JOURNALOFVIROLOGY,JUly1970, p. 94-99
Copyright @ 1970 AmericanSocietyforMicrobiology
Vol. 6, No. 1 Printed in U.S.A.
Isolation and Characterization of
a
Bacteriophage
for
Vibrio fetus
JOHN H. BRYNER, ALFRED E. RITCHIE, JOHN W. FOLEY, AND DAVID T. BERMAN
National AnimalDisease Laboratory, U.S. Department of Agriculture, Ames, Iowa50010, and Department of
Veterinary Sciences, University of Wisconsin, Madison, Wisconsin 53706
Receivedforpublication 3 March 1970
Bacteriophages were isolated from 22 of 38 strains of Vibriofetus by an
enrich-mentprocess, utilizing the donor and host strains growing together in fluid thio-glycollate medium. One phage, V-45, isolated by the conventional lawn-spot
method, was characterized by stability in broth, growthkinetics, and morphology.
It was sensitive to rapid thermal inactivation, chloroform, andpH values above
6.5.Calcium wasrequired for phage replication and stabilityinbroth. Magnesium
provided the best protection against thermal inactivation at 50C in thepHrange
of 6.5 to7.5. The minimumlatent periodwas 135 min, rise time was 75min, and
average burst sizewas 35 plaque-forming units per infected cell. Phage V-45
re-sembled Bradley's morphological group B, having a long tail without contractile
sheath. Dimensions were: head, about 50 nm; tail, about 7 by 240 nm; and tail
lumen, 2to 3 nm.
The detection of
bacteriophages
for Vibriofetus until recently was not
successful, largely
becauseofdifficultiesin
propagating
theorganism
and in adapting
phage-investigation techniques.
The need for actively
growing,
sensitive host bacteria for phagereplication
iswell known(1).
Manclark and Pickett
(10)
describedamethodforgrowing V.fetus in liquid culture medium on a
gyratory shaker which satisfies this need.
Fire-hammerand Border
(6)
firstisolatedphage
fromold broth cultures of V.
fetus.
Their method ofcross-culture
spotting
onagaryielded
additional phage from this source.They
describedplaque
types,
lytic
range, andmorphology
of some ofthese isolates and
pointed
out thedifficulty
ofobtaining
phage
by
induction withmitomycin
C.This report describes technical
improvements
in the
isolation,
preservation,
and assay of V.fetus phages, and the characterization of one representative isolate, V-45.
MATERIALS AND METHODS
Media.Fluidthioglycollatemedium(BBL,01-140) wasusedfor maintenance and enrichment cultures of
lysogenicstrainsofV.fetus. Broth for shaker cultures
andphagedilution blanks consisted of Brucella broth
(Albimi:A-114) with 3 gof sodium succinate and 0.1 g
of cysteine per liter (pH 6.9), hereafter designated
modified Albimi broth (MA broth). Brucella agar
(Albimi Laboratories, Inc. A-115) was the nutrient
base for double agar-layer cultures, and thesoft top
layer contained, per liter ofwater: agar (Colab
Ion-agar no. 2), 5 g; NH4Cl, 1 g; KH2PO4, 6.5 g;
Na2HPO4, 3.5 g;sodium succinate, 3 g; andglucose,
1 g. The agar cultures were incubated in a gas mixture
of25% air, 10% C02, and65%N2, whereasliquid
cultureswereincubatedinair (5).
Bacterial cultures. Thirty-eight strains of V. fetus from our stock culture collection were tested, including 30strainsoftype 1 and 8 strains of type 2 identified
previously (5). Type 2strains 436 and 2255 were the
bacterialhostsforprimaryphageisolation, and 2255
was the indicator host for phage assays. Log-phase
host cells, grown in nepheloculture flasks on a shaker as described by Keeler et al. (8), contained 5 X 108
colony-forming units when used in preparing plaque
assaylawns.
Phage isolation. Phage V-45/436 (V45) was iso-lated from lysogenic strain 45 on host 436 by the
lawn-spot method of Matsushiro (11). Attempts to
isolate other phages by conventional methods were
unsuccessful.
The following enrichment method was developed
forisolating phagefromlysogenicstrainsof V.fetus.
Each teststrain was inoculated into separate culture tubescontaining 9 ml of freshthioglycollate medium, and1mlofhost2255culturewasadded to each tube. Themixedcultureswereincubated for3daysat36C
and thencentrifuged at2,500 X gfor 20min to
re-movecellsfrom unadsorbed phage in the supernatant fluid. This fluid was transferred to separate 100-mi
flaskscontaining5 X 109log-phasehostcells in20ml of MA broth and incubated for 5 hr.Samples (l-ml)
fromeachflaskwereplatedinoverlayagarto testfor
plaqueformation.
Characterization ofphage V-45. After three single-94
on November 11, 2019 by guest
http://jvi.asm.org/
plaquepurifications ofthe clearplaqueform ofphage
V45, itwasreplicated insoft-agarlawnsof host2255
to atiterof1010plaque-forming units(PFU)E/ml.The
resultinglysatewashomogenizedin three volumes of
MAbroth, sedimented, andfilteredthrough0.45 g&m membrane filters (Millipore Corp. Bedford, Mass.).
This phagestock waspreservedat4 C and used inthe
followingtests.
Thephagestock wasdiluted 10-1 in tubesofMA broth and tested for sensitivity to heat, chloroform,
andvarious pH values bythemethodof McDuffetal.
(12).
Similar tests were performed to assay for phage
activation, inactivation, or protection by using MA
brothcontaining variousamountsofCaC12, MgSO4, tryptophan, cysteine, NH4-benzoate, or phosphates
(1:4ratio ofNa2HPO4andKH2PO4).Thesetest
sus-pensionswereincubated for 18 to 22 hrat36Cand
assayed for PFU/ml.
Therequirement for calciumas acofactorforphage
replication (7) was tested, by using 180 ml ofMA
brothadjustedtopH6.5 with lacticacid,induplicate
culture flasks, onecontaining CaCl2 (0.01 M) andthe
othercontainingnone. Approximately1010 host cells
infected with phageV45 at amultiplicity of0.01 were
inoculated into each flask, incubated at 36 C on a
shaker, andassayed forPFU andchanges inoptical
density(ODin0) for7hr.
Growthkinetics of phageV-45weredetermined by
methodsdescribedbyAdams (1), withexceptionsas
noted. Thephage-celladsorptionrate wasbased on the number of infected bacteria sedimented after 5-min
incubationperiods.Inthesingle-step growthtest, the
phage tocellratiowas0.06, adsorption timewas 15 min at36 C,andserumneutralization of unadsorbed
phage was10min. After appropriate dilution ofthe
infectedbacteria,thefirst growthtube and the second
growth tube consisted of100 mlofMA broth with
CaCl2 (0.01 M) in 250-ml flasks. These tubes were
assayed forPFUat5-to15-minintervalsduring7 hr
at36 C withcontinuous shaking.
Forelectronmicroscopy, phageand host cellsfrom
abroth culturewereconcentratedbycentrifugationat
30,000 X gfor30min, after stabilization with
glu-taraldehyde (0.25%, 15 min), andnegatively stained
withneutralizedphosphotungstic acidbyusing minor
modifications (14) of the Brenner and Home (4)
method. Grids wereexamined in aPhillips EM-200
electron microscope operated at 60 kv with
double-condenserilluminationand a 20- to25-,umaperture.
RESULTS
Phage isolation. Phages were
isolated
from 22of 38 V. fetus strains by
enrichment
culture ofphage from
lysogenic
strains with host 2255 insemisolid thioglycollate medium. Thenumber of
plaques
observed
atprimaryplating varied from15 in some strains to almost complete lysis in
other strains(average,250). All 22 phage isolates
were derived from V. fetus type 1 strains. This
type generally grew
poorly
in soft-agar overlay [image:2.487.244.437.69.186.2]cultures, whereas type 2 strains produced dense
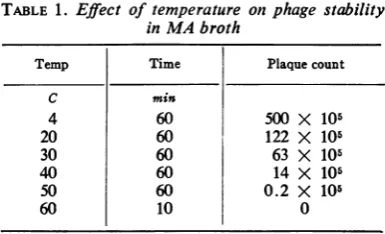
TABLE 1. Effect of temperature
inMAbroth
on phage stability
Temp Time Plaque count
C min
4 60 500 X 105
20 60 122 X 105
30 60 63 X 105
40 60 14 X 105
50 60 0.2 X 105
60 10 0
lawns, and the latter were more adaptable as
hosts.
Thelawn-spot methodyielded onlyonephage,
V-45, from various combinations of V.
fetus
strainsspottedonsoft-agar lawns. Althoughlysis
was often observed in thespots,phageswerenot
recovered from filtrates of thelysed zones.
Characterization
of phage V-45. The crude lysatefromphageV-45 onhost2255 in soft-agarcultures contained approximately 1010 PFU/ml.
After low-speed centrifugation, the supernatant
fluid contained 109 PFU/ml, and, after filtration
through
0.45-,Am filters, the final titerwas 5 x 108 PFU/ml.Attempts to extract
phage
from host cell lysates with 10% chloroform at 4 Cresulted inalmost complete inactivation (survivors, 0.008).
Oneper centchloroformin MAbroth inactivated
70% ofphage in 1 hr at 36 C.
About 76% of phage in MA broth was inac-tivated
by
incubation at20 C for 1 hr.Propor-tionately greater inactivation was observed at
temperatures from 30 to 50 C, and all phage
activitywasdestroyed in 10 minat60 C(Table1).
In tests for phage stability at various pH
values, the
optimum
was aboutpH 5.9, with arapid drop in titer belowpH 5 and abovepH 7
(Fig. 1).
Theaddition ofCaCl2toMAbrothsuspensions of
phage
V45 contributedsignificantly
to itsstability (Table 2).
One effect ofadding CaCl2
wasthe decrease in pH from 7.2to6.7. However, the addition ofphosphate
salts to bufferbroth
atpH6.7,withoutCaCl2,
hadnoeffectonphage
stability.
When5 to7.5 mgofCaCl2
permlwasadded to broth containing 5 mg of phosphate.
saltsperml, the titer of
surviving
phage wasin-creased by a factor of103.
Addition
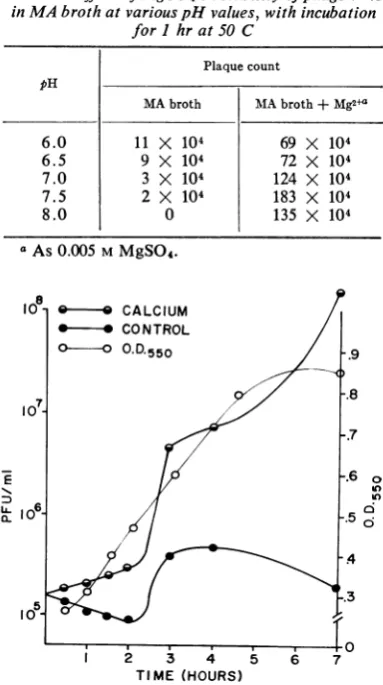
of MgSO4 to
MA broth wasbeneficialfor phage stability at 50 C, the water bath
tem-perature of thetop agar (Table 3). The
optimum
pHwith0.005 MMgSO4 was 7.5.
Tryptophan or cysteine in concentrations of
0.2 to1 mg/mlhad no effect on the phagetiter.
However, 2.5 to 5 mg/ml ofcysteine completely
on November 11, 2019 by guest
http://jvi.asm.org/
BRYNER ET AL.
l0
.
E
3-
40 5 8 9 1
a-I0
5.9
3 4 10
pH
FIG. 1. InfluenceofpHonphagetiter.TubesofMA
brothadjustedtovariouspHvalues from 3to 10 with INHCIorNaOH,containingS X 107 PFU ofphage
V-45/ml,wereincubated 24hrat36 Candassayed.
TABLE2. Effects ofCaCl2, phosphates, andpHon
phage stability inMAbroth with incubationfor 24 hrat36 C
Additiontobroth
(mg/ml) EndpH Plaquecount
CaC12 po4a
0 0 7.2 0.2X 10'
1 0 7.2 2.4X 10'
5 0 6.8 24.1 X 106
10 0 6.7 26.6 X 105
0 1 6.8 0.7 X 10'
0 5 6.7 0.5X 10'
0 10 6.4 0.4 X 10'
0 5 6.7 0.5X 105
0.1 5 6.7 0.5 X 10'
1.0 5 6.5 1.6 X 105
2.5 5 5.9 16.7 X 105
5.0 5 5.6 68.7 X 10'
7.5 5 5.4 107.0 X 10'
aP04 consisted of
parts KH2PO4.
one part Na2HPO4 to four
inactivatedthephage,sincethepHdroppedbelow
4.8.
Preparation of phage for electron microscopy
by washing in water with NH4-benzoate (2.5 to
10 mg/ml) inactivated most of the phage present. Calcium was required for phage replication in
broth cultures of the host. The titer of phage
increased by 103 PFU/ml in 7 hr with CaC12, but decreased in the absence of CaCI2 (Fig. 2).
Culture turbidity throughout the growth period
increased at the same rate in both flasks, and evidence of clearing resulting from phage lysis of
host cells was not observed ineither flask.
Antiphage serum, diluted 1:50, neutralized
90% of the phage in 10min, and the
neutraliza-tion K value was 23.
kwThe
phage-cell adsorption rate was88%
in 15 min; the adsorption K value was 5.55 X 10-11 [image:3.487.56.245.50.325.2]ml/min.
TABLE 3.Effect ofMgSO4onstability ofphage V45
inMA brothatvariouspH values, withincubation forI hrat50 C
Plaque count pH
MAbroth MAbroth + Mg2+a
6.0 11 X 104 69 X 104
6.5 9 X 104 72 X 104
7.0 3 X 104 124 X 104
7.5 2 X 104 183 X 104
8.0 0 135 X 104
aAs
0.005
MMgSO4.
0L
0
In
a .
1 2 3 4 5 6 7
TIME (HOURS)
FIG.2. Comparison of phage V-45 replication in
shaker culturesofhost2255 with and withoutCaC12.
Theopticaldensitycurve(light line),superimposedover
thePFUcurves (heavy lines), was the sameforboth
flasks.
96
J.VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:3.487.259.451.252.595.2] [image:3.487.54.247.372.606.2]U
FIG. 3. Vibrio fetusphage V-45inpotassiumphosphotungstate negativestainafterglutaraldehyde stabilization.
(A) Cellfragmentillustratingmultiplicity of receptorsitesandpredominance of "empty" heads. X 180,000. (B)
Typicalphage with "empty"
hlead,
nonsheathedtail,and ill-defined baseplate (B)associated with aknobofrecep-torsubstance (R). X 400,000. (C) Atypicalphage illustratinghexagonalsymmetry ofitspartiallyempty head membraneandsphericalsymmetry ofitsinternal residuum. X400,000. Each bar represents 100 nm.
on November 11, 2019 by guest
http://jvi.asm.org/
[image:4.487.40.440.61.556.2]BRYNER ET AL.
In the single-step growth test, the minimum
latent period was 135 min, therise time was 75
min, and the average burst size was 34 PFU/
infectedcell.
In electron micrographs, phage were usually
attached to cells or cell fragments (Fig. 3A) and
the head was "empty" regardless of whether it
had been stabilized with glutaraldehyde.
Mor-phologically, this phage resembled members of
Bradley's group B; i.e., it lacked a contractile
protein sheath on the tail (Fig. 3B). Average
dimensions of the phage werehead, ca. 50 nm,
and tail, ca. 7 by240 nm. When discernible, the
tail appeared to have a 2- to 3-nm lumen and
some form of a base plate (Fig. 3B). Free-lying phage, especially after glutaraldehyde
stabiliza-tion, often had avariable-sized knob or disc at
the tail tip (Fig. 3B, C); these were probably
receptors since they were located at a fixed
distance (ca. 10 nm) belowremnantsofthe base plate ofthe tail.
DISCUSSION
Conventional methodsofphage isolationfrom
lysogenic
bacteria were ineffective when appliedto V. fetus. Characterization of phage V-45 in
this study has permitted the development of improvements in the techniques of isolation, preservation, andassay ofadditional phagesfrom
strains of V. fetus.
Phage isolation from V. fetus was greatly enhanced by the liquid culture of lysogenic strains in the presence of host cells. Growth of
lysogenic
strains was improved by cultivation inthioglycollate medium; the growth time was
extended, resulting in a greater probability of
spontaneous inductionofalytic cycleof virulent
phage; sensitive host cells were continuously
present with the lysogenic cellstoreplicate
viru-lentphagetoa
higher
titer than couldbeobtained with the. "immune" lysogenic cells alone. The extended growth period was also important because of the slow phage-cell adsorption rateand relatively long replication cycle in the V.
fetus-phage system. The importance of mixing
the host with thelysogenic strain was shown by
thefact that no plaqueswereproducedbyeither
culture when grown alone.
Phage extractionwith chloroform is not
prac-tical, asshownbythehigh inactivation rate in as
little as 1% chloroform.
The finding that low pH enhanced phage stability led to testing ofphosphate salts in MA
broth for plaque assaydilutions, with little
bene-fit. However, addition ofCaCl2 increased phage
stability byafactorof102, andthere wasalinear
relationship between the amount of
CaCl2
andplaque counts. Furthermore, when both
CaCl2
andphosphate salts were added, the phage titer
wasincreased103,indicatingthat pH isimportant
in phage stability. Work with coliphage (1)
indicates that phosphates accelerate the rate of
heat inactivation of phage T5, whereas CaCl2
reverses the process by forming a precipitatewith
phosphates.
Although the optimum pH for phage stability
inMAbrothwas 5.9, the greatest stabilityin the
presence ofCaCl2was achieved at pH 6.5. Only
in thepresence of MgSO4 were phages stable at
pH 7.5. Althoughtests withMgSO4 utilized only
the 50 C temperature,the datain Table2indicate
thatphageV-45 should berelatively stable over
thepH range of 6 to 8 in MA broth with 0.005
MMgSO4.
Inadditiontoitsstabilizingeffect on freephage in broth, CaCl2 (probably
Ca2+)
also functionsas an essential cofactor for replication of phage
V-45inliquidcultures ofthe host.The mechanism
ofcalcium action probably involves the transfer
ofdeoxyribonucleic acid fromthephageinto the V. fetus cell as described by Lann (9) for the
Escherichia coli-T5 system. Other similarities
betweenT5 andV-45 arethe low serum neutral-izationKvalue, slowadsorptionrate, andgeneral
morphology.
The adsorption rate of V-45 was almost
identical to that reported by Firehammer and Border fora similarphage, but the latent period
waslessby45 min. Thelongerlatentperiod and
failure oftheir phage to replicate inthe second-growth tube
probably
was due to a calcium deficiency in their medium.Morphologically, phage V-45 is
indistinguish-ablefromphagesVfi-6 and Vfv-3 ofFirehammer
and Border (6) and should be included, with coliphage T5, in Bradleys morphological group
B (3). All three phages appeared to be
"empty-headed" in electron microscopic preparations,
perhapsbecause ofa
proclivity
toattachtoexcessreceptor sites (2, 15) available in thelysate, with
consequent loss ofnucleicacids. The three phages
shareacommontail
antigen
(A.
E.Ritchieetal., unpublished data), indicating a close genetic relationship between the twoprincipal
V. fetus biotypes, intestinalis and venerealis. Phageinfec-tion of V.fetus
rarely
leadstocompletedestruc-tion of the host cell, butinduces selective
altera-tion of their
enveloping membranes, revealing
anunderlying
hexagonal
pattern inthecellwall(13).
LITERATURE CITED
1. Adams,M. H.1959.Bacteriophages. Interscience Publishers, Inc.,NewYork.
2. Bradley, D. E. 1963. The structureofsomestaphylococcus and pseudomonas bacteriophages. J. Ultrastruct. Res. 8:552-565.
98 J.VIROL.
on November 11, 2019 by guest
http://jvi.asm.org/
3. Bradley D. E. 1967. Ultrastructure of bacteriophages and bacteriocins. Bacteriol.Rev.31:230-314.
4. Brenner, S., and R. W. Horne. 1959. Anegative staining methodfor high resolution electron microscopy of viruses. Biochim. Biophys. Acta. 34:103.
5.Bryner, J.H., A. H.Frank,and P. A.O'Berry. 1962. Disso-ciation studies ofvibrios from the bovine genital tract.
Amer.J. Vet. Res. 23:32-41.
6.Firehammer, B.D.,and M. Border. 1968. Isolation of tem-perate bacteriophages from Vibriofetus. Amer. J. Vet. Res. 29:2229-2235.
7. Kay, D. 1952. Theeffect of divalent metalsonthe multiplica-tion ofcolibacteriophage T5st. Brit. J. Exp. Pathol. 33:228-235.
8. Keeler, R. F., A. E. Ritchie, J. H. Bryner, and J. Elmore. 1956. The preparation and characterization of cell walls
aid the preparation offlagella of Vibrio fetus. J. Gen. Microbiol.43:439-454.
9. Lannie, Y. T. 1954. Infection by bacteriophage T5 and its intracellular growth-a study by complement fixation. J. Bacteriol. 67:640-650.
10. Manclark, C. R., and M. Pickett. 1960. Quantity production of Vibrio fetus cells. J. Bacteriol. 79:752-753.
11. Matsushiro, A. 1961. Isolation ofUV-inducible temperate
phage 80. Biken J.4:133-135.
12. McDuff, C. R., L. M. Jones, and J. B. Wilson. 1962. Charac-teristics of brucellaphage. J. Bacteriol. 83:324-329. 13. Ritchie, A. E., and J. H. Bryner. 1968.Astructuralelement
in theenvelopesystemof Vibriofetus,p.78-79. In C.J.
Ar-ceneaux (ed.), Proc. 26th Annu.Meet. ElectronMicrosc. Soc. Amer.Claitor'sPublishing Division,BatonRouge, La. 14. Ritchie, A. E., and A. L. Fernelius. 1969. Characterization of bovine diarrhea viruses. Arch. Gesamte Virusforsch. 28:369-389.
15. Weidel,W.,andE.Kellenberger. 1955. The E. coliB
recep-torfor15.II.Electronmicroscopic studies. Biochim
Bio-phys. Acta 17:1-9.