0022-538X/07/$08.00
⫹
0
doi:10.1128/JVI.00502-07
Copyright © 2007, American Society for Microbiology. All Rights Reserved.
DNA Immunization with Plasmids Encoding Fusion and Nucleocapsid
Proteins of Bovine Respiratory Syncytial Virus Induces a Strong
Cell-Mediated Immunity and Protects Calves against Challenge
䌤
Mathieu Boxus,
1* Maryle
`ne Tignon,
1Stefan Roels,
1Jean-Franc¸ois Toussaint,
1Karl Walravens,
1Marie-Ange Benoit,
2Philippe Coppe,
2Jean-Jacques Letesson,
2Carine Letellier,
1and Pierre Kerkhofs
1Veterinary and Agrochemical Research Centre (VAR), 1180 Brussel, Belgium,
1and Faculte´s Universitaires Notre-Dame de la Paix,
Unite´ de Recherche en Biologie Mole´culaire, 5000 Namur, Belgium
2Received 9 March 2007/Accepted 13 April 2007
Respiratory syncytial viruses (RSV) are one of the most important respiratory pathogens of humans and
cattle, and there is currently no safe and effective vaccine prophylaxis. In this study, we designed two
codon-optimized plasmids encoding the bovine RSV fusion (F) and nucleocapsid (N) proteins and assessed
their immunogenicity in young calves. Two administrations of both plasmids elicited low antibody levels but
primed a strong cell-mediated immunity characterized by lymphoproliferative response and gamma interferon
production in vitro and in vivo. Interestingly, this strong cellular response drastically reduced viral replication,
clinical signs, and pulmonary lesions after a highly virulent challenge. Moreover, calves that were further
vaccinated with a killed-virus vaccine developed high levels of neutralizing antibody and were fully protected
following challenge. These results indicate that DNA vaccination could be a promising alternative to the
classical vaccines against RSV in cattle and could therefore open perspectives for vaccinating young infants.
Bovine respiratory syncytial virus (BRSV) and human
respi-ratory syncytial virus (HRSV) belong to the
Pneumovirus
genus
of the
Paramyxoviridae
family (52). These two negative
single-stranded RNA viruses share common genomic, antigenic,
epidemiological, and pathological characteristics (62).
BRSV and HRSV are major causative agents of severe
res-piratory tract diseases in cattle and infants worldwide,
respec-tively (20, 31, 62). Both BRSV infection and HRSV infection
can remain asymptomatic or cause severe respiratory tract
diseases leading sometimes to death (62). Seventy percent of
calves exhibit a positive serological response against BRSV at
the age of 12 months, and mortality can reach up to 20% in
some outbreaks (31, 61). From figures available in
industrial-ized countries, the number of annual HRSV infections
world-wide can be estimated around 64 million and mortality could
be as high as 160,000 (20).
For these reasons, efficient vaccines against HRSV and
BRSV are needed. However, their development has been
ham-pered since the dramatic vaccine failure in the 1960s. In fact,
vaccination with formalin-inactivated, alum-adjuvanted virus
predisposed children to a far more serious, and sometimes
fatal, form of pathology in the case of natural infection (29).
Subsequently, it was found that a similarly inactivated BRSV
vaccine could induce strikingly similar immunopathology (47).
Further studies in mice and cattle suggested that exacerbation
of disease resulted from a polarized type 2 T-helper cell
re-sponse characterized by increased production of interleukin-4
(IL-4) and IL-5 cytokines, high levels of immunoglobulin G1
(IgG1) and IgE, and a lack of BRSV-specific CD8
⫹T cells,
resulting in enhanced pulmonary eosinophilia (10, 13, 18, 25,
27, 63, 67).
Recently, DNA vaccines have emerged as a promising
alter-native to the modified live and killed-virus (KV) vaccines.
Direct immunization with naked DNA results in the
produc-tion of immunogenic antigens in the host cell which can readily
go through processing and presentation via both class II and
class I pathways and engender long-lasting humoral and
cell-mediated immunity. Furthermore, DNA vaccines mimic live
attenuated virus in their ability to induce both humoral and
cellular responses but are considered to be safer and to offer
several technical advantages (21, 22). Finally, since the
immu-nizing protein is not present in the vaccine preparation,
plas-mid DNA is not susceptible to direct inactivation by maternal
antibodies (44).
So far, DNA vaccination against HRSV has been mainly
investigated in mice or cotton rats (6, 8, 32, 33, 58). These
studies demonstrated that plasmids encoding the HRSV fusion
(F) or attachment (G) proteins primed both humoral and
cell-mediated immunity and protected against HRSV infection
without significantly enhancing pulmonary pathology following
challenge. Despite these promising results, very few studies
confirmed the ability of DNA vaccines to protect against RSV
infection in a natural host. DNA immunization with plasmid
encoding BRSV F or G protein primed the humoral response
of young calves, reduced virus excretion, and partially
pro-tected them after experimental infection (48, 53). Similarly,
DNA immunization against BRSV F and nucleocapsid (N)
proteins was shown to be safe, immunogenic, and partially
protective in infant rhesus monkeys (64). Even if these reports
highlight the potential of DNA vaccination, it seems that the
efficacy of this strategy has to be improved in terms of the
quality and intensity of the response induced.
Codon optimization and protein boost following DNA
vac-* Corresponding author. Present address: Biologie Cellulaire et
Mole
´culaire, Faculte
´ des Sciences Agronomiques, 5030 Gembloux,
Belgium. Phone: 32 (81) 62.21.57. Fax: 32 (81) 61.38.88. E-mail:
䌤
Published ahead of print on 25 April 2007.
6879
on November 8, 2019 by guest
http://jvi.asm.org/
cination are two commonly used methods that improve the
efficacy of DNA immunization (21, 66). In this report, we
designed codon-optimized plasmids encoding BRSV F and N
proteins and assessed their immunogenicity in young calves.
MATERIALS AND METHODS
Plasmids.Full-length nonoptimized F and N genes of BRSV were amplified by reverse transcription-PCR (RT-PCR) from viral mRNA extracted from cell culture supernatant infected with the BRSV strain RB94 as previously described (7). Synthetic constructs carrying BRSV F (Fsyn) and N (Nsyn) genes were manually designed by reverse translation of the amino acid sequence of the RB94 strain in order to avoid negativelycis-acting sequences and restore bovine codon usage. The prediction of splicing sites was performed at the NetGene2 server (http://www.cbs.dtu.dk/services/NetGene2/) (9). The presence of othercis-acting elements like the polyadenylation sequence (AAUAAA) (65) or the Shaw-Kamen instability motif (AUUUA) (49) was detected manually. The CUSP software available at EMBOSS (http://emboss.sourceforge.net/) was used to re-store bovine codon frequency. The codon adaptive index (CAI), which is a measurement of the relative adaptativeness of the codon usage of a gene com-pared with the codon usage of highly expressed genes, was calculated using the CAI software available at EMBOSS with a user-defined set of codon relative adaptiveness values based on codon usage in a compendium of bovine genes (Ebovsp.cut) (ideal value of 1.0). The resulting synthetic sequences for F and N genes were submitted to GenBank under accession no. AJ971803 and AJ971804, respectively, and were provided by Geneart GmbH (Germany). Native and optimized genes encoding F (pF and pFsyn, respectively) and N (pN and pNsyn, respectively) proteins were cloned into the HindIII-BamHI window of the pCDNA3.1 plasmid (Invitrogen) with an ATG initiation codon presented in an optimized Kozak sequence context (30). All plasmids were restriction verified, sequenced, and purified with Endofree plasmid Giga kits (QIAGEN) according to the manufacturer’s instructions.
Antibodies. Monoclonal antibodies specific for BRSV F (AK13A2 and AK8D1) were produced by immunizing BALB/c mice with the human Long strain and have been previously described (35). Mab19 was kindly provided by G. Taylor (Institute for Animal Health, Compton, United Kingdom) and has been described by Arbiza et al. (4). Monoclonal antibodies specific for BRSV N (AL5G1 and MoAb␣-N) were produced by immunizing BALB/c mice with the RB94 strain by a protocol similar to that described by Matheise et al. (35). The specificity of the AL5G1 and MoAb␣-N antibodies was tested by immunoblot-ting and immunoprecipitation procedures (data not shown). The K71 bovine polyclonal anti-BRSV antibody was produced in gnotobiotic calves after re-peated inoculation with the RB94 BRSV strain and was provided by G. Welle-mans (Veterinary and Agrochemical Research Centre, Brussels, Belgium).
Cell culture.Vero and COS-7 cells were grown in minimal Eagle’s medium (Gibco) supplemented with 10% fetal calf serum, 2.5 mM glutamine, and anti-biotics.
Viruses. BRSV RB94 was grown on Vero cell monolayers. The challenge inoculum consisted of lung lavage fluid collected 6 days after intratracheal inoculation of a calf with the BRSV strain VRS 3761 (kindly provided by Gilles Meyer, Ecole Nationale Ve´te´rinaire de Toulouse, France). This strain was suc-cessively passaged on three young calves by a protocol similar to that described by Tjørnehøj et al. (56). This inoculum contained 103.5 50% tissue culture
infective doses (TCID50)/ml and was free of bovine viral diarrhea virus, bovine
herpesvirus 1, bovine parainfluenza virus 3, bovine coronavirus, bovine adeno-virus 5, endotoxins, bacteria, and mycoplasmas.
In vitro expression of plasmids in COS-7 cells.COS-7 cells were grown in 24-well plates at 8⫻104
cells per well and were transiently transfected with 400 ng of recombinant or empty pcDNA3.1 plasmids using Lipofectamine as
de-scribed by the manufacturer (Invitrogen). Twenty-four hours after transfection, monolayers were fixed in 4% paraformaldehyde and the expression of the F and N proteins was checked by an indirect immunostaining procedure using bovine polyclonal bovine anti-BRSV serum (K71) and monoclonal antibodies specific for the F protein (AK13A2, AK8D1, and mAb19) or for the N protein (AL5G1 and MoAb␣-N).
DNA immunization of mice.Four groups of five 6- to 8-week-old, BALB/c mice were housed in animal facilities. Mice were immunized twice by intramus-cular injections with 50g of plasmid construct (pCDNA3, pF, pN, pFsyn, or pNsyn). Three weeks after the second immunization, sera were collected in each group and pooled, and BRSV-specific IgG levels were measured by enzyme-linked immunosorbent assay (ELISA) performed as previously described (47). Briefly, RB94 cell cultures from classic culture were sedimented by an overnight centrifugation at 90,000⫻g. The pellet was suspended in phosphate-buffered saline (PBS) containing 0.1%N-octyl-D-glucopyranoside (Sigma) for perme-abilization of virus envelopes. A predetermined optimal dilution of this prepa-ration was adsorbed on polyvinyl microplates (Maxisorp; Nunc) overnight at 4°C. Plates were washed with PBS–0.1% Tween 20 and blocked with 100l/well of hydrolyzed casein in PBS–0.1% Tween 20 for 2 h at room temperature. Serial fivefold dilutions of murine sera (50l/well) were added to wells, and the plates were incubated for 1 h. Goat anti-mouse IgG antibody conjugated to horseradish peroxidase (Dako) was used as secondary antibody at a dilution of 1:2,000 in PBS containing hydrolyzed casein. Antibody titer was defined as the logarithm of the inverse of the dilution giving an optical density (OD) value two times over that of the corresponding preimmune serum.
Immunization of calves.Seventeen 1-month-old calves, deprived of colostrum, were housed in separate isolation rooms. The calves were free of bovine viral diarrhea virus, bovine herpesvirus 1, BRSV, and bovine parainfluenza virus 3. As shown in Table 1, three groups of four calves were immunized intramuscularly and intradermally twice at a 3-week intervals (day 0 and day 21) with either 250
g of each of the pFsyn and pNsyn plasmids (F⫹N/KV and F⫹N) or 500g of the empty pCDNA3.1 vector (pCDNA3/KV). The other calves received saline. Three weeks after the second DNA vaccination (day 42), the calves of the F⫹N/KV and pCDNA3/KV groups were vaccinated once by intramuscular im-munization with 5 ml of a commercially available, Quil-A-adjuvanted vaccine, containing 105.5
TCID50of inactivated BRSV (Bovipast; Intervet). The other
groups received saline solution. Three weeks after the third vaccination (day 63), all of the calves were challenged with BRSV strain VRS 3761 by intratracheal injection of 15 ml of inoculum. One calf (control) was not infected.
All calves were bled weekly for examination of humoral and cellular responses. Bronchoalveolar lavage (BAL) fluids were collected from days 63 to 77 by instillation and aspiration of 50 ml of PBS directly into the lungs using a 120-cm-length PVC dual-flow tube (Vygon, Belgium). On days 68, 70, and 82, one or two calves of each group were euthanized. Immediately after euthanasia, lungs were excised and macroscopic lesions were photographed and recorded on a standard lung diagram to score the extent of pneumonic consolidation as previ-ously described (56). A score of 0 was given to a lung without macroscopic lesion. A score of between 1 and 5 was given according to the extent of the lung lesions: 1 represents 1 to 5%, 2 represents 5 to 15%, 3 represents 15 to 30%, 4 represents 30 to 50%, and 5 represents⬎50%. Finally, six pieces of tissue from the right cranial, middle, and caudal lobes (two pieces in each lobe) were excised.
Evaluation of clinical signs.Clinical signs were recorded daily after challenge. Respiratory rate was measured via a stethoscope for a full minute. Rectal tem-perature, the presence of nasal discharge, spontaneous or induced cough, lung sounds, depression, and anorexia were also noted. Examination of calves was done by qualified people who were not aware of group assignments. Clinical scores were then assigned to each calf as previously described (23).
[image:2.585.42.546.81.161.2]Humoral immune response of calves.The ELISA anti-BRSV kit (CER, Bel-gium) was used to determine BRSV-specific antibody titers. IgG, IgG1, or IgG2
TABLE 1. Study design
Group (no. of calves)
Vaccination(s) Challenge
(day 63)
No. of calves euthanized
Day 0 Day 21 Day 42 Day 67 Day 69 Day 82
Mock (4)
Saline
Saline
Saline
BRSV
1
1
2
F
⫹
N/KV (4)
pFsyn
⫹
pNsyn
pFsyn
⫹
pNsyn
KV
BRSV
1
1
2
F
⫹
N (4)
pFsyn
⫹
pNsyn
pFsyn
⫹
pNsyn
Saline
BRSV
1
1
2
pCDNA3/KV (4)
pCDNA3.1
pCDNA3.1
KV
BRSV
1
1
2
Control (1)
Saline
Saline
Saline
Saline
0
1
0
on November 8, 2019 by guest
http://jvi.asm.org/
was detected by the following horseradish peroxidase-conjugated antibodies: a rabbit anti-bovine IgG antibody (Sigma), an IgG1-specific monoclonal antibody 1C8 (kindly provided by J.-J. Letesson, University of Namur, Belgium), or a sheep anti-bovine IgG2 (Serotec), respectively. All sera were tested at a 100-fold dilution, and the corrected OD (COD) was calculated for each serum by sub-traction of the OD of the serum on control antigen from the OD of the same serum on BRSV antigen. The COD was then expressed as a percentage of the COD of a positive reference serum (K71).
Neutralizing antibody titers were determined in a virus seroneutralization assay as previously described (47). Briefly, twofold-diluted sera were incubated with 100 TCID50of RB94 virus for 1 h at 37°C and added on Vero cells grown
in 96-well culture microplates. Each dilution was tested in four wells. After 4 days of incubation, monolayers were fixed in 4% paraformaldehyde and viral expres-sion on the monolayer was revealed by an immunoperoxidase assay with a bovine anti-BRSV serum (K71). Virus neutralization titers were expressed as the recip-rocal of the highest dilution of serum that completely prevented viral expression.
Lymphocyte proliferation assay.Heparinized blood samples were 10-fold di-luted in RPMI 1640 medium (Gibco) supplemented with 2 mM glutamine, 5⫻ 10⫺5M-2-mercaptoethanol, 100 U/ml of gentamicin, and 2.5g/ml ampho-tericin B (Fungizone). Two hundred microliters of diluted blood samples was added to wells of U-bottom microplates (Nunc). After 6 days of incubation with 20l of BRSV antigen (sonicated and heat-inactivated culture supernatant of Vero cells infected with RB94 virus, containing 105TCID
50/ml) or 20l of
control antigen (sonicated and heat-inactivated culture supernatant of mock-infected Vero cells), 0.8Ci of methyl [3H]thymidine in 25l RPMI 1640
medium was added to each well. After 18 h of incubation, cells were harvested and radioactivity incorporated into the DNA was measured by liquid scintillation counting (Betaplate; Pharmacia). The results were expressed as stimulation indexes (SI) corresponding to the number of cpm obtained with the BRSV antigen divided by the number of cpm obtained with the control preparation. The lymphoproliferation was considered significant when the SI was equal or superior to 2.2. All tests were set up in sextuplicate cultures.
IFN-␥assay.Gamma interferon (IFN-␥) production was measured after a 24-h in vitro stimulation of heparinized blood with 20l of the above-described BRSV or control antigens. The amounts of IFN-␥in the plasma were quantified by ELISA with the bovine IFN-␥EASIA kit (Biosource Europe). The results were expressed as the SI corresponding to the OD obtained with the BRSV antigen divided by the OD obtained with the control preparation. IFN-␥ pro-duction was considered significant when the SI was equal or superior to 1.8.
Hpt assay.The haptoglobin (Hpt) concentration was determined by measuring hemoglobin binding capacity in serum (50). Briefly, 10l of serum, blank, or standard was incubated at 25°C with 90l of methemoglobin for 10 min. Guaiacol (1.5 ml; Sigma) was added immediately followed by 0.5 ml of hydrogen peroxide. After 8 min incubation at 25°C, the OD at 470 nm was read. The Hpt concentration
in the sample was calculated as follows: [(sample absorbance⫺blank absorbance)/ (standard absorbance⫺blank absorbance)]⫻standard concentration (g/liter).
IF.For immunofluorescence (IF), frozen lung samples from the right cranial, middle, and caudal lobes were sectioned at 5m, put on slides, and fixed in acetone for 15 min at 4°C. Slides were incubated with a biotinylated bovine anti-BRSV serum (K71) for 1 h at 37°C. After washing, fluorescein isothiocya-nate (FITC)-conjugated streptavidin (Sigma) was added, the mixture was incu-bated for 1 h at 37°C, and the slides were observed on a fluorescence microscope. Slides were then examined without any information on the group assignments.
Histopathology.Samples of the macroscopic lung lesions were taken, forma-linized, embedded in paraffin wax, sectioned at 5m, and stained with hema-toxylin and eosin or using May-Gru¨nwald-Giemsa stains. Slides were then ex-amined without any information on the group assignments.
RNA extraction and cDNA synthesis.RNA extraction and cDNA synthesis were performed as previously described (7). RNA was extracted from 200l of lung homogenate or BAL fluids with an RNeasy RNA purification kit (QIAGEN) as recommended by the manufacturer. Purified RNA was dissolved in 20l of RNase-free water. Five microliters of RNA solution was used as a template for cDNA synthesis in the presence of 1l of 200 U of Moloney murine leukemia virus reverse transcriptase (200 U) (Invitrogen), 4l of 5⫻First Strand buffer (Invitrogen), 2l of 0.1 M dithiothreitol, 2l of 1⫻hexamer (Roche), 1
l of 10 mM deoxynucleoside triphosphates (dNTPs; Roche), and 5l of RNase-free water. After a 50-min incubation at 37°C, the reverse transcriptase was inactivated at 70°C for 10 min.
Real-time PCR.BRSV and bovine-actin sequences were amplified by real-time PCR as previously described (7). Each PCR was performed on 1l of cDNA mixed with 12.5l of Master Mix PlatinumTaqpolymerase (Invitrogen), 0.5 l of ROX dye (6-carboxyl-X-rhodamine; Invitrogen), 100 nM of each primer, 200 nM of probe, and RNase-free water for a final volume of 25l. Amplifications were performed as follows: 2 min at 50°C, 10 min at 95°C, and 45 cycles at 95°C for 15 s and 59°C for 1 min. The detection limit of the assay was 102
RNA copies. The standard curves demonstrated a linear range from 103
to 108copies.
For IFN-␥and IL-4, real-time PCRs were performed with Taqman Master Mix kit (Applied Biosystem) in a final volume of 25l containing 5l of cDNA, 300 nM of each primer, and 900 nM of the probes described in Table 2. Ampli-fications were performed as follows: 10 min at 95°C and 40 cycles at 95°C for 15 s and 60°C for 1 min. In each assay, control RNAs were used to construct standard curves. The detection limits for IFN-␥and IL-4 were 103RNA copies, and
standard curves demonstrated a linear range up to 108
copies (J.-F. Toussaint, unpublished data).
Statistics.The data collected were statistically analyzed using Graphpad Prism software. Two-way analysis of variance was used to compare the evolution of IgG titers, lymphoproliferation, and IFN-␥SI. Peak clinical signs and viral load values were compared by one-way analysis of variance and Tukey’s test with a family error rate of 0.05. Logarithmic transformation was applied to fulfill the condi-tions of variances in homogeneity and normality when necessary.
RESULTS
[image:3.585.41.285.87.185.2]Codon optimization of the BRSV F and N genes.
Codon
optimization of the BRSV F and N genes was performed to
restore bovine codon usage and remove potential negatively
cis
-acting elements such as splicing sites, polyadenylation signals, and
Shaw-Kamen instability motifs. As a result of codon optimization,
the GC contents of the F and N genes were increased from 35%
and 39% to 47% and 51%, respectively, and the CAI was
im-proved from 0.551 to 0.710 for the fusion gene and from 0.559 to
0.736 for the nucleocapsid gene (Table 3).
TABLE 2. Sequences of primers and probes used to detect IFN-
␥
and IL-4 by real-time RT-PCR
Target and primer or probe Sequencea
Bovine IFN-␥
Forward primer ...ATGGATATCATCAAGCAAGACATGTT Reverse primer ...TCATCCACCGGAATTTGAATC Probe...FAM-TCCTCCAGTTTCTCAGAGCTGCC
ATTCA-TAMRA Bovine IL-4
Forward primer ...TTCTGCAGGGTTGGAATTGAG Reverse primer ...TCATTCACAGAACAGGTCTTGCTT Probe...FAM-GTTCAAGCACGTGTGGCTCCTG
TAGATAC-TAMRA
a
FAM, 6-carboxyfluorescein; TAMRA, 6-carboxytetramethylrhodamine.
TABLE 3. Effects of codon optimization
Gene No. of instability sites native/optimizeda
CAI for native/ optimized
No. (%) of native nucleotides altered/total
% GC content native/optimized
Fusion
6 P, 3 SK, 4 S/0
0.551/0.710
240/1,725 (14)
35/47
Nucleocapsid
1 SK, 18 S/0
0.559/0.736
150/1,176 (13)
39/51
aP, polyadenylation signal; SK, Shaw-Kamen instability motif; S, splicing site.
on November 8, 2019 by guest
http://jvi.asm.org/
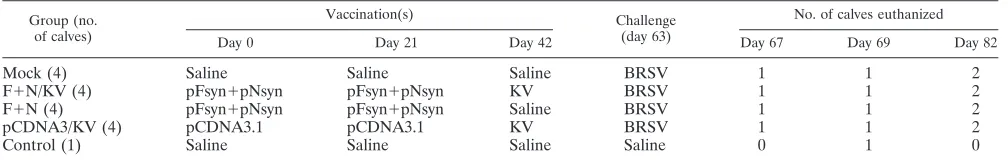
[image:3.585.38.541.660.716.2]In vitro protein expression of the pFsyn and pNsyn
plas-mids and immunogenicity in mice.
The impact of codon
opti-mization on protein expression was investigated by transfection
of COS-7 cells with equal amounts of native or optimized
plasmids encoding BRSV F and N proteins. Twenty-four hours
after transfection, protein expression was revealed by a
poly-clonal bovine anti-BRSV serum (K71) (Fig. 1). Expression of F
or N proteins was detected in cells that received native or
optimized plasmids, but no signal was observed in cells
trans-fected with the empty pcDNA3.1 plasmid. Importantly, the
results showed that expression levels were greater in cells
transfected with codon-optimized plasmids. Similar results
were obtained when monoclonal antibodies specific for
differ-ent epitopes of the fusion protein (AK13A2, AK8D1, and
mAb19) or the nucleocapsid protein (AL5G1 and MoAb
␣
-N)
were used instead of the polyclonal K71 serum (data not
shown).
The immunogenicity of the plasmids was then investigated in
mice. Five groups of four mice twice were given 50
g of
plasmid at a 3-week interval. Three weeks after the second
immunization, sera were pooled and BRSV-specific antibodies
were measured by ELISA. No antibody response was detected
after injection of mice with either the empty pCDNA3 plasmid
or nonoptimized vectors, but pFsyn and pNsyn immunizations
elicited IgG titers of 3.01 and 1.86, respectively.
Overall, these results showed that the codon optimization of
the BRSV F and N genes increased both their in vitro
expres-sion as well as their immunogenicity in mice.
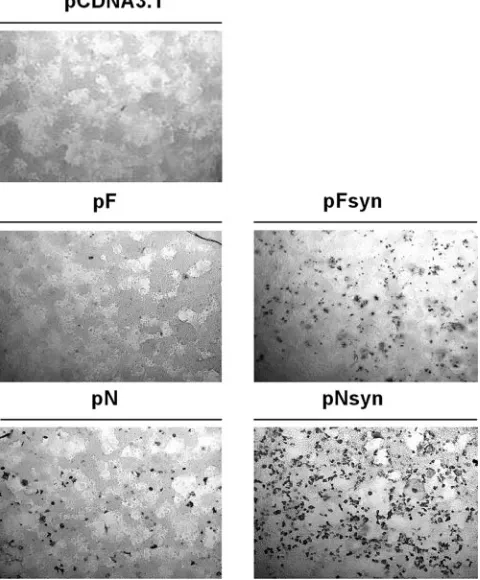
BRSV-specific immune response of calves following
vacci-nations and challenge.
The global immune response against
BRSV induced by coadministration of pFsyn and pNsyn
plas-mids was evaluated weekly by measuring levels of
BRSV-spe-cific IgG and lymphoproliferative response after antigenic
stimulation of whole blood cells. Immunization with pFysn and
pNsyn plasmids resulted in seroconversion 5 weeks after the
second vaccination (day 56), as shown by stable antibody levels
in F
⫹
N calves (Fig. 2A). KV induced the production of
BRSV-specific antibodies 2 weeks after injection in the pCDNA3/KV
groups and raised antibody levels in the F
⫹
N/KV group
com-pared to the F
⫹
N calves (
P
⬍
0.01). Ten days after challenge
(day 73), seroconversion of mock-vaccinated calves was
ob-served as well as enhanced amounts of IgG in the other groups,
especially in the pCDNA3/KV calves.
[image:4.585.324.516.70.395.2]DNA priming induced a lymphoproliferative response 5
weeks after the second vaccination (day 56) in F
⫹
N and
FIG. 1. In vitro expression of pFsyn and pNsyn plasmids in COS-7
cells. COS-7 cells in 24-well plates were transfected with 400 ng of the
plasmids described. Twenty-four hours after transfection, expression of
the BRSV F and N proteins was revealed in an immunoperoxidase
assay with a polyclonal anti-BRSV antibody (K71).
FIG. 2. BRSV-specific immune response following vaccination and
challenge. Calves were vaccinated twice with DNA (days 0 and 21)
followed by protein boost (day 42) and challenge (day 63) at 3-week
intervals. (A) IgG levels were measured by ELISA, and COD values
were transformed to percentages of a standard positive serum. (B) In
vitro lymphocyte proliferation was evaluated by measuring [
3H]thymi-dine incorporation after antigenic stimulation of whole blood cells with
control or BRSV antigen, and values were expressed as an SI. Bars
represent means (
⫹
standard deviations) in each group on days 0, 21,
42, 56, 63, 69, and 73.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.44.284.74.364.2]F
⫹
N/KV calves, as shown by higher SI compared to the
mock-vaccinated and pCDNA3/KV groups (
P
⬍
0.05) (Fig. 2B). The
KV boost was not able to stimulate lymphoproliferation in the
pCDNA3/KV group even if it induced slightly higher SI values
in F
⫹
N/KV calves compared to the F
⫹
N calves (
P
⬎
0.05).
The challenge did not enhance the SI values measured in F
⫹
N
and F
⫹
N/KV groups but it induced a lymphoproliferative
re-sponse in pCDNA3/KV and mock-vaccinated calves on day 73.
Ig isotype profile and neutralizing antibody titers.
BRSV-specific humoral immunity primed by DNA immunization and
KV boost was further investigated by analyzing the IgG isotype
profile and neutralizing antibody levels on days 63 and 69. On
these days, F
⫹
N calves had low but significant BRSV-specific
IgG1 (Fig. 3A) and IgG2 (Fig. 3B) compared to
mock-vacci-nated calves (
P
⬍
0.05). Nevertheless, these antibodies did not
possess a neutralizing activity (Fig. 3C). KV boost resulted in
significantly higher levels of IgG1 (
P
⬍
0.01), IgG2 (
P
⬍
0.01),
and neutralizing antibodies (
P
⬍
0.05) in F
⫹
N/KV and
pCDNA3/KV animals compared to F
⫹
N and mock-vaccinated
calves. Interestingly, on days 63 and 69, F
⫹
N/KV calves had
significantly lower IgG1 levels (
P
⬍
0.05) and similar amounts
of
IgG2
but
larger
neutralizing
antibody
titers
than
pCDNA3/KV calves (
P
⬍
0.05). SN/IgG
totaland IgG2/IgG1
ratios were then calculated for each calf in the F
⫹
N/KV and
pCDNA3/KV groups. These ratios could not be determined in
the two other groups because the titers before and after
chal-lenge were too weak. As shown in Fig. 3D, animals that
re-ceived the F
⫹
N priming had higher IgG
2/IgG
1and SN/IgG
totalratios than pCDNA3/KV calves (
P
⬍
0.05).
Cytokine production ex vivo and in vivo.
The effect of DNA
priming on the type of immune response induced was
exam-ined by quantifying cytokine production in vitro and in vivo.
Secretion of IFN-
␥
was measured after antigenic stimulation of
whole blood cells at challenge (day 63) and 6 days after
chal-lenge (day 69). As shown in Fig. 4A, DNA-primed calves
(F
⫹
N and F
⫹
N/KV groups) displayed significant IFN-
␥
pro-duction on the day of the challenge and this response was
slightly improved on day 69. Similarly to what we observed in
lymphoproliferation assay, KV boost did not enhance SI
mea-sured in F
⫹
N/KV calves nor did it stimulate IFN-
␥
production
in the pCDNA3/KV group.
[image:5.585.113.472.68.350.2]The amounts of IFN-
␥
and IL-4 mRNAs produced in the
right caudal lung lobes of calves euthanized four (day 67) and
six days (day 69) following challenge were measured by
real-time RT-PCR. Levels of IFN-
␥
mRNA measured in the lungs
of calves that received pFsyn and pNsyn priming (F
⫹
N and
F
⫹
N/KV) were up to 100-fold higher than those of the other
calves (Fig. 4B, left panel), confirming the results observed
after in vitro stimulation of whole blood cells. Analysis of IL-4
mRNA levels did not reveal such differences between groups
as there was no obvious IL-4 induction in the lung (Fig. 4B,
middle panel). Finally, ratios between IFN-
␥
and IL-4 were
calculated as a measure of Th bias for each calf (Fig. 4B, right
panel). These ratios were higher in calves that received the
FIG. 3. Immunoglobulin isotype profile and neutralizing antibody titers at challenge (day 63) and 6 days after challenge (day 69). IgG1 (A) and
IgG2 (B) levels were measured by isotype-discriminating ELISA, and COD values were expressed as percentages of a standard positive serum.
Neutralizing antibody titers (C) were determined by neutralization assay and expressed as the base 2 log of the maximum positive dilution. Bars
represent means
⫹
standard deviations measured in each group on days 63 and 69. Individual IgG2/IgG1 and SN/IgG
totalratios (D) were calculated
in the F
⫹
N/KV and pCDNA/KV groups on days 63 and 69.
on November 8, 2019 by guest
http://jvi.asm.org/
pFsyn and pNsyn priming compared to mock-vaccinated and
pCDNA3/KV calves, reflecting an increase in Th1 response.
Clinical signs.
After experimental infection on day 63, a
clinical score was calculated for each calf as previously
de-scribed (23). Whereas the mock-vaccinated calves had an
im-portant increase of rectal temperature and respiratory rate two
days after infection (day 65), calves from either F
⫹
N,
F
⫹
N/KV or pCDNA3/KV groups didn’t display any clinical
signs earlier than 4 to 5 days after infection (Fig. 5). In all
groups, peak clinical scores were recorded between days 68
and 70. Mock-vaccinated calves had the most severe
respira-tory disease compared to pCDNA3/KV (
P
⬍
0.01), F
⫹
N (
P
⬍
0.001) and F
⫹
N/KV (
P
⬍
0.001) animals. DNA-primed calves
displayed reduced clinical signs compared to pCDNA3/KV
group (
P
⬍
0.05), and there was no significant difference
be-tween F
⫹
N and F
⫹
N/KV groups.
Postmortem examination.
On days 67, 69, and 82, gross
pneumonic lesions were recorded and scored as previously
described (56). On day 67, only limited lesions were observed
in cranioventral lung parts of F
⫹
N, pCDNA3/KV, and
mock-vaccinated calves (Table 4). On days 69 and 82, extended
consolidated areas were seen, covering up to 50% of the lung
surface
(score
of
5)
in
the
mock-vaccinated
group.
pCDNA3/KV and F
⫹
N calves displayed reduced lesions
com-pared to mock-vaccinated calves, but consolidation was
ob-served in cranioventral and caudal regions of the lung.
[image:6.585.43.283.63.406.2]Inter-FIG. 4. Cytokine production in vitro and in vivo before and after
challenge. (A) IFN-
␥
production was measured by ELISA after
anti-genic stimulation of whole blood cells with control or BRSV antigen,
and values were expressed as SI. Bars represent means
⫹
standard
deviations measured in each group on days 63 and 69. (B) Levels of
IFN-
␥
(left panel) and IL-4 (middle panel) transcripts were quantified
by real-time RT-PCR in lung homogenates of calves euthanized on
days 67 and 69, using

-actin (

-act) as an internal reference, and
ratios between IFN-
␥
and IL-4 mRNAs were calculated for each calf
(right panel).
FIG. 5. Clinical signs following challenge. Rectal temperature,
respiratory rhythm, presence of nasal discharge, lung sounds, and
anorexia were recorded daily after challenge, and clinical scores were
calculated as previously described (56). Data represent means (
⫹
standard
deviations) in each group from day 63 (challenge) to day 77.
TABLE 4. Lesion score, peak Hpt concentration, and BRSV
detection following challenge
Group and day euthanized
Lesion scorea
Peak Hpt concn (mg/ml)
Viral
loadb,c IF detectionc,d
Mock
67
1
0
4.4
⫾
0.1
cr, ca, m
69
3
0
4.2
⫾
0.2
cr, ca, m
82
5–3
1.80–0.68
⫺
ND
F
⫹
N/KV
67
0
0
1.6
⫾
0.2
⫺
69
1
0.05
0.5
⫾
0.3
⫺
82
0–0
0.28–0
⫺
ND
F
⫹
N
67
1
0
2.2
⫾
0.2
cr
69
2
0.01
1.8
⫾
0.3
⫺
82
1–2
0.14–0.79
⫺
ND
pCDNA3/KV
67
1
0
3.0
⫾
0.6
cr, m
69
2
0
3.3
⫾
0.2
cr, m
82
4–2
1.81–0.9
⫺
ND
Control
69
0
0
⫺
⫺
aAs described by Tjornehoj et al. (56).
bMean (⫾standard deviation) viral load expressed as the log of BRSV
mRNAs per 106b-actin mRNAs detected in the right cranial, middle, and caudal
lung lobes.
c⫺, not detected.
dLung lobes in which viral expression was detected by IF. cr, cranial; m,
middle; ca, caudal; ND, not done.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.324.513.67.232.2]estingly, only one F
⫹
N/KV calf had one spot of consolidated
lung tissue in the cranial lobe. The assessment of
macroscop-ical lesions was confirmed by measuring the concentration of
an acute-phase-response protein, Hpt. Serum Hpt was
de-tected in all groups from day 68 and peaked around days 70 to
72. The highest Hpt concentrations were measured in
pCDNA3/KV and mock-vaccinated calves, and there was a
good correlation between the magnitude of the peak Hpt
con-centration and the extent of lung lesions. Overall, the observed
clinical signs were severe. Finally, viral load and expression of
the virus in cranial, middle, and caudal lung lobes were
as-sessed by real-time RT-PCR and immunofluorescence (IF),
respectively. As shown in Table 4, real time RT-PCR revealed
the presence of BRSV in all infected calves but the viral load
measured in calves that received the F
⫹
N priming was lower
than those in to mock-vaccinated and pCDNA3/KV calves.
Viral expression was detected by IF in all four lobes of
mock-vaccinated calves, in two lobes (cranial and caudal) of
pCDNA3/KV calves, and on the cranial lobe of the F
⫹
N calf
killed at day 69. Samples collected from the control calf that
was mock infected did not display gross lesions and were free
of BRSV.
Histopathological lesions.
Examination of pulmonary
histo-pathology of the control calves revealed the classical signs of
BRSV infection. Four days after infection (day 67), all sections
showed local alveolar emphysema, local lobular thickening of
the alveolar septa with syncytium formation, and localized
peribronchiolar cuffing (Fig. 6A). On day 69, pCDNA3/KV
and mock-vaccinated calves had severe bronchopneumonia
(Fig. 6B) characterized by the presence of eosinophils in the
peribronchiolar infiltrate (Fig. 6C). Eosinophils were observed
between the hyperplastic epithelium, mucosa, and submucosa.
Their presence was focal: sometimes solitary (e.g., epithelium)
and sometimes in small groups. Sections of F
⫹
N/KV calf
dis-played moderate alveolar emphysema, scattered thickening of
the alveolar septa, and syncytial formation (Fig. 6D). In the
F
⫹
N calf there was prominent inflammatory reaction with
infiltration of lymphocytes and neutrophils in the septa, near
bronchi and bronchioli, and scattered sites of consolidation
(Fig. 6E). On day 82, a prominent fibrovascular reaction,
prob-ably due to repair, was present on lung sections of
pCDNA3/KV and mock-vaccinated calves, and to a lesser
ex-tent on sections of F
⫹
N calves (Fig. 6F). Thus, the only calves
that did not display histological lesions were those from the
F
⫹
N/KV group.
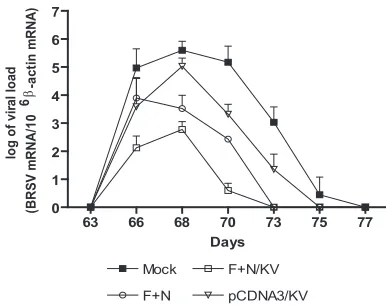
Viral replication.
The real-time RT-PCR was used to
deter-mine the viral loads in BAL fluids. As shown in Fig. 7, the
presence of BRSV was not detected before infection. Virus
load peaked around day 67 in all calves and decreased around
day 70, except in mock-vaccinated calves in which the decrease
was observed at day 72. As expected, peak viral load was
significantly higher in pCDNA3/KV and mock-vaccinated
groups compared to F
⫹
N and F
⫹
N/KV groups (
P
⬍
0.001).
Similar observations were made from the analysis of viral load
in lungs (Table 4). At day 73, F
⫹
N and F
⫹
N/KV groups were
free of BRSV, whereas viral RNA was detected up to day 75
and day 77 in pCDNA3/KV and mock-vaccinated groups,
re-spectively.
DISCUSSION
DNA vaccines are promising alternatives to the classical
modified live virus or KV vaccines. In fact, DNA immunization
often primes a Th1 immune response and provides large
num-FIG. 6. Histopathological features on lung section after challenge. (A, B, and F) Lung sections of mock-vaccinated calves on days 67 (A), 69
(B), and 82 (F). (C) Eosinophil infiltration (arrow) in pneumonic lung of a pCDNA3/KV calf on day 69. (D and E) General aspects of the F
⫹
N/KV
(D) and F
⫹
N (E) calf lung sections on day 69.
on November 8, 2019 by guest
http://jvi.asm.org/
bers of memory B and T cells. Furthermore, DNA
immuniza-tion might avoid the inhibitory effect of maternal antibodies
(for reviews, see references 21, 22, and 44). Efficacy of DNA
immunization against RSV F and G proteins has well been
described in rodents (6, 8, 32, 33, 58). However, immunization
of large animals like cattle or monkeys offered only partial
protection and require more innovative approaches, like codon
optimization and protein boost following DNA vaccination
(48, 53, 64).
In this report, we designed and constructed codon-optimized
plasmids encoding BRSV F and N proteins and evaluated the
immunogenicity of these plasmids in calves. Construction of
genes was performed in order to increase the overall GC content,
avoid negatively
cis
-acting sequence motifs, and restore codon
adaptation to frequently used bovine codons. We have shown that
codon optimization of the BRSV F and N genes led to an increase
in antigen levels expressed in mammalian cells and improved
antibody response in mice. These results are consistent with
sim-ilar codon optimization studies reported for genes encoded by
different pathogenic organisms (1, 37–39, 43).
Vaccination of young calves with a DNA plasmid containing
the BRSV F gene or recombinant vaccinia virus expressing the
F or N proteins was shown to stimulate both humoral and
cell-mediated immunity (16, 53, 54). More precisely, in these
studies, the F gene construct rather primed humoral immunity
while immunization against N rather promoted cellular
immu-nity. Consistent with these reports, two immunizations with
pNsyn elicited lower IgG levels than pFsyn but resulted in
in vitro lymphoproliferation and IFN-
␥
production, whereas
pFsyn did not (M. Tignon, unpublished results). Thus,
coim-munization against F and N proteins should stimulate both
humoral and cell mediated immunity and could be an
inter-esting vaccine approach. To answer this question, young calves
were vaccinated with both codon-optimized pFsyn and pNsyn
plasmids. Moreover, in an attempt to increase the immune
response against the virus, we boosted the DNA priming with
a heterologous boost consisting of a BRSV KV vaccine.
Re-cently, a similar strategy was shown to be efficient in the
con-text of the bovine herpesvirus 1 (57). Here, two administrations
of both pFsyn and pNsyn plasmids stimulated the two arms of
the immune system, as shown by antibody production, in vitro
lymphoproliferation, and IFN-
␥
production. Furthermore, the
use of the KV vaccine considerably increased the levels of
BRSV-specific antibody. Overall, these observations confirm
the potential of coimmunization against F and N proteins as
well as the efficacy of an alternative protein boost following
DNA immunization in large animals.
DNA priming with pFsyn and pNsyn elicited lower levels of
antibody than the KV vaccine and failed to induce the
pres-ence of a detectable neutralizing humoral response.
Neverthe-less, we have shown that pFsyn/pNsyn priming influenced the
antibody response induced by KV vaccination. Indeed, pFsyn/
pNsyn-primed calves had lower levels of IgG, especially IgG1,
but higher neutralizing antibody titers after KV vaccination
than nonprimed calves. As the presence of RSV-neutralizing
antibodies is much more important than the absolute antibody
levels in the serum (59, 60), our results suggest that pFsyn/
pNsyn priming, even if it elicits low BRSV-specific IgG titers,
improves the humoral immune response induced by the KV
boost.
The role of cell-mediated immunity is more complicated and
is a critical parameter in the outcome of RSV infection (for a
review, see reference 40). In rodents and calves, T cells are
essential for virus clearance but also appear to mediate
immu-nopathology. The CD8 cells seem to play an important role in
protection against the disease, possibly through RSV-specific
cytotoxic activity, but a strong cytotoxic T lymphocyte response
has been also implicated in the pathological outcome of
vac-cination (11, 12, 17, 25, 45, 51, 55). The role of CD4 Th cells is
more uncertain and complicated, but the imbalance of Th1 and
Th2 cytokines is likely to play a role in the pathogenesis of
RSV disease.
[image:8.585.64.257.67.220.2]DNA vaccination often promotes a Th1-biased immune
re-sponse that is characterized by high levels of antigen-specific
lymphoproliferative response and the production of large
amounts of IFN-
␥
(5). After two inoculations of the pFsyn and
pNsyn constructs, both IFN-
␥
production and
lymphoprolif-erative response were detected. In contrast to what we
ob-served for the humoral response, KV boost did not enhance
the IFN-
␥
production in calves that were DNA primed and
showed poor efficacy for the induction of both IFN-
␥
produc-tion and lymphoproliferative response in nonprimed calves
before and after challenge. In a previous report, we
demon-strated that this KV vaccine did not stimulate detectable or
strong cell-mediated immunity even after two inoculations
(28). To approach the problem of IL-4 production, we
per-formed real-time RT-PCR to quantify the amounts of IFN-
␥
and IL-4 mRNAs in the lungs of calves during the first week of
the infection. IFN-
␥
mRNA levels were increased in calves
primed with pFsyn and pNsyn plasmids, whereas there was no
clear difference in IL-4 transcripts between groups, confirming
what we observed after whole-blood-cell sensitization. Thus,
DNA priming using pFsyn and pNsyn plasmids seems to
mote a Th1-biased immune response characterized by the
pro-duction of IFN-
␥
both in vitro and in vivo. IFN-
␥
has been
shown to promote the production of IgG2, resulting in a higher
IgG2/IgG1 ratio (15). Here, in DNA-primed calves, we did not
measure higher levels of IgG2. However, higher IgG2/IgG1
FIG. 7. Evolution of viral load in BAL fluids. Virus load was
de-termined by real-time RT-PCR in BAL fluid samples collected on days
63, 66, 68, 70, 73, 75, and 77. Bars represent mean (
⫹
standard
devia-tion) standardized amounts (BRSV mRNAs/10
6
-actin mRNAs) in
each group.
on November 8, 2019 by guest
http://jvi.asm.org/
ratios were observed after challenge, further confirming that
pFsyn/pNsyn priming elicits a Th1 immune response.
The protection conferred by these vaccination protocols was
evaluated after experimental infection of calves using a BRSV
field strain (VRS 3761) successively passaged on young calves.
This method has previously been shown to induce an
excep-tionally severe pathology in mock-vaccinated calves compared
to challenge procedures based on the usage of cell
culture-passaged strains (56). In the present study, the first clinical
signs of BRSV infection were recorded 3 days following
chal-lenge and they peaked 1 week after infection. Postmortem,
consolidated areas covered up to 50% of the lung in
mock-vaccinated animals. Histopathologically, syncytia, emphysema,
alveolitis, and bronchiolitis with infiltration of numerous
neu-trophils and eosinophils appeared first, rapidly evolving in
bronchopneumonia and a prominent fibrovascular reaction
in-dicative of repair. As shown by others, the serum concentration
of Hpt, an acute-phase-response protein, peaked 7 to 9 days
after infection and the magnitude of the response was well
correlated with the severity of the clinical signs and the extent
of lung lesions (24). Thus, BRSV challenge resulted in severe
pathology similar to that seen in naturally infected calves,
allowing reliable evaluation of the protection conferred by
each vaccine strategy.
Even if it was not sufficient to fully prevent histopathological
changes and gross pneumonic lesions after challenge, the
im-mune response generated by pFsyn and pNsyn plasmids clearly
reduced both clinical signs of the infection and virus
replica-tion while KV failed to do so. In cattle and mice, secrereplica-tion of
IFN-
␥
was shown to contribute to NK activation and CD8
cytotoxic activity, resulting in viral clearance and resolution of
the infection in the airways (14, 26, 41, 55, 68). Here, we have
also shown that the production of IFN-
␥
was correlated with
viral clearance as pFsyn/pNsyn-primed calves efficiently
elimi-nated the virus after challenge. In contrast, KV vaccination,
which elicited high levels of neutralizing antibody but did not
induce IFN-
␥
production, failed to reduce viral replication and
pneumonic lesions. Thus, it seems that the induction of a
strong cell-mediated immunity is a critical parameter for the
protection against BRSV infection as the pFsyn/pNsyn
vac-cine-induced cell-mediated immunity allowed viral clearance
even in absence of a neutralizing humoral response. However,
prevention of the disease was only achieved when calves were
further vaccinated with the KV boost.
In contrast to previous reports, KV vaccination did not
in-duce a strong lymphoproliferative response and did not result
in enhanced pathology following challenge (2, 3, 19, 27, 46).
However, a single inoculation of the vaccine poorly reduced
viral load, clinical signs, histological changes, and gross
pneu-monic lesions compared to mock-vaccinated calves.
Further-more, it did not prevent eosinophilia which had previously
been linked to immunopathology following challenge (25).
These results are consistent with a previous report exploring
the efficacy of the same KV vaccine (36). This supports the
interest in DNA priming before KV vaccination. Similarly, KV
vaccines formulated with CpG oligodeoxynucleotides have also
been shown to result in a better Th1-type response and lack of
immunopathology (34, 42).
In summary, we have demonstrated for the first time that
DNA priming against BRSV F and N protein primed a
Th1-biased immune response that reduces viral replication and
partially protected calves against a highly virulent challenge.
Furthermore, KV boost potentiated the immune response
primed by DNA vaccination and this heterologous
DNA-prim-ing KV vaccine boost offered complete protection against
BRSV. In a future work, immune response induced by specific
heterologous prime-boost protocols against F or N protein as
well as strategies for improving the efficacy of DNA
vaccina-tion in cattle like microparticle encapsulavaccina-tion will be evaluated.
ACKNOWLEDGMENTS
This study was supported by the Federal Public Service Health,
Food Chain Safety and Environment (Brussels, Belgium).
The authors are grateful to K. Lentz, C. Didembourg, P. Van
Muylem, S. Durand, C. Rodeghiero, and R. Geeroms for their skilled
technical assistance. They also would like to thank all the staff of the
animal facilities of the VAR in Machelen for taking care of the
animals.
REFERENCES
1.Andre´, S., B. Seed, J. Eberle, W. Schraut, A. Bu¨ltmann, and J. Haas.1998. Increased immune response elicited by DNA vaccination with a synthetic gp120 sequence with optimized codon usage. J. Virol.72:1497–1503. 2.Antonis, A. F., E. A. Claassen, E. J. Hensen, R. J. de Groot, J. D.
Groot-Mijnes, R. S. Schrijver, and R. G. van der Most.2006. Kinetics of antiviral CD8 T cell responses during primary and post-vaccination secondary bovine respiratory syncytial virus infection. Vaccine24:1551–1561.
3.Antonis, A. F. G., R. S. Schrijver, F. Daus, P. J. G. M. Steverink, N. Stock-hofe, E. J. Hensen, J. P. M. Langedijk, and R. G. van der Most.2003. Vaccine-induced immunopathology during bovine respiratory syncytial virus infection: exploring the parameters of pathogenesis. J. Virol. 77:12067– 12073.
4.Arbiza, J., G. Taylor, J. A. Lopez, J. Furze, S. Wyld, P. Whyte, E. J. Stott, G. Wertz, W. Sullender, and M. Trudel.1992. Characterization of two antigenic sites recognized by neutralizing monoclonal antibodies directed against the fusion glycoprotein of human respiratory syncytial virus. J. Gen. Virol.73:
2225–2234.
5.Babiuk, L. A., S. L. Babiuk, and M. E. Baca-Estrada.2002. Novel vaccine strategies. Adv. Virus Res.58:29–80.
6.Bembridge, G. P., N. Rodriguez, R. Garcia-Beato, C. Nicolson, J. A. Melero, and G. Taylor.2000. DNA encoding the attachment (G) or fusion (F) protein of respiratory syncytial virus induces protection in the absence of pulmonary inflammation. J. Gen. Virol.81:2519–2523.
7.Boxus, M., C. Letellier, and P. Kerkhofs.2005. Real time RT-PCR for the detection and quantitation of bovine respiratory syncytial virus. J. Virol. Methods125:125–130.
8.Brady, R. P., C. L. Topliff, and C. L. Kelling.2004. In vitro expression of full-length and truncated bovine respiratory syncytial virus G proteins and their antibody responses in BALB/c mice. Vaccine22:3762–3768. 9.Brunak, S., J. Engelbrecht, and S. Knudsen.1991. Prediction of human
mRNA donor and acceptor sites from the DNA sequence. J. Mol. Biol.
220:49–65.
10.Bukreyev, A., I. M. Belyakov, G. A. Prince, K. C. Yim, K. K. Harris, J. A. Berzofsky, and P. L. Collins.2005. Expression of interleukin-4 by recombi-nant respiratory syncytial virus is associated with accelerated inflammation and a nonfunctional cytotoxic T-lymphocyte response following primary in-fection but not following challenge with wild-type virus. J. Virol.79:9515– 9526.
11.Cannon, M. J., P. J. Openshaw, and B. A. Askonas.1988. Cytotoxic T cells clear virus but augment lung pathology in mice infected with respiratory syncytial virus. J. Exp. Med.168:1163–1168.
12.Chang, J., S. Y. Choi, H. T. Jin, Y. C. Sung, and T. J. Braciale.2004. Improved effector activity and memory CD8 T cell development by IL-2 expression during experimental respiratory syncytial virus infection. J. Im-munol.172:503–508.
13.Connors, M., N. A. Giese, A. B. Kulkarni, C.-Y. Firestone, H. C. Morse III, and B. R. Murphy.1994. Enhanced pulmonary histopathology induced by respiratory syncytial virus (RSV) challenge of formalin-inactivated RSV-immunized BALB/c mice is abrogated by depletion of interleukin-4 (IL-4) and IL-10. J. Virol.68:5321–5325.
14.Durbin, J. E., T. R. Johnson, R. K. Durbin, S. E. Mertz, R. A. Morotti, R. S. Peebles, and B. S. Graham.2002. The role of IFN in respiratory syncytial virus pathogenesis. J. Immunol.168:2944–2952.
15.Estes, D. M., and W. C. Brown.2002. Type 1 and type 2 responses in regulation of Ig isotype expression in cattle. Vet. Immunol. Immunopathol.
90:1–10.
on November 8, 2019 by guest
http://jvi.asm.org/
16.Gaddum, R. M., R. S. Cook, J. M. Furze, S. A. Ellis, and G. Taylor.2003. Recognition of bovine respiratory syncytial virus proteins by bovine CD8⫹T lymphocytes. Immunology108:220–229.
17.Gaddum, R. M., R. S. Cook, L. H. Thomas, and G. Taylor.1996. Primary cytotoxic T-cell responses to bovine respiratory syncytial virus in calves. Immunology88:421–427.
18.Gershwin, L. J., R. A. Gunther, M. L. Anderson, A. R. Woolums, K. McArthur-Vaughan, K. E. Randel, G. A. Boyle, K. E. Friebertshauser, and P. S. McInturff.2000. Bovine respiratory syncytial virus-specific IgE is asso-ciated with interleukin-2 and -4, and interferon-gamma expression in pul-monary lymph of experimentally infected calves. Am. J. Vet. Res.61:291– 298.
19.Gershwin, L. J., E. S. Schelegle, R. A. Gunther, M. L. Anderson, A. R. Woolums, D. R. LaRochelle, G. A. Boyle, K. E. Friebertshauser, and R. S. Singer.1998. A bovine model of vaccine enhanced respiratory syncytial virus pathophysiology. Vaccine16:1225–1236.
20.Girard, M. P., T. Cherian, Y. Pervikov, and M. P. Kieny.2005. A review of vaccine research and development: human acute respiratory infections. Vac-cine23:5708–5724.
21.Gurunathan, S., D. M. Klinman, and R. A. Seder.2000. DNA vaccines: immunology, application, and optimization. Annu. Rev. Immunol.18:927– 974.
22.Gurunathan, S., C. Y. Wu, B. L. Freidag, and R. A. Seder.2000. DNA vaccines: a key for inducing long-term cellular immunity. Curr. Opin. Im-munol.12:442–447.
23.Hagglund, S., K. F. Hu, L. E. Larsen, M. Hakhverdyan, J. F. Valarcher, G. Taylor, B. Morein, S. Belak, and S. Alenius.2004. Bovine respiratory syn-cytial virus ISCOMs—protection in the presence of maternal antibodies. Vaccine23:646–655.
24.Heegaard, P. M., D. L. Godson, M. J. Toussaint, K. Tjornehoj, L. E. Larsen, B. Viuff, and L. Ronsholt.2000. The acute phase response of haptoglobin and serum amyloid A (SAA) in cattle undergoing experimental infection with bovine respiratory syncytial virus. Vet. Immunol. Immunopathol.77:
151–159.
25.Hussell, T., C. J. Baldwin, A. O’Garra, and P. J. Openshaw.1997. CD8⫹T cells control Th2-driven pathology during pulmonary respiratory syncytial virus infection. Eur. J. Immunol.27:3341–3349.
26.Hussell, T., and P. J. Openshaw.1998. Intracellular IFN-gamma expression in natural killer cells precedes lung CD8⫹T cell recruitment during respi-ratory syncytial virus infection. J. Gen. Virol.79:2593–2601.
27.Kalina, W. V., A. R. Woolums, R. D. Berghaus, and L. J. Gershwin.2004. Formalin-inactivated bovine RSV vaccine enhances a Th2 mediated immune response in infected cattle. Vaccine22:1465–1474.
28.Kerkhofs, P., M. Tignon, H. Petry, I. Mawhinney, and B. Sustronck.2004. Immune responses to bovine respiratory syncytial virus (BRSV) following use of an inactivated BRSV-PI3-Mannheimia haemolytica vaccine and a modified live BRSV-BVDV vaccine. Vet. J.167:208–210.
29.Kim, H. W., J. G. Canchola, C. D. Brandt, G. Pyles, R. M. Chanock, K. Jensen, and R. H. Parrott.1969. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epide-miol.89:422–434.
30.Kozak, M.1987. An analysis of 5⬘-noncoding sequences from 699 vertebrate messenger RNAs. Nucleic Acids Res.15:8125–8148.
31.Larsen, L. E.2000. Bovine respiratory syncytial virus (BRSV): a review. Acta Vet. Scand.41:1–24.
32.Li, X., S. Sambhara, C. X. Li, L. Ettorre, I. Switzer, G. Cates, O. James, M. Parrington, R. Oomen, R. P. Du, and M. Klein.2000. Plasmid DNA encod-ing the respiratory syncytial virus G protein is a promisencod-ing vaccine candidate. Virology269:54–65.
33.Li, X., S. Sambhara, C. X. Li, M. Ewasyshyn, M. Parrington, J. Caterini, O. James, G. Cates, R. P. Du, and M. Klein.1998. Protection against respiratory syncytial virus infection by DNA immunization. J. Exp. Med.188:681–688. 34.Mapletoft, J. W., M. Oumouna, H. G. Townsend, S. Gomis, L. A. Babiuk, and
S. van Drunen Littel-van den Hurk.2006. Formulation with CpG oligode-oxynucleotides increases cellular immunity and protection induced by vac-cination of calves with formalin-inactivated bovine respiratory syncytial virus. Virology353:316–323. [Epub 7 July 2006.]
35.Matheise, J. P., K. Walravens, A. Collard, P. Coppe, and J. J. Letesson.1995. Antigenic analysis of the F protein of the bovine respiratory syncytial virus: identification of two distinct antigenic sites involved in fusion inhibition. Arch. Virol.140:993–1005.
36.Mawhinney, I. C., and M. R. Burrows.2005. Protection against bovine respiratory syncytial virus challenge following a single dose of vaccine in young calves with maternal antibody. Vet. Rec.156:139–143.
37.Melkebeek, V., E. Sonck, F. Verdonck, B. M. Goddeeris, and E. Cox.2007. Optimized FaeG expression and a thermolabile enterotoxin DNA adjuvant enhance priming of an intestinal immune response by an FaeG DNA vaccine in pigs. Clin. Vaccine Immunol.14:28–35.
38.Mossadegh, N., L. Gissmann, M. Muller, H. Zentgraf, A. Alonso, and P. Tomakidi.2004. Codon optimization of the human papillomavirus 11 (HPV 11) L1 gene leads to increased gene expression and formation of virus-like particles in mammalian epithelial cells. Virology326:57–66.
39.Narum, D. L., S. Kumar, W. O. Rogers, S. R. Fuhrmann, H. Liang, M. Oakley, A. Taye, B. K. L. Sim, and S. L. Hoffman.2001. Codon optimization of gene fragments encodingPlasmodium falciparummerozoite proteins en-hances DNA vaccine protein expression and immunogenicity in mice. Infect. Immun.69:7250–7253.
40.Openshaw, P. J. M., and J. S. Tregoning.2005. Immune responses and disease enhancement during respiratory syncytial virus infection. Clin. Mi-crobiol. Rev.18:541–555.
41.Ostler, T., W. Davidson, and S. Ehl.2002. Virus clearance and immunopa-thology by CD8(⫹) T cells during infection with respiratory syncytial virus are mediated by IFN-gamma. Eur. J. Immunol.32:2117–2123.
42.Oumouna, M., J. W. Mapletoft, B. C. Karvonen, L. A. Babiuk, and S. van Drunen Littel-van den Hurk.2005. Formulation with CpG oligodeoxynucle-otides prevents induction of pulmonary immunopathology following priming with formalin-inactivated or commercial killed bovine respiratory syncytial virus vaccine. J. Virol.79:2024–2032.
43.Ramakrishna, L., K. K. Anand, K. M. Mohankumar, and U. Ranga.2004. Codon optimization of the Tat antigen of human immunodeficiency virus type 1 generates strong immune responses in mice following genetic immu-nization. J. Virol.78:9174–9189.
44.Robinson, H. L., and T. M. Pertmer.2000. DNA vaccines for viral infections: basic studies and applications. Adv. Virus Res.55:1–74.
45.Rutigliano, J. A., T. R. Johnson, T. N. Hollinger, J. E. Fischer, S. Aung, and B. S. Graham.2004. Treatment with anti-LFA-1 delays the CD8⫹ cytotoxic-T-lymphocyte response and viral clearance in mice with primary respiratory syncytial virus infection. J. Virol.78:3014–3023.
46.Sandbulte, M. R., and J. A. Roth.2003. Priming of multiple T cell subsets by modified-live and inactivated bovine respiratory syncytial virus vaccines. Vet. Immunol. Immunopathol.95:123–133.
47.Schreiber, P., J. P. Matheise, F. Dessy, M. Heimann, J. J. Letesson, P. Coppe, and A. Collard.2000. High mortality rate associated with bovine respiratory syncytial virus (BRSV) infection in Belgian white blue calves previously vaccinated with an inactivated BRSV vaccine. J. Vet. Med. B
47:535–550.
48.Schrijver, R. S., J. P. Langedijk, G. M. Keil, W. G. Middel, M. Maris-Veldhuis, J. T. Van Oirschot, and F. A. Rijsewijk.1998. Comparison of DNA application methods to reduce BRSV shedding in cattle. Vaccine16:130–134.
49.Shaw, G., and R. Kamen.1986. A conserved AU sequence from the 3⬘
untranslated region of GM-CSF mRNA mediates selective mRNA degra-dation. Cell46:659–667.
50.Skinner, J. G., R. A. Brown, and L. Roberts. 1991. Bovine haptoglobin response in clinically defined field conditions. Vet. Rec.128:147–149. 51.Srikiatkhachorn, A., and T. J. Braciale.1997. Virus-specific CD8⫹T
lym-phocytes downregulate T helper cell type 2 cytokine secretion and pulmo-nary eosinophilia during experimental murine respiratory syncytial virus infection. J. Exp. Med.186:421–432.
52.Stott, E. J., and G. Taylor.1985. Respiratory syncytial virus. Brief review. Arch. Virol.84:1–52.
53.Taylor, G., C. Bruce, A. F. Barbet, S. G. Wyld, and L. H. Thomas.2005. DNA vaccination against respiratory syncytial virus in young calves. Vaccine23:
1242–1250.
54.Taylor, G., L. H. Thomas, J. M. Furze, R. S. Cook, S. G. Wyld, R. Lerch, R. Hardy, and G. W. Wertz.1997. Recombinant vaccinia viruses expressing the F, G or N, but not the M2, protein of bovine respiratory syncytial virus (BRSV) induce resistance to BRSV challenge in the calf and protect against the development of pneumonic lesions. J. Gen. Virol.78:3195–3206. 55.Taylor, G., L. H. Thomas, S. G. Wyld, J. Furze, P. Sopp, and C. J. Howard.
1995. Role of T-lymphocyte subsets in recovery from respiratory syncytial virus infection in calves. J. Virol.69:6658–6664.
56.Tjornehoj, K., A. Uttenthal, B. Viuff, L. E. Larsen, C. Rontved, and L. Ronsholt.2003. An experimental infection model for reproduction of calf pneumonia with bovine respiratory syncytial virus (BRSV) based on one combined exposure of calves. Res. Vet. Sci.74:55–65.
57.Toussaint, J. F., C. Letellier, D. Paquet, M. Dispas, and P. Kerkhofs.2005. Prime-boost strategies combining DNA and inactivated vaccines confer high immunity and protection in cattle against bovine herpesvirus-1. Vaccine
23:5073–5081.
58.Tree, J. A., G. Bembridge, S. Hou, G. Taylor, E. Fashola-Stone, J. Melero, and M. P. Cranage.2004. An assessment of different DNA delivery systems for protection against respiratory syncytial virus infection in the murine model: gene-gun delivery induces IgG in the lung. Vaccine22:2438–2443. 59.Tristram, D. A., R. C. Welliver, D. A. Hogerman, S. W. Hildreth, and P.
Paradiso.1994. Second-year surveillance of recipients of a respiratory syn-cytial virus (RSV) F protein subunit vaccine, PFP-1: evaluation of antibody persistence and possible disease enhancement. Vaccine12:551–556. 60.Tristram, D. A., R. C. Welliver, C. K. Mohar, D. A. Hogerman, S. W.
Hildreth, and P. Paradiso.1993. Immunogenicity and safety of respiratory syncytial virus subunit vaccine in seropositive children 18–36 months old. J. Infect. Dis.167:191–195.
61.Valarcher, J. F., and G. Taylor. 2007. Bovine respiratory syncytial virus infection. Vet. Res.38:153–180.
on November 8, 2019 by guest
http://jvi.asm.org/
62.Van der Poel, W. H., A. Brand, J. A. Kramps, and J. T. Van Oirschot.1994. Respiratory syncytial virus infections in human beings and in cattle. J. Infect.
29:215–228.
63.Varga, S. M., X. Wang, R. M. Welsh, and T. J. Braciale.2001. Immunopa-thology in RSV infection is mediated by a discrete oligoclonal subset of antigen-specific CD4(⫹) T cells. Immunity15:637–646.
64.Vaughan, K., G. H. Rhodes, and L. J. Gershwin.2005. DNA immunization against respiratory syncytial virus (RSV) in infant rhesus monkeys. Vaccine
23:2928–2942.
65.Wahle, E., and W. Keller.1996. The biochemistry of polyadenylation. Trends Biochem. Sci.21:247–250.
66.Woodland, D. L.2004. Jump-starting the immune system: prime-boosting comes of age. Trends Immunol.25:98–104.
67.Woolums, A. R., R. A. Gunther, K. McArthur-Vaughan, M. L. Anderson, A. Omlor, G. A. Boyle, K. E. Friebertshauser, P. S. McInturff, and L. J. Gershwin.2004. Cytotoxic T lymphocyte activity and cytokine expression in calves vaccinated with formalin-inactivated bovine respiratory syncytial virus prior to challenge. Comp. Immunol. Microbiol. Infect. Dis.27:57–74. 68.Woolums, A. R., R. S. Singer, G. A. Boyle, and L. J. Gershwin.1999.
Inter-feron gamma production during bovine respiratory syncytial virus (BRSV) infection is diminished in calves vaccinated with formalin-inactivated BRSV. Vaccine17:1293–1297.
on November 8, 2019 by guest
http://jvi.asm.org/