Copyright © 1998, American Society for Microbiology. All Rights Reserved.
A Bipartite Membrane-Binding Signal in the Human
Immunodeficiency Virus Type 1 Matrix Protein Is Required
for the Proteolytic Processing of Gag Precursors
in a Cell Type-Dependent Manner
YOUNG-MIN LEE, CHUN-JUAN TIAN,ANDXIAO-FANG YU*
Department of Molecular Microbiology and Immunology, Johns Hopkins University School of Hygiene and Public Health, Baltimore, Maryland 21205
Received 11 June 1998/Accepted 13 July 1998
It is unclear whether proteolytic processing of the human immunodeficiency virus type 1 (HIV-1) Gag protein is dependent on virus assembly at the plasma membrane. Mutations that prevent myristylation of HIV-1 Gag proteins have been shown to block virus assembly and release from the plasma membrane of COS cells but do not prevent processing of Gag proteins. In contrast, in HeLa cells similar mutations abolished processing of Gag proteins as well as virus production. We have now addressed this issue with CD41T cells, which are
na-tural target cells of HIV-1. In these cells, myristylation of Gag proteins was required for proteolytic processing of Gag proteins and production of extracellular viral particles. This result was not due to a lack of expression of the viral protease in the form of a Gag-Pol precursor or a lack of interaction between unmyristylated Gag and Gag-Pol precursors. The processing defect of unmyristylated Gag was partially rescued ex vivo by coex-pression with wild-type myristylated Gag proteins in HeLa cells. The cell type-dependent processing of HIV-1 Gag precursors was also observed when another part of the plasma membrane binding signal, a polybasic region in the matrix protein, was mutated. The processing of unmyristylated Gag precursors was inhibited in COS cells by HIV-1 protease inhibitors. Altogether, our findings demonstrate that the processing of HIV-1 Gag precursors in CD41T cells occurs normally at the plasma membrane during viral morphogenesis. The
intra-cellular environment of COS cells presumably allows activation of the viral protease and proteolytic processing of HIV-1 Gag proteins in the absence of plasma membrane binding.
The infectious virions of human immunodeficiency virus type 1 (HIV-1), like those of other retroviruses, are produced by the envelopment of the retroviral capsid core. This core structure, consisting of the gag and pol gene products, is sur-rounded by the lipid membrane of the host cell, which contains viral glycoproteins encoded by the env gene (7, 26). The gag gene of HIV-1 encodes the major structural proteins of the viral core, which is initially synthesized as a 55-kDa polyprotein precursor (Pr55Gag) and subsequently cleaved to yield the
ma-trix (MAp17), capsid (CAp24), nucleocapsid (NCp7), and p6 proteins by the viral protease encoded by the pol gene region (6). The pol gene encodes three enzymatic components, includ-ing the protease (PRp11), reverse transcriptase (RTp66/51), and integrase (INp34). The HIV-1 Pol protein is synthesized as a precursor 160-kDa Gag-Pol fusion polyprotein (Pr160Gag-Pol)
by a 21 ribosomal frameshifting mechanism (8, 28). The Pr160Gag-Polprecursor is eventually processed into mature
pro-teins.
The newly synthesized Gag and Gag-Pol precursors are transported from the cytoplasm to the inner face of the plasma membrane, where viral budding takes place (7, 13, 26). In type C retroviruses, including HIV-1, assembly of the Gag and Gag-Pol precursors into capsids is thought to occur at the plasma membrane simultaneously with virus budding. In type B/D retroviruses, however, immature capsids are first formed
within the cytoplasm and then transported to the plasma mem-brane for budding. In both type C and type B/D retroviruses, the matrix domain of the Gag protein has been shown to contain a targeting signal for intracellular transport of Gag and Gag-Pol precursors to the plasma membrane (7, 13, 26).
In most retroviruses, myristic acid is cotranslationally and covalently attached to the N-terminal glycine residue of the Gag protein (21). This myristic acid modification of Gag pro-teins has been shown to be essential for their intracellular transport to the plasma membrane (7, 13, 26). In type C retro-viruses, mutations blocking this modification lead to a failure of extracellular viral particle production (1, 3–5, 16, 17, 19, 22, 25, 30). In type B/D retroviruses, immature capsids preas-semble within the cytoplasm in the absence of myristylation, but the capsids are not transported to the plasma membrane and are instead accumulated in the cytoplasm (20).
It is well established that activation of the viral protease is required for the formation of infectious retroviral particles (24). However, when and how viral protease is activated during viral morphogenesis remains largely unknown. In general, mu-tations that prevent transport to or stable association of Gag and Gag-Pol precursors with the plasma membrane block pro-tease activation (24). This observation has been well supported by experiments in which myristic acid modification of the Gag protein has been prevented in type C retroviruses (1, 19, 25) or in type D retroviruses (20). In retroviruses such as Rous sar-coma virus, in which the Gag molecule is not modified by myristic acid, mutations in the matrix protein that block plasma membrane targeting also dramatically block protease activa-tion (27).
It has been suggested that HIV-1 may be an important
* Corresponding author. Mailing address: Department of Molecular Microbiology and Immunology, Johns Hopkins University School of Hygiene and Public Health, Room E4012, 615 N. Wolfe St., Baltimore, MD 21205. Phone: (410) 955-3768. Fax: (410) 614-8263. E-mail: xfyu @jhsph.edu.
9061
on November 9, 2019 by guest
http://jvi.asm.org/
exception to this rule (5, 10, 24). In HIV-1, proteolytically processed mature Gag proteins can be detected in the cytosolic fraction of infected cells, suggesting activation of the viral protease before targeting to the plasma membrane (10). Fur-thermore, unmyristylated Pr55Gagprecursors in HIV-1, unlike
those in other retroviruses, are proteolytically processed into mature viral proteins in transfected COS cells (5, 17, 30) de-spite the presence of defects in intracellular transport (30) and virus assembly at the plasma membrane (5, 17, 30). However, proteolytic processing of unmyristylated HIV-1 Gag proteins has not been observed in transfected HeLa cells (1). Since nei-ther COS cells nor HeLa cells are natural target cells for HIV-1, it remains to be determined whether myristylation of HIV-1 Gag proteins is required for proteolytic processing of Gag precursors in clinically relevant CD41T cells.
In this study, we have investigated whether the plasma mem-brane binding signal in the matrix domain of the HIV-1 Gag protein is required for proteolytic processing of the Gag pre-cursors. We found that mutation of the membrane binding signal resulted in a defect in the proteolytic processing of the Gag precursors in HeLa cells but not in COS cells. Further-more, the Gag processing defect in the membrane binding mutants was also observed in CD41 T cells. Neither lack of
expression of the Pr160Gag-Polprecursor containing the HIV-1
viral protease nor of Pr55Gag-Pr160Gag-Pol precursor
interac-tion could explain the processing defect in the unmyristylated Gag precursor. Proteolytic processing of the unmyristylated Gag precursors in COS cells could be inhibited by HIV-1 protease inhibitors, indicating that HIV-1 protease may be activated prior to plasma membrane association in COS cells. Our findings suggest that processing of HIV-1 Gag precursors occurs normally at the plasma membrane during viral morpho-genesis, by a pathway similar to that described for other ret-roviruses.
MATERIALS AND METHODS
Construction of mutants.The wild-type infectious proviral plasmid HXB2 Hygro (Myr1) was constructed by replacing the neomycin resistance gene in the infectious proviral plasmid HXB2Neo (14) with the hygromycin resistance gene. The hygromycin resistance gene was amplified from pCEP4 (Invitrogen) by PCR with two primers containing ClaI and XhoI sites at the 59and 39ends, respective-ly. The amplified and ClaI-XhoI-digested fragments were cloned into the infectious HXB2Neo proviral plasmid, which was digested with the same enzymes. The isogenic myristylation-negative mutant proviral plasmid (Myr2) was constructed by swapping the ApaI-ApaI fragment from the MGA plasmid (30) into HXB2 Hygro.
The Pr160 plasmid expresses only the Pr160Gag-Polfusion polyprotein in the
absence of Pr55Gagprecursor expression and Pr160Gag-Polprecursor processing.
To construct this plasmid, the PstI-SalI fragment from pGPpr2(18) containing the HIV-1 viral protease active-site substitution (aspartic acid to alanine) was subcloned into the pGEM3Z vector. The subcloned pGEM3Z vector was used to change the wild-type nucleotide sequence 59-AAT TTT TTA GGG-39to 59-AAC TTC TTA AGG G-39at the21 frameshifting site. The insertion of one adeno-sine induces 100% fusion of the pol gene with the upstream gag gene, and the two thymidine substitutions were introduced to achieve expression of the Pr160Gag-Pol
fusion polyprotein without any frameshifting into the21 open reading frame. None of these mutations changed the amino acid sequence of the Gag or Gag-Pol proteins.
Cells and transfection.COS-7 cells (an African green monkey kidney cell line) were obtained from the American Type Culture Collection (ATCC). HeLa CD41b-galactosidase (b-Gal) cells (a derivative of a human cervical carcinoma cell line) were obtained from the AIDS Research and Reference Reagent Program, National Institutes of Health, Bethesda, Md. HeLa CD41b-Gal cells were used instead of the original HeLa cells in this study because HeLa CD41 b-Gal cells have a high transfection efficiency that results in a level of viral protein expression comparable to that of COS-7 cells. Hereafter, HeLa CD41 b-Gal cells are referred to as HeLa cells. Both COS-7 and HeLa cells were maintained in Dulbecco’s modified Eagle’s medium with 10% fetal bovine serum and antibiotics. The CD41T-lymphoid cell line SupT-1 was also obtained from the American Type Culture Collection and was maintained in RPMI 1640 me-dium supplemented with 10% fetal bovine serum and antibiotics.
Transfection of COS-7 cells was performed as previously described (14). For HIV-1 protease inhibitor experiments, transfected COS-7 cells were maintained
in Dulbecco’s modified Eagle’s medium with 10% fetal bovine serum and 20mM saquinavir until cell lysis and protein analysis. HeLa cells were also transfected by the DEAE-dextran method, as previously described (14) except that 200mg of DEAE-dextran per ml was used for transfection.
To generate the Myr1/SupT-1, Myr2/SupT-1, and Pr160/SupT-1 cell lines, uninfected SupT-1 CD41T cells were transfected with Myr1, Myr2, and Pr160 proviral plasmids, respectively, using Lipofectin as suggested by the manufac-turer (Gibco BRL). At 48 h after transfection, cells were selected with 0.8 mg of hygromycin B per ml (for Myr1/SupT-1 and Myr2/SupT-1) or 1.2 mg of G418 per ml (for Pr160/SupT-1) for 2 to 3 weeks until stable CD41T cells were generated. Myr1/H9 and Myr2/H9 cell lines were also generated, as described above for Myr1/SupT-1 and Myr2/SupT-1 cells. The established CD41T-cell lines were batch selected and maintained in RPMI 1640 with 10% fetal bovine serum, antibiotics, and 0.2 mg of hygromycin B or G418 per ml.
Protein analysis and sera.Immunoblotting of cell lysates or extracellular viral particles was performed as previously described (14). An HIV-1-positive human serum was obtained from an HIV-1-infected patient from Baltimore, Md. Goat anti-HIV-1 p6 antiserum has been previously described (29). Rabbit anti-CAp24 antiserum was obtained from the AIDS Research and Reference Reagent Program, National Institutes of Health. Mouse monoclonal RTp66/51 anti-body was purchased from BTI, Columbia, Md. Alkaline phosphatase (AP)-conjugated goat anti-rabbit immunoglobulin G (IgG) and AP-(AP)-conjugated rabbit anti-goat IgG antibodies were purchased from Sigma ImmunoResearch, and AP-conjugated goat anti-mouse IgG antibody was purchased from Jackson Im-munoResearch Laboratories, Inc.
Coimmunoprecipitation.The SupT-1-derived T-cell lines were lysed by incu-bation at room temperature for 5 min in phosphate-buffered saline (PBS) con-taining 1% Triton X-100, and the cell lysates were clarified by centrifugation at 14,000 rpm for 20 min. The precleared cell lysates were incubated overnight at 4°C with protein A-Sepharose beads that had been preincubated with polyclonal anti-HIV-1 p6 antiserum and then washed with PBS containing 1% Triton X-100. After incubation, the immunoprecipitated materials were obtained by centrifugation and six washes with PBS containing 1% Triton X-100. To release the immunoprecipitates, the Sepharose beads were boiled in sample loading buffer (0.08 M Tris-HCl [pH 6.8], 2.0% sodium dodecyl sulfate [SDS], 10% glycerol, 0.1 M dithiothreitol, 0.2% bromophenol blue) for 5 min and then briefly centrifuged at 12,0003g. The immunoprecipitated proteins were separated by SDS-polyacrylamide gel electrophoresis (PAGE), and viral proteins were visu-alized by immunoblotting.
RESULTS
Myristic acid modification of the HIV-1 Gag proteins is es-sential for proteolytic processing of Gag precursors in a cell type-dependent manner.It appears that myristic acid modifi-cation of HIV-1 Gag protein is not required for proteolytic processing of Pr55Gagprecursors in the African green monkey
kidney cell line COS (5, 17, 30). On the other hand, myristyl-ation is required for processing of Pr55Gagprecursors in the
human cervical carcinoma cell line HeLa (1). The discrepancy between these previous studies has not yet been resolved and could be attributed to the use of different plasmid constructs, transfection methods, levels of protein expression, or cell types to express the HIV-1 viral proteins. We examined all of these possibilities by transfecting these two cell types with the same plasmid construct by the same transfection method. The ex-pressions of the viral proteins after transfection were compa-rable in the COS and HeLa cells.
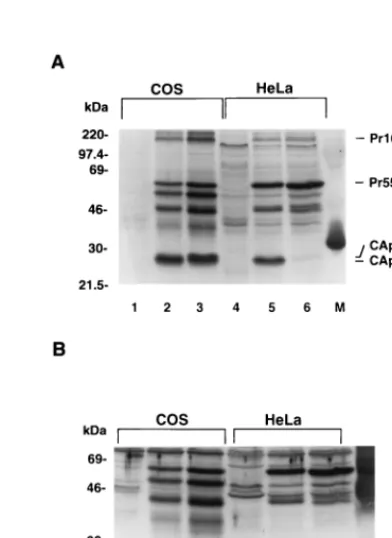
In COS cells transfected with mutant Myr2proviruses, im-munoblotting with an HIV-1-positive human serum clearly showed the presence of the mature CAp24 and cleavage-inter-mediate p25 proteins, containing CAp24 and the p2 spacer peptide (Fig. 1A). The mature CAp24 and cleavage-interme-diate p25 proteins were confirmed by immunoblotting with a polyclonal anti-CAp24 antiserum (Fig. 1B). As expected, the Pr55Gag precursors in COS cells transfected with wild-type
Myr1 proviruses were also processed into mature viral pro-teins (Fig. 1A and B). It is noteworthy that the p25 cleavage intermediate was relatively more abundant in the Myr2 trans-fected COS cells than in the Myr1transfected COS cells
(com-pare lanes 2 and 3 in Fig. 1A and B).
In contrast to the results with COS cells, proteolytic process-ing of the Pr55Gagprecursors was not detected in Myr2
trans-fected HeLa cells (Fig. 1C, lane 2), although the Pr55Gag
on November 9, 2019 by guest
http://jvi.asm.org/
cursors were processed into mature viral proteins in Myr1
transfected HeLa cells (Fig. 1C, lane 3). It is unlikely that the Pr55Gag precursors in the Myr2 transfected HeLa cells are
being processed and the cleavage products are simply being de-graded very rapidly, because there was more uncleaved Pr55Gag
in Myr2transfected HeLa cells (Fig. 1C, lane 2) than in Myr1
transfected HeLa cells (Fig. 1C, lane 3).
The difference in HIV-1 Gag processing between COS and HeLa cells was not simply due to the concentration of Gag precursor molecules. When cell lysates from equal numbers of transfected COS and HeLa cells were used, the amount of Gag protein detected in the HeLa cells was not lower than in the COS cells (compare Fig. 1A and C). Comparable expression of Gag proteins in transfected COS and HeLa cells was further demonstrated in other experiments (see Fig. 2). Therefore, the differences in proteolytic processing of unmyristylated Pr55Gag
precursors that we observed in the two cell lines probably re-flect differences in cell type rather than in other experimental conditions.
A polybasic domain at the N terminus of the HIV-1 Gag pro-teins is also required for proteolytic processing of Gag precur-sors in a cell type-dependent manner.It is not clear whether myristylation of HIV-1 Gag per se or plasma membrane bind-ing of Gag and Gag-Pol molecules is required for Gag pro-cessing in HeLa cells but not in COS cells. In addition to the importance of the myristic acid modification, a polybasic do-main at the N terminus of HIV-1 Gag protein has also been shown to play an important role in membrane binding (30, 31).
To test the role of the polybasic domain in proteolytic process-ing of HIV-1 Gag proteins, we took advantage of a mutant, B5, in which substitutions have been made in five basic residues in the polybasic domain at the N terminus of the HIV-1 Gag protein (30). The B5 mutant has been shown to have a defect in intracellular transport of the Gag precursors and production of extracellular viral particles (30).
In this experiment, COS cells or HeLa cells were mock transfected or transfected with either the wild-type or the mu-tant B5 plasmid. At 3 days after transfection, the cells were lysed and analyzed by SDS-PAGE and immunoblotting. Immu-noblotting with an HIV-1-positive human serum (Fig. 2A) and the polyclonal anti-CAp24 antiserum (Fig. 2B) demonstrated that the mutant Pr55Gagprecursors from COS cells transfected
with the B5 plasmid were processed into the mature CAp24 and p25 viral proteins, as seen in COS cells transfected with the Myr2mutant (Fig. 1). As expected, the wild-type Pr55Gag
pre-cursors from COS cells transfected with wild-type plasmid were also efficiently processed into mature CAp24 proteins.
In contrast to the results with COS cells, proteolytic process-ing of mutant Pr55Gagprecursors was dramatically inhibited in
HeLa cells transfected with the mutant B5 plasmid (Fig. 2, lane 6). On the other hand, the Pr55Gagprecursors were efficiently
processed into mature viral proteins in HeLa cells transfected with the wild-type plasmid (Fig. 2, lane 5). Therefore, these results demonstrated that, like the myristylation of HIV-1 Gag proteins, the polybasic domain of HIV-1 Gag proteins plays a critical role in productive proteolytic processing of Pr55Gag
[image:3.612.52.290.68.361.2]precursors in a cell type-dependent manner. FIG. 1. Cell type-dependent proteolytic processing of unmyristylated HIV-1
Gag proteins. COS cells (A and B) or HeLa cells (C) were mock transfected (lane 1) or transfected with the myristylation-negative mutant Myr2(lanes 2) or the wild-type Myr1(lanes 3) proviral plasmid. At 72 h after transfection, cells were lysed in radioimmunoprecipitation assay lysis buffer and separated by SDS– 12% PAGE and then transferred simultaneously to two nitrocellulose filters. Viral proteins were visualized by immunoblotting with an HIV-1-positive human serum (A and C) or a polyclonal anti-CAp24 antiserum (B).
FIG. 2. A polybasic domain at the N termini of HIV-1 Gag proteins is required for processing of Gag precursors in a cell type-dependent manner. COS (lanes 1 to 3) or HeLa (lanes 4 to 6) cells were mock transfected (lanes 1 and 4) or transfected with wild-type plasmid HXB2R3 (lanes 2 and 5) or mutant plas-mid B5 (lanes 3 and 6). At 72 h posttransfection, cell lysates were separated by SDS-PAGE and viral proteins were visualized by immunoblot analysis with an HIV-1-positive human serum (A) or a polyclonal anti-CAp24 antiserum (B). M, molecular weight standards.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.327.523.392.661.2]Myristic acid modification of the HIV-1 Gag proteins is es-sential for proteolytic processing of Gag proteins in CD41T
cells. To examine the role of myristic acid modification of the N terminus of the Gag protein in processing of Pr55Gag
precursors in CD41T cells, we generated SupT-1 CD41
T-cell lines expressing unmyristylated Pr55Gagand Pr160Gag-Pol
proteins (Myr2/SupT-1) and wild-type myristylated Pr55Gag
and Pr160Gag-Polproteins (Myr1/SupT-1).
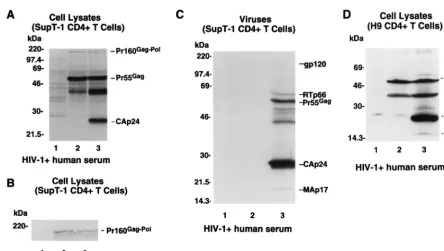
Immunoblotting with HIV-1-positive human serum demon-strated that the Pr55Gagprecursors in the Myr2/SupT-1 cells
were expressed at levels comparable to those in the Myr1/ SupT-1 cells (Fig. 3A, lanes 2 and 3). The myristylated Pr55Gag
precursors in the Myr1/SupT-1 cells were efficiently processed into mature viral proteins such as CAp24 (Fig. 3A, lane 3). In the mutant Myr2/SupT-1 cells, however, processing of unmy-ristylated Pr55Gagproteins into CAp24 was not detected (Fig.
3A, lane 2).
Immunoblot analysis with HIV-1-positive human serum (Fig. 3A) and a monoclonal RTp66/51 antibody (Fig. 3B) demon-strated that expression of Pr160Gag-Pol precursors in the
mu-tant Myr2/SupT-1 cells (Fig. 3B, lane 2) was comparable to that in the wild-type Myr1/SupT-1 cells (Fig. 3B, lane 3). Therefore, the defect in the processing of Pr55Gagprecursors could not be
explained by a lack of Pr160Gag-Polprecursor expression.
Immunoblotting with HIV-1-positive human serum also demonstrated that in SupT-1 CD41T cells, the myristic acid
modification of HIV-1 Gag proteins is essential for production of extracellular viral particles (Fig. 3C). This finding is consis-tent with results of previous studies conducted with COS and HeLa cells (1, 5, 17, 30).
In addition to our findings with SupT-1 cells, the processing defect in the HIV-1 Gag precursors in the absence of myristic
acid modification was also observed in H9 CD41T cells.
Im-munoblotting with HIV-1-positive human serum demonstrated that the Pr55Gagprecursors in the Myr2/H9 cells were
ex-pressed at levels comparable to those in the Myr1/H9 cells (Fig. 3D, lanes 2 and 3). The myristylated Pr55Gagprecursors
in the Myr1/H9 cells were efficiently processed into mature viral proteins such as CAp24 (Fig. 3D, lane 3). In the mutant Myr2/H9 cells, however, processing of unmyristylated Pr55Gag
proteins into CAp24 was not detected (Fig. 3D, lane 2). Stable T-cell lines were used for these studies because it was difficult to detect viral Gag proteins in CD41 T cells after
transient transfection. It is unlikely that the results would be substantially different in transiently transfected T cells, since we observed similar Gag processing defects in the myristylation mutants in transiently and stably transfected HeLa cells (data not shown). However, the possibility that unmyristylated Pr55Gagcould be processed in transiently transfected T cells
has not been formally excluded.
Pr55Gagprecursors are associated with Pr160Gag-Pol
precur-sors in myristylation-negative mutant CD41T cells.It is
pos-sible that the lack of Pr55Gag processing in mutant Myr2/
SupT-1 CD41T cells was caused by a lack of interaction
be-tween unmyristylated Pr55Gagand unmyristylated Pr160Gag-Pol
precursors without targeting to the plasma membrane. This possibility was tested by coimmunoprecipitation analysis. A polyclonal anti-p6 antiserum (29) recognizing only the Pr55Gag
precursor was used to coprecipitate the Pr160Gag-Polprecursor.
The p6 domain is present at the C terminus of the Pr55Gag
precursor but not the Pr160Gag-Polprecursor. If an interaction
occurred between the Pr55Gagand Pr160Gag-Polprecursors, the
[image:4.612.77.522.67.318.2]polyclonal anti-p6 antiserum would coimmunoprecipitate the FIG. 3. Defect in proteolytic processing of HIV-1 Gag precursors in Myr2CD41T cells. Myr1and Myr2CD41T-cell lines were generated as described in Materials and Methods. (A and B) Cell lysates from uninfected SupT-1 (lane 1), Myr2/SupT-1 (lane 2), and Myr1/SupT-1 (lane 3) cell lines were prepared by lysing of cells in radioimmunoprecipitation assay (RIPA) lysis buffer. The cell lysates were separated by SDS–12% PAGE and transferred to two nitrocellulose filters and then immunoblotted with an HIV-1-positive human serum (A) or with a monoclonal anti-RTp66/p51 antibody (B). (C) The supernatants from uninfected SupT-1 (lane 1), Myr2/SupT-1 (lane 2), and Myr1/SupT-1 (lane 3) cell lines were harvested after 48 h of incubation with fresh complete medium. The virion-associated proteins from harvested supernatants were concentrated by ultracentrifugation and analyzed by SDS-PAGE and immunoblotting with an HIV-1-positive human serum. (D) Cell lysates from uninfected H9 (lane 1), Myr2/H9 (lane 2), and Myr1/H9 (lane 3) cell lines were prepared by lysing the cells in RIPA lysis buffer. The cell lysates were separate by SDS–12% PAGE and the immunoblotted with an HIV-1-positive human serum.
on November 9, 2019 by guest
http://jvi.asm.org/
Pr160Gag-Polprecursor in the presence of the Pr55Gag
precur-sor but not in its absence.
As a control for the specificity of this coimmunoprecipita-tion analysis, a SupT-1 CD41T-cell line expressing only the
Pr160Gag-Pol fusion polyprotein precursor (Pr160/SupT-1)
was constructed. Immunoblotting of cell lysates from Myr2/ SupT-1, Myr1/SupT-1, and Pr160/SupT-1 cells with an HIV-1-positive human serum showed that the Pr55Gagprecursors
were detected in the Myr2/SupT-1 and Myr1/SupT-1 cells (Fig. 4A, lanes 2 and 3) but not in the Pr160/SupT-1 cells (Fig. 4A, lane 4). A monoclonal anti-RTp66/51 antibody showed that comparable amounts of Pr160Gag-Polprecursors were
ex-pressed in all three cell lines (Fig. 4B, lanes 2 to 4).
For coimmunoprecipitation experiments, cell lysates from uninfected, Myr2/SupT-1, Myr1/SupT-1, and Pr160/SupT-1 cells were first immunoprecipitated with anti-p6 antiserum. The immunoprecipitated materials were then separated by SDS-PAGE and visualized by immunoblotting. Immunoblot-ting with HIV-1-positive human serum revealed comparable quantities of Pr55Gag precursors in the immunoprecipitated
materials from the mutant Myr2/SupT-1 and wild-type Myr1/ SupT-1 cells but, as expected, not from the Pr160/SupT-1 cells (Fig. 4C). The immunoglobulin heavy chain from the goat anti-p6 antiserum which was used for immunoprecipitation was detected in lanes 1 and 4 in Fig. 4C, presumably because of a weak cross-reactivity of the alkaline phosphatase-conjugated rabbit anti-human IgG with the immunoglobulin heavy chain in the goat anti-p6 antiserum. These bands were not detected when the goat anti-p6 antiserum was not used during immu-noprecipitation (Fig. 4C, lanes 5 and 6).
Upon longer exposure of the same immunoblots, compara-ble quantities of Pr160Gag-Polprecursors were detected in the
lysates from the Myr2/SupT-1 and Myr1/SupT-1 cells (Fig. 4D, lanes 2 and 3) but not in those from the Pr160/SupT-1 cells (Fig. 4D, lane 4). The coimmunoprecipitation of Pr160Gag-Pol
with either Myr1or Myr2Pr55Gagwas further confirmed by
immunoblotting with a monoclonal anti-RTp66/51 antibody (Fig. 4E, lanes 2 and 3). The Pr160Gag-Polprecursors were not
detected by the anti-RTp66/51 antibody when the cell lysate of Pr160/SupT-1 cells was immunoprecipitated with p6 antiserum (Fig. 4E, lane 4), demonstrating that the coimmunoprecipita-tion of Pr160Gag-Pol precursors from the Myr2/SupT-1 and
Myr1/SupT-1 cell lysates could not be explained by cross-reactivity of p6 antiserum to Pr160Gag-Pol. In the absence of p6
antiserum, neither the Pr55Gagnor the Pr160Gag-Polprecursors
of the viral proteins were precipitated by protein A-Sepharose beads (Fig. 4C to E, lanes 5 and 6), indicating that the Pr160Gag-Polprecursors were specifically
coimmunoprecipitat-ed with the Pr55Gagprecursors by p6 antiserum.
In addition to the coimmunoprecipitation analysis presented here, we have recently demonstrated that unmyristylated Gag and Gag-Pol proteins of HIV-1 form an assembly-intermediate complex, which is characterized as a large oligomer that has a density of 1.10 to 1.13 g/ml and is primarily composed of Pr55Gagand Pr160Gag-Polprecursors in infected CD41T cells
(15). All of these findings are consistent with the idea that the interaction between unmyristylated Gag and Gag-Pol precur-sors may occur before their association with the plasma mem-brane.
The processing defect in the Pr55Gagprecursors in the
my-ristylation-negative mutant is rescued ex vivo by coexpression with the wild-type Pr55Gagproteins.Myristic acid modification of HIV-1 Gag proteins may alter the conformation of the Pr55Gagand Pr160Gag-Polprecursors; in the absence of this
modification, it is possible that misfolded Pr55Gagprecursors
cannot be processed into mature viral proteins. However, this
FIG. 4. Coimmunoprecipitation analysis. (A and B) Cell lysates from unin-fected (lanes 1), Myr2/SupT-1 (lanes 2), Myr1/SupT-1 (lanes 3), and Pr160/ SupT-1 (lanes 4) cells were analyzed by SDS-PAGE and immunoblotting with an HIV-1-positive human serum (A) or with a monoclonal anti-RTp66/p51 antibody (B). (C, D, and E) Cell lysates from uninfected (lanes 1), Myr2/SupT-1 (lanes 2 and 5), Myr1/SupT-1 (lanes 3 and 6), and Pr160/SupT-1 (lanes 4) cells were prepared by lysing of cells in PBS containing 1% Triton X-100, followed by immunprecipitation with a polyclonal anti-p6 antiserum (lanes 1 to 4) or without antiserum (lanes 5 and 6) overnight at 4°C. The immunoprecipitated materials were separated by SDS–12% PAGE and transferred to two nitrocellulose filters. Viral proteins on the filters were visualized by immunoblotting with an HIV-1-positive human serum (C), and the same filter was then exposed for a longer period of time (D). The other filter was probed with monoclonal anti-RTp66/p51 antibody (E) to visualize the Pr160Gag-Polprecursors.
on November 9, 2019 by guest
http://jvi.asm.org/
explanation is unlikely, since previous experiments have dem-onstrated that the lack of myristylation does not alter the conformation of Gag molecules (16) and that unmyristylated Pr55Gag precursors can be processed efficiently in vitro by
HIV-1 protease (1). Another possibility is that misfolding of unmyristylated Pr160Gag-Pol precursors generates a defective
protease that cannot be activated. This possibility is also less likely, because it has been suggested that the lack of myristyl-ation does not alter the conformmyristyl-ation of the Gag-Pol mole-cules (18). The other possibility is that the defect in processing of unmyristylated Pr55Gagprecursors might result from a lack
of targeting of the Pr160Gag-Polprecursors to the plasma
mem-brane, a prerequisite for activation of the viral protease. If this explanation is the case, providing a plasma membrane target-ing signal in trans by coexpression with wild-type myristylated Pr55Gagprecursors should activate the viral protease activity.
To distinguish among these possibilities, we examined wheth-er expression of myristylated Pr55Gagprecursors in trans could
induce processing of Pr55Gagprecursors by HIV-1 protease of
the unmyristylated Pr160Gag-Polprecursors. The myristylated
Pr55Gagprecursors were provided by theDPol plasmid, which
contains a complete deletion of the HIV-1 pol gene including the viral protease. Transfection of this construct into HeLa cells resulted in expression of unprocessed Pr55Gagprecursors
alone, as detected by the HIV-1-positive human serum (Fig. 5A, lane 4). Furthermore, CAp24 proteins were not detected in HeLa cells transfected with the mutant Myr2 construct alone (Fig. 5A, lane 3); however, mature CAp24 proteins were detected in the cells cotransfected with the mutant Myr2and
DPol plasmids (Fig. 5A, lane 6). A larger quantity of mature CAp24 proteins was detected in cells transfected with the
wild-type Myr1plasmid (Fig. 5A, lane 2) or cotransfected with the Myr1andDPol plasmids (Fig. 5A, lane 5), as determined by reactivity with the HIV-1-positive human serum. The specific-ity of CAp24 production was verified by reactivspecific-ity with the polyclonal anti-CAp24 antiserum (Fig. 5B). This finding demonstrated that the HIV-1 viral protease of the unmyristyl-ated Pr160Gag-Polprecursors could be activated when a
plas-ma membrane targeting signal was provided by myristylated Pr55Gagprecursors in trans. This finding is consistent with a
previous report that unmyristylated HIV-1 Gag proteins can interact with myristylated Gag proteins and are incorporated into released viral particles (16).
A lesser quantity of CAp24 proteins was detected in cells cotransfected with the Myr2andDPol plasmids than in those cells transfected with the Myr1andDPol plasmids (compare lanes 5 and 6 in Fig. 5), suggesting that the interaction between myristylated Pr55Gagprecursors and unmyristylated Pr55Gagor
Pr160Gag-Polprecursors expressed from two separate mRNAs
may be less efficient than that expressed from a single mRNA. Processing of unmyristylated Pr55Gag precursors in COS
cells was inhibited by HIV-1 protease inhibitors.It is possible that the HIV-1 protease in the myristylation mutant virus is activated in COS cells. Alternatively, unmyristylated Pr55Gag
precursors could be cleaved by a cellular protease in COS cells. To address this issue, COS cells were mock transfected or transfected with the wild-type, the myristylation mutant, or the HIV-1 protease mutant Pr2plasmid (15). After transfection, half of the cells were treated with the HIV-1 protease inhibitor and the other half were used as an untreated control. At 3 days after transfection, the cells were lysed and analyzed by SDS-PAGE and immunoblotting.
Immunoblotting with an HIV-1-positive human serum dem-onstrated that, in the absence of the HIV-1 protease inhibitor, the unmyristylated Pr55Gag precursors from COS cells were
processed into mature CAp24 and p25 viral proteins (Fig. 6, lane 6), as seen in the wild-type transfected COS cells (Fig. 6, lane 8). However, in the presence of the HIV-1 protease in-hibitor, the unmyristylated Pr55Gagprecursors (Fig. 6, lane 2)
as well as the wild-type Pr55Gag(Fig. 6, lane 4) from COS cells
were not processed into mature CAp24 and p25 viral proteins. Also, in the presence or absence of the HIV-1 protease inhib-itor, no processing of Pr55Gagto CAp24 was detected for the
FIG. 5. Ex vivo rescue of the processing defect in unmyristylated Gag pre-cursors by cotransfection with myristylated Pr55Gagproteins. HeLa cells were
[image:6.612.76.263.67.345.2]mock transfected (lanes 1), transfected with the wild-type Myr1(lanes 2), mu-tant Myr2(lanes 3), orDPol (lanes 4) construct, or cotransfected with the Myr1 andDPol (lane 5) or Myr2andDPol (lane 6) constructs. At 72 h after trans-fection cell lysates were analyzed by SDS-PAGE and immunoblotting with an HIV-1-positive human serum (A) or polyclonal anti-CAp24 antiserum (B).
FIG. 6. Proteolytic processing of unmyristylated HIV-1 Gag proteins in COS cells is inhibited by HIV-1 protease inhibitor. COS cells were mock transfected (lanes 1 and 5) or transfected with the myristylation-negative mutant Myr2 (lanes 2 and 6), the HXB2Pr-Neo (lanes 3 and 7), or the wild-type Myr1(lanes 4 and 8) proviral plasmid. Half of the transfected cells were treated with 20mM saquinavir (lanes 1 to 4). At 72 h after transfection, cells were lysed in radioim-munoprecipitation assay lysis buffer and separated by SDS–12% PAGE and then transferred to nitrocellulose filters. Viral proteins were visualized by immuno-blotting with an HIV-1-positive human serum.
on November 9, 2019 by guest
http://jvi.asm.org/
HIV-1 protease mutant Pr2(Fig. 6, lanes 3 and 7). These data suggest that processing of unmyristylated Pr55Gag precursors
in COS cells was due to activation of the viral protease and not the activity of a cellular protease.
DISCUSSION
In this study, we have demonstrated that myristylation of the HIV-1 Gag protein is essential for the proteolytic processing of Pr55Gagprecursors as well as for virus production in CD41T
cells. In HIV-1, the role of myristic acid modification of Gag proteins in the proteolytic processing of Gag precursors has been controversial. Some experiments have demonstrated that myristic acid modification of the HIV-1 Gag protein is re-quired for the proteolytic processing of Gag precursors in HeLa cells (1). Other experiments have shown that processing of unmyristylated HIV-1 Gag protein can occur in COS cells, suggesting that activation of the viral protease occurs prior to plasma membrane association (5, 17, 30). Our data suggest that the normal pathway for HIV-1 viral protease activation in CD41T cells requires plasma membrane targeting and
asso-ciation.
Although the expression of the unmyristylated Pr160Gag-Pol
precursors containing the viral protease and the interaction between the unmyristylated Pr160Gag-Poland the
unmyristylat-ed Pr55Gagprecursors were not affected in CD41T cells and
HeLa cells (data not shown), the processing of unmyristylated Pr55Gagprecursors into mature viral proteins was not observed
in these cells. When the plasma membrane targeting signal was provided by wild-type Gag molecules in trans, the HIV-1 viral protease of the unmyristylated Pr160Gag-Polprecursors was able
to process the Pr55Gagprecursors into mature viral proteins in
HeLa cells.
In addition to myristic acid modification, it has been shown that a polybasic domain at the N terminus of HIV-1 Gag pro-teins plays an important role in the intracellular transport and plasma membrane association of the Pr55Gagand Pr160Gag-Pol
precursors (30, 31). Mutations substituting five basic-charge residues in the polybasic domain dramatically abolished the proteolytic processing of HIV-1 Gag precursors into mature viral proteins in HeLa cells and CD41T cells (data not shown)
but not in COS cells. Since processing of membrane binding mutant Gag precursors was observed in nonhuman COS cells but not in human HeLa or CD41T cells, these findings
dem-onstrate that the two elements of the bipartite plasma mem-brane binding signal at the N terminus of the HIV-1 Gag protein, myristic acid modification and a polybasic domain, are simultaneously required for productive processing of HIV-1 Gag precursors. Altogether, these results reinforce the idea that productive processing of Gag precursors in all retrovi-ruses, including HIV-1 (11), is preceded by the targeting of Gag and Gag-Pol precursors to the plasma membrane (24).
Assembly, budding, and maturation of retroviruses are high-ly dynamic and tighthigh-ly regulated processes (7, 24, 26). Since assembly is achieved by uncleaved Gag and Gag-Pol precur-sors, premature activation of the viral protease may be detri-mental to virus assembly (2, 12). In general, activation of the retroviral protease is dependent on virus assembly and budding at the plasma membrane (24). At present, the mechanism(s) by which retroviral protease activity is regulated is largely un-known, but several mechanisms have been proposed (24).
It is possible that retroviral proteases, including the HIV-1 protease, can be activated by autoprocessing, through a mech-anism similar to that seen for pepsin (9). Dimerization of HIV-1 viral protease is known to be a prerequisite for its function (24). In this regard, dimerization of Pr160Gag-Polprecursors,
which is required for autoprocessing, may not occur until virus assembly and budding are initiated at the plasma membrane. Autoprocessing of HIV-1 Pr160Gag-Polprecursors may also
be dependent upon conformational changes in the molecules that can be influenced by conditions in the cellular environ-ment such as pH, lipid composition, or salt concentration. These conditions may differ between the cytoplasm and the plasma membrane, where virus budding occurs. Conforma-tional changes in Pr160Gag-Polprecursors could also be induced
by protein modifications such as phosphorylation or dephos-phorylation, which are accomplished by cellular enzymes at the plasma membrane.
Another possibility is that the HIV-1 protease is activated from the Pr160Gag-Polprecursors by processing with a cellular
protease, through a process similar to that seen for trypsin and other serine proteases (23). One can imagine that a cellular protease, normally localized only on the inner face of the plasma membrane, could partially cleave Pr160Gag-Pol
precur-sors and liberate the viral protease. This event would subse-quently trigger a cascade processing of viral Pr160Gag-Poland
Pr55Gagprecursors by the released viral protease.
In addition to regulation of HIV-1 viral protease activity by activation, it is also possible that the viral protease is regulated by inhibition. During the late stages of the viral life cycle, the activity of HIV-1 viral protease can be suppressed by a cellular factor(s) until virus assembly and budding take place at the plasma membrane. At the plasma membrane, the inhibitory cellular factors are removed from the budding particles, allow-ing subsequent activation of the viral protease and production of infectious virions.
Taken together, our data and other previously published observations suggest that cellular factors, which normally are restricted to the plasma membrane, are required for activation of the HIV-1 protease in CD41T cells. It remains to be
de-termined what triggers HIV-1 protease activation in COS cells in view of the apparent lack of association of Pr55Gagand
Pr160Gag-Polprecursors with the plasma membrane. Further
study will be required to elucidate the mechanism of HIV-1 protease activation and yield data that may lead to the devel-opment of effective antiviral agents.
ACKNOWLEDGMENTS
We thank Casey Morrow for the pGPpr2construct, Richard Mark-ham and David Schwartz for comments on the manuscript, and Liza Dawson for helpful discussions on the project.
This work was supported by Public Health Service grants AI-35525 and DA-09541 from the National Institutes of Health.
REFERENCES
1. Bryant, M., and L. Ratner. 1990. Myristoylation-dependent replication and assembly of human immunodeficiency virus 1. Proc. Natl. Acad. Sci. USA 87: 523–527.
2. Burstein, H., D. Bizub, and A. M. Skalka. 1991. Assembly and processing of avian retroviral gag polyproteins containing linked protease dimers. J. Virol.
65:6165–6172.
3. Freed, E. O., J. M. Orenstein, A. J. Buckler-White, and M. A. Martin. 1994. Single amino acid changes in the human immunodeficiency virus type 1 matrix protein block virus particle production. J. Virol. 68:5311–5320. 4. Gheysen, D., E. Jacobs, F. de Foresta, C. Thiriart, M. Francotte, D. Thines,
and M. de Wilde.1989. Assembly and release of HIV-1 precursor Pr55 Gag virus-like particles from recombinant baculovirus-infected insect cells. Cell
59:103–112.
5. Gottlinger, H. G., J. G. Sodroski, and W. A. Haseltine. 1989. Role of capsid precursor processing and myristoylation in morphogenesis and infectivity of human immunodeficiency virus 1. Proc. Natl. Acad. Sci. USA 86:5781–5785. 6. Henderson, L. E., T. D. Copeland, R. C. Sowder, A. M. Schultz, and S.
Oroszlan.1988. Analysis of proteins and peptides from sucrose gradient banded HTLV-III, p. 135–147. In D. Bolognesi (ed.), Human retroviruses, cancer and AIDS: approaches to prevention and therapy. Wiley Interscience, New York, N.Y.
on November 9, 2019 by guest
http://jvi.asm.org/
7. Hunter, E. 1994. Macromolecular interactions in the assembly of HIV and other retroviruses. Semin. Virol. 5:71–83.
8. Jacks, T., M. D. Power, F. R. Masiarz, P. A. Luciw, P. J. Barr, and H. E.
Varmus.1988. Characterization of ribosomal frameshifting in HIV-1 gag-pol expression. Nature (London) 331:280–283.
9. James, M. N. G., and A. R. Sielecki. 1986. Molecular structure of an aspartic proteinase zymogen, porcine pepsinogen, at 1.8 A resolution. Nature (Lon-don) 319:33–38.
10. Kaplan, A. H., and R. Swanstrom. 1991. Human immunodeficiency virus type 1 Gag proteins are processed in two cellular compartments. Proc. Natl. Acad. Sci. USA 88:4528–4532.
11. Kaplan, A. H., M. Manchester, and R. Swanstrom. 1994. The activity of the protease of human immunodeficiency virus type 1 is initiated at the mem-brane of infected cells before the release of viral proteins and is required for release to occur with maximum efficiency. J. Virol. 68:6782–6786. 12. Krausslich, H. G. 1991. Human immunodeficiency virus proteinase dimer as
a compartment of the viral polyprotein prevents particle assembly and viral infectivity. Proc. Natl. Acad. Sci. USA 88:3213–3217.
13. Krausslich, H. G., and R. Welker. 1996. Intracellular transport of retroviral capsid components. Curr. Top. Microbiol. Immunol. 214:25–63.
14. Lee, Y.-M., X.-B. Tang, L. M. Cimakasky, J. E. K. Hildreth, and X.-F. Yu. 1997. Mutations in the matrix protein of human immunodeficiency virus type 1 inhibit surface expression and virion incorporation of viral envelope glyco-proteins in CD41T lymphocytes. J. Virol. 71:1443–1452.
15. Lee, Y.-M., and X.-F. Yu. 1998. Identification and characterization of virus assembly intermediate complexes in HIV-1-infected CD41T cells. Virology
243:78–93.
16. Morikawa, Y., S. Hinata, H. Tomoda, T. Goto, M. Nakai, C. Aizawa, H.
Tanaka, and S. Omura.1996. Complete inhibition of human immunodefi-ciency virus Gag myristoylation is necessary for inhibition of particle bud-ding. J. Biol. Chem. 271:2868–2873.
17. Pal, R., M. S. Reitz, Jr., E. Tschachler, R. C. Gallo, M. G. Sarngadharan,
and F. D. Veronese.1990. Myristoylation of Gag proteins of HIV-1 plays an important role in virus assembly. AIDS Res. Hum. Retroviruses. 6:721–730. 18. Park, J., and C. D. Morrow. 1992. The nonmyristylated Pr160Gag-Pol
polypro-tein of human immunodeficiency virus type 1 interacts with Pr55Gagand is
incorporated into viruslike particles. J. Virol. 66:6304–6313.
19. Rein, A., M. R. McClure, N. R. Rice, R. B. Luftig, and A. M. Schultz. 1986.
Myristylation site in Pr65gag is essential for virus particle formation by Moloney murine leukemia virus. Proc. Natl. Acad. Sci. 83:7246–7250. 20. Rhee, S. S., and E. Hunter. 1987. Myristylation is required for intracellular
transport but not for assembly of D-type retrovirus capsids. J. Virol. 61:1045– 1053.
21. Schultz, A. M., L. E. Henderson, and S. Oroszlan. 1988. Fatty acylation of proteins. Annu. Rev. Cell Biol. 4:611–647.
22. Spearman, P., J.-J. Wang, Vander Heyden, N., and L. Ratner. 1994. Identi-fication of human immunodeficiency virus type 1 Gag protein domains es-sential to membrane binding and particle assembly. J. Virol. 68:3232–3242. 23. Stroud, R. M., A. A. Kossiakoff, and J. L. Chambers. 1977. Mechanisms of
zymogen activation. Annu. Rev. Biophys. Bioeng. 6:177–193.
24. Vogt, V. M. 1996. Proteolytic processing and particle maturation. Curr. Top. Microbiol. Immunol. 214:95–131.
25. Weaver, T. A., and A. T. Panganiban. 1990. N myristoylation of the spleen necrosis virus matrix protein is required for correct association of the Gag polyprotein with intracellular membranes and for particle formation. J. Vi-rol. 64:3995–4001.
26. Wills, J. W., and R. C. Craven. 1991. Form, function, use of retroviral Gag proteins. AIDS (Philadelphia) 5:639–654.
27. Wills, J. W., R. C. Craven, R. A. Weldon, Jr., T. D. Nelle, and C. R. Erdie. 1991. Suppression of retroviral MA deletions by the amino-terminal mem-brane-binding domain of p60src. J. Virol. 65:3804–3812.
28. Wilson, W., M. Braddock, S. E. Adams, P. D. Rathjen, S. M. Kingsman, and
A. J. Kingsman.1988. HIV expression strategies: ribosomal frameshifting is directed by a short sequence in both mammalian and yeast systems. Cell 55: 1159–1169.
29. Yu, X.-F., Z. Matsuda, Q.-C. Yu, T.-H. Lee, and M. Essex. 1995. Role of the C terminus Gag protein in human immunodeficiency virus type 1 virion as-sembly and maturation. J. Gen. Virol. 76:3171–3179.
30. Yuan, X., X.-F. Yu, T.-H. Lee, and M. Essex. 1993. Mutations in the N-ter-minal region of human immunodeficiency virus type 1 matrix protein block intracellular transport of the Gag precursor. J. Virol. 67:6387–6394. 31. Zhou, W., L. J. Parent, J. W. Wills, and M. D. Resh. 1994. Identification of
a membrane-binding domain within the amino-terminal region of human immunodeficiency virus type 1 Gag protein which interacts with acidic phos-pholipids. J. Virol. 68:2556–2569.