Copyrightq1997, American Society for Microbiology
Echovirus 1 Replication, Not Only Virus Binding to Its
Receptor, VLA-2, Is Required for the Induction of
Cellular Immediate-Early Genes
PASI HUTTUNEN,1* JYRKI HEINO,2ANDTIMO HYYPIA¨1
MediCity Research Laboratory, Department of Virology1and Department of Medical Biochemistry,2
University of Turku, Tykisto¨katu 6A, FIN-20520 Turku, Finland
Received 9 October 1996/Accepted 14 February 1997
Induction of immediate-early genes c-jun, junB, and c-fos was demonstrated during echovirus 1 infection in a human osteogenic sarcoma (HOS) cell line. Tenfold induction was seen at 10 h postinfection, corresponding approximately to the end of the first replication cycle of the virus. Echovirus 1 uses VLA-2 integrin as its cellular receptor, and ligand binding by integrin is known to trigger signal transduction pathways ultimately activating immediate-early genes. In the present study, however, VLA-2 binding alone was not sufficient to induce their expression; viral replication was needed. This conclusion was based on the observations that no stimulation of the immediate-early genes occurred in the MG-63 cell line where the virus attached only to VLA-2 but was not able to replicate and that induction of these genes was observed when the HOS cells were infected with echovirus type 7, known to use a different cellular receptor. Induction was not seen in the presence of the antiviral compound WIN 54954, which evidently inhibits the uncoating but not receptor binding of echovirus 1, suggesting that viral replication triggers the activation of the immediate-early genes. The induc-tion of these genes may have a role in viral replicainduc-tion and in the pathogenesis of infecinduc-tion.
Echoviruses of the Picornaviridae family are human entero-viruses causing meningitis, encephalitis, exanthemas, and car-ditis as well as mild respiratory and enteric diseases (16). They are the largest group of enteroviruses (28 distinct serotypes) and are genetically closely related to each other (19).
Echovi-rus 1 (EV1) is known to use a2b1integrin (VLA-2; CD49b/
CD29) for attachment to target cells (7, 8). The natural ligands of VLA-2 are several collagens and laminin 1. However, it has been shown that EV1 and extracellular matrix ligands bind VLA-2 by different mechanisms, probably at different func-tional sites (5).
Integrin occupancy and clustering are known to trigger sig-naling pathways, which activate mitogen-activated, extracellu-lar signal-regulated, and c-jun N-terminal protein kinases and ultimately immediate-early genes (36). Immediate-early gene products act as nuclear messengers converting a cytoplasmic signal into changes in gene expression. Their expression is rapidly and transiently induced by other extracellular stimuli such as growth factors, cytokines, tumor promoters, hypoxia, and acidosis but also by certain proteins of oncogenic DNA viruses (3, 4, 9, 13, 34, 38). Immediate-early genes include members of fos and jun families. Their gene products are major components of transcription factor AP-1, which is as-sumed to regulate the transcription of a set of genes crucial for cell growth and differentiation.
It has become evident that the pathogenicity of viruses is highly dependent on their receptor recognition but also on the presence of many host cell factors, which interact with viral macromolecules and influence the effectiveness of virus repli-cation. This study showed that EV1 infection can induce the expression of c-fos, c-jun, and junB genes. Since EV1 is also a
ligand for a2b1 integrin, we were interested to see whether
either binding of the virus to the receptor or subsequent events in the replication cycle are required for early gene induction.
EV1 (Farouk strain), -7 (Wallace), and -11 (Gregory) were obtained from the American Type Culture Collection. They
were propagated in LLC Mk2cells and purified by the method
described by Abraham and Colonno (1). The purified viruses were diluted in phosphate-buffered saline (PBS) containing 0.5
mM MgCl2and stored at2708C until used. The two human
osteogenic sarcoma (HOS) cell lines MG-63 and HOS were also from the American Type Culture Collection. HOS
pa2AW cells, constantly overexpressing a2integrin cDNA in
pAWneo2 vector, were established as described elsewhere (27). Control HOS pAW cells contained only the pAWneo2 expression vector. The cells were maintained in Dulbecco’s modified Eagle’s medium (Life Technologies, Inc.) supple-mented with 10% fetal calf serum (Life Technologies, Inc.)
and 400 mg of G418 (Gibco) per ml. When the cells were
infected with the virus, a culture medium containing Dulbec-co’s modified Eagle’s medium supplemented with 1% fetal calf serum was used. Antiviral compound WIN 54954 was a gen-erous gift from Sanofi Winthrop, Inc.
For binding assays, EV1 and -11 were metabolically labeled
with culture media containing [35S]methionine (Amersham,
Inc.). Cells were suspended in 20 ml of ice-cold PBS and,
incubated for 1 h on ice with the radioactively labeled virus
(10,000 cpm) in the presence or absence of an a2
-integrin-specific antibody (12F1); (26). The cells were washed three times with 1 ml of PBS to remove any unbound virus and finally
suspended in 100ml of distilled water. Radioactivity was
mea-sured in a liquid scintillation counter (Wallac).
The release of EV1 from infected HOS pa2AW cells to the
culture medium and the amount of intracellular virus were determined on confluent monolayers of the same cells in
dilu-tions of 1021to 1028. The cells were examined for cytopathic
effect (CPE) during 2 days, and the results were expressed as
end point titers. Virus RNA transfections (1 mg of purified
genomic RNA) were carried out with the Lipofectin reagent (Sigma) according to the manufacturer’s recommendations.
After transfection, the cells were incubated at 378C for 8 or
* Corresponding author. Phone: (358) 2-333 7001. Fax: (358) 2-333 7000. E-mail: [email protected].
4176
on November 9, 2019 by guest
http://jvi.asm.org/
12 h and then washed with PBS. Total RNA was extracted and used for Northern blot analysis. The susceptibility of cell lines to echovirus infection was studied by immunoperoxidase stain-ing with cross-reactive rabbit anti-EV11 primary antibody and peroxidase-conjugated secondary antibody. No reactivity of the antiserum against the uninfected cells was observed. The ratios of positively stained cells to all cells were counted under a light microscope, and the results were expressed as percentages of infected cells.
For Northern blotting, total cellular RNA was extracted by the thiocyanate-CsCl method (10). Twenty-five micrograms of total cellular RNA was separated in formaldehyde-containing 1% agarose gels, transferred to a nylon membrane (ZETA
probe; Bio-Rad), and hybridized with32P-labeled (Amersham,
Inc.) human c-jun (2), junB (35), c-fos (genomic fragment from Amersham, Inc.), and full-length EV11 cDNA (12) probes. A rat glyceraldehyde-3-phosphate dehydrogenase (GAPDH) probe (15) was used to control the proportional quantity of cellular RNA. All autoradiograms were also analyzed with the Microcomputer Imaging Device, version M4 (Imaging Re-search, Inc.), and the resulting measurements were adjusted to correspond to GAPDH mRNA levels.
EV1 binds to VLA-2 on the surfaces of HOS and MG-63 cells but replicates only in HOS cells. A binding assay using radioactively labeled virus was adapted to study the attachment of EV1 to the cell lines. Both MG-63 and HOS cells were found to bind EV1 (Fig. 1). The attachment was efficiently blocked with monoclonal antibody 12F1 (20, 26), indicating
that EV1 usesa2b1 integrin as its receptor also in these cell
lines. No significant blocking of the binding of EV11, known to use other cell surface molecules for its entry into the cells, was
observed. HOS cells express a low copy number ofa2integrin
subunit on their surfaces (11), and often the expression does
not exceed the detection level (32). Overexpression ofa2
in-tegrin chain on HOS pa2AW cells was found to significantly
increase virus binding (about 40%) (Fig. 1).
The replication of EV1 in HOS and MG-63 cells was inves-tigated with cross-reactive anti-EV11 antibody for
immunoper-oxidase detection. HOS pa2AW cells were prone to EV1
in-fection as determined by detection of intracellular viral protein synthesis, whereas MG-63 cells did not support virus
replica-tion (Table 1). In other words, the virus was able to replicate in HOS cells but not in MG-63 cells, although it also bound to the
latter cells. EV1 infection in HOS pa2AW cells was blocked by
anti-a2integrin antibody (12F1) as could be expected on the
basis of the binding assay. EV7 also infected HOS pa2AW
cells, and the infection was blocked by monoclonal anti-decay-accelerating factor (DAF) antibody (IF7), suggesting that EV7 uses DAF as its receptor (6, 40), also in this cell line.
To determine the time course of the EV1 replication cycle in HOS cells, end-point titers were measured at several time points postinfection (p.i.). A similar CPE as normally seen in
LLC Mk2cells (monkey kidney cells used for virus
propaga-tion) was observed in HOS pa2AW cells, and the replication
cycle of EV1 in this cell line was approximately 8 h, corre-sponding to that seen in other susceptible cell lines (Fig. 2). Thereafter, a second cycle appeared as was suggested by the presence of remaining uninfected cells after initial virus inoc-ulation (Table 1).
EV1 infection activates immediate-early genes. Integrin-ma-trix interaction regulates immediate-early genes (36). We have recently shown that MG-63 and HOS cell adhesion to collagen increases the expression of c-fos, c-jun, and junB genes (39). In the present study, we investigated whether these genes could
also be activated by binding of EV1 to thea2b1integrin. First,
activation was analyzed in MG-63 cells, which expressed
func-tional EV1 receptors (a2b1integrin) but were unable to
sup-port a productive infection cycle. To determine whether
acti-vation occurs after virus binding, large amounts (1 and 10mg
[image:2.612.82.275.70.251.2]corresponding to 3 and 30 PFU/cell, respectively) of virus were used. No increase in the mRNA levels of c-jun or junB genes was observed after incubation of the cells with the virus (Fig. 3),
FIG. 1. Binding of EV1 and -11 to MG-63, HOS pAW (HOS), and HOS pa2AW (HOSa2) cells. The cells were first incubated in the presence (hatched bars) or absence (solid bars) of anti-a2integrin antibody (Ab) (12F1), and then the binding of radioactively labeled virus was determined by a scintillation counter.
[image:2.612.315.556.82.130.2]FIG. 2. Replication cycle of EV1 in HOS pa2AW cells. The results of infec-tivity titration are expressed as end point titers.
TABLE 1. Proportions of immunoperoxidase-stained positive cellsa
Cell line/virus Ab2 Ab1
MG-63/EV1 0 0
HOS pa2AW/EV1 46 10
HOS pa2AW/EV7 76 19
aPercentages of cells detected with cross-reactive primary antibody (Ab) against EV11 after infection of cells in the absence or presence of anti-a2 integrin antibody (12F1 for EV1) or anti-DAF antibody (IF7 for EV7).
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.336.530.525.708.2]suggesting that binding of the virus toa2b1integrin as such is not
sufficient to activate the AP-1 components.
In HOS pa2AW cells, permissive for EV1 infection,
induc-tion of c-fos, c-jun, and junB genes was evident after a 10-h incubation period (Fig. 4). The induction was dose dependent,
since 10 mg of the virus caused a clearly more intense signal
than did 1mg. Viral replication was monitored by measuring
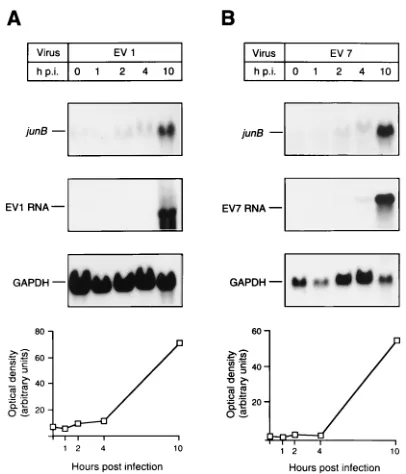
the amount of viral RNA (Fig. 4). No increase in early gene mRNA levels was observed 0.5 h p.i., confirming the finding that postattachment events in the viral replication cycle are necessary for the induction of immediate-early genes, as al-ready seen with MG-63 cells. A more detailed assessment of the time course of junB activation showed that some induction (twofold) already occurred during the first 2 to 4 h p.i. (Fig. 5). The induction of junB was 10-fold at 10 h p.i. (Fig. 5) com-pared with that of uninfected cells, which chronologically cor-responds roughly to the end of the first replication cycle of the virus.
Viral replication is necessary for early gene activation.To determine if the induction of early genes is specific for the EV1
infection cycle with thea2b1 integrin as a receptor for virus
entry, gene activation during EV7 infection was also studied. The virus is known to infect cells through an entry process with
DAF glycoprotein (6, 40). When cells were infected with 5mg
(corresponding to 250 PFU/cell) of EV7, induction similar to that seen in EV1-infected cells was observed, suggesting that active intracellular viral RNA and/or protein synthesis is needed for the activation of immediate-early genes (Fig. 5). We also studied whether immediate-early genes are induced if the receptor-mediated cell entry process is avoided by trans-fection of viral RNA (EV7) into HOS cells with the Lipofectin reagent. A clear induction of junB (10-fold) was observed 8 h p.i. (data not shown), confirming the above-mentioned finding that the activation of immediate-early genes is not mediated by virus-receptor interaction. Only low induction of junB (1.5-fold) was observed 8 h p.i. in the Lipofectin control sample, suggesting that Lipofectin itself does not significantly induce early genes.
The need for viral macromolecule synthesis in the induction of early genes was also studied by using an antiviral compound (WIN 54954), which effectively inhibited the production of infectious virus (data not shown). To study whether this
com-pound prevents the attachment of EV1 to HOS cells, 35
S-labeled EV1 was preincubated with inhibitory concentrations
(1 and 10mg/ml) of WIN 54954 and subsequently used in the
[image:3.612.334.537.67.304.2]binding assay. The WIN compound had no significant effect on the binding of EV1 to HOS cells (binding was inhibited by 6 to 15%). This finding together with those of previous studies (42) suggests that the compound inhibits the uncoating of the virus in echovirus infection. No induction of junB was seen in the
[image:3.612.63.298.68.218.2]FIG. 3. Expression of junB and c-jun genes in MG-63 cells after EV1 inoc-ulation. The cells were first incubated with 1 or 10mg of virus, and RNA was isolated 0.5 and 10 h later. junB (A) and c-jun (B) mRNA levels were determined by Northern hybridizations. GAPDH mRNA levels were used as controls. The results were compared with those obtained with uninfected cells (0 h).
FIG. 4. Northern blot analysis of c-fos, junB, and c-jun mRNA levels in EV1-infected HOS pa2AW cells. The cells were infected with 1 or 10mg of the virus, and RNA was isolated 0.5 and 10 h later. c-fos and junB (A) and c-jun (B) mRNA levels were determined by Northern hybridizations. Determinations of EV1 RNA levels were used for monitoring viral replication (B). GAPDH mRNA levels were determined and used for monitoring cellular mRNA levels. The results were compared with those obtained with uninfected cells (0 h).
FIG. 5. Time course of junB induction in EV1 (A)- and EV7 (B)-infected HOS pa2AW cells. The cells were infected with 5mg of the virus, and RNA was isolated after different time intervals. The proportional quantity of junB mRNA was assayed by Northern hybridizations. EV1 (A) and EV7 (B) RNA levels were determined for monitoring viral replication. GAPDH mRNA levels were deter-mined and used to monitor cellular mRNA levels. Optical densities measured from junB autoradiograms were corrected by means of densities determined with the GAPDH probe. The results are shown under GAPDH Northern hybridiza-tions.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.77.280.496.666.2]presence of the WIN compound 10 h p.i., and EV1 RNA could not be detected when the compound was present (Fig. 6). This reconfirms previous findings of this study that postattachment events in echovirus replication are essential for the induction of immediate-early genes.
Discussion. Human enteroviruses, including echoviruses, are thought to cause lytic infections with rapid cell destruction. In cultured cells, this process is due to active viral macromol-ecule synthesis and specific shutoff of host cell protein synthe-sis (30). However, the exact mechanisms of cell death remain to be determined. In general, the effects of the infection on cellular functions are still poorly understood. From the patho-genic point of view, it is evident that virus-induced alterations in cellular functions may have important consequences during infection. For instance, the production of virus elsewhere than in its replication site of the body may have pathogenic effects on cells and tissues expressing virus receptors even though they themselves may not be capable of supporting a complete rep-lication cycle.
The AP-1 transcription factors, including c-Jun, JunB, and c-Fos, are induced and regulated by several DNA viruses, suggesting that AP-1 may have a role in virus replication and cell transformation. In hepatitis B virus infections, the X pro-tein stimulates c-Jun N-terminal kinases, which leads to the induction and activation of AP-1 (4, 38). Human papillomavi-rus type 11 E5a oncoprotein has been reported to activate AP-1 transcription factors, including c-Jun, JunB, JunD, and c-Fos (9), whereas the human papillomavirus type 16 E7 gene product interacts with AP-1 transcription factors (3). It has also been shown that the adenovirus E1A gene product and polyomavirus middle-sized tumor antigen activate Jun/AP-1 transcription factor (13, 34). Recently, it was found that
infec-tion by an RNA virus, Theiler’s murine encephalomyelitis virus (a picornavirus, as are echoviruses), in glial cells also leads to the overexpression of c-fos and c-jun (29). In the present study, we went further and were able to show that EV1 and -7 can also regulate AP-1 gene expression. This study gives further evidence that virus-mediated induction of AP-1 transcription factor is not necessarily related only to the cell transformation process but probably also to other intracellular events.
It became evident from several viewpoints that viral repli-cation is required for the induction of c-jun, junB, and c-fos. Firstly, stimulation of these genes was not seen in MG-63 cells, which bind the virus but do not support virus replication. Sec-ondly, no induction was seen in the presence of antiviral com-pound WIN 54954, which does not prevent virus attachment but probably inhibits the uncoating of the virus. Thirdly, the induction was not integrin specific since it was also observed after infection by EV7, known to use a different cell entry route (6, 40). Finally, induction was also detected when the receptor-mediated cell entry process was completely avoided by using viral RNA transfection.
Thus, the possibility that induction is directly mediated by the interaction of viruses with their cellular receptors was ruled out, suggesting that the attachment of EV1 and collagen to VLA-2 leads to different cellular consequences. This is also in agreement with the finding that, in an adhesion assay, HOS cells were found to adhere to both EV1 and collagen but only spread on collagen was observed (data not shown). Postattach-ment events during EV1 and EV7 replication, however, effec-tively activate immediate-early genes. This may be due to the direct effects of viral macromolecule synthesis or the induction of secondary responses (for instance, cytokines), which further mediate the activation of early genes.
To study the activation of cytokines in EV1-infected cells, we tested three candidates for induction: interleukin-1 (IL-1), IL-6, and tumor necrosis factor alpha. No induction of IL-1 or tumor necrosis factor alpha was observed during EV1 or EV7 infection, but IL-6 induction was seen 10 h p.i. (data not shown). This is noteworthy, since the junB promoter has been reported to contain an IL-6 response element (24), and thus the induction of junB could be mediated by IL-6. However, the activation of immediate-early genes through this mechanism is unlikely in echovirus infections, because twofold induction of
junB could already be observed 4 h p.i., whereas IL-6 induction
did not appear until 10 h p.i. It is possible that, later during the infection, the upregulation of IL-6 enhances JunB/AP-1 induc-tion.
Whether the activation of immediate-early genes has stim-ulatory or inhibitory effects on echovirus replication remains to be determined. Other transcription factors have been impli-cated in the regulation of viral DNA replication (18). Further-more, it was shown recently that AP-1 enhances viral DNA replication by promoting unwinding of DNA in vitro in poly-omavirus infection (17). It is not known whether AP-1 has a similar role during the replication of RNA viruses.
[image:4.612.113.245.67.321.2]Ten hours p.i., when clear induction of immediate-early genes was observed, most of the cells showed typical CPE, and 46 to 76% were positive by immunostaining (Table 1). The exact role of the immediate-early gene induction in cell death remains unknown. The possibility that the induction of these genes triggers apoptosis cannot be directly concluded on the basis of the available data, but further experiments are needed to solve this question. It is noteworthy that the upregulation of c-jun, c-fos, and junB has been reported to be related to apo-ptosis (25). In some cell lines, the transcription factor AP-1 is critical for apoptosis (33), whereas in other cell lines apoptosis can take place in the absence of AP-1 components c-Fos and
FIG. 6. Effect of antiviral compound WIN 54954 on the induction of junB by EV1 in HOS pa2AW cells. The cells were incubated with 5mg of the virus, which had been preincubated in the presence (1) or absence (2) of the WIN com-pound, and RNA was isolated 10 h later. junB mRNA levels were determined by Northern hybridization. EV1 RNA levels were determined and used for moni-toring viral replication. GAPDH mRNA levels were determined and used for monitoring cellular RNA levels in the samples.
on November 9, 2019 by guest
http://jvi.asm.org/
c-Jun (28). Apoptosis, at least partially, could explain the in-duction of the early genes as detected in this study. It is known that at least poliovirus (an enterovirus, as are the echoviruses [37]), measles virus (14), varicella-zoster virus (31), human immunodeficiency virus type 1 (41), hepatitis C virus (22), influenza virus (23), and Epstein-Barr virus (21) are able to induce apoptosis in vivo or in cell cultures. In poliovirus infec-tion, specific host cell protein shutoff is assumed to have an apoptotic effect, whereas viral proteins exhibit antiapoptotic activity (37). It is possible that echovirus infection also triggers these two processes.
The study was financially supported by grants from the Academy of Finland and The Turku Graduate School of Biomedical Sciences.
We thank Sanofi Winthrop, Inc., for the WIN 54954 compound, Virgil Woods for the 12F1 antibody, and Jeffrey Bergelson for the IF7 antibody. Tapani Hovi, Ilkka Julkunen, and Antti Vaheri are acknowl-edged for stimulating discussions, and Simo Merne is acknowlacknowl-edged for checking the manuscript.
REFERENCES
1. Abraham, G., and R. J. Colonno. 1984. Many rhinovirus serotypes share the same cellular receptor. J. Virol. 51:340–345.
2. Angel, P., E. A. Allegretto, S. T. Okino, K. Hattori, W. J. Boyle, T. Hunter,
and M. Karin.1988. Oncogene jun encodes a sequence-specific trans-acti-vator similar to AP-1. Nature 332:166–171.
3. Antinore, M. J., M. J. Birrer, D. Patel, L. Nader, and D. J. McCance. 1996. The human papillomavirus type 16 E7 gene product interacts with and trans-activates the AP1 family of transcription factors. EMBO J. 15:1950– 1960.
4. Benn, J., F. Su, M. Doria, and R. J. Schneider. 1996. Hepatitis B virus HBx protein induces transcription factor AP-1 by activation of extracellular sig-nal-regulated and c-Jun N-terminal mitogen-activated protein kinases. J. Vi-rol. 70:4978–4985.
5. Bergelson, J. M., B. M. C. Chan, R. W. Finberg, and M. E. Hemler. 1993. The integrin VLA-2 binds echovirus 1 and extracellular matrix ligands by differ-ent mechanisms. J. Clin. Invest. 92:232–239.
6. Bergelson, J. M., M. Chan, K. R. Solomon, N. F. St. John, H. Lin, and R. W.
Finberg.1994. Decay-accelerating factor (CD55), a glycosylphosphatidyl-inositol-anchored complement regulatory protein, is a receptor for several echoviruses. Proc. Natl. Acad. Sci. USA 91:6245–6248.
7. Bergelson, J. M., M. P. Shepley, B. M. C. Chan, M. E. Hemler, and R. W.
Finberg.1992. Identification of the integrin VLA-2 as a receptor for echo-virus 1. Science 225:1718–1720.
8. Bergelson, J. M., N. St. John, S. Kawaguchi, M. Chan, H. Stubdal, J. Modlin,
and R. W. Finberg.1993. Infection of echoviruses 1 and 8 depends on thea2 subunit of human VLA-2. J. Virol. 67:6847–6852.
9. Chen, S.-L., Y.-P. Tsao, C.-M. Yang, Y.-K. Lin, C.-H. Huang, and S.-W. Kuo. 1995. Differential induction and regulation of c-jun, junB, junD and c-fos by human papillomavirus type 11 E5a oncoprotein. J. Gen. Virol. 76:2653–2659. 10. Chirgwin, J. M., A. E. Przybyla, R. J. MacDonald, and W. J. Rutter. 1979. Isolation of biologically active ribonucleic acid from sources enriched in ribonuclease. Biochemistry 10:5294–5299.
11. Clover, J., and M. Gowen. 1994. Are MG-63 and HOS TE85 human osteo-sarcoma cell lines representative models of the osteoblastic phenotype? Bone 15:585–591.
12. Dahllund, L., L. Nissinen, T. Pulli, V.-P. Hyttinen, G. Stanway, and T.
Hyypia¨.1995. The genome of echovirus 11. Virus Res. 35:215–222. 13. DeGroot, R., N. Foulkes, M. Mulder, W. Kruijer, and P. Sassone-Corsi.
1991. Positive regulation of jun/AP1 by E1A. Mol. Cell. Biol. 11:192–201. 14. Esolen, L. M., S. W. Park, J. M. Hardwick, and D. E. Griffin. 1995. Apoptosis
as a cause of death in measles virus-infection cells. J. Virol. 69:3955–3958. 15. Fort, P. L., L. Marty, M. Piechaczyk, S. El Sabrouty, C. Dani, P. Jeanteur,
and J. M. Blanchard.1985. Various rat adult tissues express only one major mRNA species from the glyceraldehyde-3-phosphate-dehydrogenase multi-genic family. Nucleic Acids Res. 13:1431–1442.
16. Grist, N. R., E. J. Bell, and F. Assaad. 1978. Enteroviruses in human disease. Prog. Med. Virol. 24:114–157.
17. Guo, W., W. J. Tang, X. Bu, V. Bermudez, M. Martin, and W. R. Folk. 1996. AP1 enhances polyomavirus DNA replication by promoting T-antigen-me-diated unwinding of DNA. J. Virol. 70:4914–4918.
18. Guo, Z.-S., and M. L. DePamphilis. 1992. Specific transcription factors stimulate simian virus 40 and polyomavirus origins of DNA replication. Mol. Cell. Biol. 12:2514–2524.
19. Huttunen, P., J. Santti, T. Pulli, and T. Hyypia¨. 1996. The major echovirus group is genetically coherent and related to coxsackie B viruses. J. Gen. Virol. 77:715–725.
20. Kamata, T., W. Puzon, and Y. Takada. 1994. Identification of putative ligand binding sites within I domain of integrina2b1(VLA-2, Cd49b/CD29). J. Biol. Chem. 269:9659–9663.
21. Lu, J. J.-Y., J.-Y. Chen, T.-Y. Hsu, W. C. Y. Yu, I.-J. Su, and C.-S. Yang. 1996. Induction of apoptosis in epithelial cells by Epstein-Barr virus latent mem-brane protein 1. J. Gen. Virol. 77:1883–1892.
22. Mita, E., N. Hayashi, S. Lio, T. Takehara, T. Hijioka, A. Kasahara, H.
Fusamoto, and T. Kamada.1994. Role of Fas ligand in apoptosis induced by hepatitis C virus infection. Biochem. Biophys. Res. Commun. 204:468–474. 23. Mori, I., T. Komatsu, K. Takeuchi, K. Nakakuki, M. Sudo, and Y. Kimura. 1995. In vivo induction of apoptosis by influenza virus. J. Gen. Virol. 76: 2869–2873.
24. Nakajima, K., T. Kusafuka, T. Takeda, Y. Fujitani, K. Nakae, and T. Hirano. 1993. Identification of a novel interleukin-6 response element containing an Ets-binding site and a CRE-like site in the junB promoter. Mol. Cell. Biol.
13:3027–3041.
25. Pandey, S., and E. Wang. 1995. Cells en route to apoptosis are characterized by the upregulation of c-fos, c-myc, cdc2, and RB phosphorylation, resem-bling events of early cell-cycle traverse. J. Cell. Biochem. 58:135–150. 26. Pischel, K. D., M. E. Hemler, C. Huang, H. G. Bluestein, and V. L. Woods.
1987. Use of the monoclonal antibody 12F1 to characterize the differentia-tion antigen VLA-2. J. Immunol. 138:226–233.
27. Riikonen, T., J. Westermarck, L. Koivisto, A. Broberg, V.-M. Ka¨ha¨ri, and J.
Heino.1995. Integrina2b1is a positive regulator of collagenase (MMP-1) and collagena1 (I) gene expression. J. Biol. Chem. 270:13548–13552. 28. Roffler-Tarlov, S., J. J. Brown, E. Tarlov, J. Stolarov, D. L. Chapman, M.
Alexiou, and V. E. Papaioannou.1996. Programmed cell death in the ab-sence of c-Fos and c-Jun. Development 122:1–9.
29. Rubio, N., P. Rojo, and C. Torres. 1996. Overexpression of basal c-fos and
c-jun but not of ras oncogenes after Theiler’s murine encephalomyelitis virus
infection of glial cells. J. Neurovirol. 2:167–174.
30. Rueckert, R. R. 1996. Picornaviridae: viruses and their replication, p. 636– 641. In B. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Virology. Lippincott-Raven Publishers, Philadelphia, Pa.
31. Sadzot-Delvaux, C., P. Thonard, S. Schoonbroodt, J. Piette, and B. Rentier. 1995. Varicella-zoster virus induces apoptosis in cell culture. J. Gen. Virol.
76:2875–2879.
32. Santala, P., H. Larjava, L. Nissinen, T. Riikonen, A. Ma¨a¨tta¨, and J. Heino. 1994. Suppressed collagen gene expression and induction ofa2b1 integrin-type collagen receptor in tumorigenic derivatives of human osteogenic sar-coma (HOS) cell line. J. Biol. Chem. 269:1276–1283.
33. Sawai, H., T. Okazaki, H. Yamamoto, H. Okano, Y. Takeda, M. Tashima, H.
Sawada, M. Okuma, H. Ishikura, H. Umehara, and N. Domae.1995. Re-quirement of AP-1 for ceramide-induced apoptosis in human leukemia HL-60 cells. J. Biol. Chem. 270:27326–27331.
34. Schonthal, A., S. Srinivas, and W. Eckhart. 1992. Induction of c-jun pro-tooncogene expression and transcription factor AP-1 activity by the polyoma virus middle-sized tumor antigen. Proc. Natl. Acad. Sci. USA 89:4972–4976. 35. Schu¨tte, J., J. Viallet, M. Nau, S. Segal, J. Fedorko, and J. Minna. 1989.
Jun-B inhibits and c-fos stimulates the transforming and trans-activating
activities of c-jun. Cell 89:987–997.
36. Schwartz, M. A., M. D. Schaller, and M. H. Ginsberg. 1995. Integrins: emerging paradigms of signal transduction. Annu. Rev. Cell. Dev. Biol.
11:549–599.
37. Tolskaya, E. A., L. I. Romanova, M. S. Kolesnikova, T. A. Ivannikova, E. A.
Smirnova, N. T. Raikhlin, and V. I. Agol.1995. Apoptosis-inducing and apoptosis-preventing functions of poliovirus. J. Virol. 69:1181–1189. 38. Twu, J.-S., M.-Y. Lai, D.-S. Chen, and W. S. Robinson. 1993. Activation of
proto-oncogene c-jun by the X protein of hepatitis B virus. Virology 192: 346–350.
39. Vihinen, P., V.-M. Ka¨ha¨ri, and J. Heino. Integrin-mediated cell adhesion regulates the expression of immediate early genes and modifies the effect of growth factors. Unpublished results.
40. Ward, T., P. A. Pipkin, N. A. Clarkson, D. M. Stone, P. D. Minor, and J. W.
Almond.1994. Decay-accelerating factor CD55 is identified as the receptor for echovirus 7 using CELICS, a rapid immuno-focal cloning method. EMBO J. 13:5070–5074.
41. Weller, P. F., W. L. Marshall, D. R. Lucey, T. H. Rand, A. M. Dvorak, and
R. W. Finberg.1995. Infection, apoptosis, and killing of mature human eosinophils by human immunodeficiency virus-1. Am. J. Respir. Cell. Mol. Biol. 13:610–620.
42. Zhang, A., R. G. Nanni, D. A. Oren, E. J. Rozhon, and E. Arnold. 1992. Three-dimensional structure-activity relationships for antiviral agents that interact with picornavirus capsids. Semin. Virol. 3:453–471.