0022-538X/96/$04.00
1
0
Copyright
q
1996, American Society for Microbiology
Demonstration of Binding of Dengue Virus Envelope
Protein to Target Cells
YAPING CHEN,
1TERRY MAGUIRE,
2ANDRORY M. MARKS
1*
Department of Internal Medicine, University of Michigan Medical School, Ann Arbor, Michigan 48109-0531,
1and Virus Research Group and Centre for Gene Research, Health Research Council of New Zealand,
University of Otago, Dunedin, New Zealand
2Received 17 June 1996/Accepted 8 September 1996
The nature of the initial interaction of dengue virus with target cells and the extent to which this interaction
defines tropism are unknown. Infection of some cells may involve antidengue antibody-mediated immune
adherence to cells bearing immunoglobulin Fc receptors; however, this mechanism does not explain primary
infection or the infection of cells without Fc receptors. We hypothesized that dengue virus envelope protein
mediates initial binding to target cells. To test this hypothesis, a recombinant chimeric form of dengue type 2
virus envelope protein was used as a probe to investigate binding to the surfaces of potential target cells.
Envelope protein was expressed amino terminal to the heavy-chain constant region of human immunoglobulin
G containing the Fc receptor binding motif; the binding mediated by envelope determinants was
distinguish-able from the binding mediated by immunoglobulin Fc determinants. We found that the recombinant chimera
bound to Vero, CHO, endothelial, and glial cells through envelope protein determinants and to monocytes and
U937 cells by Fc-Fc receptor interactions. The highest level of binding was to Vero cells; binding was dose and
time dependent and saturable. Examination of partial-length recombinant envelope proteins indicated that the
binding motif was expressed between amino acids 281 and 423. Recombinant envelope protein inhibited
infection of Vero cells by dengue virus, indicating the functional significance of the interaction of envelope
protein and target cells in infectivity. These results suggest that envelope protein binding to a non-Fc receptor
could explain the cell and tissue tropism of primary dengue virus infection.
Dengue virus is an arthropod-borne human pathogen that
represents a serious public health threat. Dengue virus
infec-tion is pandemic in third-world tropical areas, and the
reemer-gence of infection in developed countries, including endemic
transmission within the United States in 1995, is of increasing
public health concern (10, 19, 29). The clinical manifestations
of dengue virus infection range in severity from a simple febrile
illness to a hemorrhagic fever and a potentially fatal
hemor-rhagic shock syndrome (17, 18). There is no specific treatment
for infection, and no vaccine is yet available.
One of the most important questions about the pathogenesis
of dengue virus is the mechanism whereby the virus binds to
target cells. Dengue virus gains entry to some cell types by an
immune adherence bridging phenomenon, which is dependent
on the presence of nonneutralizing antibodies (known as
an-tibody-dependent enhancement of viral infectivity [30]). The
Fc domain of the antibody that is bound to the virus mediates
binding to cells such as macrophages that express Fc receptors;
then, fusion, internalization, and productive infection can
oc-cur. This mechanism cannot explain primary infections in
pa-tients without antibody to dengue virus, and it cannot explain
the ability of dengue virus to infect nonphagocytic cell types
that do not express Fc receptors (22).
The basis of the cell and tissue tropism of microorganisms,
including viruses, is often defined by the ability of a surface
molecule to bind to specific target cell surface receptors (40).
Viral binding is followed by membrane fusion and leads to
productive infection in suitable hosts. A wide range of
physi-ologically important cell surface receptors have been subverted
by viruses to enable initial target cell binding (42). There is no
information about the cellular tropism of dengue virus or of
any other flaviviruses; however, observations that dengue and
other flaviviruses undergo rapid binding to the surfaces of
target cells suggest a receptor-mediated process (21).
Dengue virus has a relatively simple structure; the envelope
protein is the major structural protein exposed on the surface
(33). It is therefore likely that initial binding of dengue virus to
target cells is mediated by binding of the envelope protein to a
specific, but as yet unidentified, cell surface receptor. One
previous study identified radiolabeled dengue virus envelope
protein bound to the surface of target cells (1); in this work,
envelope protein was derived from infected culture
superna-tants, and it was thus uncertain if binding was mediated
spe-cifically by envelope protein or by another viral or culture
supernatant-associated component.
In order to study the cellular and molecular mechanisms that
contribute to dengue virus tropism for target cells, we
devel-oped a unique recombinant chimeric form of dengue virus
envelope protein. The envelope protein was expressed as a
fusion with the heavy-chain constant region of human
immu-noglobulin G (IgG) that contains the motif responsible for Fc
receptor binding; binding mediated by viral envelope protein
and immunoglobulin determinants could be distinguished. We
utilized this construct to test binding to different cell types and
were able to draw conclusions regarding the presence of a
specific dengue virus envelope protein receptor on target cells.
MATERIALS AND METHODS
Envelope protein construct.The dengue type 2 virus envelope protein cDNA (Tonga 1974 strain [GenBank accession number X54319]) was obtained as a plasmid clone pTZD2E4 (7–9). The region encoding the envelope protein, ex-tending from the start codon to the beginning of the transmembrane region (nucleotides 1 to 1272), was subcloned by PCR amplification. Additional nucle-otides were included in the sense strand oligonucleotide to generate an upstream
* Corresponding author. Mailing address: Dept. of Internal
Medi-cine, Univ. of Michigan, Kresge I, Room 4570, Ann Arbor, MI
48109-0531. Phone: (313) 936-3257. Fax: (313) 763-2025. Electronic mail
address: [email protected].
8765
on November 9, 2019 by guest
http://jvi.asm.org/
NheI restriction site. The sense strand oligonucleotide primer sequence was 59-CTA GCT AGC GAT GCG CTG CAT AGG AAT ATC AAA TAG GGA-39
(the NheI site is underlined). Two oligonucleotide primers for the antisense strand were utilized sequentially for PCR in order to add sequence to the downstream region, encoding an in-frame duplex heart muscle kinase (HMK) target site (single-letter amino acid code, RRASVGRRASV [6]), followed by a BglII restriction site. The first antisense strand oligonucleotide primer had the sequence 59-ACC TAC AGA TGC ACG TCG AGA TCC AAA ATC CCA GGC TGT-39(the HMK target site is underlined). PCR was initially performed with the sense strand oligonucleotide and the first antisense strand oligonucle-otide. The product was recovered with a QIAquick PCR purification kit (Qiagen, Chatsworth, Calif.) and was used as the template for a second round of PCR, using the same sense strand oligonucleotide and the second antisense strand oligonucleotide sequence 59-CGG AAG ATC TAC TGA TGC ACG ACG ACC TAC AGA TGC ACG TCG-39. The final PCR product was digested with NheI and BglII (the HMK target site is underlined, and the BglII site is italicized).
The plasmid utilized for expression was based on pcDNA3 (Invitrogen, San Diego, Calif.), a vector designed for eukaryotic expression that incorporates enhancer-promoter sequences from the immediate-early gene of cytomegalovi-rus, polyadenylation signal and transcription termination sequences from the bovine growth hormone gene, and a simian virus 40 origin of replication. pcDNA3 was modified by ligating the cDNA for the signal peptide of CD5 and part of the heavy-chain constant region of human IgG1(the hinge, CH2, and
CH3 region exons) into the multiple-cloning site (3, 38). This modified plasmid was digested with NheI and BamHI and ligated with the envelope protein con-struct described above. This strategy allowed the envelope protein concon-struct to be ligated in frame, downstream from the sequence encoding the signal peptide of CD5, and upstream from the sequence encoding human IgG1. The CD5 signal peptide was incorporated to facilitate secretion, the IgG domains were incorpo-rated to facilitate purification and detection of expressed protein (see Discus-sion), and the HMK domains were introduced to facilitate radiolabeling by phosphorylation with32P but were not utilized in experiments reported here. The
nucleotide sequences at both the 59and 39termini of the construct were verified by chain termination sequencing, with Sequenase T7 DNA polymerase and oligonucleotides that were based on the flanking plasmid DNA sequence. Large-scale plasmid preparations were purified by Qiagen anion-exchange chromatog-raphy. The construct is named env-IgG.
In order to localize the target cell binding region, two partial-length envelope protein constructs were also generated by the same experimental strategy as was used to produce env-IgG. These constructs, named env(34/253)-IgG and env(281/423)-IgG, incorporated the envelope protein amino acids indicated in parentheses (8). A control construct was also generated and was named HMK-IgG. This construct lacked envelope protein determinants and consisted only of the CD5 signal peptide, HMK target site, and heavy-chain constant region of human IgG1. In this study, the HMK-IgG construct was used as a control for env-IgG.
Expression and purification of envelope protein.Plasmid constructs were transfected into COS-7 cells (ATCC CRL-1651; American Type Culture Collec-tion, Rockville, Md.) by calcium phosphate coprecipitation (36) and cultured posttransfection in Dulbecco’s modified Eagle medium (DMEM) containing 2% fetal bovine serum (FBS), penicillin (100 U/ml), and streptomycin (100mg/ml) (all from Life Technologies Gibco BRL, Gaithersburg, Md.). Supernatants and cell lysates were harvested after 96 h. Cells from each 10-cm-diameter dish were scraped into 0.75 ml of cation-free phosphate-buffered saline (PBS) (Gibco) containing 0.5% Triton X-100 (PBS-T) and protease inhibitors (leupeptin, apro-tinin, soybean trypsin inhibitor, and pepstatin [all 1mg/ml] and phenylmethyl-sulfonyl fluoride [1 mM], all from Sigma, St. Louis, Mo.), sonicated for 1 min, and clarified by centrifugation at 14,0003g for 30 min at 48C. Protease inhibitors were added to culture supernatants, which were clarified as described above.
Recombinant proteins were purified by affinity chromatography over protein A, utilizing its high avidity interaction with the Fc region of IgG. Protein A-Sepharose (Zymed, San Francisco, Calif.) was incubated with 100 ml of cell lysate on a rotating platform overnight at 48C and packed into a column, and nonbound protein was removed by washing the column with 10 bed volumes of PBS-T and then 10 bed volumes of PBS without detergent. Bound protein was eluted in 4 bed volumes of Actisep elution medium (Sterogene, Carlsbad, Calif.) and dialyzed against three changes of PBS. The presence of env-IgG and HMK-IgG was confirmed by denaturing sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) using a Laemmli buffer system (27) and by West-ern blotting (immunoblotting) with antibodies to dengue virus envelope protein and human IgG; the purity of the preparations was assessed by silver staining after SDS-PAGE, and the protein concentrations were measured by a colori-metric assay (Bio-Rad, Hercules, Calif.).
Western blotting.Proteins were resolved on the basis of molecular weight by denaturing SDS-PAGE (7% polyacrylamide) and electrotransferred to nitrocel-lulose membranes (Schleicher & Schuell, Keene, N.H.). The equivalent of 50,000 lysed cells was loaded per lane. Membranes were blocked with 5% nonfat dry milk in PBS. Human IgG epitopes were detected by incubating membranes with peroxidase-conjugated goat antibody to human IgG (Bio-Rad), diluted 1/5,000 in blocking solution, for 1 h at room temperature. After the membranes were washed, antibody binding was detected with an enhanced chemiluminescence kit (Amersham, Arlington Heights, Ill.). Envelope protein epitopes were detected
by incubating membranes with mouse monoclonal antibody 152 to dengue type 2 virus envelope protein (generated by F. Austin, Virus Research Unit, Health Research Council of New Zealand, University of Otago, New Zealand), diluted 1/100 in PBS, for 1 h at room temperature and then with peroxidase-conjugated goat antibody to mouse IgG (Bio-Rad), diluted 1/10,000, for 1 h at room tem-perature. Antibody binding was detected as described above.
Deglycosylation of env-IgG.For N-glycosylation analysis, 4mg of purified env-IgG was denatured by boiling in 0.5% SDS–1%b-mercaptoethanol for 10 min; the preparation was incubated with 23103
U of endo-b -N-acetylglu-cosaminidase F (PNGaseF) (New England Biolabs, Beverly, Mass.) in buffer made to 50 mM sodium phosphate (pH 7.5)–1% Nonidet P-40 for 1 h at 378C. For O-glycosylation analysis, 4mg of purified env-IgG was incubated with 4 mU of O-glycosidase (Boehringer Mannheim, Indianapolis, Ind.) in PBS, pH 6.0, for 6 h at 378C. After enzyme treatment, SDS-PAGE and Western blotting were performed, as described above.
Cell preparation.The following cell types were cultured in the media indi-cated: COS-7, Dulbecco’s modified Eagle medium with 10% FBS; Vero (ATCC CCL-81), medium 199 with 5% FBS; glial (ATCC CCL-107), Ham’s F-10 me-dium with 15% heat-inactivated horse serum and 5% FBS; Chinese hamster ovary (CHO-K1, ATCC CCL-61), a-MEM with deoxyribonucleotides, ribo-nucleotides, and 10% FBS; and U937 (ATCC CRL-1593), RPMI 1640 medium with 10% FBS. U937 cells were induced to differentiate by treatment with 1.63
1028M phorbol myristate acetate (Sigma) for 5 days, and Fc receptor expression
was further increased by treatment with 100 U of recombinant human gamma interferon (Boehringer Mannheim) per ml for 18 h. Human umbilical vein endothelial cells were isolated from freshly harvested umbilical cords and cul-tured in medium 199 with 20% FBS, 10 ng of recombinant human basic fibroblast growth factor (Synergen, Boulder, Colo.) per ml, and 100 ng of bovine lung heparin (Sigma) per ml in gelatin (Sigma)-coated tissue culture dishes (25). All culture media were supplemented with penicillin (100 U/ml) and streptomycin (100mg/ml), and cells were grown in 10-cm-diameter tissue culture dishes (Corn-ing, Corn(Corn-ing, N.Y.). All culture media and additives were obtained from Gibco, unless otherwise stated.
Peripheral venous blood was obtained from healthy human donors, after informed consent was obtained by a protocol approved by the University of Michigan Medical School Institutional Review Board. Leukocytes were fraction-ated into mononuclear and polymorphonuclear (principally neutrophil) cell pop-ulations by density gradient centrifugation (Polymorphprep; Gibco). Leukocytes were washed twice with PBS by centrifugation at 2003g for 7 min at 48C, kept at 48C, and used within 1 h of preparation.
Envelope protein cell binding assay.Adherent cells were detached from the culture surface by incubation with PBS–10 mM EDTA (Gibco) for 10 min at 378C followed by gentle agitation. Cells were washed twice by centrifugation through DMEM and then 33105cells were incubated with env-IgG, control
HMK-IgG, or control normal human IgG (Sigma) for 1 h at 48C in 30ml of PBS–1% normal goat serum (PBS-GS) (Gibco). Cells were washed twice by centrifugation through PBS-GS at 2003g for 10 min at 48C. Cells were then incubated with the R-phycoerythrin-labeled F(ab)2fragment of affinity-purified
goat antibody to human IgG Fc (5mg/ml; Jackson Labs, West Grove, Pa.) for 1 h at 48C and washed as described above. In some experiments, cells were prein-cubated with mouse IgG1 (1 mg/ml; Sigma) to occupy Fc receptors, before incubation with env-IgG or control proteins, and binding was then assayed as described above.
Cellular fluorescence and forward- and side-angle light scatter were quanti-tated by flow cytometry (FACScan; Becton Dickinson, Mountain View, Calif.). A minimum of 10,000 cells was assessed from each treatment group. Data were accumulated only for viable cells; nonviable cells were excluded from analysis on the basis of low-level forward-angle light scatter. Results were expressed as contour plots of light scatter and fluorescence per cell and as frequency histo-grams of fluorescence per cell. Fluorescence was measured on a 104-U
logarith-mic scale. A value for median fluorescence intensity was derived from the fluorescence frequency histograms for each sample and used to compare fluo-rescence between samples. In each experiment, controls consisted of untreated cells, cells incubated with only the secondary fluorochrome-labeled antibody, and cells incubated with either normal human IgG or HMK-IgG followed by the secondary fluorochrome-labeled antibody.
Inhibition of infection with dengue virus.Vero cells were seeded at 500 cells per well and cultured overnight in 96-well plates (Nunc, Naperville, Ill.) in MEM (Sigma) with 10% FBS, penicillin (100 U/ml), and streptomycin (100mg/ml; Gibco). A 60-mg/ml concentration of env-IgG, control HMK-IgG, control human IgG, or buffer used in preparation of recombinant proteins was added to wells and incubated for 20 min at 378C. Additional wells were left untreated. Tenfold serial dilutions, from 1021to 1027, of a stock of freshly prepared dengue type 2
New Guinea strain C virus were then added and incubated for 5 min at 48C. Nonbound virus was removed by washing each well three times with PBS. Fresh culture medium (300ml) was then added, and the cells were incubated at 378C in a humidified gassed (5% CO2) incubator and observed daily for cytopathic
effects for 7 days. Dengue virus infection was confirmed as the cause of cyto-pathic effects in selected wells by immunofluorescence analysis using a dengue type 2 virus-specific monoclonal antibody. A total of 16 wells were assessed for each experimental condition, and 32 wells were assessed for the untreated cells. The titer of the 50% infectious dose (ID50titer) was determined as the viral
on November 9, 2019 by guest
http://jvi.asm.org/
dilution at which 50% of the wells demonstrated cytopathic effects. The mean infectious titers and standard deviation were derived by cumulative averaging (13, 31).
RESULTS
Recombinant proteins.
The genomic structures of dengue
virus and the derived envelope protein expression construct
are illustrated in Fig. 1. The amino terminus of env-IgG is
made up of the signal peptide of CD5 followed by the
424-amino-acid ectodomain of the dengue type 2 virus envelope
protein (hydrophobic transmembrane domains deleted). This
is followed by an HMK target site (introduced to allow
high-level activity radiolabeling with
32P, but not utilized in these
experiments). The carboxy terminus of the molecule is made
up of part of the heavy-chain constant region of human IgG1
(including the region required for Fc receptor binding).
Cell lysates and supernatants were prepared from
trans-fected COS cells. Recombinant env-IgG and HMK-IgG
pro-teins were detected by Western blotting following gel
electro-phoresis under reducing conditions, with antibodies specific for
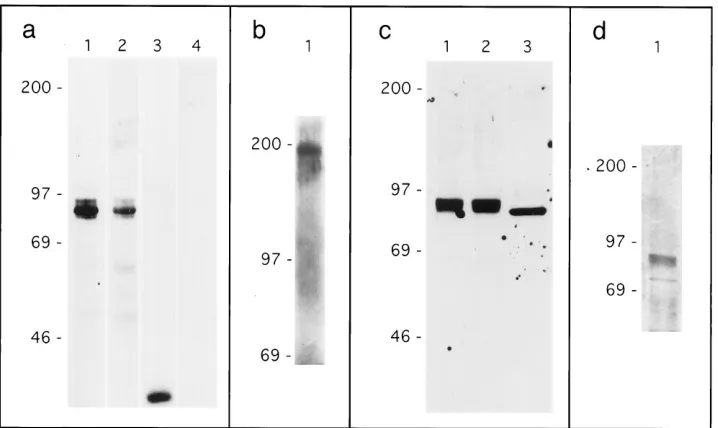
envelope protein and for human IgG (Fig. 2a). Multiple
env-IgG isoforms, migrating with apparent molecular weights
be-tween 90,000 and 96,000, were detected by both antibodies in
the env-IgG-transfected cell lysate (lanes 1 and 2), as expected
for a recombinant protein expressing both envelope protein
and immunoglobulin determinants. No significant env-IgG was
detected in the supernatant, and all recombinant env-IgG was
purified from cell lysates. A similar Western blot assay was
performed with the two partial-length envelope protein
con-structs, env(34/253)-IgG and env(281/423)-IgG. The results
in-dicated that dengue virus antibody 152 recognized an epitope
expressed by amino acids 281 to 423 but not by amino acids 34
to 253 (not shown). A single band with an apparent molecular
weight of 41,000 was detected in the HMK-IgG-transfected cell
supernatant, but only by the antibody to human IgG (lanes 3
and 4), as expected for a recombinant protein expressing
im-munoglobulin but not envelope protein determinants.
HMK-IgG used for subsequent studies was purified from cell
super-natants. Control nontransfected cell lysates and supernatants
were negative by Western blotting with both antibodies (not
shown).
[image:3.612.64.296.69.125.2]The human IgG1 determinants expressed by the
recombi-nant chimeric env-IgG contain the site of an interchain
disul-fide bond responsible for expression of IgG as a homodimer.
To determine if env-IgG was also expressed in homodimeric
form, env-IgG was subjected to gel electrophoresis under
non-reducing conditions followed by Western blotting with
anti-body to human IgG (Fig. 2b). The predominant band observed
migrated with an apparent molecular weight of 190,000, twice
FIG. 1. Dengue virus genome and envelope protein construct. (a) 59region of dengue virus genome. The three dengue virus structural proteins capsid (C), premembrane (PrM), and envelope (Env.), followed by nonstructural proteins (NS), are encoded at the 59end of the RNA genome. Numbers and arrows indicate nucleotides encoding each domain. Envelope protein makes use of the carboxy terminus of the premembrane protein as a signal peptide (solid bar). The hydrophobic transmembrane region of the envelope protein is indicated by the hatched area. (b) Dengue envelope protein expression construct. Nucleotides 937 to 2208 of the dengue virus cDNA (8) were subcloned into an expression construct downstream from the signal peptide of CD5 (CD5 sig. pep.) and upstream from a duplex HMK target site followed by part of the heavy-chain constant region of human IgG1. Expression was driven by cytomegalovirus-derived promoter and enhancer sequences (CMV P/E). Polyadenylation signal and transcription termination sequences were derived from the bovine growth hormone gene (BGH pA).
FIG. 2. Characterization of recombinant proteins. (a) env-IgG and HMK-IgG epitope expression. Cell lysates from COS cells transfected with pcDNA3-env-IgG, were subjected to gel electrophoresis (SDS-PAGE) under reducing conditions followed by Western blotting with antibodies to human IgG (lane 1) and dengue virus envelope protein (lane 2). Supernatants from pcDNA3-HMK-IgG transfectants were similarly analyzed with antibodies to human IgG (lane 3) and dengue virus envelope protein (lane 4). (b) env-IgG expression as a homodimer. Cell lysate from COS cells transfected with pcDNA3-env-IgG was subjected to gel electrophoresis under nonreducing conditions followed by Western blotting with antibody to human IgG (lane 1). (c) Deglycosylation analysis. Protein A-purified env-IgG was digested with endoglycosidases; samples were subjected to gel electrophoresis under reducing conditions followed by Western blotting with antibody to human IgG. Lane 1, mock digestion; lane 2, digestion with O-glycosidase; lane 3, digestion with PNGaseF. (d) Silver staining of env-IgG. Protein A-purified env-IgG was resolved by gel electrophoresis followed by silver staining. For all panels, molecular weight markers (in thousands) are indicated on the left.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.128.487.444.658.2]that of env-IgG exposed to reducing conditions, which is
con-sistent with the expression of env-IgG as a homodimer.
Based on amino acid content, the predicted molecular
weight of the env-IgG construct is 89,000; however, multiple
isoforms of env-IgG with apparent molecular weights ranging
from 90,000 to 96,000 were detected (Fig. 2a). Both the
enve-lope protein and human IgG contain consensus sites for
N-linked glycosylation. To determine if a posttranslational
mod-ification such as glycosylation could account for the apparent
additional molecular weight, protein A-purified env-IgG was
digested with the endoglycosidases PNGaseF (cleaves all
classes of N-linked carbohydrate chains at the
peptide-carbo-hydrate linkage [15]) and O-glycosidase (cleaves some
O-gly-cosidic linkages between N-acetylgalactosamine and serine or
threonine [41]) and then subjected to gel electrophoresis and
Western blotting with antibody to human IgG (Fig. 2c).
Mock-digested env-IgG is shown in lane 1. O-Glycosidase had no
effect on the migration of env-IgG (lane 2); in contrast,
diges-tion with PNGaseF reduced the apparent molecular weight to
89,000, consistent with the additional molecular weight, 7,000,
being due to N-linked glycosylation (lane 3). In other
experi-ments (not shown), digestion of HMK-IgG with PNGaseF
resulted in a reduction of molecular weight of 3,000; this result
indicates that the envelope protein sequence was associated
with a glycan content that accounted for a molecular weight of
4,000.
To generate sufficient quantities for cell binding studies,
env-IgG was batch purified from transfected cell lysates by
protein A affinity chromatography. To assess the purity of the
preparation, env-IgG was subjected to gel electrophoresis,
fol-lowed by silver staining (Fig. 2d). env-IgG isoforms (molecular
weight, 90,000 to 96,000 [previously identified by Western
blot-ting]) represented approximately 80% of total protein present.
Env-IgG binding to cultured cells.
Vero cells, derived from
African green monkey kidney, are frequently used as a target
for studying dengue virus infection in vitro (23, 37); thus, these
cells were selected as the principal target for cell binding
stud-ies. Cells were grown as adherent cultures, resuspended
with-out the use of proteases, incubated with env-IgG or control
normal human IgG or left untreated, and then were incubated
with a fluorochrome-labeled antibody to human IgG; binding
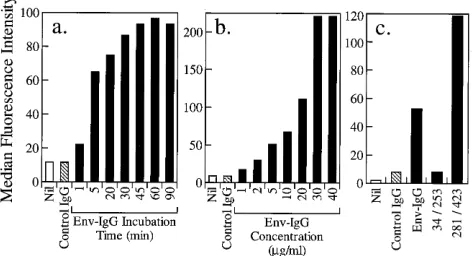
was assessed by flow cytometry. Figure 3a presents the time
course of binding of env-IgG to Vero cells at 4
8
C and
demon-strates strong env-IgG-specific binding that becomes saturated
after 45 min. Binding of control human IgG was no greater
than the background level (secondary antibody alone),
indicat-ing that there was no IgG Fc-mediated bindindicat-ing to Vero cells
and that env-IgG binding was attributable solely to envelope
protein determinants. Figure 3b demonstrates the effects of
increasing concentrations of env-IgG on Vero cell binding at
4
8
C. A dose-dependent increase in binding, which became
saturated at a concentration of 30
m
g/ml, was observed.
Bind-ing of control human IgG (20
m
g/ml) was no greater than the
background level (secondary antibody alone).
The binding activity of the partial-length envelope protein
constructs, env(34/253)-IgG and env(281/423)-IgG, was also
tested on Vero cells (10
m
g/ml for 60 min at 4
8
C). The
con-struct that expressed amino acids 34 to 253 did not bind to
Vero cells. In contrast, the construct that expressed amino
acids 281 to 423 exhibited greater Vero cell binding than did
the full-length env-IgG construct, thus indicating that the
tar-get cell binding motif resides in the envelope protein
carboxy-terminal region (Fig. 3c).
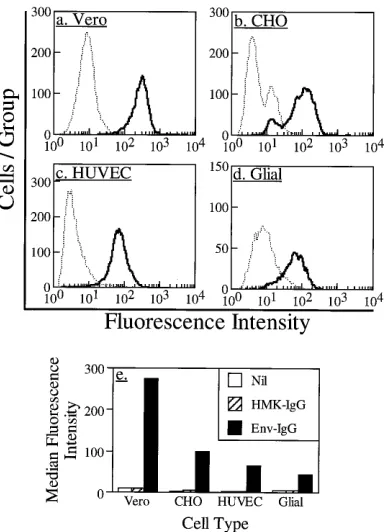
The binding of env-IgG to the following three additional cell
lines was then assessed: (i) CHO cells, derived from hamsters,
which are susceptible to dengue virus infection (39); (ii) human
endothelial cells, which can be infected in vitro and could be a
natural target of dengue virus infection (2); and (iii) human
glial cells, which are relatively resistant to dengue virus
infec-tion (24). Binding to all these cell types was observed; however,
there were distinct and reproducible differences in the binding
profiles (in order from strongest to weakest binding, Vero,
CHO, endothelial, glial). Figures 4a to d are fluorescence
his-tograms of the binding of env-IgG and control HMK-IgG to
each cell type; Fig. 4e demonstrates the median fluorescence
intensity derived from each histogram. Only env-IgG bound
significantly to these cells; binding of control HMK-IgG was
not significantly greater than the background level (secondary
antibody alone), indicating that binding was due to envelope
protein and not immunoglobulin determinants.
Env-IgG binding to leukocytes.
Mononuclear leukocytes are
[image:4.612.319.555.75.203.2]probable natural targets for dengue virus infection in vivo and
can be infected in vitro (18); however, there is still considerable
uncertainty about whether binding of dengue virus to these
cells is solely due to antibody-dependent binding to Fc
recep-tors or whether it can also be mediated by a specific viral
receptor (12). Binding of env-IgG and control human IgG to
blood-derived human leukocytes was examined to determine if
envelope protein-specific binding could be detected; the results
are summarized in Fig. 5. Preparations of neutrophils were
98% pure by morphologic criteria. Low-level and equivalent
binding of both env-IgG and human IgG to these cells was
observed. Preparations of mononuclear cells contained only
lymphocytes and monocytes by morphologic criteria.
Mononu-clear cells were incubated with env-IgG or control human IgG,
FIG. 3. Binding of env-IgG to Vero cells. (a) Time course. env-IgG binding to Vero cells is time dependent and saturable. env-IgG was incubated with Vero cells at a concentration of 10mg/ml at 48C for time intervals ranging from 1 to 90 min (solid bars). Flow cytometry was used to detect and quantify binding. Ab-scissa, time interval. Nil refers to cells incubated without primary reagent. Con-trol IgG refers to cells incubated with conCon-trol human IgG (10mg/ml, 90 min, 48C). Ordinate, median fluorescence intensity, derived from fluorescence histo-grams for each sample analyzed. (b) Dose response. env-IgG binding to Vero cells is dose dependent and saturable. Vero cells were incubated with concen-trations of env-IgG ranging from 1 to 40mg/ml for 60 min at 48C (solid bars). Flow cytometry was used to detect and quantify binding. Abscissa, env-IgG concentration. Nil refers to cells incubated without primary reagent. Control IgG refers to cells incubated with control human IgG (20 mg/ml, 60 min, 48C). Ordinate, median fluorescence intensity, derived from fluorescence histograms for each sample analyzed. (c) Partial-length envelope proteins. The env-IgG binding motif for Vero cells is expressed by carboxy-terminal amino acids. Vero cells were incubated with full-length or partial-length recombinant envelope proteins (10mg/ml, 60 min, 48C). Flow cytometry was used to detect and quantify binding. Abscissa, envelope protein preparation. Nil refers to cells incubated without primary reagent. Control IgG refers to cells incubated with control human IgG (10mg/ml, 60 min, 48C). Env-IgG refers to full-length envelope protein, 34/253 refers to partial-length envelope protein expressing amino acids 34 to 253, and 281/423 refers to partial-length envelope protein expressing amino acids 281 to 423. Ordinate, median fluorescence intensity, derived from
fluores-cence histograms for each sample analyzed.
on November 9, 2019 by guest
http://jvi.asm.org/
lymphocytes and monocytes were distinguished during flow
cytometric analysis by using standard forward- and side-angle
light scatter criteria (35), and binding results for lymphocyte
and monocyte subpopulations were expressed independently.
No binding of either env-IgG or control human IgG to
lym-phocytes was observed. In contrast, there was significant
bind-ing to monocytes that was equivalent for both env-IgG and
control human IgG; this pattern suggested that binding was
mediated by their common immunoglobulin determinants and
not by envelope protein elements.
In order to further explore the mechanism of envelope
pro-tein binding to monocytes, additional experiments were
per-formed with U937 cells, cells from a histiocytic
lymphoma-derived cell line that can be chemically induced to undergo
terminal monocytic differentiation and exhibits many
mono-cyte-like characteristics, including expression of
immunoglob-ulin Fc receptors (16). As with monocytes, there was low-level
but significant and equivalent binding of both env-IgG and
control human IgG to U937 cells. Binding increased as the
cells were differentiated with phorbol myristate acetate and
activated with interferon gamma, consistent with the known
effects of these treatments on increasing expression of Fc
re-ceptors. Figure 6a demonstrates the median fluorescence
in-tensity derived from the fluorescence histogram of each cell
preparation. env-IgG and control human IgG bound
signifi-cantly and equally to these cells in all treatment groups,
con-sistent with binding mediated by common immunoglobulin but
not by envelope determinants.
Additional experiments were performed to confirm that
env-IgG binding to U937 cells was mediated by env-IgG Fc
determi-nants. U937 cells were first incubated with mouse IgG1 to
saturate Fc receptors or were not treated; cells were then
incubated with env-IgG, control human IgG, or no added
ma-terial and were washed, and binding was detected by a final
incubation with a fluorochrome-labeled secondary antibody
specific for human (but not mouse) IgG. Results of this assay
(Fig. 6b) indicated that preincubation with mouse IgG resulted
in virtually complete inhibition of binding of both env-IgG and
human IgG, consistent with env-IgG binding to U937 cells
being attributable entirely to immunoglobulin determinants.
env-IgG inhibits dengue virus infection.
To assess the
func-tional role of the binding of envelope protein in the infection
of target cells, the ability of recombinant env-IgG to inhibit
FIG. 4. Binding of env-IgG to four different cell types. Cells were incubated with env-IgG or control HMK-IgG (10mg/ml, 60 min, 48C); then, flow cytometry was used to quantify binding. Panels a to d represent fluorescence histograms for each cell type. Abscissa, fluorescence intensity on a 104-U logarithmic scale;
[image:5.612.84.276.70.336.2]ordinate, number of cells in each fluorescence intensity group. Results for env-IgG are indicated by the solid line. Results for control HMK-env-IgG are indicated by the broken line. (a) Vero cells; (b) CHO cells; (c) human umbilical vein endo-thelial cells (HUVEC); (d) glial cells. (e) Summary of results obtained for each cell type. Abscissa, cell type; ordinate, median fluorescence intensity for cells incubated without primary reagent (Nil), with control HMK-IgG, or with env-IgG, all followed by incubation with secondary fluorochrome-labeled antibody.
FIG. 5. Binding of env-IgG to blood-derived leukocytes. Neutrophils (N) and mononuclear cells were purified from human peripheral blood. Cells were then incubated without primary reagent (Nil), with control human IgG, or with env-IgG; this was followed by incubation with secondary fluorochrome-labeled anti-body, and then binding was detected by flow cytometry analysis. Lymphocytes (L) and monocytes (MØ) within the mononuclear cell preparation were resolved by forward- and side-angle light scatter characteristics, and fluorescence data were expressed for each individual cell type. Abscissa, cell type; ordinate, median fluorescence intensity for each sample analyzed.
FIG. 6. Binding of env-IgG to U937 cells. (a) Effect of cell activation. Cells were treated with phorbol myristate acetate (PMA), phorbol myristate acetate and gamma interferon (PMA1IFNg), or left untreated (abscissa). Cells were then incubated without primary reagent (Nil), with control human IgG, or with env-IgG; this was followed by incubation with secondary fluorochrome-labeled antibody, and then binding was detected by flow cytometry analysis. Ordinate, median fluorescence intensity values for each sample analyzed. (b) Effect of blocking Fc receptors. U937 cells were preincubated with mouse IgG to saturate Fc receptors or left untreated (abscissa). Cells were then incubated without primary reagent (Nil), with control human IgG, or with env-IgG. Cells were washed and incubated with secondary fluorochrome-labeled antibody that binds to epitopes expressed by human IgG but not by mouse IgG. Binding was detected by flow cytometry analysis. Ordinate, median fluorescence intensity values for each sample analyzed.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.319.555.73.191.2]dengue virus infection of Vero cells was examined. Vero cells
were preincubated with env-IgG, control HMK-IgG, control
human IgG, or recombinant protein buffer or were left
un-treated; then, serial dilutions of a fresh dengue virus
prepara-tion were added. Unbound virus was removed by washing, and
the cells were observed for cytopathic effects. The ID
50titer for
untreated cells was 1.4
3
10
24. env-IgG treatment reduced the
ID
50titer to 8.9
3
10
24
, a 6.3-fold reduction that was 5
stan-dard deviations below the ID
50titer for the untreated wells. In
contrast, control human IgG had no effect on the ID
50titer
(1.6
3
10
24), and control HMK-IgG and buffer were both
associated with an increase in the ID
50titer (6.3
3
10
25
and
5.0
3
10
25, respectively).
DISCUSSION
Several factors have limited progress in the delineation of
the pathogenesis of dengue virus infection. Research has been
hampered both by the lack of facilities for clinical studies in
third-world areas where disease is prevalent and also by the
limitations of available animal models. For example, although
subhuman primates can be infected with dengue virus, they do
not develop the disease symptoms that occur in humans (20,
23). Rodents can be infected and killed by dengue virus and
are widely used to investigate immune responsiveness to
den-gue virus antigens; however, rodents are most effectively
in-fected by intracerebral injection and do not manifest major
features of the human disease (23). Although simple tissue
culture models are limited in the information they can provide
about potentially complex pathophysiological mechanisms,
such models can yield valuable insights about cellular and, by
inference, tissue tropism. Our results, derived from in vitro
studies, provide important new information about the
molec-ular mechanism by which dengue viruses bind to target cells.
Our results, demonstrating specific binding of recombinant
envelope protein to target cells, are consistent with findings of
an electron microscopy study of dengue virus interaction with
target cells which demonstrated very rapid association of
viri-ons with the cytoplasmic membranes of cells (21). The rapidity
of the observed interaction and the apparent cell surface
bind-ing are congruent with the current understandbind-ing that the
initial interaction of viruses and other microorganisms with
target cells is based on specific receptor-ligand interactions
between surface structures. Viruses utilize a wide range of cell
surface molecules as binding targets (42), and in some cases,
the restricted tissue distribution of the target molecule explains
the tropism of the virus and disease pathogenesis; a notable
example is the binding of human immunodeficiency virus
viri-ons to CD4 molecules, which are expressed by subsets of
lym-phocytes (11).
Flaviviruses, including dengue virus, have a single envelope
protein as the major surface structural molecule (33). It is
therefore likely that target cell binding motifs will be found
within the envelope protein, although none have previously
been described. In this study, a reagent consisting of the
ectodomain of dengue type 2 virus envelope protein was
gen-erated as a recombinant chimeric protein in a eukaryotic
ex-pression system and used as a probe to investigate binding to
target cells. This reagent bound to Vero cells, a target cell line
that is readily infected by dengue viruses. Binding was
satura-ble over both time and dose, which is characteristic of a specific
receptor-ligand interaction. Binding was attributable to
enve-lope protein determinants; control proteins that lacked the
envelope sequence did not bind to these and other target cells.
Studies with endoglycosidases suggested that the decoration
of the recombinant envelope protein with carbohydrates was
responsible for an increase in molecular weight of 4,000, in
agreement with an investigation of dengue virus envelope
pro-tein expressed with a baculovirus-derived vector in insect cells
(14). The consensus sites for N-linked glycosylation occur at
amino acids 67 and 153. The partial-length construct
express-ing amino acids 34 to 253, that included both of these sites, did
not bind to Vero cells, whereas the construct expressing amino
acids 281 to 423 bound at high levels. This finding indicated
that envelope protein glycosylation is not required for target
cell binding; however, these results do not exclude a role for
glycosylation in other elements of pathogenicity.
Localization of the target cell binding motif to
carboxy-terminal residues is also congruent with results of a recent
study that defined the crystal structure of the envelope protein
of tick-borne encephalitis virus (32), a flavivirus closely related
to dengue virus. This analysis demonstrated that domain III of
the envelope protein, situated at the carboxy terminus, formed
an immunoglobulin-like module. The presence of this
struc-ture, which is common to many proteins with an adhesive
function (4), provides additional support for the hypothesis
that the carboxy terminus of the envelope protein is
responsi-ble for cell attachment.
In a clinical context, evidence of endothelial cell binding
attributable to the envelope protein is of particular interest
because these cells are a likely target of dengue virus infection
in vivo. While it has been difficult to identify dengue virus
within endothelial cells in infected human tissue (34), cultured
endothelial cells are readily infected with dengue virus (2).
Moreover, the prominent vascular manifestations of dengue
virus infection, including the erythematous rash in dengue
fe-ver, the hemorrhagic rash in dengue hemorrhagic fefe-ver, and
the cardiovascular collapse in dengue hemorrhagic shock
syn-drome, all suggest tropism of dengue virus for the vasculature.
The specific binding of envelope protein to cultured human
vascular endothelial cells demonstrated in this study supports
the hypothesis that dengue virus is endotheliotropic. The
re-combinant envelope protein that was utilized as a probe of cell
surface binding could also be useful for the isolation and
iden-tification of the endothelial receptor molecule(s).
Additional evidence to support the pathophysiological role
of the envelope protein in determining tissue tropism was
provided by the cellular specificity of binding. Glial cells, which
can be infected by dengue virus but are relatively resistant to
infection (24, 28), displayed much lower levels of envelope
protein binding than the infection-permissive Vero cells,
con-sistent with the hypothesis that binding of envelope protein to
target cells has an important role in controlling infectivity.
More direct support for the hypothesis was provided by the
results of the experiment in which the impact of soluble
re-combinant envelope protein on viral infectivity was assessed;
the preincubation of Vero cells with the envelope protein
re-duced their susceptibility to infection by dengue virus.
Al-though the extent of the reduction observed, 6.3-fold, was
relatively small, it was clearly significant in view of the lack of
any inhibitory effect of the controls used and the very large
number of assay replicates. An experimental factor that could
explain the relatively small inhibitory effect observed was the
use of soluble recombinant env-IgG that was in a nonnative
conformation and was also probably partially denatured and
thus not able to compete efficiently with the native molecule,
expressed on the virion surface, for target cell binding sites.
The recombinant chimeric protein utilized for this study was
engineered to express part of the heavy-chain constant region
of human IgG1 at its carboxy terminus. Immunoglobulin
chi-meras have a number of properties that make them useful as
soluble probes. (i) The chimeric proteins can be highly purified
on November 9, 2019 by guest
http://jvi.asm.org/
and concentrated in a single step by affinity chromatography
over protein A. (ii) The protein chimeras are expressed as
polyvalent homodimers, leading to increased ligand-binding
avidity. (iii) Binding of these chimeras to targets can be readily
detected with the large range of secondary reagents available
to detect human IgG determinants. (iv) The Fc region of these
chimeras can be utilized to precipitate and isolate
ligand-re-ceptor complexes by binding to immobilized protein A, in a
procedure similar to immunoprecipitation.
The secretion of the envelope-immunoglobulin chimera into
the culture medium was expected, but no protein could be
recovered from the culture supernatant. Functional
determi-nants of the envelope protein could account for this finding.
The flavivirus membrane protein could function by preventing
intracellular binding and fusion of envelope protein (33), and
it is possible that coexpression with membrane protein would
be required for secretion to occur. We also observed that
env-IgG bound to the surface of COS cells, the cell type used
for expression, and it is likely that any secreted protein became
cell bound in this way.
In this study, a particular advantage of utilizing a chimeric
probe expressing both envelope protein and immunoglobulin
determinants was that it enabled us to compare the relative
roles of envelope protein binding to unknown cellular
recep-tors and immunoglobulin Fc region binding to cellular Fc
re-ceptors. The data indicated that binding to Vero, CHO, glial,
and endothelial cells was mediated by envelope protein
deter-minants whereas binding to U937 cells and human monocytes
was mediated exclusively by the immunoglobulin Fc
determi-nants. While the results presented do not exclude the
possibil-ity that monocytes and U937 cells express an envelope protein
receptor, none could be demonstrated. Previous reports have
indicated that dengue virus can infect monocytes and
leuko-cyte-derived cell lines in the absence of infection-enhancing
antibody, implying that viral receptors could exist on these cells
(5, 12, 26). It is possible that the flow cytometry technique we
utilized for receptor detection was insufficiently sensitive for
detection of low-abundance or low-affinity receptors or that
binding mediated by immunoglobulin Fc determinants
gener-ated a signal too high to allow the detection of relatively
lower-level envelope determinant-mediated binding.
In summary, our results demonstrated that dengue virus
envelope protein expresses motifs that bind to as yet unknown
surface structures that are expressed by a range of target cell
types, in a manner consistent with a specific
receptor-ligand-mediated interaction; binding to these cell types is
indepen-dent of immunoglobulin Fc determinants. Evidence that the
envelope protein inhibited infection of target cells suggests
that envelope protein binding is an important and limiting
determinant of infectivity. Envelope protein-target cell
inter-actions probably contribute to the tissue tropism and
patho-genesis of dengue virus and other flaviviruses.
Further work directed at identifying both the envelope
pro-tein motifs responsible for target cell binding and the target
cell receptor molecules will lead to an enhanced understanding
of the pathophysiology of flavivirus infections. The
character-ization of these epitopes could yield important molecular
tar-gets both for vaccine development and for the development of
therapeutic strategies based on the blockade of target cell
binding. Finally, investigation of the target cell receptors
uti-lized by other families of viruses has revealed that they exploit
a wide range of physiologically important cell surface
mole-cules. We speculate that the identification of flavivirus
recep-tors could lead to the discovery of novel, physiologically
im-portant cell surface molecules.
ACKNOWLEDGMENTS
R. M. Marks is supported by a grant from the Pew Scholars program
and Public Health Service grants PO1AI33189, P50AR41703,
MO1RR000420758, and P60AR20557. T. Maguire is supported by a
grant from the Health Research Council of New Zealand.
We acknowledge the generous assistance of John B. Lowe for access
to flow cytometry services, Vishva M. Dixit and Haining Shao for
providing the CD5-IgG expression plasmid, and Yvonne Coughlan for
technical assistance.
REFERENCES
1. Anderson, R., A. D. King, and B. L. Innis. 1992. Correlation of E protein binding with cell susceptibility to dengue 4 virus infection. J. Gen. Virol.
73:2155–2159.
2. Andrews, B. S., A. N. Theofilopoulos, C. J. Peters, D. J. Loskutoff, W. E.
Brandt, and F. J. Dixon.1978. Replication of dengue and Junin viruses in cultured rabbit and human endothelial cells. Infect. Immun. 20:776–781. 3. Aruffo, A., I. Stamenkovic, M. Melnick, C. B. Underhill, and B. Seed. 1990.
CD44 is the principal cell surface receptor for hyaluronate. Cell 61:1303– 1313.
4. Bork, P., L. Holm, and C. Sander. 1994. The immunoglobulin fold. Struc-tural classification sequence patterns and common core. J. Mol. Biol. 242: 309–320.
5. Brandt, W. E., J. M. McCown, F. H. Top, Jr., W. H. Bancroft, and P. K.
Russell.1979. Effect of passage history on dengue-2 virus replication in subpopulations of human leukocytes. Infect. Immun. 26:534–541. 6. Chen, P. C., and T. Hai. 1994. Expression vectors for affinity purification and
radiolabeling of proteins using Escherichia coli as host. Gene 139:73–75. 7. Chen, W.-B. 1992. Ph.D. thesis. University of Otago, Dunedin, New Zealand. 8. Chen, W.-B., and T. Maguire. 1990. Nucleotide sequence of the envelope glycoprotein gene of a dengue-2 virus isolated during an epidemic of benign dengue fever in Tonga in 1974. Nucleic Acids Res. 18:5889.
9. Chen, W.-B., X.-Y. Qu, and T. Maguire. 1992. A simple and rapid method for preparing large fragments of dengue virus cDNA from replicative-form RNA using reverse transcriptase and PCR. J. Virol. Methods 39:197–206. 10. Clark, G. G. 1996. Personal communication.
11. Dagleish, A. G., P. C. Beverley, P. R. Clapham, D. H. Crawford, M. F.
Greaves, and R. A. Weiss.1984. The CD4 (T4) antigen is an essential component of the receptor for the AIDS retrovirus. Nature (London) 312: 763–767.
12. Daughaday, C. C., W. E. Brandt, J. M. McCown, and P. K. Russell. 1981. Evidence for two mechanisms of dengue virus infection of adherent human monocytes: trypsin-sensitive virus receptors and trypsin-resistant immune complex receptors. Infect. Immun. 32:469–473.
13. Davis, B. D., R. Dulbecco, H. N. Eisen, H. S. Ginsberg, and W. B. Wood, Jr. 1968. The nature of viruses, p. 629–671. In Principles of microbiology and immunology. Harper and Row, New York.
14. Deubel, V., M. Bordier, F. Megret, M. K. Gentry, J. Schlesinger, and M.
Girard.1991. Processing, secretion, and immunoreactivity of carboxy termi-nally truncated dengue-2 virus envelope proteins expressed in insect cells by recombinant baculoviruses. Virology 180:442–447.
15. Elder, J. H., and S. Alexander. 1982. Endo-beta-N-acetylglucosaminidase F: endoglycosidase from Flavobacterium meningosepticum that cleaves both high-mannose and complex glycoproteins. Proc. Natl. Acad. Sci. USA 79: 4540–4544.
16. Grattage, L. P., I. F. C. McKenzie, and P. M. Hogarth. 1992. Effects of PMA, cytokines and dexamethasone on the expression of cell surface Fc receptors and mRNA in U937 cells. Immunol. Cell Biol. 70:97–105.
17. Halstead, S. B. 1988. Pathogenesis of dengue: challenges to molecular biol-ogy. Science 239:476–486.
18. Halstead, S. B. 1989. Antibody, macrophages, dengue virus infection, shock, and hemorrhage: a pathogenetic cascade. Rev. Infect. Dis. 11:S830–S839. 19. Halstead, S. B. 1990. Global epidemiology of dengue hemorrhagic fever.
Southeast Asian J. Trop. Med. Public Health 21:636–641.
20. Halstead, S. B., H. Shotwell, and J. Casals. 1973. Studies on the pathogen-esis of dengue infection in monkeys. I. Clinical laboratory responses to primary infection. J. Infect. Dis. 128:7–14.
21. Hase, T., P. L. Summers, and K. H. Eckels. 1989. Flavivirus entry into cultured mosquito cells and human peripheral blood monocytes. Arch. Virol.
104:129–143.
22. He, R.-T., B. L. Innis, A. Nisalak, W. Usawattanakul, S. Wang, S.
Kalay-anarooj, and R. Anderson.1995. Antibodies that block virus attachment to Vero cells are a major component of the human neutralizing antibody response against dengue virus type 2. J. Med. Virol. 45:451–461. 23. Henchal, E. A., and J. R. Putnak. 1990. The dengue viruses. Clin. Microbiol.
Rev. 3:376–396.
24. Imbert, J. L., P. Guevara, J. Ramos-Castaneda, C. Ramos, and J. Sotelo. 1994. Dengue virus infects mouse cultured neurons but not astrocytes. J. Med. Virol. 42:228–233.
25. Jaffe, E. A. (ed.). 1984. Biology of endothelial cells, p. 1–13. Martinus Nijhoff, The Hague, The Netherlands.
on November 9, 2019 by guest
http://jvi.asm.org/
26. Kurane, I., U. Kontny, J. Janus, and F. A. Ennis. 1990. Dengue-2 virus infection of human mononuclear cell lines and establishment of persistent infections. Arch. Virol. 110:91–101.
27. Laemmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680–685.
28. Lucia, H. L., and D. Kangwanpong. 1994. Identification of dengue virus-infected cells in paraffin-embedded tissue using in situ polymerase chain reaction and DNA hybridization. J. Virol. Methods 48:1–8.
29. Monath, T. P. 1994. Dengue: the risk to developed and developing countries. Proc. Natl. Acad. Sci. USA 91:2395–2400.
30. Porterfield, J. S. 1986. Antibody-dependent enhancement of viral infectivity. Adv. Virus Res. 31:335–355.
31. Reed, L. J., and H. Muench. 1938. A simple method of estimating fifty per cent end points. Am. J. Hyg. 27:493–497.
32. Rey, F. A., F. X. Heinz, C. Mandl, C. Kunz, and S. C. Harrison. 1995. The envelope glycoprotein from tick-borne encephalitis at 2 A resolution. Nature (London) 375:291–298.
33. Rice, C. M. 1996. Flaviviridae: the viruses and their replication, p. 931–959. In B. N. Fields, D. M. Knipe, P. M. Howley, et al. (ed.), Fields virology, 3rd ed. Lippincott-Raven, Philadelphia.
34. Sahaphong, S., S. Riengrojpitak, N. Bhamarapravati, and T.
Chira-chariyavej.1980. Electron microscopic study of the vascular endothelial cell in dengue hemorrhagic fever. Southeast Asian J. Trop. Med. Public Health
11:194–204.
35. Salzman, G. C., J. M. Crowell, J. C. Martin, T. T. Trujillo, A. Romero, P. F.
Mullaney, and P. M. LaBauve.1975. Cell classification by laser light scat-tering: identification and separation of unstained leukocytes. Acta Cytol.
19:374–377.
36. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. In Molecular cloning: a laboratory manual, 2nd ed., p. 16.1–16.81. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
37. Schlesinger, R. W. 1977. Dengue viruses. Springer-Verlag, New York. 38. Shao, H., L. Lou, A. Pandey, E. B. Pasquale, and V. M. Dixit. 1994. cDNA
cloning and characterization of a ligand for the Cek5 receptor protein-tyrosine kinase. J. Biol. Chem. 269:26606–26609.
39. Tarr, G. C., and A. S. Lubiniecki. 1976. Chemically induced temperature-sensitive mutants of dengue virus type 2: comparison of temperature sensi-tivity in vitro with infecsensi-tivity in suckling mice, hamsters, and rhesus monkeys. Infect. Immun. 13:688–695.
40. Tyler, K. L., and B. N. Fields. 1996. Pathogenesis of viral infections, p. 173–217. In B. N. Fields, D. M. Knipe, P. M. Howley, et al. (ed.), Fields virology, 3rd ed. Lippincott-Raven, Philadelphia.
41. Umemoto, J., V. P. Bhavanandan, and E. A. Davidson. 1977. Purification and properties of an endo-alpha-N-acetyl-D-galactosaminidase from Diplococ-cus pneumoniae. J. Biol. Chem. 252:8609–8614.
42. Wimmer, E. (ed.). 1994. Cellular receptors for animal viruses, p. 1–13. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.