Copyright01975 American Society for Microbiology Printed in U.S.A.
Occurrence of the
Bacteriophage Lambda Receptor
in
Some
Enterobacteriaceae
MAXIME SCHWARTZ* ANDLtON
LE MINORUnite deGenetique Moleculaire and Service des Enterobacteries, Institut Pasteur, Paris, France Received forpublication 25 November 1974
In
Escherichia coli K-12, the
receptorforphage lambda is
anoutermembrane
protein which inactivates the phage
in vitro.Lambda
receptoractivitywasfoundin extracts from
all
wild strains of E. coli tested, although mostofthem failtosupport
growth
of thephage.
Insome casesthis
failure isdue
toamasking of the receptorinvivo, the bacteria being unable
toadsorb the
phage orto reactwithantireceptor antibodies.
Inother
cases,adsorption
doesoccur,and
thenatureofthe block in
phage growth
wasnotinvestigated.
Most Mal+ strains of Shigellahave
lambda
receptor,whereasmostMal
-strains donothave it.Synthesis of thelambda
receptor inShigella
is thuspresumably controlled by
the positiveregulator
geneof the maltose regulon
asisthe
caseinE.coli
K-12.Phage
lambdaadsorbs on many Mal+ strains of
Shigella and
evenyields plaques
on some ofthem, althoughatalowfrequency. No lambdareceptoractivity could be found in extractsof
several strains
ofSalmonella and
Levinea.
Bacteriophage
lambda
wasoriginally isolated
from
aculture of Escherichia coli K-12, which
happened
tobe lysogenic
forthis
phage
(8).
Since
its
discovery,
phage lambda
wastusually
propagated
onnonlysogenic derivative of
E. coli
K-12
ormorerarely
inE.
coli C
(2),
instrains ofShigella
flexneri (5),
and in
hybrids
between E.
coli and
Shigella (5,
11)
orSalmonella strains
(1).
There
islittle information
inthe literature
on
the host range
ofthis
bacteriophage.
Wefound that
plaques of
phage lambda could be
obtained
ononly five
outof
70randomly
se-lected strains of E. coli. The
inability
of moststrains of
E.
coli
tosupport
growth
ofthe
phage
could
be due
to one or moreofseveral reasons,
the
mostobvious
being
that those strains do
notadsorb the phage,
perhaps because
they
lack the
specific receptor.
However,
possession
ofthe
receptor may
be
suspected
toconfer
aselective
advantage since this receptor has
been shown
tobe
aprotein of
the
outermembrane
(15)
in-volved in
the active transport
ofmaltose
(S.
Szmelcman and M.
Hofnung,
manuscript
inpreparation)
and in chemotaxis towards thissugar
(G. Hazelbauer,
J.Bacteriol.,
inpress).
We show in this
paper that lambda receptor
activity is
present
in extracts of most wild strainsof E.coli,
aswellasinextractsofseveralShigella
strains. In some of those strains thereceptor
isapparently
notaccessible
to thephage,
sothat the lack
ofphage
growth
canstill
be
attributed
to a lack ofadsorption.
In otherstrains
adsorption
ofthephage
doesoccur, andthe
block
musttherefore lie at a further step in
the infection
process.MATERIALS AND METHODS
Bacterial strains. All the strains used are listed in Table 1, except for E. coli K-12 CR63 used to plate phage XVh (15). HfrG6 is our standard wild-type, lambda-sensitive strain of E. coli K-12, while pop 1730 is a spontaneous lambda-resistant mutant of
HfrG6,
carryinga deletion (malBA17)encompassinglamB, the structural gene for the lambda receptor protein (7, 15). The other strains, except for E. coli ML30 (3), were chosenatrandom inthe collection of theService des Enterobacteries de l'Institut Pasteur. The E. coli 0 strains are reference strains for 0
serotypes. The other strains have been isolated from various sources,mainly from hospital patients. Char-acterizationofthe strainswasdoneaccording tothe classicaltechniques(4).
Phage strains. Phage XV isavirulent mutant of
lambda. Although this is probably not relevant for this work, it carries a b2 deletion. Phage XVh is a spontaneoushost rangemutantofXV isolated in this
laboratoryby plating XVonE. coli K-12CR63 which islambda resistant(15). XCI57is a b2+clearmutant ofX.
Media.
Shigella strains,
which areauxotrophic,
were grown in complete ML medium (1% tryptone [Difco], 0.5% yeast extract, 0.5% NaCl, pH 7.2, supplemented with 0.4% maltose). The other strains were grown in minimalmediumM63 (7, 15) supple-mented with 0.4% maltose, as wellas0.01% L-histi-dinein thecase ofHfrG6.To growpop 1730, which cannotutilizemaltose,0.4%glycerolwasaddedtothe above medium. The complete medium usedfor plat-679
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Lambdareceptoractivity incholate EDTA extractsofvarious enterobacteria
Ability to Protein concn in Receptoractiv- Receptoractivityasmeasured
Abil
nlity
to cholate EDTA itymeasured byinactivation
ofXVc
Bacterial strains utilize extracts byinactivation I rsneo nasneomaltosea (mg/ml)b of
XVhC
Inpresenceof Inabsenceof1 ~~~~20%,ethanol ethanol
Escherichia coliK-12 Hfr G6 + 0.80 +-+
E.coliK-12pop1730 _ 0.85 _
E.coliML30 + 1.20 +
E. coli13.73 + 0.50 +
E. coli14.72 + 0.30 +
E. coli18.72 + 0.40 + +
E. coli63.71 + 0.65 + + +
E. coli66.71 + +
E. coli67.71 + +
E. coli 16.72 + + + +
E.coli19.72 + +
E. coli02 + 0.40 + +
E.coli04 + 1.00 t
E.coli 06 + 0.35 + + +
E. coli 07 + 0.50 +
E. coli 010 + 0.20 + +
E.coli 018 + 0.30 + +
E. coli 028 + 0.15 +
E. coli075 + 0.55 + +
E.coli081 + 0.65 +
E.coli 083 + 0.85 +
Alkalescensdispar4.74 + 0.30 + + +
A.dispar25.73 + 0.30 + +
Shigella sonnei22.70 + 0.25 + +
S. sonnei 20.73 + 0.15 +
S. sonnei 25.73 + 0.20 +
S.sonnei3.74 + 0.10 +
S.sonnei 28.73 + 0.15 + +
S.sonnei 27.70 + 0.10 + +
S.sonnei28.70 + 0.10 + +
S.sonnei29.70 + 0.15 + +
S. sonnei30.70 + 0.20 + +
S.sonnei 46.70 + 0.21 +
Shigella dysenteriae R.16.72 + 0.45 +
S.d'ysenteriae8.39.74 + 0.35 +
S.dysenteriae1.30.74 0.50 _
S.dysenteriae2.31.74 0.25 _
S.dysenteriae4.32.74 0.30 _
S.dysenteriae7.33.74 0.10 _
S.dysenteriae8.34.74 - 0.35 _
Shigella flexneri1.29.73 0.20 _
S.flexneri2.5.74 _
S.flexneri1.8.68 0.45 _
S.flexneri2.8.70 - 0.35 +
S.flexneri6.15.66 - 0.10 _
S.flexneri3.16.70 + 0.25 +
S.flexneri4.28.74 + |+
S.flexneri1.15.73 _ 0.10 _
S.flexneri1.15.73 Mal+d + 0.25 +
S.flexneri2.21.73
S.flexneri2.21.73Mal+d + +
S.flexneri2.26.74
S.flexneri2.26.74 Mal+d + +
S.flexneri1.29.74
S.flexneri 1.29.74 Mal+d + +
S.flexneri3.27.74
on November 10, 2019 by guest
http://jvi.asm.org/
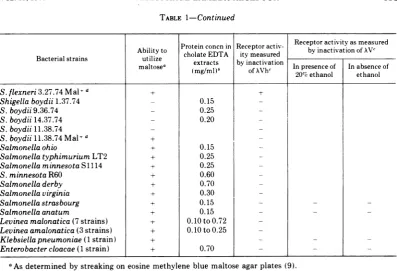
TABLE 1-Continued
..Proteinconcnin Receptoractiv- Receptor
activity
asmeasured [image:3.507.54.452.67.338.2]Ability to
tiliz
e Prti.occholateEDTA
nRcpoitymeasuredci by inactivation of XVc Bacterial strains utlze extractsby
inactivation|maltosea
maltosea(mg/ml)b(mg/ml)5 of XVhcofXVh~ 20/C
In presence ofethanol|Inabsence
ethanolofS.flexneri3.27.74Mal+d
Shigella boydii 1.37.74 - 0.15
-S.boydii9.36.74 - 0.25
-S.boydii14.37.74 - 0.20
-S.boydii11.38.74 _ _
S.boydii11.38.74Mal+d +
Salmonella ohio + 0.15
-SalmonellatyphimuriumLT2 + 0.25
-Salmonella minnesota S 1114 + 0.25
-S. minnesota R60 + 0.60
-Salmonelladerby + 0.70
-Salmonellavirginia + 0.30
-Salmonella strasbourg + 0.15 - _
Salmonella anatum + 0.15 - _
Levineamalonatica (7 strains) + 0.10to0.72
-Levinea amalonatica (3 strains) + 0.10to0.25
-Klebsiellapneumoniae (1 strain) + - _
Enterobacter cloacae (1 strain) + 0.70 _ _
aAsdeterminedbystreakingoneosine methylene blue maltose agar plates (9). ,Proteinconcentration wasdetermined according toLowryet al. (10).
c+, Extract contains at least
10'%
asmuch activity as does an extract of E. coli K-12 Hfr G6.dThisstrain isaspontaneous Mal+ mutant isolated from the Mal- strain listedon the preceding line. By growthin amaltose-containing medium suchMal+ mutants canbe obtainedfrom many of the Mal- Shigella strains.
ingphage had the following composition: 1% tryptone (Difco),0.25%NaCl, and 1% agar (Difco).
Extraction and assayof the phage lambda re-ceptor. Extraction ofthe receptor from whole cells with 1% sodium cholate and 2 x 10' M ethylenediaminetetraacetate (EDTA), as well as assay ofreceptor activity, weredescribed previously (15). The rate constantforphage XVh inactivation is proportionaltothe concentrationofreceptorpresent.
Preparation of antiserum directed against the lambda receptor. Partially purified receptor was preparedasdescribed (15) exceptforminor modifica-tionduetothe scalingupofthe procedure. Themain modification is the introduction of a precipitation with 66%ethanolbefore the chloroform-ethanol treat-ment.Protein (150 ug)fromthemostpurified fraction (representing about 100 jtg of receptor), emulsified with complete Freund adjuvant and 5
Mg
of methyl-ated bovine serumalbumin, was injected subcutane-ouslyinto arabbit. The injectionwasrepeatedonce a weekover 4 weeks. The fifthweek,300tig
ofproteins was injected intravenously and the serum was col-lected 10days later. Exhaustion ofthe serum from antibodiesreacting with E. colisurfaceantigensother than thereceptor wasdoneby incubatingthe serum with 2 volumes ofa pop 1730 suspension (10lbac-teria/ml) insaline (0.9% NaCl) for 1 h at 37 C and then 10h at4C,andby eliminationofthebacteriaby
centrifugation. Lambda receptoractivity isinhibited in extracts by this antiserum, but not by sera from
nonimmunized rabbits. Protection of the receptor from inactivation by the antiserum is provided by an extract fromCR63 (a lambda-resistant mutant with a point mutation in lamB) and not by an extract from pop1730(a strainwhose lamB gene is deleted). These results, which will be described in detail elsewhere, demonstratethe specificityofreceptor inactivation by the antiserum used inthiswork.
Immunofluorescentlabeling. The technique used consists in first reacting the bacteria with rabbit anti-lambda receptor antiserum prepared as de-scribed above and then adding fluorescein-labeled goatimmunoglobulins directed against rabbit
immu-noglobulins(13). Thistechniqueclearly differentiates between cells of HfrG6, which become labeled with fluorescent antibodies, and cells of pop 1730, which remain unlabeled. In control experiments, the anti-lambda receptor antiserumwasomitted andno label-ingwasobserved.
Electronmicroscopy. Exponentially growing cells werecentrifuged andresuspendedat a concentration of
10'
bacteria/ml in 10-2 M MgSO,. Purified XV phage was added at a multiplicity of 400 active particles per bacterium. After 15-min incubation at 37C, most oftheunadsorbedphagewasremoved bycentrifugation,and thecellswereresuspendedin102 M MgSO4 and fixed by adding 0.5% formaldehyde.
Thebacteriawerethendepositedoncarboned, Form-var-coatedgridswhich hadpreviously been submitted to a glow discharge. The preparations were stained
VOL. 681
on November 10, 2019 by guest
http://jvi.asm.org/
with 1%//. uranyl acetate and examined under a Sie-mensElmiskop 101 electron microscope.
RESULTS
Lambda receptor activity in extractsfrom various enterobacteria. Extracts of E. coli K-12 inactivate host range mutants of phage lambda, such as the XVh strain used in this
work (15). They also inactivate wild-type phage lambda if saturating amounts of chloroform
(15)or20%ethanol (M. Schwartz, unpublished
data) were present duringthe reaction. Inacti-vationofbothX and Xh isduetothepresencein
the extractsof lambdareceptor, aprotein from
theouter membrane. Asignificant activitywas
found in 21 randomly selected E. coli strains includingtwo strains from the
Alkalescens
dis-pargroup. An apparently less clearcutresult is
obtained with
Shigella,
since extracts fromsome strains have lambda receptor activity
whereas extracts from other strains do not.
Although all testedE. coli strains are Mal+, as are most strainsofthisspecies, thesame isnot true of Shigella strains. An almost perfect
correlation is observed between the ability of
Shigella strains to utilize maltose and the
presence in their extracts of lambda receptor
activity. Hence, 19Mal+strains outof 20make
an active lambdareceptor, and 18Mal- strains
outof 19 do not make it.
Furthermore
someofthe Mal+strainsofShigella listed in Table1are in fact Mal+ variants of strains which were
originally Mal- anddevoid of lambda receptor.
Such results regarding the Shigella strains
ex-tend the observation of Gemski et al. (5) and strongly suggest that, in Shigella as in E. coli
K-12, synthesis of the enzymes of maltose
metabolism andofthe lambdareceptorprotein is regulated by asingle positive regulatorgene,
which would be inactive in most naturally occurring Mal- strains. Extracts of several Mal+ strains of Salmonella and Levinea areall
devoid oflambdareceptoractivity.
The results in Table 2 demonstrate that the
activity found inextractsofE.coli andShigella
strains can be inhibited by an antiserum
di-rected against the lambda receptor ofE. coli
K-12. The activityinsuchextractsisthus likely
dueto aprotein similartothe lambdareceptor
ofE. coli K-12.
Ascan beseen inTable 1, inactivation ofXV
by extracts of several E. coli and Shigella
strains does not require thepresenceof
chloro-form or ethanol. Whether this results from a property ofthe lambda receptorprotein itself,
which wouldbeslightly different in such strains
from what it is in E. coli K-12, or from the
TABLE 2. Inhibitionoflam bdareceptoractivity from
variousstrainsby antiserumdirectedagainstthe lambdareceptorfrom E.coli K-12a
Untreated Extract treated extract withantiserum Bacterialstrains Pla- Pla- Pla-
Pla-quesat quesat quesat quesat
t =O t= 12 t=O t = 12
min min min min
Noextract 490 436 555 560
Escherichia coli K-12 250 53 548 515 Hfr G6
E.coli66.71 338 55 515 445
E.coli 67.71 179 29 502 354
E.coli 19.72 278 47 481 423
E.coli 13.73 283 46 488 294
Shigella sonnei 22.70 526 41 470 457
S.sonnei 3.74 560 40 432 357
aThe antiserum against lambda receptorwasused
at a 1/30 dilution in saline. E. coli extracts were diluted 100-fold, and S. sonnei extracts 10-fold, in 10-2 M Tris buffer, pH 7.5. These diluted extracts were incubated for 1 h at 37C with 100 ul ofsaline (untreated extracts) or with 100
pl
of the diluted antiserum (extract treated withantiserum). The vol-umes werethencompleted to 1 ml with 10-2 M Tris bufferand, at time (t) =0min, 1ml of a suspension ofXVh in 10-2 M MgSO4 was added. One-hundred-microlitersamples were withdrawnimmediately (t= 0min)and 12 min later (t = 12min) andaddedto100 uI ofindicatorbacteria (CR63)for
plating.
presence of some other component in the ex-tracts still remainsto befound.
Absence of receptor activity in extracts of strains other than E. coli and Mal+ Shigella could be due either to an actual absence of receptor in these strains, or to a lack of extrac-tion ofthe receptor by 1% sodium cholate and 2
X 10-3M EDTA. In
fact,
as seeninTable1,
the amount ofprotein
extracted variesdepending
on the strain. However, no obvious correlation was foundbetweenthe amount of
protein
found in the extracts and the presence orabsence
of receptoractivity.
Also,
even sonic extracts of two Salmonella strains and five Mal- Shigella strains, supplemented with 1% sodium-cholate and 2 x 10-3 M EDTA, were found totallydevoid of lambdareceptor
activity. (Extracts
of E. coli K-12 HfrG6 prepared in this way have the same activity as regular cholate EDTA extracts.)Consequently
it would seem that absence of receptor activity in cholate EDTA extracts of a strain really reflectsabsence
of active receptor in these strains, rather than a lack ofextractibility.Accessibility
oflambda
receptor in vivo.Several of the strains listed in Table 1, mainly
J.VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
those
which
wereshown above
tohave
receptor,were tested
fortheir ability
toadsorb
phagelambda.
As seen inTable
3,all
Shigella
sonnei
strains, as well as some ofthe E.
coli
strains,do
adsorb
XVh, as would be expected since theyhave receptor. Most of the E. coli strains, however, do not adsorb the phage, although
their
extracts contain receptoractivity.
Thisresult could
beexplained if, insuchstrains, thereceptor
was not accessible to the phage. Anindependent way of studying the accessibility of
the
receptor in vivo consists of testing its ability to react with antibodies. Contrary to whathappens with
antibodies directed againstsev-eral other
surface antigens, antibodies againstlambda receptor
do not induce an easilydetect-able clumping
of bacteria. Some differentialclumping
of sensitive versuslambda-TABLE 3. Adsorption of phage lambda and of anti-lambda receptor antibodies on someenterobacteria
Bacterial strains AdsorptionofXVhmeas- Adsorption ofXVdetected Antibodylabeling uredby centrifugationa byelectron
microscopyb
(immunofluorescence)c
Escherichia coli K-12 HfrG6 E.coli K-12 pop 1730
E. coli
ML30E. coli 13.73 E.coli 14.72 E.
coli
18.72E.coli 63.71 E.coli 66.71
E.coli 67.71 E.coli 16.72 E.
coli
19.72E.coli 02
E.
coli
04 E.coli
06 E.coli 07 E.coli010E. coli 018 E.coli 028 E.coli 075
E.
coli 081
E.
coli
083Alkalescensdispar4.74
A.dispar 25.73
Shigella
sonnei 22.70S.sonnei20.73
S.sonnei 25.73 S.sonnei 3.74 S.sonnei 28.73 S.sonnei27.70
S.sonnei28.70
S.sonnei 29.70 S.sonnei 30.70 S.sonnei 46.70
Shigella
dysenteriae
1.30.74Shigella
flexneri
1.15.73 Shigella boydii1.35.74 SalmonellatyphimuriumLT2 Salmonella ohioSalmonellaanatum
+
+
+ +
+ + + +
+ +
+
+
± +
+
+ + + + + + + + -4-+
+
+ +
+
aAbout 5 x 103 active
AVh
particles were added toabout 2 x 108 bacteriasuspended in 1 ml of10-2 MMgSO,. The same amount ofphage was added to a control tube containing 1 ml of 10-2 M MgSO,but no bacteria.Aftera 15-minincubationat 37C,thesuspensionswerecentrifugedfor5min at 10,000 rpmand the
supernatantwasassayedfor activephageparticles.+,Indicatesthat the supernatantcontainedless than 10% phageasfoundin thecontroltube; -, indicates that thesupernatantcontainedmorethan 50%phageasfound inthecontroltube.
"Thebacteriawereeither devoidofadsorbedphageparticles (-)orcoveredwithatleast100particles (+). cThebacteriawereeitherclearlyfluorescent (+)oralmostinvisible (-) underthemicroscope.
683
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.507.59.453.193.592.2]resistant bacteria upon addition of suitably
diluted antiserum
can bedetected
under themicroscope,
but this reaction does not have asufficient degree of
reliability.
For this reasonimmunofluorescence
waspreferred,
because
itgave
unambiguous results.
Out of 14 strains ofbacteria which
did notadsorb the
phage
al-though they have lambda receptor, 12 cannotbe
labeled
with theantibodies,
a result favoringinaccessibility
of the receptor in such strains(Table 3). On the other
hand,
outof 15E.
coliand
Shigella strains which
doadsorb the
phage14 reactwith the
antibodies.
Plating
efficiency
ofphage lambda
onvari-ous E. coli and
Shigella strains.
All the strainslisted inTable
1had firstbeen tested for theirability
tosupportgrowth
ofphage
lambda,by cross-streaking
against XVandXVh.
Only
E.coli K-12 HfrG6 gave a
positive
result.Once
itbecame
apparent that several of the strainscould
adsorb
thephage
a furthercheck of theirability
to support its growth seemed desirable.No
plaques
of XVorXC
157 couldbe
obtained
onany ofthe sevenstrains ofE.
coli which
adsorblambda,
nor on four out of'the 10strains ofS.
sonnei
which also adsorb thephage.
Theeffi-ciency
ofplating
ofphage lambda
on thesestrains must
be
less than 10-8 or 10-9.On
thesix
remaining
S.sonnei
strains,
however,
plaques
of XV andXCI57
wereobtained,
atfrequencies ranging
between
10-5and 10-7.Experiments
are in progress todetermine
whether the rare plaques
obtained
in thiscaseare formed
by lambda
mutants present in thephage
stock, orby wild-type
particles whichescape a restriction system.
DISCUSSION
Active
lambda
receptorcanbe
extracted f'rom most Mal+ strain of E.coli
andShigella.
On the otherhand,
nolambda
receptor was found inextracts of several Mal- strains of'
Shigella,
a result suggesting that regulation of maltose metabolism and lambda receptor synthesis is thesame inShigella
as it is in E.coli
K-12 (7). Lambda receptor activity could not be found in extracts of severalstrains
of'Salmonella
andLevinea, although
all the strainstested
wereMal-'. No searchhasbeen done in such bacteria
f'or a protein cross-reacting
immunologically
with lambda receptor. Such a protein may be present, still endowed
with
a f'unction inmal-tose transport andchemotaxis towards maltose,
but
unable toinactivate phage lambda. Extracts of' several strains of' E.coli
andShigella inactivate wild-type lambda even
inabsence of chloroform or
ethanol,
contrary
towhat is found withextractsofE. coli K-12. This observation may be of
importance
for thestudy
of theinteraction between
phage
lambda and itsreceptor.
Although
all wild strains of' E. coli testedduring
the course of this worksynthesize
alambda receptor molecule active in
vitro,
they
do not support
growth
of thephage.
In manycasesthis is due to
inaccessibility
oftherecep-tor in
vivo,
since thebacteria donot adsorb thephage
and do not react withantireceptor
anti-bodies. In suchbacteria the receptor islikely
tobe buried under the
lipopolysaccharide
(5).
However, although
the receptor is accessible inatleast one
rough
strain, i.e.,
E. coliK-12(14),
it is also accessible inseveral smooth strains of
well-characterized
0-antigenic specificity.
Apossible interpretation
would be that thelipo-polysaccharide
of the different strains have0-specific
chains ofdifferent sizes which inturnhavedifferent effectsonthe
accessibility
ofthe receptor.In
sevoral
strains of E.coli
andShigella
onwhich
phage
lambda canadsorb,
it stillcannotgrow.
Why
this is sohas not beeninvestigated.
The
phage
may be unable toinject
its DNA(16),
its DNA may be restricted (2, 12),and/or
someofthe
phage
geneproducts
may be unabletointeract
correctly
withsomespecificproducts
present in these bacteria (1, 6).
Conceivably,
elucidation of' what blocks
phage
development
in such bacteria could
yield
interesting resultsconcerning
the interaction of thephage
with itshost.
Particularly
suitable for such studies would be the strains of'S. sonnei shown heretoplate
thephage
at a low but measurablefre-quency.
ACKNOWLEDGMENTS
We gratefully acknowledge the skillful technical help of Madeleine Jolit, Celestine Derval, and Christine Charle-Marsaines. We express our thanks to Claude Frehel for examiningourpreparationsunder theelectronmicroscope.
This work wassupported by grants from the Centre
Na-tionalde la RechercheScientifique, theDelegationGenerale
a la RechercheScientifique et Technique, and the National
Institutesof Health.
LITERATURE CITED
1. Baron, L. S., I. R. Ryman, E. M. Johnson, and P. Gemski,Jr.1972.Lytic replicationofcoliphage lambda in Salmonella typhosa hvbrids. J. Bacteriol. 110:1022-1031.
2. Bertani, G., and J. J. Weigle. 1953. Host controlled
variation inbacterial viruses. J.Bacteriol.65:113-121.
3. Dessein, A., and M.Schwartz. 1974. Is there a pullula-naseinEscherichiacoli?Eur. J.Biochem.45:363-366. 4. Edwards,P.R., and W. H.Ewing. 1972.Identificationof
enterobacteriaceae, 3rd ed. Burgess Publishing Co., J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
BACTERIOPHAGE LAMBDA RECEPTOR
Minneapolis.
5. Gemski, P., Jr., J. A. Alexeichik, and L. S. Baron. 1972. Behavior of coliphage lambdainShigella flexneri2a.J. Virol. 10:668-674.
6. Georgopoulos, C.P., R. W. Hendrix, S.R.Casjens, and A. D.Kaiser. 1973. Host participationinbacteriophage lambda head assembly.J.Mol. Biol.76:45-60. 7. Hofnung, M. 1974. Divergent operons and the genetic
structureofthe maltose B region in Escherichia coli K12. Genetics76:169-184.
8. Lederberg, E. M. 1951. Lysogenicity in E. coli K12. Genetics 36:560.
9. Lederberg, J. 1947. Gene recombination and linked segregation inEscherichia coli. Genetics 32:505-525. 10. Lowry, 0. H., N. J. Rosebrough, A. J. Farr, and R.J.
Randall. 1951. Protein measurement with the Folin phenolreagent.J. Biol.Chem. 193:265-275.
11. Luria, S. E., and J. W. Burrous. 1957. Hybridization between Escherichia coli and Shigella. J. Bacteriol. 74:461-476.
12. Meselson, M., R. Yuan, and J. Heywood. 1972. Restric-tion and modificationofDNA. Annu. Rev. Biochem. 41:447-466.
13. Nairn, R. C. 1969. Fluorescent protein tracing, 3rd ed. Livingstone, Edinburgh.
14. 0rskov, I., and F. Orskov. 1960. The H. antigenof the "K12" strain. A new E. coli H antigen: H48. Acta Pathol. Microbiol.Scand. 48:47.
15. Randall-Hazelbauer, L., and M. Schwartz. 1973. Isola-tionofthe bacteriophage lambdareceptorfrom Esche-richia coli. J.Bacteriol. 116:1436-1446.
16. Scandella, D., and W. Arber. 1974.AnEscherichia coli mutant which inhibits the injectionofphageADNA. Virology58:514-525.
VOL. 15, 1975