Vol. 45, No. 3 JOURNALOFVIROLOGY,Mar.1983,p.1118-1139
0022-538X/83/031118-22$02.00/0
CopyrightC)1983,American SocietyforMicrobiology
Coliphage
P1
Morphogenesis: Analysis of Mutants by Electron
Microscopy
JEAN T. WALKERANDDONALD H.WALKER, JR.* DepartmentofMicrobiology, University of Iowa,IowaCity, Iowa52242
Received 1 July1982/Accepted19October 1982
Weused electron microscopyandserumblockingpowerteststodetermine the phenotypes of 47 phage P1 ambermutants that havedefects inparticle morpho-genesis. Elevenmutantsshowed head defects, 30 showed tail defects, and 6 hada
defect inparticle maturation (which could be either in the head or in the tail).
Consideration of previous complementation test results, genetic and physical positions of the mutations, and phenotypes of themutantsallowed assignment of mostof the 47 mutationstogenes.Thus,aminimum of 12 tailgenes,4headgenes,
and 1particle maturationgeneare nowknown for P1. Of the 12 tailgenes,1 (gene
19,located within the invertible C loop) codes for tail fibers, 6 (genes 3, 5, 16, 20, 21, and 26) code for baseplate components (although one of these genes could
code for the tail tube), 1 (gene 22) codes for the sheath, 1 (gene 6) affects tail length, 2 (genes 7 and 25)areinvolvedin tailstability, and 1 (gene 24) either codes
forabaseplatecomponentorisinvolved in tailstability. Of the four headgenes, gene9 codes foraprotein required for DNA packaging. The function of headgene
4is unclear. Headgene8probably codes foraminor headprotein, whereas head gene 23 could code for either a minor head protein or the major head protein.
Excluding theparticle maturationgene(gene 1), the 12 tailgenesareclustered in
threeregions of the P1 physicalgenome.Thefourheadgenesareatfourseparate locations. However, someP1headgeneshavenotyetbeendetected and could be located intworegions (for which therearenoknowngenes) adjacenttogenes4 and 8.The P1 morphogeneticgeneclustersareinterrupted bymanygenesthatare
expressed in the prophage.
Coliphage P1 consists of an icosahedral, DNA-containing head and a complex tail (47;
Fig. 1). The tail has a tube surrounded by a
contractile sheath. At the distal end of the
sheath is an ill-defined baseplate to which
kinked tailfibers(probably six)are attached. At the proximal end of the sheath is a cap
(occa-sionally seen ontails unattachedto heads).The
head is attached to the tail via a neck which protrudes from a vertex of the head (47). The neck is attached to the head via a head-neck connector(which isonlyseenwhen the head is empty or has beendisrupted).
Oneoftheunique characteristics ofP1 is that it normallyproduces isometricheadsof at least threedifferent sizes(PlB,P1S,andPlM,having
diameters of 86, 65, and 47 nm, respectively
[47]). As part of an approach to determine how the different head sizes are assembled, we exam-ined 103 P1 amber mutants to identify those
defective in head structure.
Ageneticmap for Pl (Fig. 2), which includes these 103ambermutations,hasbeenestablished
(49). Two-factor crosses permitted assignment
ofthemutationsinto 10lineararrays, orlinkage clusters, I through X, with mutations in cluster X behaving as two subgroups, X-1 and X-2. However, cistrondesignations,apart from those
originally
assigned
by Scott (39), were notas-signed because
complementation
tests inliquid
frequentlygave ambiguous results.
With other phages, amber mutants can be
divided into two major classes
depending
on whetherthey have a genedefectivein afunctionwhichisrequired early or one which isrequired
latein phage particle development. Late genes are involved in particle
morphogenesis
(e.g., production ofproteinswhich become structuralcomponents of the particle) or in cell
lysis.
Generally,earlygene mutants(i.e.,those defec-tive inregulation of transcription,DNA synthe-sis, orrecombination) do not allowsynthesis of any of the late gene products, including those
causing
lysis.
Thus, those mutants that cause celllysis
should havemorphological
defects. Therefore, we determined which of the 103 P1amber mutants could cause cell
lysis
(50)
andexamined55ofthembyelectronmicroscopyfor
1118
on November 10, 2019 by guest
http://jvi.asm.org/
MORPHOLOGY OF P1 MUTANTS 1119
B
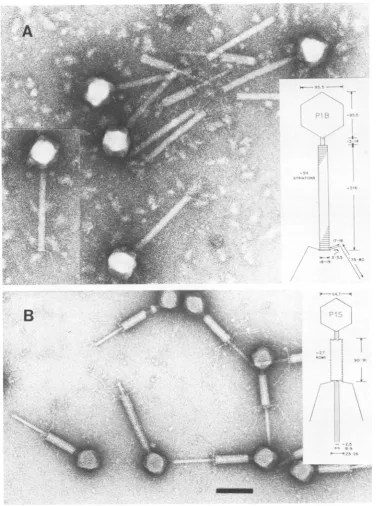
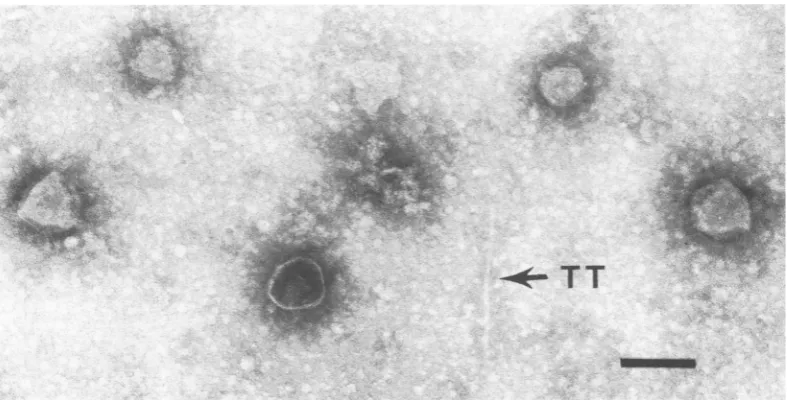
FIG. 1. Micrographs showing normalPlparticles.(A) LysateofP1vir * Sh hasbig heads(P1B),full ofDNA, andtails, themajority of whichhavesheaths.(B)Particles from aCsCIgradient. The full small heads(P1S)have
tails with sheaths invaryingstages ofcontraction; baseplates and tail fibers canbeeasily seen. Insets show diagrammatic scale drawings ofPl;dimensionsare innanometers.Bar, 100 nm.
structural defects(this paper). In doing so, we identified those mutants whose defects are in early functions, in lysis functions, and in head
and tail morphology. In addition, because the
genetic (39, 50)and
physical
(21, 34,38,
43,45)
locations of these mutations on the P1 genome VOL. 45,1983
i. ..
4i
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.489.60.437.68.575.2]1120 WALKER AND WALKER
areknown, wedetermined theextentof cluster-ing of the functions represented by these
mu-tants.
MATERIALS AND METHODS
Bacterial and phage strains. Bacterial strains were Shigella dysenteriae
sup'
strains Sh-15 and Sh-16(which isastreptomycin-resistant Sh-15 strain) (5) and
the tetracycline-resistant sup+ strain Sh-16supF con-structedby Van Montagu (50). Phage strainswereP1
vir(thesupervirulent mutantP1 kcvirs of Sarkar that grows on strains lysogenic for P1 [39]), P1 amber
mutants(Fig. 2)13, 16, 19, and 21 of Scott (39), and 51 P1 ambermutants of Walker and Walker(49). These P1 ambermutantsalso have the vir mutation (i.e., they are vir amber double mutants); for simplicity these
mutantswill be referredtoasaml3,etc.,without the virdesignation.
Media, phage assays, and thepreparation of phage stocks. Media, phage assays, and the preparation of phage stocks were asdescribedpreviously (50).
Preparation of defective lysates of amber phage-infected cells. At 0min the desired ambermutantwas added(atamultiplicity of infection of4to6)toa log-phase L-broth cultureof S.dysenteriae
sup'
strain Sh-15. vir ambermutantswereusedtoavoid lysogeniza-tion (32), andamber'P1 virwasincludedas acontrol. After allowing 10 min at 37°C for adsorption in theLINKAGE CLUSTER CISTRON
I
presence of 5 mM CaCl2, the culture was filtered to
remove unadsorbed phage, diluted 2.5-fold in fresh medium containing 5 mM MgSO4, and incubated at
37°C with vigorous shaking. With the onset oflysis (usuallyat60to70min when it occurred), DNase and RNasewereaddedtofinalconcentrations of 1 ,ug/ml.
At 110min, when lysiswascomplete,asample of the lysatewasprepared immediately for electron
micros-copyasdescribedbelow, and thelysatewasassayed, withsupoandsup' strains Sh-16 and Sh-16 supF, for viable phage and revertants. Under these conditions titers formutantsonsup'cellswerelowerbyafactor of104than those obtained usinga P1 virlysate as a control. This iscomparabletoresults obtained for T4 (55) and P2 (32). Such titers for mutants are due primarilytotransmission of ambers.
The remaining lysate was chilled, and 5 ml was centrifuged (at 35,000rpmfor 2sat4°CinanSW50.1 rotor) to pellet unlysed cells and cell debris. The
supernatantfluidswerecentrifuged (at 22,000rpmfor
2 h in thesamerotor)topelletanyphagecomponents present. The pellet was left at 4°C for 2 days to
resuspend in 0.25 ml of culture fluid (i.e., a 20-fold concentration of phage components), and a sample wasprepared for electron microscopy.
Serumblockingassays.Those defectivelysates used for electron microscopywerealsoassayedforserum blocking antigen (SBA). Themethod fordetermining
m
4 17 2
3
AMBER 47141 3734 4 17357 129 7 . 244731 13420 17 U72 6649 74 75 6171
MUTATION 132_
170 155
LYSIS NORMAL 11 NO Il-vJI NORMAL I
PHENOTYPE LYSIS PREMATURE
SERUM
BLOCKING I POSITIVE 'L-
NEGATIVE'1I
POSITIVEPOWER
Vll VlilX11-2 I- 1
5 6 7 6 10 9
ItW I .. .11_ ..i a. . .1.. . M...W 1la i( 111 ..._ ..Si .1 (I _t, , , J.mI 11 .,!i .,1 ill im (!l,1
I35Io7 45 28 19 1622L9 6 32 14 395UI102 IQ 40 29139 51 71 13 178 36 16 1524664 48 43
iBO 115 138 61 109 27 67 116 42 101151 68 11 104 54 36 58 06
179 110 125 160 113 4 140 70 . 131 112
175
NORMAL JI 11 JI-NORMAL-I
LYSIS
POSITIVE POSITIVE
NEGATIVE
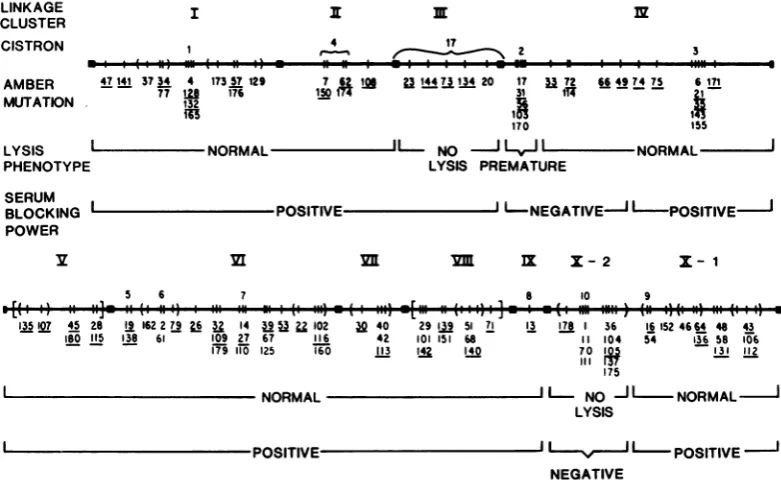
FIG. 2. GeneticmapofPlambermutations, modified fromWalker and Walker(50), showingresultsofhost
celllysisandserumblockingpowertests.Regionsdemarcatingthe 10linkageclustersarerepresented bysolid blocks.Thecistrondesignationsabove the map line include theoriginal1to10of Scott(39)and 17ofWalkerand
Walker(50).Groups ofambermutationsvertically stackedunder very close hash marksorwithinparenthesesdo
not showcomplementation (49). The orientationof groups ofamber mutations within parentheses orwithin
bracketswith respecttomutations outsidethese groups is unknown. Thespacingofthe hash marks isnot meant toindicate eithergeneticorphysicaldistance. Underlinedmutations indicate themutantsthatwereused in the
experiments reportedin this paper.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.489.53.444.349.592.2]MORPHOLOGY OF P1 MUTANTS 1121
SBAwas amodification of the DeMars(9)methodas used by Israeletal. (22) for P22 and Wardetal. (52) for T4;itcomparesratesrather thanfinallevels ofserum inactivation byblocking antigens.
Serial twofold dilutions were made of the lysates
afterthey had been treated with chloroform for 20 min
at 37°C to kill any viable cells. Cells infected with
mutants(in linkage clusters III and X-2; Fig. 2) that do
not cause cell lysis were lysed by sonication at the time when lysisoccurred with amber'-infectedcells. Cells infected with othermutantsand withP1 virwere
notsonicated,exceptasdescribed below for Pl vir,to
obtainastandardcurve. Reaction mixturescontained 0.45 ml ofalysatedilution and 0.1 ml oftesterphage,
towhichwassubsequently added 0.45 ml of antiphage serum.ThetesterphagewasP1 kcatafinal
concentra-tion of 3 x 105 PFU/ml; the turbid plaques could be
distinguished from the clear plaques ofanyrevertants presentinthe P1 v'iramberlysates.Thefirst-orderrate constant (k) for phage inactivation of the undiluted hyperimmune serum at 37°C was 1,500 min-'. This serumhadak of 0.1min-' in the finaldilution of the reaction mixture. Thereaction mixtureswereassayed (onstrain Sh-16) for surviving testerphageatthe end of 46min,whentesterphage survivalwasabout 1% in theabsence of SBA. This control without SBA was always included in eachgroupoftestsdoneon
differ-entdays. This inactivation was assumed tobe expo-nentialthroughout the 46-min incubation.
To obtain standardcurves,lysatesofamber' Plvir
grown on strain Sh-15 wereused as SBA;the phage wereinactivated by either sonicationorUVtreatment
sotheplaques wouldnotobscure those of the
compet-ing P1 kctesterphage.
The results of the serum blocking assays were plottedaspercentresidualk= (1 - kfinal/kinitial) X 100
=(1 -lnSISd/lnSISO) x 100,whereS/SO= survival of
thetesterphage in the control after 46min withserum but noSBA, and SISd = survival of the testerphage
after 46min with both serumand SBAatdilution dof the latter. Rather than showing graphs for the 55
mutants tested, comparisons were made using the dilutionatwhichresidual k= 50%.
Electronmicroscopy. A 0.4-misample of
concentrat-edorunconcentratedlysateof thephage-infected cells wasdialyzed for 15min by placing it on amembrane filter (Millipore Corp.; average pore size, 25 nm)
floatingon P1buffer(rather than water,which caused head disruption). This dialysis was done at room
temperature todecrease tail contractions obtained by dialysisat4°C. A drop of samplewasplacedon a glow-discharged, carbon-coated, 400-meshcoppergrid and allowed to stand for 2min. A small drop of sodium phosphotungstate (1%; pH 7.0)wasaddedandleft for 30 sbeforetheexcessliquidwasremoved with filter paper.A JEOL 100B electronmicroscope withan anti-contamination devicewasusedat80kV.Micrographs weretakenatinitial magnifications of10,000to50,000
onKodak 4463 film.
RESULTS
We examined, by electron microscopy, ly-sates of 55 P1 amber mutants grown under nonpermissive conditions to determine their phenotypes. Insuch lysates, mutantsdefective
in some aspect of tail assembly should produce
normal heads andoneofavarietyof tail compo-nentcombinations. Similarly, mutantsdefective in some aspect of headassemblyshouldproduce
normal tails and either no heads or one of a variety ofdefectivehead structures. Inaddition,
we used serum blocking assays to detect the presence oftail fibers in the defectivelysates.
Of103 P1 ambermutantspreviouslytested for lysisof nonpermissive cells (50), 83 caused cell
lysis at the normal lysis time for Pl. These 83
mutationsareinlinkage clusters(Fig. 2) 1,II,
V,
VI, VII,VIII, and IX and partsofIVand X(i.e.,
X-1). By analogy with other phages,these mu-tants would be defective in late genes involved in particlemorphogenesis.
The numberof cistrons represented by these 83 mutants is unknown. Therefore, 47 mutants
(underlined in Fig. 2) were chosenforstudy by electron microscopy from the maximum
possi-ble number of cistrons in the following way. These 47 mutants represent (i) at least one mutation from each ofthe groupings of
muta-tions(showninFig.2 asverticallystackedunder very close hash marks or within parentheses) that had been found (49) not to show comple-mentation and (ii) each ofthe single mutations
outside thesegroupings. In addition, eight mu-tants (also underlined in Fig. 2), representing
those that cannot cause cell lysis (linkage clus-ters III and X-2) or that cause premature cell lysis (linkage cluster IV), were also included in this study.
Serumblocking assays. P1 tailfibersareonly1 to2 nminthickness(47) and soareonlyseenby electron microscopywhenthey are attachedto
phageparticlesand when thestaining conditions
aresuchastogive optimalcontrastbetweenthe
fibers and the background (Fig. 1). Therefore, because in other phages with contractile tails,
suchasT4, themajority of SBAcanbe attribut-ed to tail fibers, serum blocking assays were done to confirmthe presence of attached fibers or todetect the presenceofunattached fibers in the P1 defectivelysates.Resultswereplottedas percent residual k versus lysate dilution as
de-scribedabove, and thedilutiongiving 50%
resid-ual kwas determinedforeach defective lysate.
Thesedilutions were designated SBA values. As would be expectedforaphage suchasP1 that has a complex morphology and therefore
would codefor various structures inaddition to tail fibers, the majority ofmutants showed sig-nificantserumblockingpower(similartothatof the control Pl vir lysates), indicating that tail fibers werepresent(Table 1; Fig. 2).
Thosemutantsthat gavenegative results(i.e.,
SBA values ofless than 1) have mutations in
linkage cluster X-2 and part of cluster IV. Be-cause cluster X-2 mutants didnot lyse the host VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
1122 WALKER AND WALKER
cells, these lysates were tested for SBA both before and after sonication to cause cell lysis;
negativeresults wereobtainedinboth instances,
indicating that these negative results were not
dueto afailure of the cells tolyse. (Mutants in
cluster III also did not lyse the host cells, but gave positive SBA results, indicatingthat soni-cation does notadversely affect SBA activity.) ClusterX-2mutations have been shown to be in the samecistron(cistron10),andmutants
repre-senting this cistronareprobablydefective in the
expression of late genes (50). Since SBAs are
lategeneproducts,this wouldexplain why nega-tive results areobtainedwithdefectivelysates of
these mutants.
Allof those mutants (74, 75, 21, 35, and 171)
located in the right part of linkage cluster IV gave normal levels of SBA. However, mutants (31, 56, 33, 72, 66, and 49)locatedinthe leftpart
of cluster IV gave negative SBA results (i.e., SBA values of lessthan1),indicatingthat these mutants produce few, if any, tail fibers. The
significance of these results obtained with the latter six mutants is discussed in the next
sec-tion.
Serumblockingassaysconfirmed electron
mi-croscopic examination (see below) of the mu-tants in the cases in which normal amounts of sheathed tailswereproduced.Thatis, tail fibers
were seen by electron microscopy on particles
produced by all ofthe mutants that produced normalamountsof sheathed tails (either extend-ed orcontracted), except mutants 128, 150, 33,
72, 66, and 138. The preparations of mutants 128, 150, and 138 were not good enough to
determine the presence of tailfibers, but these
mutants all showed serum blockingpower, and
for each there are allelic mutants (i.e., mutants that didnot show complementation [49])where tail fibers could be seen. Therefore, these mu-tants arepresumedtohavetailfibersattached to
their tails. Preparations ofmutants, 33, 72, and 66 in linkage cluster IV were good enough to detect any tail fibers ifpresent, and these mu-tantsdid not show serumblocking power, con-firming that these mutants do not produce tail fibers.
SBA was produced by all mutants (except
am49; see next section) that did not produce
sheathed tails (and where tail fibers therefore
could notbe detected byelectron microscopy), indicatingthattail fibers were made eventhough
no sheathed tail was available for their attach-ment.
Electron microscopy of P1 amber mutants. Defective lysatesof the 55 mutants were exam-ined by electronmicroscopy before, as well as
after, differential centrifugation to concentrate any phage structures present.
Amber'
P1 vir controls(Fig. 1) preparedatthesametimeasthedefective lysateswere examined in parallel. The types ofphage structures seen before and after
centrifugation were similar except that where unattached tails had sheaths which had slipped downfromthe top of the tail before
centrifuga-tion(mutantsin cluster VII and the right part of VI; see below), there was a decrease in the
proportion of tails relative to polysheath after
centrifugation. This decrease isprobably caused
bythe sheath subunits dissociating from the tail tubes after centrifugation and polymerizing to
formpolysheath during resuspension ofthe
cen-trifuged pellet at 4°C for 2 days, conditions which allowsheath subunits to polymerize in T4 (1). Where tails (either attached to or free from
heads) had extended sheaths before
centrifuga-tion, the majority had contracted or semicon-tracted sheaths after the concentration steps. Lysateswhich showedfull heads before
centrif-ugation showedfullheads aftercentrifugation if the heads were attached to tails. However, a
high proportion ofempty heads were seen after centrifugationif thefullheads were notattached
to tails before centrifugation. Results are sum-marized in Table 1 and Fig. 17.
Mutations affecting tail assembly. Mutations affecting tail assembly are located in linkage
clusters IV through VIII. Each tail-defective
mutant produces complete heads. The heads of
mutants33, 72, and 66(in linkagecluster IV) are
attachedtotails whicharecomplete except that they are missing tail fibers (Fig. 3). These mu-tantsdonot show serumblockingpower, which confirms that fibers are not present either
at-tachedorunattached to theparticles. However,
it is unclear whether the whole tail fiber is missingorjustthedistal portion, becauseshort
projections, which could be part of the base-plate,areoccasionallyseenprotruding fromthe base of the sheath, and the serum blocking
assaysdid not distinguish between these possi-bilities. Therefore, itremainsunknown whether the kinkedP1tail fiber is made of more than one
polypeptide and whether the mutations of mu-tants 33, 72, and 66 are in the same gene.
Mutants49, 74, 75, 21, 35, and 171 in cluster IV and mutants 135, 107, and 45 in cluster V
produce completeheads andpolysheath(Fig.4). In T4phage, completed baseplatesare required forthe tube
polypeptide
to initiatepolymeriza-tion to form the tube, and then the sheath
polypeptide polymerizes around the tube to form the tail (26). However, unlike the tube
polypeptide, the sheath polypeptide can
poly-merize by itself to formpolysheath. Therefore,
thepresenceof polysheathalone meansthat the defect is in thebaseplate, whereas thepresence of polysheath and baseplates means that the defect is in the tube (or baseplate, if the defect has not damaged the structural integrity of the J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
MORPHOLOGY OF P1 MUTANTS 1123
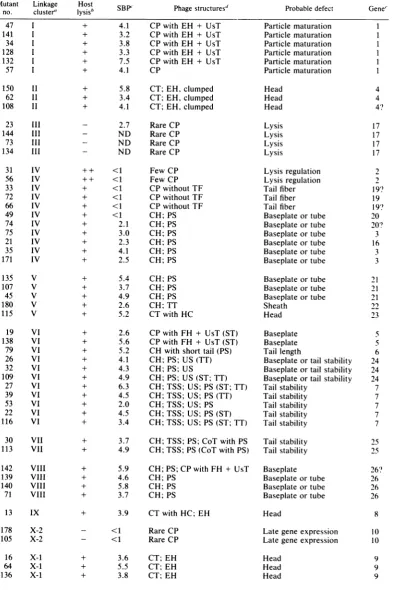
TABLE 1. Phenotypic classification ofP1 ambermutants
Mutant Linkage Host Probable defect Gene
no. cluster' ysisb SBP Phage structuresP
+ 4.1 CP with EH + UsT
+ 3.2 CP with EH + UsT
+ 3.8 CP with EH + UsT
+ 3.3 CP with EH + UsT
+ 7.5 CP with EH + UsT
+ 4.1 CP
+ 5.8 CT; EH, clumped
+ 3.4 CT; EH, clumped
+ 4.1 CT; EH, clumped
- 2.7 RareCP
- ND RareCP
- ND RareCP
- ND Rare CP
++ <1 ++ <1 + <1 + <1 + <1 + <1 + 2.1 + 3.0 + 2.3 + 4.1 + 2.5 Few CP Few CP CP without TF CP without TF CP without TF CH; PS CH; PS CH; PS CH; PS CH; PS CH; PS Particle maturation Particle maturation Particlematuration Particlematuration Particlematuration Particle maturation Head Head Head Lysis Lysis Lysis Lysis Lysis regulation Lysis regulation Tail fiber Tail fiber Tailfiber
Baseplateortube
Baseplateortube
Baseplateortube
Baseplateortube
Baseplateortube Baseplate or tube
+ 5.4 CH; PS
+ 3.7 CH;PS
+ 4.9 CH; PS
+ 2.6 CH;TT
+ 5.2 CT with HC
+ 2.6 CPwith FH+ UsT(ST)
+ 5.6 CP with FH+ UsT (ST)
+ 5.2 CH with shorttail (PS)
+ 4.1 CH;PS;US(TT)
+ 4.3 CH; PS; US
+ 4.9 CH; PS; US (ST;TT)
+ 6.3 CH;TSS; US; PS (ST; TT)
+ 4.5 CH;TSS;US;PS(TT)
+ 2.0 CH;TSS;US;PS
+ 4.5 CH;TSS; US; PS (ST)
+ 3.4 CH; TSS; US; PS (ST; TT)
+ 3.7 CH; TSS; PS; CoT withPS
+ 4.9 CH;TSS;PS(CoT withPS)
+ 5.9 CH;PS; CPwith FH+ UsT
+ 4.6 CH;PS
+ 5.8 CH;PS
+ 3.7 CH;PS
+ 3.9 CT withHC; EH
- <1 RareCP
- <1 RareCP
+ 3.6 CT;EH
+ 5.5 CT; EH
+ 3.8 CT; EH
Baseplateortube
Baseplateortube
Baseplateortube
Sheath Head
Baseplate Baseplate
Tail length
Baseplateor tailstability Baseplateortailstability
Baseplate or tail stability Tailstability Tail stability Tail stability Tail stability Tail stability Tailstability Tail stability Baseplate Baseplateortube
Baseplate or tube
Baseplateortube
Head
Late gene expression Lategeneexpression
Head Head Head 47 I 141 I 34 I 128 I 132 I 57 I 150 1I 62 1I 108 II 23 III 144 III 73 III 134 III 31 IV 56 IV 33 IV 72 IV 66 IV 49 IV 74 IV 75 IV 21 IV 35 IV 171 IV 4 4 4? 17 17 17 17 2 12 19? 19 19? 20 3 16 3 3 135 V 107 V 45 V 180 V 115 V 19 VI 138 VI 79 VI 26 VI 32 VI 109 VI 27 VI 39 VI 53 VI 22 VI 116 VI 30 VII 113 VII 142 VIII 139 VIII 140 VIII 71 VIII 13 IX 178 X-2 105 X-2 16 X-1 64 X-1 136 X-1 21 21 21 23 5 6 24 24 24 7 7 7 7 7 25 25 26? 26 26 26 8 10 10 9 9 9
VOL.45,1983
l
l l
on November 10, 2019 by guest
http://jvi.asm.org/
1124 WALKER AND WALKER
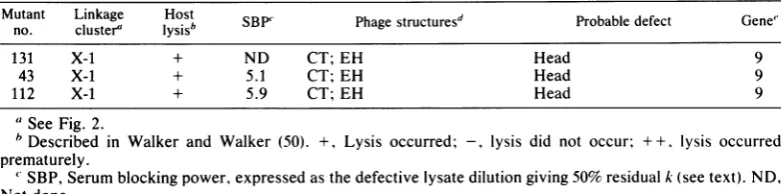
TABLE1-Continued
Mutant Linkage Host
SBP'
PhagestructureSd
Probable defect Gene"no. cluster"
1ysisb
B'Paesrcue131 X-1 + ND CT; EH Head 9
43 X-1 + 5.1 CT; EH Head 9
112 X-1 + 5.9 CT; EH Head 9
aSee Fig. 2.
bDescribed in Walker and Walker (50). +, Lysis occurred; -,
lysis
did not occur; ++,iysis
occurredprematurely.
' SBP, Serum blockingpower,expressedasthedefective
lysate
dilution giving 50%residual k (see text). ND, Notdone.dCH, Complete heads; CP, complete particles; CT, complete tails; CoT, contracted tails; EH,emptyheads;
FH,full heads; HC, head-neckconnectors; PS, polysheath;ST, stackedtails; TF,tail fibers; TSS, tails with slipped sheaths; TT, tail tubes; US, unit-length sheaths; UsT, unstable tails (tails in various stages of
contraction). Parentheses denote thatfew ofthesestructures were seen (see text).
eNumbers up to 10 assigned byScott(39);numbers 17and19through26 wereassigned by WalkerandWalker
(50; thispaper). Queries denote uncertain genedesignations.
baseplate but only the ability of the tube to be
built upon it.) The baseplate of P1 (Fig. 1) is muchthinner than that of T4 and has not been seenunattached to the P1 particle; therefore, the
defect ofthesecluster IV and V mutants could be in either baseplate or tube formation. In
addition to this defect, am49 does not show serumblocking power, indicating that tail fibers arenotproduced. This mutation is located next to mutations
33,
72,
and66,
where the only. b
!hI.1T.i.t
,,,,<Fa' l't.>,''.~~4at
{@-, N.
'..Z
^9,jGx..bvi~Ajk
~~~~~~~~I
^ _...O...-.defect is tail fiber production; therefore, am49 may be polar on these mutations (see below).
InT4, at least 15 genes are involved in base-plateformation (53); therefore, if these nine P1 mutants(49, 74, 75, 21,35, 171, 135,107,and45) are defective in baseplate formation, they may be defective in different components of the
base-plateand represent several genes. In support of this is the fact that mutants 49, 74, 75, 21, 35, and 171 are in one linkage cluster (IV) and
-:
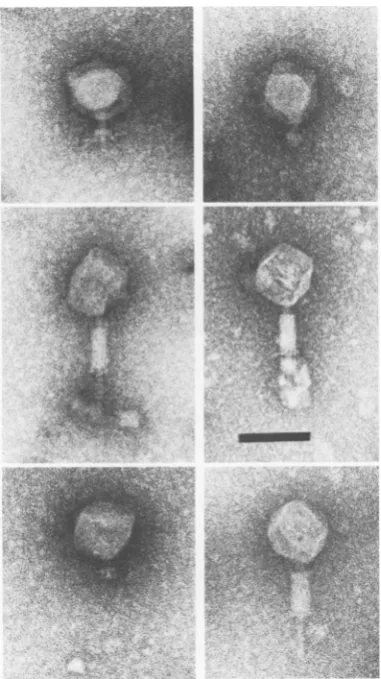
FIG. 3. MicrographofP1am66defectivelysate.InsetshowsasingleparticlefromP1am72defectivelysate. Allparticles lack tailfibers. Contractionofsheathsisanartifactduetopreparation procedures.Bars, 100nm.
J. VIROL.
"i
n . t
l?. 4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.489.50.441.83.180.2] [image:7.489.51.445.375.644.2]MORPHOLOGY OF P1 MUTANTS 1125
FIG. 4. Micrograph ofP1am35defectivelysate.Thislysateis characterizedbypolysheath(PS) andnormal
heads,manyof whichareempty due topreparation procedures. Bar, 100nm.
mutants135, 107, and 45 are in another (V), and the restriction fragments that rescue these two groupsofmutants areseparated by other
restric-tion fragments that carry for other genes (see
Fig. 17). Based on complementation tests (49), these mutants in cluster IV represent at least three genes (genes 3, 16, and 20; see Fig. 17).
:.
X..:.*
t.'s~~~~~~~4
;w-~~~~Zk-j--Therefore, the nine mutantsprobably represent at least four genes (the fourth being gene 21), someofwhichcouldcodeforbaseplate compo-nents, butonly oneof whichcouldcode forthe
tube, as it is probably (by analogy with other phages) composed of onlyonepolypeptide spe-cies.
0;..ilt--71.l.
.*l.I..
4
N
FIG. 5. Micrograph ofP1aml80defectivelysateshowing a normal length tail tube (TT) as well as full heads of both PlB and PlS.Bar, 100 nm.
VOL. 45. 1983
I
4
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.489.45.443.75.312.2] [image:8.489.49.443.448.648.2]1126 WALKER AND WALKER
Mutant 180(in cluster V; Fig. 5) shows com-plete heads and tail tubes without sheaths.
Therefore, the defect of am180 is probably in
sheath subunit production. No baseplates were seenattachedtothe tubes. This is probably due to thebaseplates detachingfrom the completed tubes because, in
amber'
P1lysates, contracted tails without baseplates are often seen (47).Alternatively,because am45, which has a defect in either the baseplate or the tube, does not complement am180, it may be that am180 is
polar on am45 and is therefore missing both
sheathsand baseplates.
Mutants19 and 138(bothincistron 5 in cluster VI) show particles which are complete except that all ofthe tails are semicontracted or con-tracted (Fig. 6). The defect of these mutants is
probablyin abaseplatecomponentthat is neces-sary for baseplate stability. That is, either this componentisof minor importance forbaseplate
assembly orthe mutation occurs at a location in the gene that allows formation of an almost
complete polypeptide that does not prevent
baseplateassembly. Ineithercase,tailassembly andhead-tailjoining couldoccur,but the
result-antbaseplate would be unstable and cause tail contraction.Mutantswith mutationsin gene 9 in T4,whichhaveadefectinbaseplatecompletion
(25, 27), showaphenotypesimilar (12)tothatof
these P1 mutants. Because the phenotype of these P1 mutants is different from that of the nine mutants (in clusters IV and V) discussed
above, they could be defective in a different
baseplate component and represent a different gene.This issupportedby the fact that theyare in a different linkage cluster and are physically separated by other P1 genome restriction
frag-ments that carry other genes (see Fig. 17). Mutant 79 incluster VI produces polysheath
and complete heads attached to short tails. Thesetailshave sheathsofvariouslengths
(Fig.
7)withbaseplates and tail fibersattached, andit
is apparent that the defect of am79 affects tail length. In some instances the tails are so short that although a baseplate is seen, it cannot be determined whether any sheath subunits are present.
Mutants 26, 32, and 109 in clusterVIproduce complete heads, polysheath, and unit-length
sheaths (Fig. 8). These sheaths (seen only in lysatesofthesemutantsand mutants27, 39, 53,
22, and 116;nextparagraph)are similarinlength
tothesheathsoncontractedtails,buttheyhave no tail tubeorbaseplate. These structures indi-catethat baseplates and tubes wereformed but subsequently slipped off the sheath. Such a
phenotypesuggestseither thatanunstable base-plate was formed and that the defect of these mutants is in a baseplate component or that thereis adefect at the top of the tail similarto
FIG. 6. Micrograph of P1 aml9 defective lysate
showingparticles with tailsthatalways have
contract-ed sheaths. Bar, 100 nm.
one of the defects described for the following mutants.
Mutants27, 39, 53, 22, and116(incluster VI) producecomplete heads, some polysheath, and
unattachedtails that havebaseplates, tail fibers,
and normal-length tubes(Fig. 9). However, the sheathonthetubes is contracted and hasslipped down from the top of the tubes, and in some cases some of the sheath material is lost. In
addition, unit-length sheaths are also seen, which could arise from the sheaths slipping
completely off the tubes. This phenotype sug-gests that the defect ofthese mutants is in tail
stability. Mutants 30 and 113 (which arein the J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.489.261.444.73.472.2]MORPHOLOGY OF P1 MUTANTS 1127
FIG. 7. Micrographs ofP1 am79 defective lysate showing particles with shorter than normal length tails. Bar, 100nm.
samecistron incluster VII) have a similar
phe-notype (although no unit-length sheaths were
seen), suggesting that their defect is in tail stabilityalso. However,inadditiontocomplete heads, polysheath, and tails with slipped sheaths, these mutants alsoproduce contracted tails with polysheath attached (Fig. 10). These twogroupsofmutations (27, 39, 53, 22, and116, and 30 and 113) are located in two different linkage clusters (Fig. 2); therefore, they are probably in two different genes. In T4, two genes (3 and 15) are required for tail stability. The gene 3 product is added to the top ofthe tube, where itacts astheterminator for sheath
polymerization.Thegene15productthenforms
a stable bond between thetube topand thetop
of the sheathand, atthe sametime, creates the headattachmentsite (25,29). Thetwo(orthree, ifmutations26, 32,and 109areincluded)groups
of P1mutationsaffectingtailstabilitymay
repre-sentgenes with functions similarto those ofT4 genes 3and15.Forexample, theP1 genescould code for (i) the tail cap structure occasionally
seen onP1 tails unattached toheads and (ii)the ringlike structure (seen attached to the base of
the neck on tails where abnormal contraction
has leftthe proximal part of the tube exposed; Fig. 9, inset), which is thought to stabilize the
extended sheath and anchor it to the tail tube (47). In this regard, the contracted tails with polysheath attached seen in the cluster VII mutants could arise if the tail cap was missing and sheath subunits continued to polymerize
onto the tail,past the topofthe tube.
Tail tubes separate from sheaths are seen in lysatesofsomeofthe mutantsin linkage cluster VI,where thetailsare notattached to the heads (Table 1; Fig. 9 and 11). However, since they tend to occurinaggregates, they are not distrib-uted evenly on the grid; thus, their presence may
be overlooked in lysates ofother mutants. This
might explain why none were seen in am32 and am53 lysates. However, with these mutants,
unit-length sheaths with or without tail tubes were seen, which confirms that tubes are made.
Therefore, the absence of tail tube aggregates doesnotchange ourconclusions concerning the
defects of these mutants. Additional structures seeninlysatesofsomeofthe mutants in cluster VI are stacked tails (Table 1; Fig. 6 and 9). These were seen infrequently, and their signifi-cance is unknown.
Mutant 142 (in cluster VIII)
produces
com-plete particles with semicontracted or contract-ed tails (similar to those seen with aml9 andam138), complete heads,andpolysheath.
There-fore, the defect of am142 is
probably
in base-plate formation. Other mutants in cluster VIII(am139, aml40, and am7l) only produce com-plete heads and polysheath. Their defect could thereforebe inbaseplateortubeformation. Itis not known how many cistrons these four mu-tants represent. If they are all in the same
cistron, their defect couldbein
baseplate
forma-tionwitham142 located in the distal partofthiscistron.
Mutations
affecting
head assembly. Mutationsaffecting head assembly are located in linkage
clustersII, V, IX,and X-1. Mutants in cluster II (150, 62, and 108; Fig. 12) and cluster X-1 (16, 64, 136, 131, 43, and 112; Fig. 13) produce complete tails and unattached empty heads in head/tail ratios of 1:1 to 1:3. (A total of245 to 873 particles were counted per mutant,
using
electronmicrographsofrandomlyselectedfields from several preparations.)
In complementation tests (49), aml50 and am62give negative results. Because of this and because the phenotypes of thetwo mutants are
similar, the mutations that they represent are VOL.45,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.489.48.239.73.413.2]1128 WALKER AND WALKER
t~~~~~
s
P
1~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~J1FIG. 8. Micrograph ofP1 am26defectivelysate. Inadditiontonormalfull heads, therearepolysheath(PS)
andunit-lengthsheaths (US)orfragments thereof.The long curvedstructuresarebacterialflagella. Bar, 100nm.
probably in the same gene, which we will refer to as gene 4, a designation originally made by Scott (39) for am7. (Complementation results with am62 and amlO8 were ambiguous;
there-fore, we are unsure whether the mutation in amlO8 is located in this gene.) For similar rea-sons and because the mutants in linkage cluster X-1 are rescued from two adjacent fragments of theP1 genome (see Fig. 17), one ofwhich also contains at least part ofthe P1
ci
geneand the otherof whichisflanked bya0.62-kilobase-pair (kb) fragment which contains at least part of gene10, themutationsinlinkage clusterX-1 areassigned to asingle gene(gene 9, adesignation originally made by Scott [39] for aml6), al-though there is a possibility that these mutants could represent more than one gene.
The empty heads produced by gene 9 and cluster IImutants are similar (insize and in the thickness of the capsid shell) to empty mature heads seen in
amber'
lysates. Therefore, they are presumed to be mature heads that arelack-ingDNA, rather than proheads.
InT4, mature, empty, unattached heads (and complete tails) are produced by mutants with defects either in various head completion genes
(8, 11, 26)orin gene49. Headcompletiongenes are involved in neckassembly (gene 13), DNA
packaging (genes 16 and 17), or unknown func-tions (genes4and65). Gene49codesforDNase and is required for DNA packaging (14, 33).
Heads of mutants in these genes are empty,
eitherbecause they were not filled with DNA or because they were unstably filled and subse-quently lost their DNA (8, 17). Genes in other phages that are involved in head completionare genes4, 10, and 26in P22 (6, 36), which code for threeminor structural proteins required for head stability, and gene D inX(24), which codes fora protein that is added to the mature capsid shell and whose presence is also required for DNA
cutting (4, 23).
From theirphenotype, the P1 gene 9mutants could bedefective in one of these functions. In
thisregard, Sternberg (personalcommunication)
recently foundthat,unlikemutantsin genes 4, 8, and23, gene 9 mutantscannotcleaveP1 DNAat pac, the site for initiation of DNA packaging
(see Fig. 17); therefore, like genes 2 and 3 in phage P22 (31), the P1 gene 9product is involved in the recognition and cleavage of P1 DNA at pac.With otherphages, conversionofproheads to mature heads usually occurs concomitantly
with DNApackaging. Gene 9 mutants apparent-ly cannot initiate DNApackaging. Therefore, in P1 either conversion of proheads to mature headscanoccurindependently of DNA
packag-ing or P1 proheads resemble mature heads in size and shell thickness.
Thecluster IImutantscouldalsobedefective
in one of these functions (except for DNA
cleavage because, as mentioned above,
Stern-bergfound thatclusterIImutants cancleave P1 DNAat the pacsite).
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.489.56.447.73.300.2]MORPHOLOGY OF P1 MUTANTS 1129
PS
,
* . '-4
.4
IN .;.
t '.4
'.'
444
r .JE ~ ~
i~S- T
[image:12.489.43.446.73.438.2]taST~
FIG. 9. Micrographof Pl am27defectivelysate showingseveralkinds of tailstructures. Note the tail tube
aggregate(TT), polysheath (PS),tails withslippedsheaths(TSS),and stacked tails(ST).Inset showsanamber' Pt particle fromaCsClgradient;afterprolongedexposuretoCsCl,there is abnormal tailcontraction,i.e.,the tailsheathhascontracted and sliddown thetube,butaringlikestructurewithorwithoutsomesheathsubunits remainsatthejunctionbetween theproximal end of the tube and the neck(see text).Bar,100nm.
FIG. 10. Micrograph of Pt am3Odefective lysate.
Polysheathappearstobe attachedtotheproximalend of thischaracteristicallycontractedtail, which other-wiseappears tobenormalin having bothabaseplate and tailfibers. Bar, 100nm.
Like the mutantsinlinkageclusters IIand X-1,a single mutant (amll5)in cluster V and the
mutant (aml3) that represents cluster IX (i.e.,
gene 8) produce complete tails and unattached
empty heads. With aml3, the head/tail ratio is 1:2 (1,967 particles were counted). However,
with amll5, very few heads were seen (the
head/tailratiowas1:30,with431particles
count-ed), and input phage could account for these
heads. Attached tothe tailsof both amll5 and
amt3 mutants are structures resembling the neck and head-neck connector (Fig. 14). The neck and head-neck connector are part of the head.Thissuggeststhat the headswere
original-ly assembled, filled withDNA, andattached to
the tails, but were unstable and subsequently eitherdisintegrated (amllS)ordissociated from
p
'-p--,-, -Lt-
*X-VOL. 45,1983
I
Z"
on November 10, 2019 by guest
http://jvi.asm.org/
[image:12.489.49.229.503.611.2]1130 WALKER AND WALKER
FIG. 11. Micrograph of P1 amll6 defective lysate showing aggregates of tail tubes. No single tube is longer thannormal length. Bar, 100 nm.
the tail, leaving behind at least some of the neck andhead-neck connector (aml3). Such instabil-ity could be due to the absence of a minor head
protein,e.g., aminor head shell protein (amllS) or ahead-neck connectorprotein(aml3).
Alter-natively, because so few heads are seen in amllS lysates, amllS could bedefective in the major head protein. In thiscase, the structures seenwouldnotbe a neck andhead-neck connec-torbutanaberrant structure similartothat seen in T4(calledtheextension[8]),which consistsof anaberrant accumulationonthe topofthe tail of five proteins normally assembled into the head aspart ofthe neck andhead-neck connectoror added to thehead-neckconnectorafter head-tail
joining.
Mutationsaffecting particle maturation. Muta-tions affecting particle maturation (47, 141, 34, 128,132, and 57) comprise linkage clusterI.The mutants in cluster I donot show complementa-tion(49) andarerescuedfroma0.92-kb restric-tion fragment of the P1 genome flanked by
fragments carrying other genes (see Fig. 17);
[image:13.489.57.450.71.214.2]therefore, these six mutations are probably in the same gene (gene 1). Particles produced by the six mutants are complete except that, inall of themutants except57, heads tendtobeempty andthetails are in various stagesofcontraction (Fig. 15). Mutant 57 produces completeparticles withat leastpartially filled heads and extended sheaths (Fig. 16). These phenotypessuggestthat thedefectivepolypeptideallowsassemblyofthe
FIG. 12. Micrograph of P1 amlO8 defectivelysate.Thetails appeartobenormal;heads,not seenhere,are
alsoproduced. Bar, 100 nm.
J.VIROL.
-V
Gkll-.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:13.489.57.450.430.643.2]MORPHOLOGY OF P1 MUTANTS 1131
FIG. 13. MicrographofP1am64defectivelysate.Theheadsarethesamesizeasmatureheads,buttheyare
always empty;thetailsarenormal, their contractionbeinganartifact ofpreparationprocedures. Bar, 100nm.
complete particles but that these particles are either morphologically complete but noninfec-tive(am57) or unstable. Morphologically normal
particlesareproduced by other phages, e.g., P22
-RI i.^ .. .
._l S ...
FIG. 14. Micrograph of P1 aml3 defective lysate. Tailsarenormalexceptforaknobatthe proximal end (arrow). Bar, 100nm.
mutants defective in genes 7, 16, and 20 (6,36) andT4mutants(11, 12, 26) defective ingenes2, 4, 11, 12, 50, 64, and 65 (which produceat least somemorphologically normal phages). Genes 11 and 12 are tail genes, coding for a baseplate component and short tail fibers, respectively. Allofthe remaining gene products are incorpo-ratedinto the head at early stages of assembly, but theirfunctions vary; e.g., some are involved inthe DNA injection process, and some (in T4) are involved in cleavage of head proteins. The P1 mutantscould have one of these head gene defects (although it isnotknown whetherthere is cleavage of Pl head proteins). Alternatively, because of the instability of the tail, they could have a defect in a tail gene, e.g., a baseplate gene. The difference observed between am57 and the other mutants in this cluster could be
explained by the am57 mutation being in the distal part of the gene togive a more complete
polypeptide andhence a more stable particle. In supportof this is thefact that am57 is located at one endofthe clusterwith respect to the other five mutantsexamined (Fig. 2).
Mutationsaffecting host celllysis. Cells infect-ed separately with eight mutants that cannot causecell lysis (gene 17, linkage cluster III, or gene 10, cluster X-2) or that cause premature cell lysis(gene2, cluster IV) were examined by electronmicroscopy at the incubation time (110 min) at which lysis would be complete in cells infected with lysis-positive mutants.
VOL.45,1983
4
.1
4. -f
... .7 -,-.
IS6
on November 10, 2019 by guest
http://jvi.asm.org/
[image:14.489.45.235.430.639.2]1132 WALKER AND WALKER
FIG. 15. Micrographof P1 am47 defective lysate. Particles appearto be normalexcept forempty heads (mature size); normal tailsareinvariousstagesofcontraction. Bar, 100nm.
Mutants (23, 144, 73, and 134) having
muta-tions ingene17 showedintactcellsasexpected
and,occasionally, somecellsthathadlost some
oftheircontentsbutwerefull ofphageparticles. Veryrarely,somecomplete phage, which could
have been input phage, were seen outside the
cells. These resultsareconsistent with the find-ings(50) thatartificially lysed cellsinfected with these mutants produce viable phage particles and give positive results in serum blocking as-says and that phage particles are seen in thin sectionsof unlysed cells infectedwitham144(or am2O, which has a mutation in the same gene)
and support the conclusion that these mutants are defective ina lysisfunction.
Mutants (178 and 105) having mutations in
gene 10 showed intact cells and, very rarely, some complete phage, which were probably
inputphage.These mutantsdidnotshowserum
blockingpower, evenaftersonicationtodisrupt the unlysed cells and to release any tail fiber antigenthatmightbepresent. These results are
consistent with thefindings (50) that artificially lysed cells infected with these mutants do not
produce viable phageparticles andthat nophage
particles are seen in thin sections of unlysed,
infected cells. Therefore, theresults are
consis-tent with the conclusion (50) that gene 10 mu-tantscannot synthesize late proteins (including
thelysisproteins)and that gene 10 may codefor
aregulatoroflate gene expression forP1. Mutants (31 and 56)having mutationsin gene 2 showed very few particles, and these were complete (except that the presenceoftail fibers could not bedetermined). These mutants cause premature lysisand may have adefect in alysis regulator (50). In addition to very few particles
being seen by electron microscopy, serum blocking assaysarenegativewith these mutants, and theburst size is low (50). Theseresultscan all beexplained by too few phageparticles being producedby the time that lysis occurs.
DISCUSSION
Gene designations of mutations affecting P1
morphogenesis. We determined the phenotypes of 47 Pl amber mutants by use of electron microscopy and serum blocking assays. Eleven J. VIROL.
.i6&
on November 10, 2019 by guest
http://jvi.asm.org/
[image:15.489.55.446.72.389.2]MORPHOLOGY OF P1 MUTANTS 1133
FIG. 16. Micrograph of Pl am57defectivelysate. Particles appear normal; irregular outline of heads may possiblybe due toflattening as a consequence of being only partially filled with DNA. Bar, 100 nm.
mutants (Fig. 17) showed head defects, thirty showed tail defects, and six had a defect in
particle maturation that could be either in the
head or in the tail. When we consider the
unambiguous results from complementation tests, the genetic and physical positions of the mutations, and the phenotypes of the mutants, we can assign most of these mutations to genes
(Table1 andFig. 17). Exceptions are mutations 108, 33, 66, 74, and 142, which could be located in different genes than those indicated (i.e., genes4, 19, 20, and 26).am108gives ambiguous complementation test results with other gene 4 mutants and thus may be in a different gene. Results of serum blocking assays did not
distin-guish betweenthe possibilities that mutants 33, 72,and66could bein the same gene (gene 19) or indifferent genes.Although mutations 139, 140, and 71(incluster VII; gene 26) result in a slightly
different phenotype than that ofam142 (Table 1), these four mutations could be in the same gene because they are located in a relatively shortregion of the genome (Fig. 17). However, itis not known whether mutations 74 and 49 are in the same gene (gene 20) because, although they show similar phenotypes, they give ambig-uousresults in complementation tests (49), and thepreciselocation of am74 on the physical map is unknown.
Mutationsaffecting head morphogenesis. As a resultof these gene designations, it is seen that the11 mutations causing head defects are
locat-ed in four(orat mostsix) genes(genes 4, 23, 8,
9, and perhaps the particle maturation gene 1 andagene represented by amlO8). In T4, head
assembly involves the products ofat least 20 genes, about half of which constitute the
fin-ished particle (53). P22 head assembly requires
theproducts of11 genes,about6ofwhicharein the finished
particle
(53). With its head shell, head-neck connector, and neck structures, P1heads are at least as complex as P22 heads. In supportofthis, results fromgel
electrophoresis
examination of P1
particles
indicate thatthey
consist of28 proteins, of which 15 are in the
head,9arein thetail,and 4areeitherin the head orthe tail(51). Ofthe 15(to
19)
headproteins,
4 are coded by a region ofthe genome, between mapunits24 toabout 32,thatisnotessential for particle formation. However, it is not knownwhether any of the remaining 11 to 15 head proteins result from cleavage orfusion of
poly-peptides duringheadassembly. In
addition,
it is notknown whether thereareanyproteins
(suchas the scaffolding protein in P22) that are not componentsofthe finalvirionstructure. Never-theless, it is likely that more than four (to
six)
genesarerequiredfor P1 headassembly. Inthis
regard it is of interest to note that
adjacent
to head genes4and 8(Fig. 17),thereareregions
of the genome for which no functions are known and which might carry further head genes. Razza et al. (38) have described temperature-sensitive mutants in two new genes (genes 13 VOL.45,1983on November 10, 2019 by guest
http://jvi.asm.org/
[image:16.489.47.441.74.300.2]1134 WALKER AND WALKER
and 14) that are locatednear gene 4 and that are required for lytic growth ofP1. These could be head genes. (Another new gene described by Razza et al., gene 15, is located near genes 22 and 23 and could be either a head or a tailgene.)
Onlythree phenotypes are produced byamber mutations in the four head genes. These pheno-types are (i) complete tails and separate empty
heads (genes 4 and 9), (ii) complete tails with necks and head-neck connectors and separate empty heads (gene 8), and (iii) complete tails with necks and head-neck connectors (or exten-sions) (gene 23). As mentioned above, gene 9 mutants are defective in a function required for DNA cleavage at pac (N. Sternberg, personal
communication). The defect of cluster II mu-tantsisunclear, but could be in a minor protein
(suchas aneckprotein)orinaproteinrequired
forDNApackaging(other than thatrequiredfor DNA cleavage at pac). The second and third phenotypes suggest either a defect in a minor head protein that leads to a filled, attached, but unstable head or a defect in the major head
proteinthat leads to anaccumulation of the neck and head-neck connector proteins at the top of the tail toform a structure called the extension by Coombs and Eiserling (8). Head-defective phenotypes not seen, which might represent
headgenes not as yetdetected, are asfollows.(i) Proheads. Unless P1 proheads resemble mature heads, none were seen. Other phages produce
proheadsin defectivelysates ofmutantshaving
mutations representing various different genes. Therefore we would expect to have more than one gene in which the mutant allele results in proheadproduction. In thisregard, it could be, asmentionedabove, that Plproheads are unsta-ble and can convert to mature heads
indepen-dently of DNA packaging. (ii) No heads. In otherphages, mutations in themajorheadgene result in mutants that do not produce heads.
Although P1 mutants that do not produce any heads at all have not been found, the gene 23 mutant produces very few heads(1 head per 30
tails), so thisgene might code forthe major P1 headprotein. (iii) Spiral head structures, such as those seen in mutants of P22 defective in scaf-foldingprotein (28) orin mutantsof X defective either in the minor headproteinsBand C(24) or in the scaffolding protein Nu3 (37). (iv) Poly-heads, such as those seen in T4 mutants defec-tive either in the core protein, an internal pro-tein, the vertex protein, or in proteins required for correct capsid initiation (12, 13, 30, 40, 56). Polyheads are also seen inKmutantsdefective in genes B and C. (v) Particles with only small heads(P1S particles). AuniquepropertyofPl is that, in addition tonormal, infectious,
big-head-ed (P1B) particles, it also produces 10 to 20% noninfectious, small-headed PlS
particles (47).
It would be useful in studying head size determi-nation to have a mutant that produces P1S
particles only. So far, the only Pl phage that produce an increasedproportionofP1Sparticles (i.e.,50% P1Sand50%PlB particles) are those that are deleted for aregionof the genome that is not essential for particle morphogenesis (i.e., mapunits 24 to 32) (Fig. 17) (20). The determi-nantinthisregion for thischange in thePlS/PlB
ratio is designated vad for viral architecture
determination (20).
Mutationsaffectingtailmorphogenesis.The 30 mutations causing P1 tail defects are located in atleast 12 genes (genes 19, 20, 16, 3, 21, 22, 5, 6, 24, 7, 25, and 26; Fig. 17).There couldbeup to five additional tail genes (represented by mu-tants 72, 66, 49, and 142and perhaps including gene 1,which is involved in particle maturation).
In T4, tail assembly (including tail fibers)
re-quirestheproducts of29 genes,andall but 7of thembecome partofthefinalstructure. The Pl tail is less complex than that of T4 (i.e., the
baseplate is thinner and less well defined). Re-sults from gel electrophoresis studies indicate
that theP1 tail iscomposed of between9and13 proteins (51). However, it may be that some of thebaseplatecomponents are small andrequired
inlowquantitiesand so were notresolvedbygel electrophoresis.
Ofthestructuraltail genesinT4, 1 genecodes forthetube;1forthesheath;4forthe tailfibers;
14forthe baseplate; and 2forthe structures that add to thetopof thetail tostabilize it.The 12 to 17P1 tail genesconsist of1 (gene22)coding for
thesheath;between 6(genes 20, 3, 16,21, 5,and 26) and 10 (the additional genes being gene 1, gene 24, and genes represented by am49 and am142) coding for baseplate ortube production (of which only 1 would code for tube produc-tion); 2or3 (genes7, 24,and25)codingfor tail
stabilization; 1(gene6)affectingtaillength;and 1(gene19,represented by am72)or more
(repre-sentedbyam33 andam66)coding fortailfibers. In addition, Razza et al. (38) found two genes (genes 11 and 12, represented by temperature-sensitivemutants) that are located in thevicinity
of gene 19 and maycodefor tail fiberstructural proteins. In summary, we may not know for all of the tail genes which code for structural pro-teins and which codefor nonstructuralproteins,
but mutations affecting each of the tail compo-nentsvisible by electron microscopyare repre-sented (with a possible exception of the tail tube).
Gene19 mutants do not have tail fibers and do notshow serumblockingpower(forthis discus-sion we shall assume that mutations33, 72, and 66 are all in gene 19, although this may not be so). Therefore, gene 19 isastructuralgene that
probably codes for the distal part of the tail J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
3 ~J- c
FIG. 17. Map of the bacteriophage P1 genome, modified from that ofYarmolinsky (57). The inner circle represents the physical genome, divided into the numbered digestion fragments generated by restriction
endonuclease EcoRI (2). Genetic symbols within this circle represent functions not required for phage morphogenesis (except pac); for acomplete list of these functions and their references, see Yarmolinsky (57). pac is the site where DNA packaging initiates (arrow indicates direction ofpackaging). Other symbols and functions: c genes, establishment and maintenance oflysogeny(c6 and c8 were found in phageP7); inmmCand
imml, bipartite immunity system; resand mod, restriction and modification; dar, defense against restriction; vad, viral architecture determination; gta, generalized transduction affected; lyd, lysis delay; cin, C loop inversion; virs, virulence;sim, superimmunity; ant orreb, antagonism or bypassofcl repressor; par, plasmid partition; incA and incB, plasmid incompatability; rep, plasmid replication; ori, origin of replication; cop,
plasmidcopynumber;ban, dnaB analog; loxP, locus ofcrossover inplasmid; andcre,site-specific recombina-tion. Outercircle, 100 map units (equivalentto percentof the 90-kb genome). On this circle, C denotesthe invertible C loop; ISJ denotes insertion sequence 1. Between the outer circle and the arcs are numbers representing amber mutations. Thelocations of these amber mutations on the physical map were determined by marker rescue experiments, using cloned fragments ofP1 DNA (21, 34, 38, 43, 45). The arcs denote extentof
linkage clusters (roman numerals). Numbers outside these arcs denote genes (gene numbers 5, 6, 24, 7, and 25 are repeated above their relevant functions). A query denotes the uncertain gene designation of am74.
Diagrammaticrepresentations of theamber mutant phenotypes and functions of the genes are noted. Asterisks besidebaseplate notations mean that one of these genes could code for the tail tube. Imprecise locations of genes oramber mutationsare denoted byparentheses around the relevant symbol or number. Genes 11 through 15 are represented by temperature-sensitive mutations only (not shown) and have not been phenotyped. Rescue tests and two-factor crosses have shown amber mutant 75 (gene 16) to be located between mutants 74 and 35 and not around map unit 55 as indicated previously.
1135
on November 10, 2019 by guest
http://jvi.asm.org/
[image:18.489.51.438.48.448.2]1136 WALKER AND WALKER
fiber. Mutant 72 is missing a
105,000-molecular-weight (MW) protein that, unless it is a fusion product, is theproduct ofgene 19 (unpublished
data). Toussaint et al. (45) located the gene 19
mutations within or near the P1 C loop (an invertible DNA region) because (i) this is the
only regionofhomology shared with phage Mu
(i.e., the Mu G loop) and (ii)the
amber'
alleles of theseP1 mutants are rescuedfrom Mu. The Pl C loopis 3 kb long and flanked by two 0.62-kb inverted repeats. The 105,000-MW proteincoded bygene 19 would require a coding length
of2.9 kb ofDNA. Gene 19 probably does not extend more than halfway (1.5 kb) into the G loop (by analogy with Mu [46]); therefore, the remaining portion (1.4 kb) of the gene would include one of the 0.62-kb inverted repeats and extend at least 0.78 kb outside the inverted repeat. If the105,000-MW proteinisnot a fusion product, even more of gene 19 could be located
outside the C loop. This is because
amber'
alleles of two Mu genes, one of which (U) is wholly within the G loop and another of which (S) ispartially within theG loopand partiallyin theotregion(16), arerescued from P1 (45). This suggests that there is another gene inadditionto gene 19 in the P1 C loop.Products ofboth the U andSgenes arerequired forMuadsorption(46);
therefore, in P1 thetwoanalogous genescould
codeforthedistal (gene 19) and proximal parts
oftheP1 tail fiber.
Thefollowing findings support the suggestion
that gene 19 is partially outside theC loop and that it includesoneoftheinverted repeats. The
P1-related phageP7,whichhas a0.2-kb deletion
(orsubstitution) within therightinverted repeat (7), produces a 100,000-MW protein instead of the105,000-MWP1tail fiberprotein (51). (These findingsalsosuggest that gene 19 extends to the right rather than to the left of the C loop.)
Transcriptionofgene 19 is leftward fromam49
(becauseam49ispolarongene19;Fig. 2). Also,
no polypeptide fragment of the 105,000-MW protein is seen after sodium dodecyl
sulfate-polyacrylamide
gelelectrophoresis
ofam72(un-published data). Therefore, unless the amber fragment has beendegraded, thismutation must be near the end ofgene 19 that is proximal to wheretranscription isinitiated.
Themodelofgene 19asbeingpartiallywithin and partially outside the C loop is consistent
with thefollowingfindings: (i) Mu and P1 have
partial serological cross-reactivity (45), and (ii)
the host range specificities of these two phages
are within(or near) theseinvertibleregions (45,
46). By analogy with T4 (3), these host range
specificitiesareprobablylocated in the tail fiber genes of thephages.
Assembly of P1 particles. By examining the structures obtained in defective lysates, it is
seen that P1 heads and tails can be assembled independently. Thissuggeststhat, like T4, these Pl structures have independent assembly path-ways. However, in P1,tail fibers are often seen attached to headless tails. Therefore, unlike T4 tail fibers, P1 tail fibers can attach to the tail before head-tail joining occurs. (This had al-readybeen suspected because the P1 tail fibers are not longenoughto reach from thebaseplate
to thehead, such a length being required for the addition of tail fibers to T4particles.)
Distribution of mutations. TheP1 amber muta-tions affecting particle morphogenesis are not equallydistributedamong thedesignatedgenes. Forexample,including mutationsnotstudiedin the work reported here (Fig. 2), there are 13
mutations in gene 1 andonly 1 mutation ineach
ofgenes 22, 23,and8. We do not knowthe sizes ofthesegenes,butit maybethat there are "hot
spots" of mutation (obtained by
hydroxyl-amine), as wasfoundin P22 (36). In support of
this, we do know thatthere are 10mutations in gene 10,whichcodesforaprotein of64,000 MW (18)involvedin late geneexpression (50), and a
maximum of 3 mutations in gene 19, which
codes fora 105,000-MW tail fiber protein. The existenceofhotspotsof mutationcould explain
why some of the head-defective phenotypes described abovehave not yetbeen seen.
Correlation ofphenotypes withthe P1 genetic
andphysical maps. The 103P1 ambermutations
have been assigned (49) to 10 linear arrays or linkage clusters (I through X; Fig. 2). Because
complementation tests in liquid often gave
am-biguous results and not all mutants were tested in allcombinations, only some of themutations were put into groups that showed no comple-mentation. The 55 amber mutant phenotypes (i.e., including mutations ingenes17, 2, and 10)
that we determined correlate with the linear arrays ofthecorresponding mutations. For
ex-ample,inlinkage clusterVI, mutants 26,32,and 109showonephenotypeand mutants27, 39, 53,
22, and 116 show another. In addition, except
for mutants 180 and 45
(which
may be polarmutants), allmutants tested that were found not tocomplement (Fig. 2) showed the same
pheno-type.
We previously suggested (49) that the
phe-nomenon of linkage clustersmight resultfroma variety ofunderlyingcauses, such as
recombina-tion hotspots, long segments of silent DNA, or
regions for which there were no known muta-tions. The physical position of the mutations
comprising the linkage clusters is now known
(Fig. 17)(57), and it can beseenthattheclusters consist of single genes or groups of genes and that the demarcations between some of these linkage clusters (i.e.,
II/IlI,
VI/VII, VII/VIII,
VIII/IX,
and X-2/X-1)are due torecombinationJ. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
MORPHOLOGY OF P1 MUTANTS 1137
between these genes or gene groups. The reason why some linkage clusters are comprised of single genes and some are comprised of gene groupsis unknown, unless hot spots of
recombi-nation are invoked. Other demarcations be-tween linkage clusters, such as I/IT, III/IV, IV/V, V/VI, and IX/X-2, are due to long seg-ments of DNA that either contain other genes (e.g., coding for immunity) or for which no mutations are known, or a combination of these two reasons. (It may be that there are no long segments of silent DNA because, for example,
some morphogenetic genes [especially head
genes] are still undetected.) Between linkage
clusters X-1 and I (i.e., at the ends of the vegetative map), there is a shorter segment of DNA that codes for part of the P1 immunity system. However, this region has also been shown tocontainarecombination hotspot(loxP [44]), whichalone could accountforthe
demar-cationbetween these two clusters.
Clustering of functionally related genes. The
physical andgeneticmaps ofP1 show consider-ableclustering ofgeneshavingrelatedfunctions. Suchclusteringcould beadvantageousfor
regu-latingtheassemblyofsubstructures, such as,for example, baseplates. We do not know whether gene 1isaheador atailgene.Excludinggene1,
theknown tail genesarelocated in three regions. The approximate physical positions of these regions are map units 34 to 49, 56 to 59, and 78 to
84.5. Within these regions, some similar func-tions are located together. In the first region there are three to five tail fibergenesand three to four baseplate (or tail tube) genes. In the
secondregionwefoundonetailsheath geneand
one baseplate (or tail tube)gene. It may be, by analogywith T4 (54), that the single P1 tail tube gene is located in this region beside the sheath gene. In the third region we found one or two
baseplate genes,atail lengthgene, twoorthree
tail stability genes, and one or two baseplate
or tail tube genes. In this region the baseplate
genes are separated by other tail genes. How-ever, unlike genes 3 and 15 of T4 (54), the two to three P1 tail stability genes are located
to-gether.
The fourto five known Pl head genes are in
four regions ofthe genome. This would argue against clustering of functionally related head genes. However, some expected P1 head genes have not yet been detected and could be located in two main regions for which there are few known genes. One of these regions comprises map units 10 to 20 and could theoretically code for about 10 genes, including head gene 4, the head gene represented by am108 (if different from gene 4), and genes 13 and 14 found by Razza et al. (38). The other region comprises map units 84.5to 94,which could code for 9 or
10 genes,including head gene 8. Inaddition, it is interesting to note that head gene 9, which is adjacent to pac, the site at which DNA packag-ing is initiated, has nowbeen found tocode for P1 DNA cleavage at this site (N. Sternberg, personal communication).
Inaddition to lysis gene 17, P1 probablyhasa second lysis gene (lyd[20]), analogous to the S gene of X, located somewhere between map units 24 andabout 32.Therefore, thesetwolysis
genes and a lysis regulator gene (gene 2), al-though not all adjacent toeach other,arelocated within a region spanningless than 15% of the P1 genome (map units 20 to about 33).
Gene 10,which regulates late gene expression (50), is located approximately between map units 94 and 96. Gene 10 is probably analogous to gene Q in X, which is adjacent to genes (O and P) involved in DNA synthesis. If similar clustering occurs in P1, such DNA
synthe-sis genes could be to the left of gene 10, in a region for which no genes have as yet been found.
We do not know thefullextentofclusteringin P1 because of the regions of the genome for which no genes are known. That is, the regions
mentioned above (between genes 8 and 10 and genes1and 4) and anotherregion approximately
between map units 67 and 76.
In addition, very little is known about the transcription patterns in P1, so wedo not know how much the clustering offunctionally related genes is related to their cotranscription. (An exception is that am49 is polarongene 19, and the gene it represents is therefore probably cotranscribed withgene 19.)
Genomeorganization of P1comparedwith that ofotherphages. The functionally related genes
ofP1 arenotclusteredasconservativelyasthey
are, forexample, in X, in which all head genes arein oneregion,adjacent to all of thetailgenes in another region, and in which all the genes
determiningotherfunctions(suchasintegration, excision, recombination, gene regulation, repli-cation, and lysis)are in the remaining region of
the genome (10). The organization of the P1 genome more closely resembles thatofT4 (54), inwhich the tailgenes arelocatedin atleastsix regions and the head genes in at least five regions. InT4, thesemorphogenetic gene clus-ters areinterspersedwith genes concernedwith, forexample, DNA metabolism. InP1,
morpho-genetic gene clusters are interrupted by many genesthat are expressed in the prophage, such asthe genesfor establishment andmaintenance
of lysogeny, plasmid replication, immunity, or restriction and modification of DNA. As for the
precisefunctions of the genes forwhichwehave amber mutants,furtherinvestigationswith other techniques are required.
VOL. 45,1983