Copyright © 2010, American Society for Microbiology. All Rights Reserved.
The Globoside Receptor Triggers Structural Changes in the B19 Virus
Capsid That Facilitate Virus Internalization
䌤
Claudia Bo
¨nsch,
1Christoph Zuercher,
1§ Patricia Lieby,
2Christoph Kempf,
1,2and Carlos Ros
1,2*
Department of Chemistry and Biochemistry, University of Bern, Freiestrasse 3, 3012 Bern,1and CSL Behring AG, Wankdorfstrasse 10,3000 Bern 22,2Switzerland
Received 28 May 2010/Accepted 26 August 2010
Globoside (Gb4Cer), Ku80 autoantigen, and␣51 integrin have been identified as cell receptors/coreceptors for human parvovirus B19 (B19V), but their role and mechanism of interaction with the virus are largely unknown. In UT7/Epo cells, expression of Gb4Cer and CD49e (integrin alpha-5) was high, but expression of Ku80 was insignificant. B19V colocalized with Gb4Cer and, to a lesser extent, with CD49e. However, only anti-Gb4Cer antibodies could disturb virus attachment. Only a small proportion of cell-bound viruses were internalized, while the majority became detached from the receptor. When added to uninfected cells, the receptor-detached virus showed superior cell binding capacity and infectivity. Attachment of B19V to cells triggered conformational changes in the capsid leading to the accessibility of the N terminus of VP1 (VP1u) to antibodies, which was maintained in the receptor-detached virus. VP1u became similarly accessible to anti-bodies following incubation of B19V particles with increasing concentrations of purified Gb4Cer. The receptor-mediated exposure of VP1u is critical for virus internalization, since capsids lacking VP1 could bind to cells but were not internalized. Moreover, an antibody against the N terminus of VP1u disturbed virus internal-ization, but only when present during and not after virus attachment, indicating the involvement of this region in binding events required for internalization. These results suggest that Gb4Cer is not only the primary receptor for B19V attachment but also the mediator of capsid rearrangements required for subsequent interactions leading to virus internalization. The capacity of the virus to detach and reattach again would enhance the probability of productive infections.
Human parvovirus B19 (B19V) belongs to theErythrovirus
genus of the Parvoviridaefamily. The virus has a worldwide distribution and typically causes a mild childhood febrile illness known as erythema infectiosum or fifth disease. In patients with underlying immunologic and hematologic disorders, B19V has been associated with more severe manifestations, such as arthropathies, aplastic anemia, hydrops fetalis, and fetal death (4).
B19V has a single-stranded DNA genome encapsidated in a T⫽1 nonenveloped icosahedral capsid. The capsid is assem-bled from two structural proteins, VP1 (83 kDa) and VP2 (58 kDa). VP1 is identical to VP2, with the exception of 227 amino acids (aa) in the N-terminal part, the so-called VP1 unique region (VP1u) (9, 26). Despite VP1u being the minor compo-nent of the capsid, the dominant immune response against B19V is elicited by the VP1u region, which harbors strong neutralizing epitopes (2, 31, 41). A secreted phospholipase A2 (PLA2)-like activity has been located in the VP1 unique region
of B19V (12), which is required for infection (13, 17, 40). Despite all these properties, we recently showed that VP1u is not accessible to antibodies. However, brief exposure to mild temperatures or low pH can render this region accessible (30).
In this sense, B19V is similar to other parvoviruses in which VP1u is not accessible but can become exposedin vitroby mild heat or low-pH treatment (10, 21). In all parvoviruses tested so far, VP1u becomes exposed during the intracellular trafficking of the virus (18, 23, 28, 32, 33). However, B19V VP1u harbors strong neutralizing epitopes, meaning that its accessibility to antibody binding should occur prior to uptake by cells. In line with this hypothesis, we have demonstrated that incubation of B19V with red blood cells, which allow virus binding but not virus internalization, can trigger the externalization of VP1u in a proportion of the bound particles (3).
The glycosphingolipid globoside (globotetraosylceramide [Gb4Cer]) is the cellular receptor of B19V (5, 6). Gb4Cer is largely expressed in human erythroid progenitor cells in the bone marrow, which are the main target cells for the virus. However, the pathogenicity and tropism of B19V cannot be explained if Gb4Cer is the only receptor. Previous studies have suggested that Gb4Cer is necessary for B19V to bind to cells but is not sufficient for cell entry (35). Subsequently, ␣51 integrin (36, 37) and the Ku80 autoantigen (25) were identified as coreceptors for B19V infection. While Ku80 might assist in virus attachment (25),␣51 integrin is thought to be required for internalization (36, 37). In line with a complex mechanism of internalization based on multiple receptors is the observa-tion that B19V does not stably bind membrane-associated glo-bosidein vitro(20), indicating that B19V probably binds glo-boside jointly with other molecular structures present on cell membranes.
In the present studies, the interaction of B19V with cell surface receptors and the implication of this interaction for the capsid structure were examined. The cells chosen for this study
* Corresponding author. Mailing address: Department of Chemistry and Biochemistry, University of Bern, Freiestrasse 3, 3012 Bern, Swit-zerland. Phone: 41 31 6314349. Fax: 41 31 6314887. E-mail: carlos.ros @ibc.unibe.ch.
§ Present address: Institute of Veterinary Virology, University of Bern, La¨nggass-Str. 122, P.O. Box 8466, CH-3001 Bern, Switzer-land.
䌤Published ahead of print on 8 September 2010.
11737
on November 8, 2019 by guest
http://jvi.asm.org/
were of the erythropoietin (Epo)-dependent bone marrow megakaryoblastic leukemia UT7/Epo cell line, which is com-monly used to study B19V infection. UT7/Epo cells support viral DNA replication and protein expression; however, intra-cellular factors limit the production of infectious progeny virus. The results indicate that B19V interacts dynamically with cell surface receptors and that internalization is a complex process involving sequential steps. B19V binds initially to Gb4Cer, which triggers the externalization of VP1u. The modified cap-sid is then ready for a subsequent interaction. Whenever the second interaction is not possible, the virus detaches from Gb4Cer and is ready for another attempt. This mechanism of “detachment-reattachment” is repeated until the required sec-ond interaction occurs, after which the virus is internalized.
MATERIALS AND METHODS
Cells and viruses.UT7/Epo cells were cultured in RPMI 1640, 10% fetal calf serum (FCS), and 2 U/ml of recombinant human Epo (Janssen-Cilag, Midrand,
South Africa) at 37°C and 7.5% CO2. A B19V-infected plasma sample was
obtained from our donation center (genotype 1; CSL Behring AG, Charlotte, NC). The viremic serum sample did not contain B19V-specific IgM or IgG antibodies. B19V was concentrated from the infected plasma sample by ultra-centrifugation through 20% sucrose. The viral pellets were washed and resus-pended in phosphate-buffered saline (PBS). B19V titers were determined by quantitative PCR and expressed as numbers of DNA-containing particles per microliter. To obtain receptor-detached virus, UT7/Epo cells were infected with B19V at 4°C for 1 h. The cells were repeatedly washed with PBS to remove unbound virus and further incubated in RPMI 1640 at 37°C for 1 h. The cells were pelleted, the supernatant containing the detached virus was recovered, and titers were determined as specified above. Baculovirus-expressed B19V-like empty particles (VLPs) containing VP1 and VP2 were kindly provided by R. Franssila (Helsinki, Finland) and were produced in High-5 cells as described elsewhere (15). VP2-only empty particles lacking VP1 were kindly provided by G. Elliott (Biotrin Technologies Ltd., Dublin, Ireland).
Antibodies.A rabbit polyclonal antibody against VP1u (aa 142 to 163; referred to as anti-VP1u) was obtained as previously described (3). Two monoclonal antibodies (MAbs), one specific for the most N-terminal region of VP1u (aa 30 to 42) (MAb 1418-1; referred to as anti-N-VP1u) and the other recognizing only intact capsids (MAb 860-55D; referred to as anti-Caps), were kindly provided by S. Modrow (Regensburg, Germany). The two MAbs were produced from pe-ripheral blood mononuclear cells from healthy individuals with high titers of serum antibodies against B19 virus proteins (16) and were shown to be highly neutralizing (12, 16, 30). For detection of B19V proteins by immunoblotting, a mouse anti-B19V VPs antibody was used (U.S. Biologicals, Swampscott, MA). A globoside-specific mouse IgM MAb (AME-2; referred to as anti-Gb4Cer) was kindly provided by J. de Jong (Netherlands Red Cross, Amsterdam, Nether-lands) (34). Mouse IgG MAbs against Ku80 and CD49e were purchased from BD Biosciences (San Jose, CA). Mouse nonspecific IgM/IgG isotype control antibodies were purchased from Sigma (St. Louis, MO).
Analysis of cell surface receptors.The presence of B19V receptors or core-ceptors on the surfaces of UT7/Epo cells was analyzed quantitatively by flow cytometry. UT7/Epo cells were incubated with either an Ku80 or anti-Gb4Cer antibody at 4°C for 1 h in PBS containing 2% fetal calf serum, followed by incubation with fluorescein isothiocyanate (FITC)-conjugated rat anti-mouse IgG or IgM, respectively (BD Biosciences). Additionally, UT7/Epo cells were stained with R-phycoerythrin-conjugated anti-human CD49e (BD Biosciences). The cells were analyzed on a BD FacsCanto II flow cytometer (Becton Dickin-son, San Jose, CA). Data acquisition and analysis were conducted with BD FacsDiva software (BD Biosciences).
For detection of CD49e by immunofluorescence, cells were fixed with
acetone-methanol (1/1 [vol/vol]) for 5 min at⫺20°C before being stained with antibodies.
For detection of Gb4Cer, cells were stained before fixation. After fixation, the cells were air dried, washed with PBSA (PBS plus 1% bovine serum albumin [BSA]), and stained with anti-CD49e (diluted 1:400 in PBSA) for 1 h at room temperature. The cells were washed three times with PBSA for 5 min each, and secondary antibodies were added (diluted 1:400 in PBSA) (Santa Cruz Biotech-nology, Santa Cruz, CA) for 1 h at room temperature. For detection of Gb4Cer, the procedure was the same, but the antibodies (same dilutions and incubation conditions) were added before fixation. After final washings with PBSA, the cells
were washed and mounted with Mowiol (Calbiochem, La Jolla, CA) containing 30 mg/ml of Dabco (Sigma) as an antifading agent and were examined by fluorescence microscopy.
Colocalization of cell surface receptors with B19V capsids.UT7/Epo cells
were infected for 1 h at 4°C (2⫻104
virions per cell) and then washed extensively to remove unbound viruses. Detection of CD49e and Gb4Cer was performed as described above, but the human MAb against intact capsids (MAb 860-55D) was also applied (diluted 1:400 in PBSA). Before being mounted with Mowiol, nuclei
were stained with DAPI (4⬘,6-diamidino-2-phenylindole; Invitrogen, Eugene,
OR). Uninfected cells and nonspecific IgM/IgG isotype control antibodies were used as controls. Cells were examined by confocal laser scanning microscopy.
Quantitative PCR.Amplification of B19V DNA and real-time detection of PCR products were performed by using a LightCycler system (Roche Diagnos-tics, Rotkreuz, Switzerland) with SYBR green (Roche). PCR was carried out using a FastStart DNA SYBR green kit (Roche) following the manufacturer’s instructions. Primers used for B19V DNA amplification were as follows:
B19V-forward, 5⬘-GGGCAGCCATTTTAAGTGTTT-3⬘; and B19V-reverse, 5⬘-GCA
CCACCAGTTATCGTTAGC-3⬘. Plasmids containing the genome of B19V
were used at 10-fold dilutions as external standards. PCR specificity was verified using melting curve analysis. The number of cells used for each experiment was
determined by quantification of the cell-actin gene, as previously described
(27).
B19V binding assays.UT7/Epo cells (3⫻105
) were incubated with antibodies specific for the cellular receptors globoside, CD49e, and Ku80 or with a similar concentration of nonspecific IgM/IgG isotype control antibodies (Sigma). After
1 h at 4°C, B19V was added (3⫻104virions per cell) and further incubated at
4°C for 1 h. Following washings to remove unbound virus, the cells were lysed and total DNA was extracted by using a DNeasy tissue kit (Qiagen, Valencia, CA). Viral DNA was quantified from three independent experiments as specified above.
Analysis of B19V dynamics on the cell surface.The dynamics of B19V binding, detachment, and internalization were examined in UT7/Epo cells by quantitative
PCR. B19V was added to cells (3⫻104virions per cell). After 1 h at 4°C, the
cells were washed repeatedly to remove unbound virus and then transferred to 37°C to allow virus internalization. At progressive times, the cells were pelleted at 4°C and the supernatant was transferred to a new tube and used to quantify receptor-detached viruses. The cell pellets were washed and used to quantify the cell-associated virus. To detect internalized virus, the cell pellets were resus-pended in a trypsin-EDTA solution and incubated for 4 min at 37°C. The cell pellets were washed and used to quantify the internalized virus. Total DNA was extracted from the cell and supernatant fractions by using a DNeasy tissue kit (Qiagen). Viral DNA was quantified from three independent experiments by using real-time PCR as explained above.
Infectivity assay.Virus infectivity was examined by quantification of NS1 mRNA. Cells were transferred at 24 h postinfection to RNase-free tubes (Safe-Lock 1.5-ml tubes; Eppendorf Biopur) and then pelleted. The pellet was washed twice with PBS
and stored at⫺20°C until use. Total poly(A) mRNA was isolated with an mRNA
isolation kit (Roche). Following reverse transcription, cDNAs from three indepen-dent experiments were quantified by using a LightCycler 2.0 system (Roche) and a LightCycler FastStart DNA Master SYBR green I kit (Roche). The following prim-ers were chosen to amplify a 133-nucleotide (nt) NS1 cDNA fragment: NS1 forward,
5⬘-GGGGCAGCATGTGTTAAAG-3⬘(nt 1017 to 1035); and NS1 reverse, 5⬘-CC
ATGCCATATACTGGAACACT-3⬘(nt 1129 to 1150).
Analysis of VP1u accessibility by immunoprecipitation.UT7/Epo cells were infected at 4°C with B19V as described above. After being washed to remove unbound virus, the cells were lysed in NP-40 lysis buffer (50 mM Tris-HCl, 150 mM NaCl, 1% NP-40, 5 mM EDTA) supplemented with protease inhibitor cocktail (Complete Mini; Roche). B19V capsids were immunoprecipitated with a human MAb against intact capsids (MAb 860-55D) or with a rabbit polyclonal antibody against VP1u. After overnight incubation at 4°C in the presence of 20
l of protein G agarose beads, the supernatants were discarded and the beads
were washed four times with PBSA. Immunoprecipitated virions were analyzed by SDS-PAGE. Additionally, the immunoprecipitated virions from four inde-pendent experiments were quantified by quantitative PCR.
B19V-globoside interactionin vitro.Purified Gb4Cer (Sigma) was resuspended
in chloroform-methanol (2:1). B19V native capsids (3⫻109
) were incubated
overnight at 4°C with increasing concentrations of Gb4Cer (0 to 20M).
Fol-lowing 2 h of additional incubation at 37°C, viral particles were immunoprecipi-tated at 4°C with a human MAb against intact capsids (MAb 860-55D) or with a rabbit polyclonal antibody against VP1u. Immunoprecipitation and quantifica-tion were performed for three independent experiments as described above.
Analysis of B19V capsid stability.The stability of B19V capsids was examined by use of native capsids, capsids attached to the cellular receptor, and capsids
on November 8, 2019 by guest
http://jvi.asm.org/
that became detached from the receptor. Equivalent amounts of the capsid preparations were exposed to increasing temperatures for 3 min and then cooled on ice. Subsequently, in order to examine viral DNA externalization, DNase I was added and samples were incubated overnight at 4°C. Total DNA was ex-tracted, and viral DNAs from three independent experiments were quantified as specified above.
RESULTS
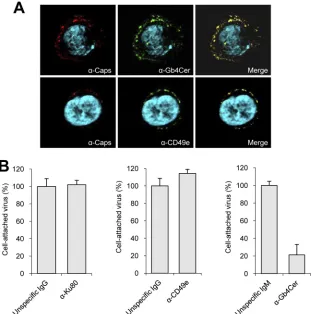
Globoside is the primary attachment receptor of B19V in UT7/Epo cells.The expression of B19V receptors in UT7/Epo cells was analyzed quantitatively by flow cytometry. Expression levels of CD49e (integrin alpha-5 subunit, which forms a het-erodimer with the beta-1 subunit) were high (66%⫾ 0.6%). Gb4Cer was also abundant (29%), but the expression level varied considerably with cell passage (data not shown). Expres-sion of Ku80 in UT7/Epo cells was insignificant (0.25% ⫾ 0.07%) (Fig. 1A), confirming the results obtained in previous studies (39). Therefore, the Ku80 autoantigen does not play a role in B19V binding to UT7/Epo cells. Immunofluorescence staining of cells with antibodies against Gb4Cer confirmed that Gb4Cer was abundantly expressed, although it was not distrib-uted homogeneously among the cells (Fig. 1B). Double
stain-ing with an antibody against the viral receptor and an antibody against intact capsids showed extensive colocalization of B19V with Gb4Cer and, to a lesser extent, with CD49e (Fig. 2A). Virus attachment was disturbed in cells preincubated with anti-Gb4Cer; in contrast, no effect was observed in cells preincu-bated with anti-Ku80 or anti-CD49e (Fig. 2B), indicating that preliminary attachment of B19V to UT7/Epo cells is achieved through Gb4Cer.
A large proportion of cell-bound capsids are not internal-ized and instead dissociate from the receptor.Binding of na-tive capsids to Gb4Cer was not stable for a large proportion of capsids, which were rapidly detached when the temperature was changed from 4°C to 37°C. The capsids that remained stably bound became progressively internalized (Fig. 3A). Vi-rus detachment was also observed after incubation of cells with decreasing amount of viruses, from 3⫻104to 103viruses/cell
(data not shown), indicating that viral load was not critical. Virus-receptor dissociation kinetics were slower at 4°C than at 37°C; however, independent of the temperature, the total amount of viruses that finally became detached was approxi-mately two-thirds of the total amount initially bound (Fig. 3B). Tendencies diverged with prolonged incubation times. At 4°C, where internalization and receptor recycling are impaired, a progressive and slow detachment of all viruses was observed. However, prolonged incubation at 37°C resulted in a slow and progressive reattachment of the receptor-dissociated capsids, probably to newly expressed and/or recycled receptors (Fig. 3B). These data suggest that rebinding of viruses to the same receptors from which they dissociated does not occur.
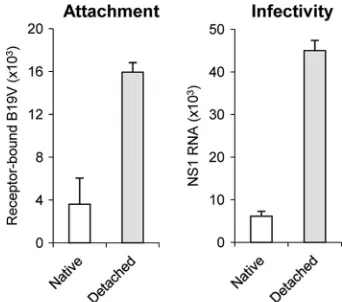
Receptor-detached virus shows superior cell binding capac-ity and infectivcapac-ity.Cell binding and infectivity were compared between receptor-detached and native viruses. When added in equal numbers to uninfected cells, the receptor-detached virus showed a superior cell binding capacity as well as a superior infectivity to that of the native virus (Fig. 4). This result sug-gests the existence of beneficial structural differences in the detached particles improving the infection.
Binding of B19V to UT7/Epo cells triggers conformational changes in the capsid leading to the exposure of VP1u. We have previously shown that VP1u of B19V, similar to those of other parvoviruses, is not accessible to antibody binding. However, low pH, temperature treatment, or incubation with red blood cells can trigger the externalization of this region (3, 30). Incubation of B19V with UT7/Epo cells trig-gered the accessibility of VP1u (Fig. 5A). Quantification analysis of the immunoprecipitated capsids indicated that fewer than 10% of the native particles had VP1u already externalized. In contrast, the proportion of capsids with exposed VP1u increased in the receptor-bound particles, to approximately 80%. A similar proportion was also found in the detached virus, indicating that the receptor-induced VP1u exposure was irreversible (Fig. 5B). When the receptor-bound capsids were further incubated for 1 h at 4°C, to avoid virus internalization and to initiate virus de-tachment (as shown in Fig. 3B), all capsids remaining stably bound to the cells had VP1u accessible (Fig. 5C), suggesting that the external position of VP1u assists in stabilizing the receptor-capsid complex.
[image:3.585.61.265.68.379.2]Studies of capsid disassemblyin vitrosuggest that the con-formational transitions that lead to the exposure of VP1u also
FIG. 1. Detection of B19V receptors in UT7/Epo cells by flow cytometry and immunofluorescence. (A) The presence of the B19V receptors Gb4Cer, Ku80, and CD49e (integrin alpha-5) on the surfaces of UT7/Epo cells was determined quantitatively by fluorescence-acti-vated cell sorter (FACS) analysis. The proportion of positive cells is shown in the upper right quadrant of each panel. The background control consisted of cells stained with the conjugated secondary anti-body. (B) Immunofluorescence picture of a large field showing UT7/ Epo cells (nuclei stained with DAPI) and Gb4Cer (green).
on November 8, 2019 by guest
http://jvi.asm.org/
lead to the externalization of the viral genome (10, 11, 29). It was therefore of interest to examine whether the receptor-induced externalization of VP1u would also result in accessi-bility of the viral DNA. Cell-bound viruses were immunopre-cipitated as described above. The immunopreimmunopre-cipitated capsids were treated with DNase I prior to DNA extraction and quan-tification. The results showed that the viral DNA remained encapsidated (Fig. 5D). Additionally, receptor-detached cap-sids, which also have VP1u accessible, were directly incubated with DNase I or AccI, which cleaves a sequence within the 3⬘ palindromic sequence. These treatments had no effect on viral infectivity, indicating that despite the structural changes lead-ing to VP1u exposure, the viral genome remained fully encap-sidated and infectious (Fig. 5E).
Purified globoside can trigger the externalization of VP1u.
In addition to Gb4Cer, Ku80 and integrin ␣51 have been recognized as coreceptors of B19V (25, 36). However, expres-sion analysis of these receptors in UT7/Epo cells by flow cy-tometry indicated that only a negligible subset of UT7/Epo cells expressed Ku80 (Fig. 1A). Additionally, blocking CD49e with an antibody disturbing ligand binding had no effect on B19V attachment (Fig. 2B), indicating that CD49e does not play a critical role in initial virus attachment, but rather in subsequent steps. These results are in agreement with previous studies indicating that Gb4Cer is the primary receptor for B19V attachment (5, 35). However, the observed
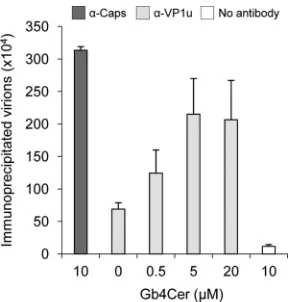
conforma-tional changes might also result from an interaction of B19V with Gb4Cer jointly with other structures on the cell mem-brane. In order to examine whether Gb4Cer alone is able to trigger the exposure of VP1u, B19V capsids were incubated with purified Gb4Cer receptor. As shown in Fig. 6, increasing amounts of B19V capsids externalized VP1u following incuba-tion with increasing concentraincuba-tions of purified Gb4Cer.
Receptor-bound capsids acquire superior stability com-pared to receptor-detached and native particles. We previ-ously showed that heat-induced DNA accessibility was remark-ably more pronounced in B19V than in any other parvovirus, explaining the lower resistance of B19V to inactivation treat-ments (24). This difference was also evident under conditions mimicking endosomal acidification (pH 5 to 6.5), which trig-gered the externalization of B19V DNA but not of minute virus of mice (MVM) DNA (29). Accordingly, trafficking of incoming B19V through the acidic endosomal compartment would presumably cause the degradation of the exposed viral DNA by lysosomal enzymes. We therefore analyzed whether the receptor-induced structural changes in the B19V capsid would also modify the overall stability of the virion. Cell-bound, receptor-detached, and native capsids (alone or in the presence of cells) were exposed to increasing temperatures for 3 min. Subsequently, the samples were cooled and incubated with DNase I at 4°C. Total DNA was extracted, and the intact viral DNA was quantified. The results showed that
receptor-FIG. 2. Role of cell receptors in B19V attachment. (A) Immunofluorescence scanning microscopy of representative cells showing colocalization of B19V (red) with Gb4Cer or CD49e (green). A merged image is shown. (B) Quantitative comparison of B19V binding to UT7/Epo cells in the presence of antibodies against Ku80, CD49e, and Gb4Cer.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:4.585.137.453.69.383.2]bound capsids were markedly more resistant to heat than the native or detached particles (Fig. 7). Interestingly, although VP1u externalization was also detected in the detached cap-sids, the stability of the capsid was only slightly superior to that
of native capsids. This observation suggests that B19V traffick-ing through low-pH compartments could be assisted by an increased stability of the Gb4Cer-bound capsids, minimizing virus degradation. The fact that the increased stability is lost upon virus-receptor dissociation would facilitate uncoating.
[image:5.585.79.249.69.327.2]Receptor-induced externalization of VP1u facilitates virus internalization.Apart from Gb4Cer, B19V might interact with additional structures at the cell surface, notably integrins, and such interactions are thought to play a role in virus internal-ization (36, 37, 38). It is tempting to speculate that the newly exposed VP1u domains interact with integrin subunits and that virus internalization ensues as a consequence. In order to ex-amine the role of the exposed VP1u in virus internalization,
FIG. 3. Dynamics of B19V receptor detachment and internaliza-tion in UT7/Epo cells. (A) At progressive times following incubainternaliza-tion at 37°C, total virus (}; virus in the supernatant and cell pellet) and cell-detached (f; virus in the supernatant), cell-associated (Œ; virus in the cell pellet), and internalized (E; virus in the cell pellet after trypsinization) virions were quantified. (B) Kinetics of receptor de-tachment at 37°C (f) and 4°C (䡺). The upper, discontinuous line represents the total number of cell-bound virions before incubation. The lower dotted line represents the maximum amount of receptor-dissociated virus at 37°C.
[image:5.585.319.520.72.397.2]FIG. 4. Binding and infectivity of native and detached viruses. UT7/Epo cells were incubated with equal amounts of native and de-tached B19V at 4°C for 1 h. Following washings to remove unbound virus, total DNA was extracted and viral DNA was quantified by quantitative PCR. Virus infectivity was examined by quantification of NS1 mRNA at 24 h postinfection.
FIG. 5. Analysis of VP1u accessibility and viral DNA exposure following binding to cells. UT7/Epo cells were infected at 4°C with B19V. After being washed to remove unbound virus, the cells were lysed. B19V capsids were immunoprecipitated with MAb 850-55D (against intact capsids) or with a rabbit polyclonal antibody against VP1u. Native particles were immunoprecipitated in the presence of lysis buffer. As a control, an unspecific IgG control antibody was used. (A) Immunoprecipitated capsids were analyzed by SDS-PAGE. (B) Immunoprecipitated virions were quantified by real-time PCR. (C) Quantification of VP1u exposure after prolonged incubation at 4°C for 1 h. (D) Analysis of viral DNA externalization in receptor-bound virions. Following B19V binding to UT7/Epo cells, the cells were washed, lysed, and incubated with DNase I. Subsequently, intact viral DNA was quantified by real-time PCR. As a control for DNase I digestion, capsids pretreated at 65°C for 3 min were used. (E) Infec-tivity of receptor-detached virus following incubation with DNase I or the AccI restriction enzyme.
on November 8, 2019 by guest
http://jvi.asm.org/
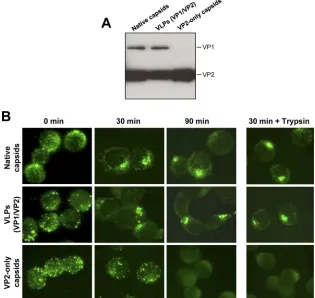
[image:5.585.76.247.516.667.2]the binding and internalization of native virions and VLPs (empty capsids composed of VP1 and VP2) were compared to those of VP2-only particles (empty capsids devoid of VP1) by immunofluorescence microscopy. Western blot analysis of the different capsid preparations confirmed that VP2-only parti-cles did not contain VP1 (Fig. 8A). All partiparti-cles were able to bind UT7/Epo cells at 4°C, with apparently similar efficiencies. However, following incubation at 37°C to allow virus internal-ization, VP2-only particles were not internalized and were instead progressively detached from the cells, being hardly detectable after 90 min. In contrast, native virus and VLPs were internalized rapidly and accumulated in a region of the cytoplasm, with no detectable differences (Fig. 8B). To further confirm this observation, following virus internalization for 30 min at 37°C, the cells were shortly trypsinized to remove un-internalized capsids. While in cells infected with native virus and VLPs the capsids appeared with typical intracellular accu-mulation, no significant signal was detected in the cells incu-bated with VP2-only particles (Fig. 8B).
The most N-terminal part of VP1u is involved in binding events required for internalization.The deficient internaliza-tion of VP2-only particles suggests that the presence of VP1 as a component of the capsid is dispensable for cell binding but required for the internalization step. We next analyzed whether VP1u is required for internalization. The mechanism of neutralization by a MAb that targets the most N-terminal part of VP1u (MAb 1418-1; referred to as anti-N-VP1u) was investigated and compared with that of a MAb against intact capsids (MAb 860-55D; referred to as Caps). Both anti-bodies were previously found to be highly neutralizing (12, 16, 30). In our assay, the antibodies were highly neutralizing when added during virus binding. However, in contrast to anti-Caps, anti-N-VP1u was not neutralizing when added after virus bind-ing (Fig. 9A). This result suggests that the epitope recognized by anti-N-VP1u is involved in binding events that hinder its accessibility to the antibody. In our previous studies, we have shown that anti-N-VP1u does not influence the PLA2activity of the virus (3) and does not disturb virus binding to UT7/Epo cells (30). In order to verify the infection step disrupted by
anti-N-VP1u, we analyzed the capacity of B19V to be inter-nalized in the presence of this antibody. The antibody par-tially disturbed virus internalization, but similar to the re-sults obtained with the infectivity assay, the effect was evident only when the antibody was present during virus binding, not when it was present after virus binding (Fig. 9B), confirming that the N terminus of VP1u is involved in interaction events required for virus internalization but not for initial virus attachment. The strong neutralizing effect by anti-N-VP1u could not be explained exclusively by its ability to disturb the internalization step, which was only partial. Accordingly, the antibody might extend its neutralizing ac-tivity beyond internalization to intracellular trafficking.
DISCUSSION
Binding of B19V to the cellular receptor globoside (Gb4Cer) induces structural changes in the capsid leading to accessibility of the N-terminal region of VP1 (VP1u). Al-though such large receptor-induced structural changes have not yet been observed in other parvoviruses, a slight opening of the 5-fold axis pore has been detected following binding of adeno-associated virus 2 (AAV-2) to heparin (22). The expan-sion of the 5-fold axis pore is believed to facilitate the exter-nalization of VP1u, which in all parvoviruses studied so far occurs during the intracellular trafficking of the capsid medi-ated by the acidified endosomal environment (18, 23, 28, 32, 33). Therefore, B19V would be unique among parvoviruses in that the conformational changes leading to the accessibility of VP1u sequences do not occur inside the cell but prematurely, at the cell membrane. The question remains whether VP1u is not accessible because it is buried within the capsid, similar to those of other parvoviruses, or because it is already external but not accessible to antibodies due to a particular “unattain-able” conformation. An argument against the latter hypothesis is that the 5-fold channel, which is suggested to be used during VP1u externalization, is closed in the B19V particle (1, 8). However, it cannot be excluded that structural rearrangements during infection, similar to those observed in AAV-2 capsids bound to heparin (22), may eventually open or expand a flex-ible pore. It is also not known whether the observed
[image:6.585.91.238.67.218.2]rearrange-FIG. 7. Analysis of B19V capsid stability. Similar amounts of var-ious capsid preparations (capsids bound to cells, detached capsids, and capsids from native virus, with or without cells) were exposed to in-creasing temperatures for 3 min. Following incubation of the capsids with DNase I, total DNA was extracted and quantified.
FIG. 6. B19V-globoside interactionin vitro. Increasing concentra-tions of purified Gb4Cer were incubated with viral particles. Subse-quently, capsids were immunoprecipitated with a human MAb against intact capsids (860-55D) or with a rabbit polyclonal antibody against VP1u. Quantification of the immunoprecipitated virions was per-formed by real-time PCR.
on November 8, 2019 by guest
http://jvi.asm.org/
[image:6.585.350.493.68.205.2]ments concern the complete VP1u region or some domains. In our previous studies, we examined the accessibility of two dis-tant regions of VP1u: one region in the N-terminal part, which contains numerous neutralizing epitopes, and a second region in the opposite part, where the PLA2 motif is located. Both regions were not accessible to antibodies in the native particles but became exposed upon mild heat and low-pH treatments without capsid disassembly (3, 30).
The early VP1u exposure prior to virus internalization and the fact that it remains stably exposed in receptor-dissociated virus would explain how originally inaccessible regions of the capsid can harbor numerous neutralizing epitopes (2, 31, 41). Accordingly, neutralizing antibodies raised against those epitopes would not target circulating native viruses but would exclusively target viruses in the process of cell attachment and receptor-dissociated viruses.
Similar to the case with MVM (10, 11), physicochemical treatments of B19V capsidsin vitrolead simultaneously to both VP1u and viral genome exposure without capsid disassembly (29). However, an important difference is that receptor binding leads to VP1u exposure but not to viral genome externalization (Fig. 5). Therefore, physicochemical treatmentsin vitrodo not entirely mimic or are not able to fully dissect the sequential structural transitions of the virion during the process of cell entry. Unexpectedly, VP1u exposure did not facilitate the
ex-ternalization of the viral genome in response to increasing temperatures (Fig. 7). On the contrary, receptor-bound capsids were remarkably more resistant than native or detached cap-sids. Increased capsid rigidity might assist incoming viruses during entry. In our previous studies, we showed that the B19V genome, in contrast to other parvovirus genomes, can easily be externalized upon mild acidification similar to that found in endosomes/lysosomes (29). Therefore, the natural resistance of B19V capsids would apparently be insufficient to prevent the externalization of the viral DNA in the acidic endosomal en-vironment. Our results indicate that following binding to glo-boside, alone or in concert with other molecular structures at the cell surface, B19V acquires a remarkably superior stability. The increased resistance, in contrast to VP1u exposure, was reversible upon receptor detachment (Fig. 7). In this way, B19V capsids would become more resistant when needed but then become flexible again upon receptor dissociation and endosomal escape to facilitate uncoating.
[image:7.585.134.449.71.369.2]It is reasonable to expect that receptor-induced conforma-tional changes are necessary for subsequent interaction events leading to virus internalization. In line with this assumption is the fact that VP2-only capsids, lacking VP1 (Fig. 8A), were unable to be internalized (Fig. 8B). Moreover, an antibody (MAb 1418-1; referred to as anti-N-VP1u) recognizing an epitope in the N-terminal part of VP1u (aa 32 to 40) (12)
FIG. 8. Binding and internalization of native virus and VLPs (with or without VP1). (A) SDS-PAGE of native virus, VLPs (VP1/VP2), and VP2-only particles. (B) UT7/Epo cells were incubated with the different viral particles for 1 h at 4°C, washed intensively, and further incubated at 37°C to allow virus internalization. At increasing times, the cells were washed, fixed, and stained with MAb 860-55D against intact capsids. Alternatively, after 30 min at 37°C, the cells were incubated with trypsin-EDTA to remove uninternalized capsids. Subsequently, the cells were washed, fixed, and stained with MAb 860-55D.
on November 8, 2019 by guest
http://jvi.asm.org/
blocked B19V infectivity and hampered virus internalization, but only when present during binding, not when added after virus binding (Fig. 9A and B). This result suggests that the externalized N-terminal part of VP1u is engaged in binding events required for internalization, preventing antibodies from binding due to ligand occupancy. Since anti-N-VP1u could only partially block internalization, the epitope recognized by the antibody (aa 30 to 42) might not be the region directly implicated in binding events. Growing evidence indicates that ␣51 integrin might be required for virus internalization (36, 37). The␣51 integrin binding motif RGD is present in the N-terminal part of VP1u regions from rhesus (aa 141 to 143) and simian (aa 139 to 141) erythroviruses and aligns with RGE and RGA motifs in these regions from B19V (aa 104 to 106) and pig-tailed erythrovirus (aa 106 to 108), respectively. While RGD is the classical ␣51 integrin binding motif, RGE and RGA motifs have also been found to interact specifically with
␣51 integrin (7, 19). Although the mere presence of RGD or RGD-like motifs does not warrant an interaction with integrin, the fact that these sequences are conserved in all members of the Erythrovirusgenus is rather suggestive. Interestingly, this region is adjacent to the epitope recognized by anti-N-VP1u (aa 30 to 42). It was previously shown that B19V infection was not inhibited by RGD-containing peptides (36), raising the possibility that B19V-integrin interaction differs from those involving classical RGD motifs. In order to elucidate this ques-tion, further experiments are in progress to examine binding of native and mutated VP1u domains with integrin subunits.
[image:8.585.92.236.67.417.2]Previous studies demonstrated that B19V does not bind to isolated Gb4Cer (20), suggesting that B19V binds Gb4Cer at the cell surface as part of a complex involving other structures. However, incubation of B19V particles with increasing concen-trations of purified Gb4Cer triggered changes in the original inaccessible configuration of VP1u, which became increasingly accessible on the capsid surface (Fig. 6). A possible explana-tion for our results is that binding of B19V to isolated Gb4Cer is highly unstable due to the lack of required stabilizing core-ceptors. This might also explain the detachment of a large proportion of cell-bound particles observed in this study (Fig. 3). Interestingly, reattachment of receptor-dissociated B19V to the receptor from which it detached was not detectable at 4°C and occurred only after several hours at 37°C, probably with newly expressed and/or recycled receptors (Fig. 3B). In con-trast, when the detached virus was added to uninfected cells, these particles bound with a higher efficiency and were more
[image:8.585.299.542.68.314.2]FIG. 9. Role of the N-terminal part of VP1u in virus internaliza-tion. (A) Neutralization activity of two MAbs against B19V. One MAb recognizes intact capsids (MAb 860-55D;␣-Caps), and the other rec-ognizes an epitope in the N-terminal part of VP1u (MAb 1418-1; ␣-N-VP1u). The antibodies were added during (preattachment) or after (postattachment) virus binding or after virus internalization (postinternalization). Virus infectivity was examined by quantification of NS1 mRNA at 24 h postinfection. (B) Capacity of B19V to be internalized in the presence of anti-N-VP1u added after virus binding (postattachment) or during virus binding (preattachment).
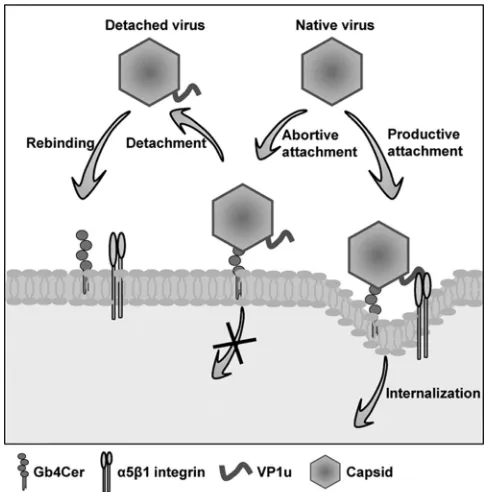
FIG. 10. Schematic representation of the proposed mechanism of B19V binding and internalization in UT7/Epo cells. B19V binds to the Gb4Cer receptor. The binding triggers the exposure of VP1u, which interacts with the coreceptor. However, whenever the interaction with the coreceptor is not possible, the virus detaches from Gb4Cer while keeping VP1u accessible at the surface. Cycles of binding and detach-ment are repeated until the interaction with the coreceptor occurs, after which the virus is internalized.
on November 8, 2019 by guest
http://jvi.asm.org/
infectious than the native virus (Fig. 4). This observation might represent a strategy to avoid repetitive “abortive” attachments to the same receptor and in this way increase the probability of productive bindings leading to virus internalization.
An understanding of the events preceding virus internalization is critical for establishing strategies for the prevention and treat-ment of B19V infections. Our results suggest a complex internal-ization mechanism involving various steps in a sequential manner, as illustrated in Fig. 10. B19V binds initially to Gb4Cer, which triggers changes in the VP1u conformation. The structurally al-tered capsid is ready for a subsequent interaction, which involves binding of the N-terminal part of VP1u with the coreceptor. However, in situations where interaction with the coreceptor is not possible, the virus detaches from Gb4Cer. The procedure is repeated until the second interaction is possible, and virus inter-nalization ensues as a consequence. The mechanism of “detach-ment-reattachment” would enhance the infection by minimizing the consequences of abortive attachments that do not lead to virus internalization.
ACKNOWLEDGMENTS
We are grateful to S. Modrow (Regensburg, Germany) for kindly pro-viding MAb 1418-1 against the N-terminal region of VP1u and MAb 860-55D against intact capsids and to J. de Jong (Netherlands Red Cross, Amsterdam, Netherlands) for providing the globoside-specific IgM monoclonal antibody (AME-2). We gratefully acknowledge the gifts of baculovirus-derived B19V capsids from R. Franssila (Helsinki, Finland) and of VP2-only particles from G. Elliott (Dublin, Ireland).
REFERENCES
1.Agbandje, M., S. Kajigaya, R. McKenna, N. S. Young, and M. G. Rossmann.
1994. The structure of human parvovirus B19 at 8 A resolution. Virology
203:106–115.
2.Anderson, S., M. Momoeda, M. Kawase, S. Kajigaya, and N. S. Young.1995. Peptides derived from the unique region of B19 parvovirus minor capsid
protein elicit neutralizing antibodies in rabbits. Virology206:626–632.
3.Bo¨nsch, C., C. Kempf, and C. Ros.2008. Interaction of parvovirus B19 with human erythrocytes alters virus structure and cell membrane integrity. J.
Vi-rol.82:11784–11791.
4.Broliden, K., T. Tolfvenstam, and O. Norbeck.2006. Clinical aspects of
parvovirus B19 infection. J. Intern. Med.260:285–304.
5.Brown, K. E., S. M. Anderson, and N. S. Young.1993. Erythrocyte P antigen:
cellular receptor for B19 parvovirus. Science262:114–117.
6.Brown, K. E., J. R. Hibbs, G. Gallinella, S. M. Anderson, E. D. Lehman, P. McCarthy, and N. S. Young.1994. Resistance to parvovirus B19 infection due to lack of virus receptor (erythrocyte P antigen). N. Engl. J. Med.
330:1192–1196.
7.Chillon, M., and E. J. Kremer.2001. Trafficking and propagation of canine adenovirus vectors lacking a known integrin-interacting motif. Hum. Gene
Ther.12:1815–1823.
8.Chipman, P. R., M. Agbandje-McKenna, S. Kajigaya, K. E. Brown, N. S. Young, T. S. Baker, and M. G. Rossmann.1996. Cryo-electron microscopy studies of empty capsids of human parvovirus B19 complexed with its cellular
receptor. Proc. Natl. Acad. Sci. U. S. A.93:7502–7506.
9.Cotmore, S. F., V. C. McKie, L. J. Anderson, C. R. Astell, and P. Tattersall.
1986. Identification of the major structural and nonstructural proteins en-coded by human parvovirus B19 and mapping of their genes by procaryotic
expression of isolated genomic fragments. J. Virol.60:548–557.
10.Cotmore, S. F., A. M. D’abramo, Jr., C. M. Ticknor, and P. Tattersall.1999. Controlled conformational transitions in the MVM virion expose the VP1
N-terminus and viral genome without particle disassembly. Virology254:169–181.
11.Cotmore, S. F., S. Hafenstein, and P. Tattersall.2010. Depletion of virion-associated divalent cations induces parvovirus minute virus of mice to eject
its genome in a 3⬘-to-5⬘direction from an otherwise intact viral particle.
J. Virol.84:1945–1956.
12.Dorsch, S., B. Kaufmann, U. Schaible, E. Prohaska, H. Wolf, and S.
Mod-row.2001. The VP1-unique region of parvovirus B19: amino acid variability
and antigenic stability. J. Gen. Virol.82:191–199.
13.Farr, G. A., L. G. Zhang, and P. Tattersall.2005. Parvoviral virions deploy a capsid-tethered lipolytic enzyme to breach the endosomal membrane
dur-ing cell entry. Proc. Natl. Acad. Sci. U. S. A.102:17148–17153.
14. Reference deleted.
15.Franssila, R., K. Hokynar, and K. Hedman.2001. T helper cell-mediated in vitro responses of recently and remotely infected subjects to a candidate
recom-binant vaccine for human parvovirus B19. J. Infect. Dis.183:805–809.
16.Gigler, A., S. Dorsch, A. Hemauer, C. Williams, S. Kim, N. S. Young, S. Zolla-Pazner, H. Wolf, M. K. Gorny, and S. Modrow.1999. Generation of neutralizing human monoclonal antibodies against parvovirus B19 proteins.
J. Virol.73:1974–1979.
17.Girod, A., C. E. Wobus, Z. Za´dori, M. Ried, K. Leike, P. Tijssen, J. A. Kleinschmidt, and M. Hallek. 2002. The VP1 capsid protein of adeno-associated virus type 2 is carrying a phospholipase A2 domain required for
virus infectivity. J. Gen. Virol.83:973–978.
18.Harbison, C. E., J. A. Chiorini, and C. R. Parrish.2008. The parvovirus capsid odyssey: from the cell surface to the nucleus. Trends Microbiol.
16:208–214.
19.Humtsoe, J. O., R. A. Bowling, Jr., S. Feng, and K. K. Wary.2005. Murine lipid phosphate phosphohydrolase-3 acts as a cell-associated integrin ligand.
Biochem. Biophys. Res. Commun.335:906–919.
20.Kaufmann, B., U. Baxa, P. R. Chipman, M. G. Rossmann, S. Modrow, and R. Seckler.2005. Parvovirus B19 does not bind to membrane-associated
globoside in vitro. Virology332:189–198.
21.Kronenberg, S., B. Bottcher, C. W. von der Lieth, S. Bleker, and J. A. Kleinschmidt.2005. A conformational change in the adeno-associated virus type 2 capsid leads to the exposure of hidden VP1 N termini. J. Virol.
79:5296–5303.
22.Levy, H. C., V. D. Bowman, L. Govindasamy, R. McKenna, K. Nash, K. Warrington, W. Chen, N. Muzyczka, X. Yan, T. S. Baker, and M. Agbandje-McKenna.2009. Heparin binding induces conformational changes in
adeno-associated virus serotype 2. J. Struct. Biol.165:146–156.
23.Mani, B., C. Baltzer, N. Valle, J. M. Almendral, C. Kempf, and C. Ros.2006. Low pH-dependent endosomal processing of the incoming parvovirus minute virus of mice virion leads to externalization of the VP1 N-terminal sequence (N-VP1), N-VP2 cleavage, and uncoating of the full-length
ge-nome. J. Virol.80:1015–1024.
24.Mani, B., M. Gerber, P. Lieby, N. Boschetti, C. Kempf, and C. Ros.2007. Molecular mechanism underlying B19 virus inactivation and comparison to
other parvoviruses. Transfusion47:1765–1774.
25.Munakata, Y., T. Saito-Ito, K. Kumura-Ishii, J. Huang, T. Kodera, T. Ishii, Y. Hirabayashi, Y. Koyanagi, and T. Sasaki.2005. Ku80 autoantigen as a
cellular coreceptor for human parvovirus B19 infection. Blood106:3449–
3456.
26.Ozawa, K., and N. Young.1987. Characterization of capsid and noncapsid proteins of B19 parvovirus propagated in human erythroid bone marrow cell
cultures. J. Virol.61:2627–2630.
27.Ros, C., C. Burckhardt, and C. Kempf. 2002. Cytoplasmic trafficking of MVM. Low pH requirement, routing to late endosomes and proteasome
interaction. J. Virol.76:12634–12645.
28.Ros, C., and C. Kempf.2004. The ubiquitin-proteasome machinery is essen-tial for nuclear translocation of incoming minute virus of mice. Virology
324:350–360.
29.Ros, C., C. Baltzer, B. Mani, and C. Kempf.2006. Parvovirus uncoating in vitro reveals a mechanism of DNA release without capsid disassembly and
striking differences in encapsidated DNA stability. Virology345:137–147.
30.Ros, C., M. Gerber, and C. Kempf.2006. Conformational changes in the VP1-unique region of native human parvovirus B19V lead to exposure of internal sequences playing a role in virus neutralization and infectivity.
J. Virol.80:12017–12024.
31.Saikawa, T., S. Anderson, M. Momoeda, S. Kajigaya, and N. S. Young.1993. Neutralizing linear epitopes of B19 parvovirus cluster in the VP1 unique and
VP1-VP2 junction regions. J. Virol.67:3004–3009.
32.Sonntag, F., S. Bleker, B. Leuchs, R. Fischer, and J. A. Kleinschmidt.
2006. Adeno-associated virus type 2 capsids with externalized VP1/VP2 trafficking domains are generated prior to passage through the cytoplasm
and are maintained until uncoating occurs in the nucleus. J. Virol.80:
11040–11054.
33.Vihinen-Ranta, M., D. Wang, W. S. Weichert, and C. R. Parrish.2002. The VP1 N-terminal sequence of canine parvovirus affects nuclear transport of
capsids and efficient cell infection. J. Virol.76:1884–1891.
34.Von dem Borne, A. E., M. J. Bos, N. Joustra-Maas, J. F. Tromp, M. B. van’t Veer, R. van Wijngaarden-du Bois, and P. A. Tetteroo.1986. A murine monoclonal IgM antibody specific for blood group P antigen (globoside).
Br. J. Haematol.63:35–46.
35.Weigel-Kelley, K. A., M. C. Yoder, and A. Srivastava.2001. Recombinant human parvovirus B19 vectors: erythrocyte P antigen is necessary but not sufficient for successful transduction of human hematopoietic cells. J. Virol.
75:4110–4116.
36.Weigel-Kelley, K. A., M. C. Yoder, and A. Srivastava.2003. Alpha5beta1 inte-grin as a cellular coreceptor for human parvovirus B19: requirement of
func-tional activation of beta1 integrin for viral entry. Blood102:3927–3933.
37.Weigel-Kelley, K. A., M. C. Yoder, L. Chen, and A. Srivastava.2006. Role of integrin cross-regulation in parvovirus B19 targeting. Hum. Gene Ther.
17:909–920.
on November 8, 2019 by guest
http://jvi.asm.org/
38.Weigel-Van Aken, K. A.2009. Pharmacological activation of guanine nucleotide exchange factors for the small GTPase Rap1 recruits high-affinity beta1 integrins as coreceptors for parvovirus B19: improved ex vivo gene transfer to human
erythroid progenitor cells. Hum. Gene Ther.20:1665–1678.
39.Wong, S., N. Zhi, C. Filippone, K. Keyvanfar, S. Kajigaya, K. E. Brown, and N. S. Young.2008. Ex vivo-generated CD36⫹erythroid progenitors are highly
permissive to human parvovirus B19 replication. J. Virol.82:2470–2476.
40.Zadori, Z., J. Szelei, M. C. Lacoste, Y. Li, S. Gariepy, P. Raymond, M. Allaire, I. R. Nabi, and P. Tijssen.2001. A viral phospholipase A2 is required
for parvovirus infectivity. Dev. Cell1:291–302.
41.Zuffi, E., E. Manaresi, G. Gallinella, G. A. Gentilomi, S. Venturoli, M. Zerbini, and M. Musiani. 2001. Identification of an immunodominant peptide in the parvovirus B19 VP1 unique region able to elicit a
long-lasting immune response in humans. Viral Immunol.14:151–158.