertheless, ifthe frequency of RSN-2) complemented mutants ts2 (A2), ts7
TABLE 4. Interstraincomplementation,ts7(A2) versus tsl(RSN-2), tsl7(RSN-2),tsl9(RSN-2), and
ts2O(RSN-2)
Complementation indexes in
Incuba- mixed infection' withRSN-2mutants:
Astuntion
timemutant
ati
at 39 C (htime (RSN-tsl (RSN-tsl7 (RSN-ts19 (RSN.ts2O2) 2) 2) 2)
ts7(A2) 43 76 26 17 3.3
49 ND 25 13 5.8
60 17 5.3 8.3 2.3
67 4 4 2.3 1.4
'Meanof threeexperiments.Theoptimum values are indicated initalics.ND, Notdone.
escapeddetection in the earlier work.Therefore,
theyieldfromthecomplementationexperiment
illustrated in Fig. 2b was examined to
determine whether thegeneration ofwild-type
virusbyrecombination had contributed tothe
highyields obtainedinthisexperiment.Aseries
ofcloneswasisolated from thecomplementation
yieldsat69, 75,88, and 100h,and theseclones
were screened for ability to form plaques at
39°C. All317clones examinedweretemperature
sensitive like the parental virus; therefore, the
wild-type recombinant frequency, even under
conditions favoring the recovery of wild-type
virus, couldnothaveexceeded 0.03%.
DISCUSSION
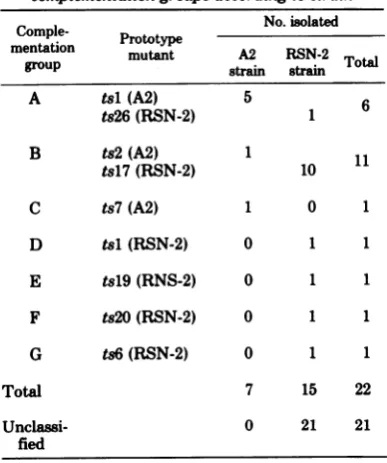
Table 7 summarizes the classification ofRS virusts mutants intocomplementation groups.
Thereisamarked difference in the distributinn
sandstrain
ofogin
Forinstance o the7
stami
iunts intocompleme
ngroup
A.whereasonly1Of-the
5 tr mut-antsappearsinthisgroul.
Similarly,
10 ot tne 15 RSN-2strainmutan`ts
wereclassifiedingroupB,butonly1of the7A2
strain mutants. Group C is unique to the A2
strain,andgroupsD,E, F,andGareuniqueto the RSN-2 strain. These differencesmay indi-cate inherentdifferencesin themutabilityof the twostrains.Ontheotherhand,theA2 mutants 462
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 5. The sevencompiementationgroupsdefined by inter-andintrastrain mixedinfections
Complementationindexes ingroup:
Group Mutant A B D E F G
ts26 tUl7 tsl tsl9 ts2O ts6
(RSN-2) (RSN-2) (RSN-2) (RSN-2) (RSN-2) (RSN-2)
A tsl(A2) 1.5 72 96 555 67 3
B ts2(A2) 14 1.6 50 78 86 5
C ts7(A2) 11 26 76 21 6 28
A ts26(RSN-2) - 11 6 10 4 23
B tsl7(RSN-2) - - 550 536 100 7
D tsl (RSN-2) - - - 150 9 23
E tsl9(RSN-2) - - - - 68 114
F ts20(RSN-2) - - - 60
[image:5.501.50.452.82.339.2]aHighest valuein three tofiveexperiments.
TABLE 6. Assiwnmentof RSN-2mutants togroupB ComplementationindeX
Mutant
ts17
tsl9
ts3 ts4 | 5 |sl2MU t16 tsN8 ts23 ts25 ts37 (RSN-2) (RSN-2) (RSN-2) (RSN-2) (RSN-2) (RSN-2)(R)N-
(RSN- (RSN-2) (RSN-2) (RSN-2)______________________________________________ 2) 2)
tsU7(RSN-2) (2) 650 0.4 0.2 0.7 1 1.5 0.5 1.2 0.6 0.3
(3) (160) (2) (120) (0) (1) (0) (2,000) (300) (89) tResult ofsingle
experiment;
optimum
timeofincubation for RNS-2mutants.Figuresinparentheses arenumberof PFU in self-infection.
TABLE 7. Distributionofmutantsinto complementation groupsaccordigto strain
Comple- No.isolated
mentaonmentation Prototypemutant A2 RSN-2
grOUp strain1 strainl Total
A tsl(A2) 5
ts26(RSN-2) 1 6
B ts2(A2) 1
tsl7(RSN-2) 10
C ts7(A2) 1 0 1
D tsl (RSN-2) 0 1 1
E tsl9(RNS-2) 0 1 1
F ts2O(RSN-2) 0 1 1
G ts6(RSN-2) 0 1 1
Total 7 15 22
Unclassi- 0 21 21
fied
were isolatedfrom HeLa cells and the RSN-2
mutants from BS-C-1 cells, and it is possible that the host cell may influence the type and frequency ofmutants. Similardifferenceshave
been observed with ts mutants of the human
rhabdovirus Chandipura virus after
5-fluorour-acilmutagenesisindifferent hostcells(Gadkari
and Pringle, unpublished data). However, the
differentfrequenciesmaymerely reflect the
dif-ferentprocedures employed ineachlaboratory.
A
high_proE2rtion
(21/36)
of the RSN-2 strain_______not-e
cassifed
intoomen
~~~asuggesf~L
etp
mutationalevents or alesion
affecting
fusible
geneDroduct.
Two of thethreeunnamedcomplementation groupsof theRSN-2 straindescribed by
Faulk-ner et al. are equivalent to group B (mutant
ts23) andgroup E(mutanttsl9).Theidentityof
thegrouprepresented bymutanttsl5 is obscure
because themutanttsl5 usedby Faulkneretal.
(4) couldnotberecovered fromstorage.
The absenceof recombination and the
appre-ciable number of complementation groups (7
from22 mutants) arecharacteristic features of
otherparamyxoviruses.Forexample,tsmutants
ofSendai virushave been classified intoseven
groups (8), fiveand apossible sixth complemen-tation group have been described for measles virus(2), and five have been claimed for New-castlediseasevirus(10). Therefore the size and
structure.ofthe RS virusgenome may be similar
tothatof other
paramyxoviruses.
The.datecion
ofcomplementAtinnbetLeen
mutantsofRSvirusdependedonseveralfa
whichfincluded
thenos1cee
type,multiplicity of 27,on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.501.50.244.374.606.2]infection, durationofpreliminaryincub at
permissive-temperature.
andperiod Of incuba-tionatrestrctve temperAttempts
toim-prove
complemen
by creation ofviralag-gregates using
poly-L-ornithine,
nucleohistone,orcoprecipitation byPEG-6000wereineffective.
It wasconfirmedthat HeLa cels wereabetter IMt tha
M-U-_
-ceffs
e ada that this was aconseauenceof the greatertemperature
sensitiv-it-y
(lowerleak) ofmutantsinthesecell(dt
not shown). The multipcy ofi onused
was thehcouldbeachieved
(about
1PFU/cell
at the best), but the low intectivity --Sed
-w'ititers obtaine w Ith S viru were
probal
resposbefor teproiDiemencoutrdn
~mplementtionanalysis.Aperiodof
absorp-tionat3100of 7 h increased the
complementa-tionidex-witho_ut
increaigtes-yielda
otmutantsofbothstrainsequally.
However,
theoptimal
timeof incubationat39°C differed (Fig. 2). The
rea-son for this difference is not clear since the growthcyclesof the wild types of thetwostrains
in HeLa cells at
390C
did not show obviousdifferences (Fig. 1).
Possibly
amplification
ofgene productswas required inthe case of the
RSN-2 mutants, whichhad lowereffective
mul-tiplicitiesofinfectioninHeLa cellsthenthe A2
strainmutants.Althouh22tsmutantshave been
grouped
satisfactorily
into seven groups, it ispossible that these conditionswere not
appro-priate for complementation between mutants
withlesionsinothergeneproducts. The
example
of RS virus may be instructive in the
genetic
analysisof other animal viruseswhere
difficul-ties have been encountered in
demonstrating
complementation.
Some phenotypicproperties of the A2 strain
mutants in groups A,
B,
and C have beende-scribed (1, 6, 9), the most
striking being
thenonsyncytial
cytopathology
associated withmu-tant ts2
(group
B).
It may besignificant
thatnoneof theRSN-2mutants
presently
classifiedingroupBinduced the
nonsyncytial
cytopathic
effect in
HEp-2
cellsreported
for ts2(A2).
Therefore the homologyof thesemutants maybespurious,and absence of
complementation
inthiscase mustbe treated with caution. Thesame
reservationmayapplytogroupA,sincetsl(A2)
is thermosensitive and ts26 (RSN-2) is ther-mostable(unpublished data).Therefore seven is the minimum number of complementation groups resolved in the present study, if
intracis-troniccomplementationisdiscounted. ACKNOWLEDGEMENTS
This workwasasistedby financial supporttoH. B. Gi-menezfromthe British Council, the Medical Rearch Coun-cil, and the World Health Organization.
LTERATURE CTD
1.Belshe, R. B., L S.Richardson, T. J.Schnitzer,D.A. Prevar, E. Camargo, and R. M. Chanock. 1977. Furthercharacterization of the complementation group Btemperature-sensitive mutant ofrespiratory syncytial virus. J.Virol. 24:8-12.
2. Breswhkin, A. M., F. Rapp, and F. E. Payne. 1977. Complementationanalysis of measles viru ts mutants. J.Virol. 21:439-441.
3. Cash,P., W. H.Wunner,andC. R.Pringle.1977.A companson of thepolypeptides of human and bovine respiratory syncytial virume and murine pneumonia virus.Virology 82:369-379.
4.Faulkner, G. P., P. V. Shirodarla,E. A. C.Follett, and C. R.Pringle.1976.Respiratorysyncytial virusts mutants and nuclear immunofluorescence. J. Virol. 20:487-500.
5. Gharpure,ALA.,P. F.Wright,and R. LCho 1969. Temperature-sensitive mutants of respiratory syncytial virus. J. Virol. 8:414 421.
6.Kalica, A. R., P. F. Wright, F. MLHetrick, and R. IL Chanock.1973.Electronmicroscopicstudies of respi-ratorysyncytial virus temperature-sensitive mutants. Arch.GesamteVimrforach.41:248-258.
7. Levine, S. 1977. Polypeptidesof respiratory syncytial virus. J.Virol. 21:427431.
8. Portner,A., P. A. Marx, and D. W. Kingsbury. 1974. Isolation andcharacterizationofSendaivirus temper-ature-sensitive mutants. J. Virol.18:298-304. 9.Schnitzer, T. J., L. S. Richardson, and R. M. Chanock.
1976.Growth and genetic stability of thetM-Imutant of respiratory syncytialvirusatrestrictivetemperature. J. Virol.17:431438.
10.Tslpis, J. E., and AL A. Bratt. 1976. Isolation and preliminary characterisation oftemperature-sensitive mutants of Newcastle disease virus. J. Virol. 18:8484866.
11. Wright, P. F., M. A.Gharpure,D.S.Hodes, and R. M.Chanock.1973.Genetic studies of respiratory syn-cytial virustemperature-sensitive mutants. Arch. Ge-samteVirusforsch. 41:238-247.
12.Wunner, W. H., G. P.FaulEker,and C.R.Pringle.
1975.Respiratorysyncytial virus: some biological and biochemicalproperties, p. 193-202. In B. W. J. Mahy andR.D.Barry (ed.),Negative strand virses,vol.I. AcademicPrees,London.
13.Wunner,W.H.,and C.R.Pringle. 1976.Respiratory syncytialvirusproteins. Virology 73:228-243.
on November 10, 2019 by guest
http://jvi.asm.org/