0022-538X/80/08-0542/05$02.00/0
NOTES
Comparative Analysis of the Genomes of Feline Leukemia
Viruses
ZEDAF. ROSENBERG,"2FINN S. PEDERSEN,2" AND WILLIAM A. HASELTINE'2*
Departmentof Microbiology,HarvardSchoolof PublicHealth,'andSidney Farber CancerInstitute,
CharlesA.DanaCancerCenter,2Boston, Massachusetts, 02115, and Department of MolecularBiology,
University of Aarhus,Aarhus,Denmark3
The genomesofseveralstrains of felineleukemiavirus (FeLV) werecompared
by two-dimensional polyacrylamide gel electrophoresis of the large RNase
T,-resistant oligonucleotides ofthe 70S RNA. Differences between each strain of
FeLV tested were detected by this method. We estimate that the degree of
sequenceidentitybetween the viruses is: FeLV A(Glasgow-1)toFeLV B
(Snyder-Theilen),52%;FeLV A (Glasgow-1) toFeLV C (Sarma), 66%; FeLV B
(Snyder-Theilen) toFeLV C (Sarma),37%. The fingerprints of two independent isolates
ofFeLV strainsofsubgroupA(Glasgow-1andRickard) weredetectablydifferent.
We conclude that the RNase
T,
oligonucleotide fingerprint pattern provides a usefultool for identification of FeLVstrains.Feline leukemiaviruses (FeLV's) are a hori-zontally transmittedgroupof type Cretroviruses
that causelymphoma,
lymphoblastic leukemia,
fibrosarcoma, and
aplastic
anemia incats(1, 2).
This class of viruses ishorizontally transmitted,
as demonstrated by epidemiological studies of multiple-cathouseholds(3)andbyexperimental
infections under laboratory conditions (4, 5).
TheFeLV's have been classified into three
dis-tinctsubgroups,A (FeLV A), B (FeLV B), and
C (FeLV C), on the basis of
type-specific
neu-tralizationandinterferencetests (14, 15).Tests ofhomology by nucleic acidhybridization
showthat these viruses are closely related. At least
85%of the
complementary
DNApreparedfromanyone of thesubgroups willanneal toany of
theothersubgroups. Moreover,themelting
tem-perature of the hybrids formed between the
complementaryDNA ofonesubgroup and the
RNA of another is within 1°C of that of the
homologoushybrids (10).
Although the FeLV's appeartobe veryclosely
relatedbyhybridization techniques, they differ
markedlyin their abilitytoinfectheterologous
celllines.SubgroupAvirusesreplicateprimarily
in feline cells, whereas viruses ofsubgroups B
andCreplicate additionallyin humanand mink cells (8, 16). These viruses also differ in their
naturalpatternsofoccurrence. All field isolates
of FeLV contain FeLV A either alone or in a
mixture ofsubgroups (8, 15). FeLV C is rarely
isolated innature and hasneverbeendetected
in the absence ofother FeLV subgroups(7). In diseased cats under natural conditions, no ob-served correlation exists between the FeLV
sub-group isolated and the type of disease that is
produced(7).
The differences in the biology of the FeLV's
motivatedus toinvestigatethe structure ofthe
viral genomesusing a method capable of
detect-ing differences between closely related viruses.
For thisstudy, we used a high-resolution two-dimensional RNA fingerprinting technique
which permits direct comparison of
approxi-mately 15% of the viral genome (11, 12).
Ge-nomic 70S RNAwasprepared fromvirus strains
propagated in fibroblasts. The fingerprints of
three prototype strains, FeLV A (Glasgow-1),
FeLV B(Snyder-Theilen),andFeLVC (Sarma),
areshowninFig. 1.
About60largeRNase
T1-resistant
oligonucle-otideswereidentified in thefingerprintof each
virus.Theuniqueoligonucleotides areidentified
asthosespecies that migrate slowly in the
sec-ond dimension and that appear to be present in
unimolar amounts asjudged by the amount of
radioactivity in the gel corresponding to the
individualoligonucleotides.Ourprevious studies
indicate that the labeling methods used here
resultinuniformlabelingof theoligonucleotides
(11, 12). Thecomplexity of thefingerprint
pat-ternsfor these threevirusesis that anticipated
ifeach isolatewere apuresinglespecies andnot
amixtureoftwo or morevirusstrains.
542
on November 10, 2019 by guest
http://jvi.asm.org/
NOTES 543
,,
-4
:w vl
*M
*o*O
A
ow I
B
C
FIG. 1. Autoradiograms of the two-dimensional
RNA fingerprints ofFeLVsubgroups. Viruses and
cellswereprovided by0.Jarrett. FeLVA
(Glasgow-1)wasisolatedfromacatwith spontaneous
alimen-tary lymphosarcoma (9). FeLV B (Snyder-Theilen)
waspurified from amixture of subgroupsAand B
presentasthe helperviruses in the Snyder-Theilen
strain ofFeSV (14). FeLV C(Sarma) waspurified
fromamixtureof subgroupsA and C isolatedfrom
FL-237 cells (14). Viruswas harvestedfrom
super-natant culturefluidscollectedfrom confluentroller bottlesonce ortwicedailyand concentratedby
ultra-centrifugation.Viruspelletswerethensuspendedand
pelleted through30%sucrosein TNE (50mMTris,
pH 7.5, 100 mMNaCl, 1 mM EDTA), followed by
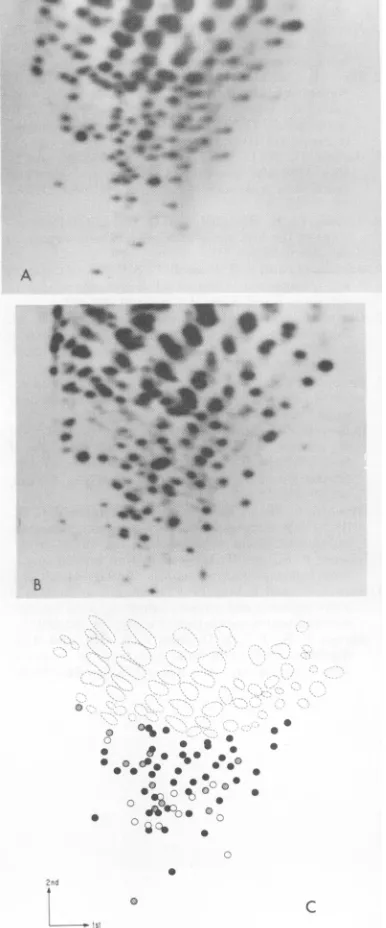
Inspection ofFig. 1 revealssome similarities
and somedifferencesinthe fingerprint patterns ofeachstrain. To obtainqualitative estimates of thesimilarityinthefingerprints, equalamounts of3P-labeled oligonucleotidesderivedfrom each
virus were mixedinpairs and then analyzed by
two-dimensional gel electrophoresis. To
deter-mine thenumber ofunique oligonucleotides that
hadthesame electrophoretic mobility, the fin-gerprintpattern ofthe mixture was then com-pared withindependent
fingerprints
of each vi-rus.Aschematicrepresentation ofonesuch
exper-imentis picturedinFig. 2Afor FeLV A
(Glas-gow-1) and FeLV B (Snyder-Theilen). The S unique
oligonucleotides
that have thesame mo-* bility in the two virus strains are indicated as solid circles, those that areunique to FeLV A (Glasgow-1) are indicated by open circles, and * those thatareunique
toFeLVB(Snyder-Thei-len) are indicated
by
shaded circles. Of the 61 uniqueoligonucleotides
of FeLVA(Glasgow-i),
32(52%) comigrate with those ofFeLV B
(Sny-der-Theilen),
and32of the53unique
oligonucle-otides of FeLV B
(Snyder-Theilen) comigrate
withthoseof FeLVA
(Glasgow-i).
The extent of comigration permits a rough
estimation of the sequence
identity
of the two viral strains (Table 1). The estimate assumes that the sequenceidentity
of theoligonucleo-tides is
representative
ofthe sequenceidentity
of the entiregenome.Calculationsbased on
co-migrationmayslightly overestimate the degree
of sequence identity, since
oligonucleotides
whichcomigratemayhave thesamebase
com-digestionwithproteinase K (E.M. Biochemicals). 70S
viral RNA-containing fractions,pooledfromalinear
15 to30%sucrosegradientin TNEplus 0.2% sodium
dodecyl sulfate,werepassedover an
oligodeoxythy-midylic acid-cellulose column (Collaborative
Re-search, T-3). Thepolyadenylic acid-containing 70S
RNAwaseluted with10mMTris-hydrochloride(pH
7.5),precipitated twice with ethanol, and suspended
in 20 mMTris (pH 8.0)-2 mM EDTA. In vitro
32p
labelingofviral RNA wasperformed according to
themethodof Pedersen and Haseltine (11, 12). A
0.2-to 0.4-pg sample ofviral RNA was digested with
RNaseT, (Calbiochem)and bacterialalkaline
phos-phatase(RNasefree; WorthingtonBiochemicalCo.).
The digested RNA was subsequently labeled with
[y-32P]ATP
and polynucleotide kinase (P.L.Bio-chemicals). They-32P-labeled oligonucleotides were
electrophoresedin a first-dimension 10%
polyacryl-amidegelcontaining6M ureaand25mM citric acid
(pH3.5), followed by electrophoresis in a
second-dimension 22.5%polyacrylamide gel containing 50
mM Tris,50mM boricacid, and1 mM EDTA (pH
8.3). Thegelwasthenautoradiographed.(A) FeLVA
(Glasgow-i);(B)FeLVB(Snyder-Theilen); (C)FeLV
C(Sarma).
VOL. 35,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.504.39.229.67.507.2]544 NOTES
0
0
0
00
2nd
1iSt
0 0
3S
0 0
0 0
0 2nd
A
0ist
B
,.:,*
,*S
0
0
o00
o
o
e
o
o
0
.
o0
lb@0
0 0*
0
c0
0cP@
c
,
0
0
0
S 0
*
ceD
FIG. 2. Comparison ofthegenomes ofFeLV.Equalamountsoflabeled RNaseT,-resistant oligonucleotides
fromtwodifferentstrainsweremixedtogetherimmediately before electrophoresisin thefirstdimension.
Two-dimensionalgelelectrophoresiswasperformedaspreviouslydescribed. (A)Schematicrepresentationofthe
mixtureofFeLV A (Glasgow-i)and FeLV B (Snyder-Theilen). RNaseT1-resistantoligonucleotides: (open
circles) found onlyinFeLVA; (shadedcircles), found onlyinFeLVB; (black circles),commonto both FeLV
A and FeLVB. (B)Schematicrepresentation ofthe mixtureofFeLV A (Glasgow-i)andFeLV C(Sarma).
RNase T1-resistant oligonucleotides: (open circles) found onlyin FeLVC; (shadedcircles), found only in
FeLVA;(black circles),commontoboth FeLVA and FeLVC.(C)Schematicrepresentationofthemixtureof
FeLV B (Snyder-Theilen) and FeLV C(Sarma). RNase T1-resistantoligonucleotides: (open circles) found onlyin FeL VC; (shaded circles)found onlyinFeLVB; (blackcircles)commontobothFeLV B and FeLV C.
2nd
L1st
c
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.504.62.456.58.553.2]TABLE 1. Calculatedsequence identities of the FeLV's
No. (%) showingsequence identity with viral genome"
Viralgenome FeLV A
(Glasgow-i) FeLV B FeLV C FeLV A (Rickard)
FeLVA(Glasgow-1) 59/59(100) 32/61 (52) 37/58(64) 46/60(76)
FeLV B(Snyder-Theilen) 32/53(60) 55/55 (100) 21/56(37) ND
FeLVC(Sarma) 37/56(66) 21/56 (37) 56/56(100) ND
FeLV A(Rickard) 46/59(78) ND ND 60/60(100)
aNumberof RNase T,-resistant oligonucleotides found in common between the two viral genomes/total
number ofuniqueoligonucleotides.The number of unique oligonucleotides scored for a particular viral strain
variedslightlyfromexperiment toexperimentasaresult of the degree of resolution of each fingerprint. The
numbers inparenthesesarethe percentages of sequenceidentity. ND, Not done.
* 0
P.:s
oll
41
0 C* 0 .
0
0- Wc1
S-0.1
position butnotnecessarily thesamesequence. On the other hand, this calculation maybe an underestimate, since the mobility of an oligo-nucleotidecanbealteredbyachange inasingle nucleotide.
Similar analysiswasdone onmixtures of 32P-labeledoligonucleotides ofFeLV A
(Glasgow-i)
and FeLV C
(Sarma)
(Fig. 2B) andofFeLV B (Snyder-Theilen) and FeLV C (Sarma) (Fig. 2C). The number of comigrating oligonucleo-tideswasdeterminedasdescribed above and is listed in Table 1 for eachexperiment. Weesti-matethat64 to66%of thesequencesofFeLV A
(Glasgow-i)
and FeLV C(Sarma)
areidentical.ThegenomesofFeLV B (Snyder-Theilen) and
FeLV C (Sarma) are more distantly related. Only about one-third (37%) of the unique oligo-nucleotides have the same electrophoretic
mo-bility.
To determine whether the fingerprint of an FeLV strainwasaffected by thespecies of fibro-blastsonwhich the viruswasgrown,the finger-print of FeLV B
(Snyder-Theilen)
grown on feline fibroblastswascompared with that of the samevirusgrown oncanine fibroblasts. No dif-ferences in thefingerprints
of the two prepara-tions were detected in this experiment (not shown).Thepreceding experiments demonstrate that viruses of thedifferent
subgroups
canbedistin-guished
from one another byanalysis
of the unique large RNaseT1-resistant
oligonucleo-tides. Todetermine whether viruses of thesame subgroup could also be
distinguished
from each otherby thismethod,thefingerprint
of the viral RNA ofFeLV A(Glasgow-1)wascompared
with thatofanothersubgroupAvirus,
FeLVA(Rick-ard) (Fig. 3). We estimate that the degree of
sequence identity between thesetwo strains is
0
0
0
C
FIG. 3. Comparison ofthe two subgroupA virus
genomes.FeLVA (Rickard)wasisolatedfromF422 cells, a spontaneous lymphoidcelllineprovided by M. Essex(13). (A)FeLVA (Glasgow-1); (B)FeLVA (Rickard); (C)Schematicdiagram. RNase
T,-resist-ant oligonucleotides; (open circles) found only in FeLV A (Rickard); (shaded circles) found only in FeLVA(Glasgow-1);(black circles)commontoboth. B
NOTES 545
VOL. 35,1980
0
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.504.52.245.200.664.2]546 NOTES
76 to 78% (Table 1). It is possible that the
background spots on theautoradiogram of the
fingerprint of FeLV A (Rickard) represent
se-quencesfromasecondviral orsubviral
compo-nent.
The extent of sequence identity detected
among the FeLV strains was anticipated from
previousstudiesofnucleicacidhomology. These
studiesindicated an overallhomology ofabout
85% among all FeLVstrainstested. This
homol-ogy is consistent with the degree of sequence
identityrepeatedhere, with thepossible
excep-tion of therelatively lowdegreeofidentity,37%,
found between FeLV B (Snyder-Theilen) and
FeLVC (Sarma).
Theresults presented here demonstrate that
afingerprint of the 70SgenomicRNAprovides
auseful method foridentifyinganFeLV strain.
No two FeLV strains tested had the same
fin-gerprint,includingtwoindependentsubgroup A isolates.Thefingerprints presentedhereshould provide a standard for future FeLV
identifica-tion. The subgroup type of each of the isolates
used here wasconfirmed in blindtests of
sub-groupspecificity by OswaldJarrett.
Asanexampleof theutilityof this method for virological studies of FeLV, we have used the RNAfingerprinting techniquetocorrectly
iden-tifythe FeLVhelpercomponentof a mixture of
FeLV and defective feline sarcoma virus
ge-nomesasbelonging to subgroup B. The helper
virus had been previouslythought to belong to
subgroup C. We have alsofingerprinted avirus describedassubgroup B and detectedasecond subgroup A viral component in the analysis.
Thisviruswassubsequentlytyped as two viruses
with both subgroup A and subgroup B
specific-ities.(Apreliminaryreportof thisworkhasbeen
published[6]).
We thankMyronEssex,Oswald Jarrett,and William Hardy for helpful discussions and Robert Crowther for technical assistance.
W.H. isthe recipientof an American Cancer Society Fac-ulty Research Award. F.S.P. is a Fellow of the Leukemia Society of America, Inc. and the Danish Natural Science ResearchCouncil. Z.F.R. issupportedby Public Health Ser-vicegrant AID 0014 from theNational InstitutesofHealth.
LITERATURE CITED
1. Cotter, S.M.,W. D.Hardy,Jr.,and M. Essex. 1975. Theassociationof the felineleukemiavirus with lym-phosarcoma and other disorders. J. Am. Vet. Med. Assoc.6:447-455.
2.Essex,M. 1975. Horizontallyandvertically transmitted oncornaviruses ofcats.Adv. Cancer Res.21:175-248. 3. Essex,M.,S. M.Cotter,A. H. Sliski,W. D.Hardy,
Jr.,J. R.Stephenson, S.Aaronson, and0.Jarrett. 1977.Horizontal transmission of feline leukemia virus under natural conditions in afeline leukemia cluster household.Int. J.Cancer19:90-96.
4.Hardy, W.D.,Jr.,P.W.Hess,E.G.MacEwen, A. K. McClelland,E. E.Zuckerman,M.Essex, S.M. Cot-ter,and0.Jarrett. 1976.Biologyoffeline leukemia virus in thenatural environment. CancerRes. 36:582-588.
5. Hardy, W.D., Jr., L.J.Old,P. W.Hess, M. Essex, and S. M. Cotter. 1973. Horizontal transmission of felineleukemiavirus. Nature(London)244:266-269. 6. Haseltine,W.A.,F.S.Pederson,B.G.Sahagan,Z. F.
Rosenberg, andJ.Koslov. 1979.Comparative anal-ysisof RNA tumor virusgenomes,p. 529-552. In R. Neth and R. Gallo (ed.), Modem trends in human leukemia,vol.III.Springer-Verlag,Berlin.
7. Jarrett, O., W. D.Hardy, Jr.,M.C.Golder,and D. Hay. 1978.Thefrequency of occurrenceof feline leu-kemia virussubgroupsin cats. Int.J. Cancer 21:334-337.
8. Jarrett,O., H. M.Laird,andD.Hay. 1973. Determi-nants of the host range offeline leukemiaviruses. J. Gen.Virol.20:169-175.
9. Jarrett,O., andP. H.Russell.1978.Differentialgrowth and transmissionin catsof felineleukemia viruses of subgroupsAandB. Int.J.Cancer21:466-472. 10. Levin, R., S.K. Ruscetti, W. P. Parks, and E. M.
Scolnick. 1976. Expression of feline type-C virusin normalandtumortissues of the domesticcat.Int. J. Cancer18:661-671.
11. Pedersen, F.S.,andW.A.Haseltine. 1980.A micro-method fordetailed characterization ofhigh molecular weight RNA.MethodsEnzymol. 65:680-687. 12. Pedersen,F.S.,andW.A. Haseltine.1980.Analysisof
the genomeofanendogenous, ecotropic retrovirusof theAKR strainofmice:micromethodfordetailed char-acterization of high-molecular-weight RNA. J. Virol. 33:349-365.
13. Rickard, C. G., J. E. Post, F. Noronha, and L. M. Barr. 1969. Atransmissible virus-inducedlymphocytic leukemia ofthe cat. J.Nat.CancerInst.42:987-1014. 14. Sarma, P. S., and T.Log. 1971. Viral interference in
feline leukemia-sarcomacomplex.Virology44:352-358. 15. Sarma,P., andT.Log.1973.Subgroupclassificationof feline leukemiaand sarcoma virusesby viral interfer-enceand neutralizationtests.Virology54:160-169. 16. Sarma,P. S.,T.Log,D.Jain, P. R.Hill, and R. J.
Huebner. 1975. Differential host range of viruses of feline leukemia-sarcomacomplex. Virology64:438-446.