Copyright © 1999, American Society for Microbiology. All Rights Reserved.
Moloney Murine Sarcoma Virus Genomic RNAs Dimerize via a
Two-Step Process: a Concentration-Dependent Kissing-Loop
Interaction Is Driven by Initial Contact between
Consecutive Guanines
HINH LY,
1DONALD P. NIERLICH,
2,3JOHN C. OLSEN,
4,5ANDANDREW H. KAPLAN
1,4,6*
Departments of Microbiology & Immunology
1and Medicine,
4Lineberger Cancer Center,
6and Cystic Fibrosis/Pulmonary
Research and Treatment Center,
5School of Medicine, The University of North Carolina at Chapel Hill, Chapel Hill,
North Carolina, and Department of Microbiology, Immunology, & Molecular Genetics
2and Molecular Biology
Institute,
3University of California Los Angeles School of Medicine, Los Angeles, California
Received 10 February 1999/Accepted 25 May 1999
Retroviruses contain two plus-strand genomic RNAs, which are stably but noncovalently joined in their 5
ⴕ
regions by a dimer linkage structure (DLS). Two models have been put forward to explain the mechanisms by
which the RNAs dimerize; each model emphasizes the role of specific molecular determinants. The kissing-loop
model implicates interactions between palindromic sequences in the DLS region. The second model proposes
that purine-rich stretches in the region form purine quartet structures. Here, we present an examination of the
in vitro dimerization of Moloney murine sarcoma virus (MuSV) RNA in the context of these two models.
Dimers were found to form spontaneously in a temperature-, time-, concentration-, and salt-dependent
manner. In contrast to earlier reports, we found that deletion of neither the palindrome nor the consensus
purine motifs (PuGGAPuA) affected the level of dimer formation at low concentrations of RNA. Rather,
different purine-rich sequences, i.e., consecutive stretches of guanines, were found to enhance both in vitro RNA
dimerization and in vivo viral replication. Biochemical evidence further suggests that these guanine-rich
(G-rich) stretches form guanine quartet structures. We also found that the palindromic sequences could
support dimerization at significantly higher RNA concentrations. In addition, the G-rich stretches were as
important as the palindromic sequence for maintaining efficient viral replication. Overall, our data support a
model that entails contributions from both of the previously proposed mechanisms of retroviral RNA
dimer-ization.
A unique feature of retroviruses is that they encapsidate a
dimer of identical plus-strand genomic RNAs. Electron
mi-croscopy and sedimentation studies indicate that the two
RNAs are held together noncovalently near their 5
⬘
ends in a
union referred to as the dimer linkage structure (DLS) (3, 21,
27, 35). This region of the genome also encodes major
regu-latory features important to the viral life cycle, including the
primer binding site, the major splice donor, the
gag
start codon,
and the
cis
-acting encapsidation (
) signal. Thus, the DLS
directly or indirectly plays roles in reverse transcription of viral
genomic RNA, viral RNA splicing specificity, translational
control, and determination of the specificity of viral RNA
encapsidation (40).
Although it has been recognized for some time that the DLS
resides near the 5
⬘
end of the genome, the mechanism of
dimerization remains obscure. Currently, two models for
dimerization have been put forward. The kissing-loop model
for DLS formation proposes that palindromic sequences (Pal)
in a hairpin loop interact through Watson-Crick base pairing.
An RNA containing the human immunodeficiency virus type 1
(HIV-1) DLS was shown to dimerize in vitro through
interac-tions of a 6-base Pal (GUGCAC) (23, 34, 38, 41, 46).
Further-more, studies with synthetic RNAs and antisense
oligonucleo-tides implicated a 16-base, self-complementary sequence of the
Moloney murine leukemia virus (MuLV) as part of a putative
DLS region (16, 17, 42, 43, 49). However, mutations
engi-neered to ablate Pal in MuLV did not affect the stability of
genomic RNAs isolated from virions (4, 7, 18, 24, 45).
Fur-thermore, the kissing-loop model cannot explain some unusual
features of the retroviral RNA dimers, e.g., their resistance to
denaturing conditions and the inability of antisense DLS RNA
to form dimers (29, 34). Finally, the formation of heterodimers
between the DLS sequences of HIV-1 and other retroviruses
that lack homology between their Pal sequences suggests a role
for other determinants of dimerization (29).
For the second model of dimerization, phylogenetic analyses
of putative retroviral DLSs revealed a unique purine-rich motif
(PuGGAPuA) that is conserved among retroviruses (29). It
has been proposed that these consensus purine motifs may
form purine-base tetrads, in part because they are stabilized by
monovalent cations such as those observed in human telomeric
DNA (53). A similar kind of structure that involves consecutive
guanine stretches has also been proposed (1, 48). Structural
probing and biochemical studies support the contention that
the HIV-1 DLS is maintained by these guanine-rich (G-rich)
sequences (1, 2, 29, 48). In addition, in vitro and in vivo studies
of rat retrotransposon VL30 and Rous sarcoma virus DLS
RNAs provide evidence that G-rich sequences can function in
dimer pairing (12, 25, 50, 51).
Here, in an attempt to synthesize these apparently disparate
findings, we characterize each of the signals that constitute the
two different models of retroviral dimerization. Our results
suggest that the G-rich and Pal sequences both play roles in
* Corresponding author. Mailing address: CB#7030, 547
Burnett-Womack, UNC-Chapel Hill, Chapel Hill, NC 27599-7030. Phone:
(919) 966-2536. Fax: (919) 966-6714. E-mail: [email protected].
7255
on November 9, 2019 by guest
http://jvi.asm.org/
pLN plasmid (32). The modification involved subcloning the retroviral sequences of pLN into a pBluescript II KS(⫹) vector to permit a higher vector yield during propagation inEscherichia coli(36). MuSV is a replication-defective virus that was derived from MuLV and in which most of thepolandenvopen reading frames have been replaced by that of the c-mosgene. The MuSV DLS sequence possesses greater than 95% homology to that of the MuLV (44). The nucleotides are numbered according to the MuSV sequence recently submitted to GenBank (accession no. AF033813) (Fig. 1A and B).
By using PCR, DNA fragments were synthesized with the MuSV DLS region of pLNBS as the template, a forward primer identical to the primer binding site at nucleotide (nt) 105 and carrying a 5⬘EcoRI site (5⬘GAATTCTGGGGGCT CGTCCGGGAT3⬘), and a reverse primer beginning at position 557 (5⬘TATTT TCAGACAAATACAGAAAC3⬘). The PCR products were then digested with
EcoRI andPstI (position 503) and cloned into the vector pGEM3Zf(⫹) (Pro-mega) behind its T7 promoter, which was used in transcribing the inserted sequences. In a similar fashion, several deletion mutations were constructed with primers that incorporated the desired deletions (Fig. 1C) (20). The sequences of all the constructions were confirmed by sequencing by the Sanger dideoxy ter-mination method with Sequenase 2.0 (USB).
Preparation of RNA samples.Prior to transcription, template DNAs (⬃2.0 g) were linearized with various restriction enzymes (PstI,BsiEI, andTthlllI) that subsequently define the 3⬘ ends of the products. The linearized DNAs were phenol-chloroform extracted and ethanol precipitated, and the pellets were re-suspended in 2.0l of water. Transcription mixtures (20l) contained 1.0g of DNA template, 0.5 mM (each) unlabeled ATP, GTP, and UTP, 50M unla-beled CTP, and 2.5M [␣-32P]CTP. The reaction was carried out with a T7
MAXIscript kit as suggested by the manufacturer (Ambion). All RNA tran-scripts starting at position 310 were the result of transcription of PCR products of pLNBS with the forward primer carrying the T7 promoter sequence and the reverse primer as above.
RNA transcripts were separated by electrophoresis on 5% polyacrylamide–8 M urea gels at 250 V for 2 h in 1⫻TBE buffer (0.09 M Tris-borate, 0.002 M EDTA). RNA samples were eluted from the gels with elution buffer as suggested by the manufacturer (Ambion). The purified RNAs were then ethanol precipi-tated, resuspended in 100l of diethylpyrocarbonate-treated water, and stored at⫺70°C. The products were quantitated by counting in a Beckman LS6500 scintillation counter.
RNA dimerization assay.For the dimerization assay, 8-l portions of⬃1.5 nM [␣-32P]RNA were heated at 95°C for 3 min and chilled on ice for 3 min, and 2l
of D5X buffer (500 mM NaCl, 250 mM Tris-HCl [pH 7]) was added. The samples were then incubated at the temperatures indicated in the text for 1 h. At the end of the incubation period, the samples were chilled and separated by electro-phoresis on 5% nondenaturing polyacrylamide gels in the presence of 1⫻TBE buffer. The gels were dried and exposed directly on a PhosphorImager screen for 18 to 20 h. The amount of dimer formation was quantitated with a 445SI PhosphorImager system and ImageQuaNT image analysis software. The fraction of RNA in dimeric form was estimated by taking the ratio of the amount of dimeric RNA to the amount of the sum of both the monomeric and dimeric species. Concentration-dependent, time-dependent, and salt-dependent mea-surements were made in a similar manner, varying the appropriate parameters. In the salt-dependent dimerization assays of f587–985, the reaction mixtures were incubated for 30 min rather than 1 h. The time-dependent dimerization comparisons of f105–503, f105–503⌬Pal, and f105–503⌬G5 were done in a man-ner similar to the standard dimerization reaction, except that f105–503⌬Pal RNA was incubated at 50°C whereas the other RNAs were incubated at 60°C.
Cation-dependent thermal dissociation.The RNA sample f792–1040 (52.5 nM) was incubated for 3 h at 60°C in dimerization buffer containing a final concentration of 100 mM NaCl and 50 mM Tris-HCl (pH 7). At the end of the incubation, the RNA was precipitated by addition of 2 volumes of ethanol and 0.1 volume of 3 M sodium acetate (pH 5.2), and after centrifugation, the pellet
colony formation as described previously (37). The deletions of the palindromic and the G-rich stretches are outlined in Fig. 1C.⌬Psi was made by deleting part of thesequence from theSpeI site (position 243) to theEcoRI site (41 nt upstream of theneoAUG site).
RESULTS
MuSV DLS dimerizes in a temperature-, concentration-,
and salt-dependent manner.
Initial experiments were designed
to evaluate the effects of reaction conditions on spontaneous
dimerization of the DLS region of MuSV RNA (Fig. 1).
Dimerization of a 398-nt fragment containing the Pal as well as
purine-rich sequences (f105–503) was optimal at 60°C and was
not supported by temperatures lower than 40°C or higher than
70°C (Fig. 2). In addition, we found that f105–503 dimer
for-mation was RNA concentration, time, and salt dependent
(data not shown).
Neither the Pal sequences nor the consensus purine motifs
(PuGGAPuA) are required for dimerization.
RNAs lacking
part or all of the self-complementary sequence were tested to
determine whether the Pal sequence is required for dimer
pairing (Fig. 2). The f105–503
⌬
Pal fragment (Fig. 1A and C),
which carries a deletion of 10 of the 16 nt of Pal, was still able
to dimerize (Fig. 2A). The optimal temperature for dimer
formation of this fragment was 50°C, 10°C lower than that of
the full-length f105–503 fragment (compare lanes 2 and 6).
Also, at optimal temperatures, f105–503
⌬
Pal dimerized only
50% as efficiently as the full-length molecule did. These
ob-servations imply that the palindromic sequence may help
sta-bilize dimeric RNA.
To examine whether the palindrome itself was sufficient for
dimer formation, an RNA fragment (f105–310) spanning the
Pal sequence was tested under conditions identical to those
used for the full-length construct. Figure 2B(ii) shows that Pal
alone was not sufficient for dimerization. In contrast, a
down-stream fragment (f310–557), lacking Pal altogether, dimerized
in a manner similar to that seen for the full-length fragment
[Fig. 2B(i)]. This region carries multiple purine-rich sequences
(Fig. 1C).
A close examination of the f310–557 sequence reveals two
stretches of the consensus purine motif (GGGAGA [Gcon1] at
positions 319 to 324 and AGGAGA [Gcon2] at positions 417
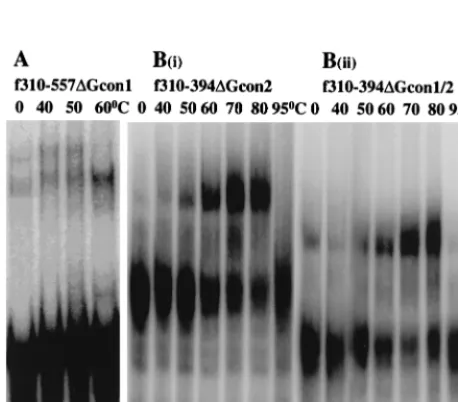
to 422) (Fig. 1C). Deletion of either one or both Gcon motifs
did not inhibit dimer formation (Fig. 3), suggesting that these
phylogenetically conserved sequences do not function in
dimerization.
At different concentrations of RNA, both guanine-rich and
Pal sequences promote dimerization.
The f310–557 sequence
also carries stretches of consecutive guanines at positions 340
on November 9, 2019 by guest
to 344 (G5) and 371 to 373 (G3) (Fig. 1C). Since similar G-rich
sequences are involved in initiating dimerization of HIV-1
DLS RNA (2, 48), the influence of these G-rich sequences was
tested with MuSV RNA. As shown in Fig. 4, the deletion of
either of these sequences virtually eliminated dimer formation.
By contrast, extending the RNA fragment carrying only Pal
(f105–310), which by itself did not dimerize [Fig. 2B(ii)], to
include these two G-rich sequences restored the ability of the
FIG. 1. MuSV RNA sequences involved in dimerization. (A) Representation of the MuSV genome. Numbering is with respect to the viral mRNA, which begins at the transcriptional initiation site and ends at the polyadenylation site (i.e., 5⬘R to 3⬘R) (GenBank accession no. AF033813). Viral long terminal repeats,gag,pol, and v-mosgenes, and restriction endonuclease cleavage sites are shown. (B) Diagram of the RNAs used in this study. Gcon1, first consensus purine motif; Gcon2, second consensus purine motif; G5 and G3, consecutive guanines; PBS, primer binding site; SD, splice donor site; AUG,gaginitiation site. (C) Sequence of the DLS region of MuSV. The locations of restriction sites of interest, the primer binding site (PBS), and the splice donor site (SD) are indicated by lines above the sequence. Pal, the consecutive guanines (G5 and G3), and the consensus purine motifs (PuGGAPuA) are shown in boldface and are noted above the sequences. The deletions of the Pal, G-rich, and first consensus purine motif sequences are underlined.on November 9, 2019 by guest
http://jvi.asm.org/
fragment to dimerize (f105–394) (Fig. 5A). No obvious
struc-tural rearrangement was predicted for f105–310 compared to
f105–394 as calculated with MFold, a program to predict RNA
secondary structures based on free-energy minimization (data
not shown) (52, 55). Moreover, when G5 or G3 was
individu-ally deleted from f105–394, dimerization was severely
dimin-ished (Fig. 5B and C). The percentage of dimeric RNA at
optimal temperature was reduced from 65% to as low as 15%.
These data suggest that the G-rich stretches play an important
role in dimer formation.
The observation that Pal is dispensable for RNA
dimeriza-tion is at odds with results obtained by others (9, 10, 16, 23, 41,
49). Since we used an assay system with 10
3-fold less viral RNA
than was used previously, we decided to examine the effects of
RNA concentration on the role played by the different
deter-minants. In these experiments, increasing amounts (0.25 to 2.0
[image:4.612.316.545.72.273.2]
g, equivalent to 0.2 to 1.5
M) of the identical but unlabeled
FIG. 2. Pal is dispensable in dimer formation but is required to stabilize the dimeric structure. (A) An RNA fragment lacking 10 of the 16 nt of Pal (f105– 503⌬Pal) dimerizes less efficiently than the full-length fragment (f105–503). (B) The RNA fragment containing Pal alone (f105–310) does not form dimers [B(ii)], whereas [B(i)] one lacking Pal but carrying the downstream sequence (f310–557) forms dimers efficiently [B(i)].M, monomeric RNA;D, dimeric RNA;
M1, monomeric f310–557 RNA;M2, monomeric f105–310 RNA;, an RNA marker of 330 nt was transcribed from the-globin gene (Ambion).
[image:4.612.57.290.72.268.2]FIG. 3. The consensus purine motifs (Gcon) are dispensable in MuSV dimer-ization. Temperature-dependent dimerization of RNA fragments carrying dele-tions of either one [(A) and B(i)] or both [B(ii)] motifs was carried out as described in Materials and Methods, except that f310–557⌬Gcon1 was incubated for 30 min rather than 60 min.
FIG. 4. The consecutive guanine stretches (G-rich) are essential in MuSV dimerization. Dimerization of an RNA fragments carrying a deletion of the five consecutive guanines (f310–557⌬G5) (A) and the three consecutive guanines (f310–557⌬G3) (B) is severely inhibited. L, the RNA marker of 300, 400, and 500 nt was transcribed from the Century-Plus DNA templates (Ambion);M, mono-meric RNA;D, dimeric RNA.
FIG. 5. The G-rich sequences downstream from Pal rescue the dimerization defect of f105–310, and the deletions of the G-rich sequences greatly diminish dimer formation. (A) An extended molecule (f105–394) including Pal and down-stream purine-rich sequences dimerizes efficiently. (B and C) Dimers of f310– 557⌬G5 (B) and of f310–557⌬G3 (C) which lack the G-rich sequences are greatly reduced. All samples were assayed under the same conditions and separated on the same gel. L, the RNA marker of 300, 400, and 500 nt was transcribed from Century-Plus DNA templates (Ambion);M, monomeric RNA;D, dimeric RNA.
on November 9, 2019 by guest
[image:4.612.314.545.469.659.2] [image:4.612.59.288.478.679.2]RNA were added to the standard assay mixture containing a
fixed amount of [
␣
-
32P]RNA (1.5 nM). Figure 6A shows that
although the fragment carrying Pal alone, f105–310, was not
able to form dimers at 1.5 nM (lane 2), it did dimerize at higher
concentrations. A similar experiment was carried out with the
full-length f105–503 fragment (Fig. 6B). The result mirrors the
concentration dependence of the shorter fragment, except that
a small amount of dimer was observed even at the lowest RNA
concentration (Fig. 6B, lane 2). These data emphasize that
dimerization of Pal is concentration dependent across the
range of concentrations tested. Also, the observation that
f105–310 RNA at 1.5
M dimerized 10% less efficiently than
did the full-length f105–503 suggests that the dimeric structure
of this short fragment of RNA is less stable than that of the
full-length molecule (Fig. 6A and B, lanes 6).
Dimers of the guanine-rich sequences are preferentially
sta-bilized by monovalent cations with the smallest radius.
DNA
or RNA containing runs of consecutive guanines may form
four-stranded structures (G tetrads) by the formation of
Hoogsteen base pairs. These structures are markedly stabilized
by specific monovalent cations, apparently by reducing the
electrostatic repulsion of the bases across the central tube of
the molecules (6, 15, 19, 22, 54). Depending on their primary
sequence, dimers containing quartet structures dissociate at
various temperatures and also are stabilized to various degrees
in different monovalent cations. These considerations
prompted us to define the melting temperature of the G-rich
f310–557 RNA in the presence of different monovalent cations.
This RNA shows a preference for the cation with the smallest
radius (Li
⫹) (Fig. 7). The melting temperature of f310–557 in
the presence of Li
⫹was higher than 80°C, about 10°C higher
than observed in the presence of cations with larger radii. This
preference for a small cation suggests that DLS RNA forms an
ordered structure which contains G tetrads.
Viral replication is reduced by mutations in the Pal and
G-rich sequences.
Finally, we examined the relative roles
played by the Pal and G-rich elements of MuSV DLS in the
context of a single round of virus replication. To do this,
se-quences containing various mutations were placed into the 5
⬘
leader region of pLNBS, an MuLV-MuSV-derived retroviral
vector containing a
neo
gene. The effect of the mutations on
vector production following transfection into the GP
⫹
E86
ret-roviral packaging cell line was determined. The results
ob-tained from three independent experiments for each of the
mutants indicate the replication ability of each of the viral
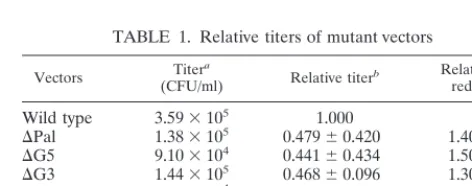
constructs (Fig. 1C) relative to the wild type (Table 1). While
deletion of either the Pal sequence (
⌬
Pal) or the G-rich
stretches (
⌬
G5 or
⌬
G3) alone slightly reduced the titer of the
virus, deletion of both signals (
⌬
Pal/
⌬
G5 or
⌬
Pal/
⌬
G3)
pro-duced up to a sevenfold-reduction in titer. Also, there was an
additive effect of viral reduction when both signals were
de-leted. These data demonstrate that both the Pal and G-rich
sequences play important roles in the MuSV viral life cycle.
DISCUSSION
[image:5.612.317.546.72.262.2]Several lines of observations have been made that support a
two-stage model of MuSV-RNA dimerization. First, G-rich
sequences, and not the phylogenetically conserved purine-rich
motifs, drive the initial dimer formation, perhaps via the
for-mation of an ordered quartet structure. Second, kissing-loop
FIG. 6. f105–310 dimerizes in a concentration-dependent manner. [␣-32P]RNA (1.5 nM) was incubated with increasing amounts of unlabeled RNA
at 60°C for 1 h before being separated on a nondenaturing 5% polyacrylamide gel as described in Materials and Methods. (A) f105–310 fragment. For this experiment, a 0.25-g addition in a 10-l reaction mixture represents 3.8⫻104
[image:5.612.57.288.72.254.2]nM RNA. (B) f105–503 fragment. All samples were assayed under the same conditions and separated on the same gel.M, monomeric RNA; D, dimeric RNA.
FIG. 7. The f310–557 dimeric structure is preferentially stabilized by lithium cations. Samples of f310–557 RNA were allowed to dimerize, and the dimers were precipitated, washed with ethanol, and resuspended in water as described in Materials and Methods. Aliquots were then incubated at 40 to 95°C in the presence of the indicated salts.M, monomeric RNA;D, dimeric RNA; L, the RNA marker of 300, 400, and 500 nt was transcribed from the Century-Plus DNA templates (Ambion).
TABLE 1. Relative titers of mutant vectors
Vectors (CFU/ml)Titera Relative titerb Relative fold reduction
Wild type
3.59
⫻
10
51.000
⌬Pal
1.38
⫻
10
50.479
⫾
0.420
1.40–2.90
⌬G5
9.10
⫻
10
40.441
⫾
0.434
1.50–3.50
⌬G3
1.44
⫻
10
50.468
⫾
0.096
1.30–2.30
⌬Pal/⌬G5
6.60
⫻
10
40.198
⫾
0.104
4.70–6.80
⌬Pal/⌬G3
8.10
⫻
10
40.266
⫾
0.256
2.80–7.20
⌬Psi
1.60
⫻
10
30.005
⫾
0.005
210–266
aTiters of the vectors from one representative experiment.
bTiters normalized to the wild type. Data were taken from three independent assays. Relative titers are given as mean⫾standard deviation.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.314.550.610.703.2]formation (Fig. 7). Also, Torrent et al. found that the VL30
dimeric RNA was stabilized in the presence of LiCl (50), and
similar results were obtained with RSV RNA (25). In contrast,
the proposed HIV-1 quartet structure is stabilized by KCl and
is thought to involve the phylogenetically conserved
purine-rich motifs (PuGGAPuA) (1, 48). These differences in the
stabilizing cation, Li
⫹versus K
⫹, may reflect structural
differ-ences: deletion of the MuSV consensus purine-rich motifs did
not affect dimerization (Fig. 3). Similarly, the deletion of all
five consensus purine motifs in the HIV-2 DLS did not alter
the level of dimerization (5).
An important finding of this study is the demonstration that
the influence of the Pal and the G-rich regions on dimerization
varies with concentration. While RNA fragments carrying the
G-rich regions effectively dimerized at low, possibly more
phys-iologically relevant, concentrations, RNA fragments carrying
the Pal sequence alone paired at only very high concentrations
[Fig. 2B(ii) and 6A]. The observation of MuSV Pal sequence
dimerization at high concentrations corroborates the features
of MuLV Pal sequence dimerization observed by others (42,
43). In addition, it appears that the kissing loop serves as an
important factor in determining dimer stability (Fig. 2A). Our
findings are also consistent with the observation that
unchar-acterized sequences downstream from the kissing loop in
MuLV play a role in dimer stability (49).
Viral replication assays with a vector system carrying the
MuSV
region show that removal of either the Pal or the
G-rich sequence affects viral replication. Of note, deletions of
both signals (Pal and G-rich sequences) reduced viral
produc-tion to a much greater extent than did deleproduc-tion of either
se-quence alone (Table 1). These data corroborate recent
obser-vations indicating a dispensable but nonetheless significant
role of the Pal sequence in several other viral systems. It was
previously noted that Pal mutations affect steps other than
dimerization in the viral life cycle (40). For instance, mutations
in the palindrome of HIV-1 decreased viral infectivity (8, 24,
26, 39) and slowed viral replication (4, 18, 39). Most
charac-teristically, the palindromic mutations greatly affected RNA
encapsidation (4, 7, 8, 24, 30, 31, 39) and the synthesis of
proviral DNA (39). However, no effect of these mutations on
the synthesis of viral proteins (4) or the genomic RNA dimers
isolated from the virions (7, 24, 45) was observed. Similarly,
Mougel et al. found that a deletion spanning the Pal sequence
in an MuLV vector moderately reduced viral RNA
encapsida-tion and reverse transcripencapsida-tion of the genomic RNA (33).
Re-cently, Doria-Rose and Vogt observed that the Pal sequence is
preferred but not required to maintain stable viral replication
in RSV (11).
strategies may be used by other viral systems to induce stable
dimer formation (13, 14, 47).
Our viral infectivity data provide, for the first time, direct
evidence that in addition to Pal, the guanine stretches play
important functional roles in dimerization and/or other steps
of the viral life cycle. Although we have identified the
impor-tance of both molecular determinants in efficient MuSV
rep-lication, the mechanisms through which the mutations exert
their effect remain unknown. We are currently examining in
more detail the precise biological and molecular roles of these
elements.
ACKNOWLEDGMENTS
This work was supported by NIH grant K11 AI01107 to A.H.K. and
a seed grant from the UCLA AIDS Institute to D.P.N. H.L. was
supported in part by the UCLA Presidential Research Fellowship, the
College Honors Naumburg Research Scholarship, and the AAP
Drown Foundation Research Scholarship.
We thank John Sechelski, Nan N. Lee, Amy T. A. Tran, Liza Kim,
Yanli Yang, and James Matsunaga for their help. We also thank Steve
Pettit for his critical reading of the manuscript.
REFERENCES
1.Awang, G., and D. Sen.1993. Mode of dimerization of HIV-1 genomic RNA. Biochemistry32:11453–11457.
2.Baudin, F., R. Marquet, C. Isel, J.-L. Darlix, B. Ehresmann, and C. Ehres-mann.1993. Functional sites in the 5⬘region of human immunodeficiency virus type 1 RNA from defined structural domains. J. Mol. Biol.229:382– 397.
3.Bender, W., Y.-H. Chien, S. Chattopadhyay, P. K. Vogt, M. B. Gardner, and N. Davidson.1978. High-molecular-weight RNAs of AKR, NZB, and wild mouse viruses and avian reticuloendotheliosis virus all have similar dimer structures. J. Virol.25:888–896.
4.Berkhout, B., and J. L. van Wamel.1996. Role of the DIS hairpin in replication of human immunodeficiency virus type 1. J. Virol.70:6723–6732. 5.Berkhout, B., B. B. Oude Essink, and I. Schneveld.1993. In vitro dimeriza-tion of HIV-2 leader RNA in the absence of PuGGAPuA motifs. FASEB J.
7:181–187.
6.Cheong, C., and P. B. Moore.1992. Solution structure of an unusually stable RNA tetraplex containing G- and U-quartet structures. Biochemistry31:
8406–8414.
7.Clever, J. L., M. L. Wong, and T. G. Parslow.1996. Requirements for kissing-loop-mediated dimerization of human immunodeficiency virus RNA. J. Virol.70:5902–5908.
8.Clever, J. L., and T. G. Parslow.1997. Mutant human immunodeficiency virus type 1 genomes with defects in RNA dimerization or encapsidation. J. Virol.71:3407–3414.
9.Darlix, J.-L.1990.Ciselements andtrans-acting factors involved in dimer formation of murine leukemia virus RNA. J. Virol.64:774–783.
10. Darlix, J.-L., C. Gabus, M.-T. Nugeyre, F. Clavel, and F. Barre-Sinoussi.
1990.ciselements andtrans-acting factors involved in the RNA dimerization of the human immunodeficiency virus HIV-1. J. Mol. Biol.216:689–699. 11. Doria-Rose N., and V. M. Vogt.1998. In vivo selection of Rous sarcoma virus
mutants with randomized sequences in the packaging signal. J. Virol.72:
8073–8082.
on November 9, 2019 by guest
12.Feng, Y. X., W. Fu, A. J. Winter, J. G. Levin, and A. Rein.1995. Multiple regions of Harvey sarcoma virus RNA can dimerize in vitro. J. Virol.69:
2486–2490.
13.Fu, W., and A. Rein.1993. Maturation of dimeric viral RNA of Moloney murine leukemia virus. J. Virol.67:5443–5449.
14. Fu, W., R. J. Gorelick, and A. Rein.1994. Characterization of human im-munodeficiency virus type 1 dimeric RNA from wild-type and protease-defective virions. J. Virol.68:5013–5018.
15. Gellert, M., M. N. Lipsett, and D. R. Davies.1962. Helix formation by guanylic acid. Proc. Natl. Acad. Sci. USA48:2013–2018.
16. Girard, P. M., B. Bonnet-Mathoniere, D. Muriaux, and J. Paoletti.1995. A short autocomplementary sequence in the 5⬘leader region is responsible for dimerization of MoMuLV genomic RNA. Biochemistry34:9785–9794. 17. Girard, P. M., H. de Rocquigny, B. P. Roques, and J. Paoletti.1996. A model
of psi dimerization: destabilization of the C278-G303 stem-loop by the nu-cleocapsid protein (NCp10) of MoMuLV. Biochemistry35:8705–8714. 18. Haddrick, M., A. L. Lear, A. J. Cann, and S. Heaphy.1996. Evidence that a
kissing loop structure facilitates genomic RNA dimerization in HIV-1. J. Mol. Biol.259:58–68.
19. Hud, N. V., F. W. Smith, F. A. Anet, and J. Feigon.1996. The selectivity for K⫹versus Na⫹in DNA quadruplexes is dominated by relative free energies
of hydration: a thermodynamic analysis by 1H NMR. Biochemistry
35:15383–15390.
20.Kunkel, T. A., K. Bebenek, and J. McClary.1991. Efficient site-directed mutagenesis using uracil-containing DNA. Methods Enzymol.204:125–139. 21. Kung, H. J., S. Hu, W. Bender, J. M. Bailey, N. Davidson, M. O. Nicolson, and R. M. McAllister.1976. RD-114, baboon, and wolly monkey viral RNAs compared in size and structure. Cell7:609–620.
22. Laughlan, G., A. I. Murchie, D. G. Norman, M. H. Moore, P. C. Moody, D. M. Lilley, and B. Luisi.1994. The high-resolution crystal structure of a parallel-stranded guanine tetraplex. Science265:520–524.
23. Laughrea, M., and L. Jette.1994. A 19-nucleotide sequence upstream of the 5⬘major splice donor is part of the dimerization domain of human immu-nodeficiency virus 1 genomic RNA. Biochemistry33:13464–13474. 24. Laughrea, M., L. Jette, J. Mak, L. Kleiman, C. Liang, and M. A. Wainberg.
1997. Mutations in the kissing-loop hairpin of human immunodeficiency virus type 1 reduce viral infectivity as well as genomic RNA packaging and dimerization. J. Virol.71:3397–3406.
25. Lear, A. L., M. Haddrick, and S. Heaphy.1995. A study of the dimerization of Rous sarcoma virus RNAin vitroandin vivo. Virology212:47–57. 26. Louis, D. C., D. Gotte, E. Sanders-Buell, D. W. Ritchey, M. O. Salminen,
J. K. Carr, and F. E. McCutchan.1998. Infectious molecular clones with the nonhomologous dimer initiation sequences found in different subtypes of human immunodeficiency virus type 1 can recombine and initiate a spread-ing infection in vitro. J. Virol.72:3991–3998.
27. Maisel, J., W. Bender, S. Hu, P. H. Duesberg, and N. Davidson.1978. Structure of 50 to 70S RNA from Moloney sarcoma viruses. J. Virol.25:
384–394.
28. Markowitz, D., S. Goff, and A. Bank.1988. A safe packaging cell line for gene transfer: separating viral genes on two different plasmids. J. Virol.62:1120– 1124.
29. Marquet, R., F. Baudin, C. Gabus, J.-L. Darlix, M. Mougel, C. Ehresmann, and B. Ehresmann.1991. Dimerization of human immunodeficiency virus (type 1) RNA: stimulation by cations and possible mechanism. Nucleic Acids Res.91:2349–2357.
30. McBride, M. S., and A. T. Panganiban.1996. The human immunodeficiency virus type 1 encapsidation site is a multipartite RNA element composed of functional hairpin structures. J. Virol.70:2963–2973.
31. McBride, M. S., M. D. Schwartz, and A. T. Panganiban.1997. Efficient encapsidation of human immunodeficiency virus type 1 vectors and further characterization ofciselements required for encapsidation. J. Virol.71:
4544–4554.
32. Adam, M. A., and D. Miller.1988. Identification of a singnal in a murine retrovirus that is sufficient for packagaing of nonretroviral RNA into virions. J. Virol.62:3802–3806.
33. Mougel, M., Y. Zhang, and E. Barklis.1996.cis-active structural motifs involved in specific encapsidation of Moloney murine leukemia virus RNA. J. Virol.70:5043–5050.
34. Muriaux, D., P.-M. Girard, B. Bonnet-Mathoniere, and J. Paoletti.1995. Dimerization of HIV-1Lai RNA at low ionic strength: an autocomplemen-tary sequence in the 5⬘leader region is evidenced by an antisense oligonu-cleotide. J. Biol. Chem.270:8209–8216.
35. Murti, K. G., M. Bondurant, and A. Tereba.1981. Secondary structural features in the 70S RNAs of Moloney murine leukemia and Rous sarcoma viruses as observed by electron microscopy. J. Virol.37:411–419. 36. Olsen, J. C.Unpublished data.
37. Olsen, J. C., and J. Sechelski.1995. Use of sodium butyrate to enhance production of retroviral vectors expressing CFTR cDNA. Hum. Gene Ther.
6:1195–1202.
38. Paillart, J. C., R. Marquet, E. Skripkin, B. Ehresmann, and C. Ehresmann.
1994. Mutational analysis of the bipartite dimer linkage structure of human immunodeficiency virus type 1 genomic RNA. J. Biol. Chem.269:27486– 27493.
39. Paillart, J. C., L. Berthoux, M. Ottmann, J. L. Darlix, R. Marquet, B. Ehresmann, and C. Ehresmann.1996. A dual role of the putative RNA dimerization initiation site of human immunodeficiency virus type 1 in genomic RNA packaging and proviral DNA synthesis. J. Virol.70:8348– 8354.
40. Paillart, J. C., R. Marquet, E. Skripkin, C. Ehresmann, and B. Ehresmann.
1996. Dimerization of retroviral genomic RNAs: structural and functional implications. Biochimie78:639–653.
41. Paillart, J. C., E. Skripkin, B. Ehresmann, C. Ehresmann, and R. Marquet.
1996. A loop-loop “kissing” complex is the essential part of the dimer linkage of genomic HIV-1 RNA. Proc. Natl. Acad. Sci. USA93:5572–5577. 42. Paoletti, J., M. Mougel, N. Tounekti, P. M. Girard, C. Ehresmann, and B.
Ehresmann.1993. Spontaneous dimerization of retroviral MoMuLV RNA. Biochimie75:681–686.
43. Prats, A.-C., C. Roy, P. Wan, M. Erard, V. Housset, C. Gabus, C. Paoletti, and J. L. Darlix.1990.cis-elements andtrans-acting factors involved in dimer formation of murine leukemia virus RNA. J. Virol.64:774–783.
44. Reddy, P. E., M. J. Smith, and S. A. Aaroson.1981. Complete nucleotide sequence and organization of the Moloney murine sarcoma virus genome. Science214:445–450.
45. Sakuragi, J.-I., and A. T. Panganiban.1997. Human immunodeficiency virus type 1 RNA outside the primary encapsidation and dimer linkage region affects RNA dimer stability in vivo. J. Virol.71:3250–3254.
46. Skripkin, E., J. C. Paillart, R. Marquet, B. Ehresmann, and C. Ehresmann.
1994. Identification of the primary site of the human immunodeficiency virus type 1 RNA dimerization in vitro. Proc. Natl. Acad. Sci. USA91:4945–4949. 47. Stoltzfus, C. M., and P. N. Snyder.1975. Structure of B77 sarcoma virus
RNA: stabilization of RNA after packaging. J. Virol.16:1161–1170. 48. Sundquist, W. I., and S. Heaphy.1993. Evidence for interstrand quadruplex
formation in the dimerization of human immunodeficiency virus type 1 genomic RNA. Proc. Natl. Acad. Sci. USA90:3393–3397.
49. Tapia, M. D., V. Metzler, M. Mougel, B. Ehresmann, and C. Ehresmann.
1998. Dimerization of MoMuLV genomic RNA: redefinition of the role of the plaindromic stem-loop H1 (278–303) and new roles for stem-loops H2 (310–352) and H3 (355–374). Biochemistry37:6077–6085.
50. Torrent, C., T. Bordet, and J. L. Darlix.1994. Analytical study of rat retro-transposon VL30 RNA dimerization in vitro and packaging in murine leu-kemia virus. J. Mol. Biol.240:434–444.
51. Torrent, C., C. Gabus, and J. L. Darlix.1994. A small and efficient dimer-ization/packaging signal of rat VL30 RNA and its use in murine leukemia virus-VL30-derived vectors for gene transfer. J. Virol.68:661–667. 52. Tounekti, N., M. Mougel, C. Roy, R. Marquet, J.-L. Darlix, J. Paoletti, B.
Ehresmann, and C. Ehresmann.1992. Effect of dimerization on the confor-mation of the encapsidation Psi domain of Moloney murine leukemia virus RNA. J. Mol. Biol.223:205–220.
53. Wang, Y., and D. J. Patel.1993. Solution structure of the human telomeric repeat d[AG3(T2AG3)3] G-tetraplex. Structure1:263–282.
54. Williamson, J. R., M. K. Raghuraman, and T. R. Cech.1989. Monovalent cation-induced structure of telomeric DNA: the G-quartet model. Cell59:
871–880.
55. Zuker, M.1989. On finding all suboptimal foldings of an RNA molecule. Science244:48–52.
on November 9, 2019 by guest
http://jvi.asm.org/