Copyright 0 1975 AmericanSocietyforMicrobiology Printed in U.S.A.
Termination
Sites for Adenovirus
Type
2
DNA
Replication
ASLIHAN TOLUN AND ULF PETTERSSON*
Department ofMicrobiology, The Wallenberg Laboratory, Uppsala University, Uppsala, Sweden
Received for publication 6 May 1975
Termination sites for replication of adenovirus type 2 DNA have been
demonstrated at both ends of theviral chromosome by the procedure of Danna
and Nathans (1972). Single-stranded DNA from replicating intermediates was
also characterized by hybridization with separated strands of viral DNA. The
results indicate that both strands areexposed during replication.
Adenovirus type 2 (Ad2) DNA is a
nonper-muted linear duplex molecule with no
single-strand discontinuities (8, 25). The molecular
weight isabout 23 x 106 (4, 8, 25) and there are
inverted terminal repetitionsatthe ends(7,26).
Ad2 DNA labeled for short time periods was
observedto exhibit some propertiesnottypical
for mature molecules: it sediments with afast
rate in sucrose gradients and bands at a high
density in CsCl equilibrium gradients (12, 21,
24). Pulse chase experiments have shown that
these properties are due to the presence of
single-stranded branches on replicating
mole-cules (10, 12, 24). Replicating DNA was also
studiedbyelectronmicroscopy:Y-shaped
mole-cules with a single-stranded arm and linear
molecules of unit length with single-stranded
gaps ofvarying length were observed (19, 21,
24), suggestinga"stranddisplacement model"
for replication of adenovirus DNA (19, 21).
Replication starts at one end and the progeny
strand, which issynthesized first, displacesone
of the parental strands, thus leading to a
Y-forked intermediate. Afterreplicationofone
strand is completed, thereplicating
intermedi-ate separates to give a linear duplex DNA
molecule and a free single strand which is
replicated separately. Sometimes replication
initiates on the displaced strand before
dis-placement synthesis iscompleted (5, 19).
The strand displacement model raises the
question whether replication starts from one
specific end of the molecule always displacing
the same strand.Sussenbach et al. (19) studied
replication of Ad5 DNA in vitro in isolated
nuclei. Hybridization experiments and analysis
ofpartially denatured replicating molecules by
electron microscopy suggested that onespecific
strand always is displaced and thatreplication
starts from the AT-rich end of Ad5 DNA.
Experiments reported by Lavelle et al. (10)
in contrast, showed thatsingle-stranded DNA,
which was isolated from cells infected with
759
Ad2, hybridizes equally well with both strands
oftheviral DNA.
Inthe present study we have investigated the
termination points for Ad2 DNAreplication by
the method of Danna and Nathans (3).
Termi-nation sites at both ends have been detected.
We have also characterized the single-stranded branches that are exposed during replication.
MATERIALS AND METHODS
Cell cultures and virus. HeLa or KB cells were concentrated to 107cells/mlinEagle spinnermedium with 2% calf serum and infected with Ad2 at a
multiplicityof 104particles/cell.After 30 minthe cells were diluted to 2 x 10' cells/ml with warm spinner medium with 7% calf serum. At 13 to 14h the cells wereconcentrated to 1 to 2 x 106cells/ml andpulse
labeled for 5, 10, 45, and 60 min. To label thenewly synthesized DNA,50 to150ACiofmethyl-['H
Hthymi-dine(New EnglandNuclear; 40 to 60mCi/mM)per ml wasused. At the end of eachpulsethecellswere mixed with crushed frozenphosphate-bufferedsaline and the DNAwasextracted.
32P-labeled virus and DNA was madeaccordingto Pettersson andSambrook(15).
DNA extraction. Cells were pelleted and
resus-pended in 0.65%Nonidet P-40 inphosphate-buffered
saline at a concentration of 5 x 10' cells/ml or less. After incubation at 0C for 10 min the nuclei were spun down, washed with phosphate-buffered saline, lysed with 0.5% sodium dodecyl sulfate, and digested with 1 mg of Pronase per ml in a buffer which contained 0.1 M EDTA, 0.1 M NaCl, and 0.01 M
Tris-hydrochloride (pH 7.9) for 2 h at 37 C. The sample was then extracted twice with phenol (satu-rated with 0.5 MTris-hydrochloride, 10 mM EDTA, pH 7.9) and once with chloroform:isoamylalcohol
(24:1). All steps except the Pronase digestion were
carried outat 4C.
Restriction enzymes. EndoR-EcoRI waspurified from Escherichia coli strain RY13 as described by Pettersson andPhilipson (14). Digestions were made in0.1 MNaCl,0.01MMgCl2,and 0.01 M Tris-hydro-chloride (pH 7.9).
EndoR-HpaIwaspurified by the method of Sharp et al. (18) and was a kind gift from R. Kamen,
on November 10, 2019 by guest
http://jvi.asm.org/
760 TOLUN AND PETTERSSON
ImperialCancer Research Fund, London. Incubation mixturescontained0.1M KCl, 0.006 MMgCl2, 0.006 M ,B-mercaptoethanol, and 0.006 M Tris-hydrochlo-ride (pH 7.5).
EndoR HindIIIwaskindly supplied by Ernst Win-nacker from the University of Cologne, West Ger-many. Digestions were carried out in 0.06 M NaCl, 0.007 M MgCl2, 0.001 M 3-mercaptoethanol, and 0.Q06 M Tris-hydrochloride (pH 7.9).
Polyacrylamidegelelectrophoresis. After diges-tionwas completed samplesfor electrophoresis were extracted withphenol,precipitated with ethanol, and dissolvedin0.1 ml of0.01 M Tris-hydrochloride (pH 7.9) with 10- M EDTA (TEbuffer).
Composite gels which contained 0.7% agarose (Sigma) and2.2or1.1%acrylamide(Serva,Germany) were prepared asdescribed by Pettersson et al. (13). Electrophoresis was carried out in cylindrical gels at 4.5 V/cm in a buffer which contained 5 mM sodium acetate, 1mM EDTA, and 0.04 M Tris-hydrochloride (pH 7.9). Fragments obtained after cleavage with EndoR.HpaI and EndoR.EcoRI were separated on 13-cmgels and fragments after cleavage with EndoR-HindJII were separated on 18-cm gels. Bands were visualized by staining with ethidium bromide (18) and DNA was eluted electrophoretically from gel slices as described by Pettersson et al. (13). To achieve separationbetween fragments HindIII-D and -E and between fragments HindIII-F and -G, the combined fragments were eluted from gel slices and subjectedto asecondcycle of electrophoresisfor36 to 48h.
Hybridizations. Hybridizations of replicating DNA with isolated single strands of Ad2 DNA were carried out in 0.14 M phosphate buffer (equimolar amounts of Na2HPO4 and NaH2PO4) with 0.4% so-diumdodecylsulfate and 1MNaClat 65C for4to 5 days.Sampleswerethen assayedby chromatography on hydroxylapatite columns (Bio Gel HTP) as de-scribedbySambrooketal.(16).Single-stranded DNA was eluted with 0.14 M phosphate buffer with0.4% sodium dodecyl sulfate and double-stranded DNA with 0.4 M phosphate buffer with 0.4% sodium dodecyl sulfate.
S3P-labeled DNA was determined as Cerenkov radiation. Fractions with 3H-labeled DNAwere
pre-cipitated on membrane filter disks (Millipore Corp.)
with10%trichloroacetic acid before counting. Separation of the complementary strands from Ad2 DNA. Unlabeled strands were prepared by equilibrium centrifugation inCsClofdenatured viral DNA which had been complexed withribopoly(U,G) as described in detail by Tibbetts et al. (23). 2P-labeled strandswerepreparedby incubationof dena-tured labeled DNA with an excess of one unlabeled DNA strand under annealing conditions. Labeled complement-specific DNAwasisolatedby fractiona-tion of the incubation mixture on hydroxylapatite. Themethodforstrandseparationof32P-labeledDNA has beendescribedindetailby Tibbetts and Petters-son(22).
Thestrand whichhas a higherbuoyant densityin CsCl whencomplexedwithpoly(U,G)is referredto as
the H-strand. The strand with a lighter buoyant
density when complexed with poly(U, G) is accord-ingly designated the L-strand.
Equilibrium centrifugation in CsCl. 3H-labeled DNA and 32P-labeled marker Ad2 DNA was mixed with TE buffer and thedensity was adjusted to 1.705 g/ml with solid CsCl. Centrifugation was performed in aSpinco Ti5OorTy5O fixed angle rotorforat least 42 hat 40,000 rpm at 20C. Gradients were fractionated from thebottom and aliquots were assayed for trichlo-roacetic acid-insoluble radioactivity.
In some experiments the DNA was sheared by passage twicethrough a 25-gauge syringe needle prior toequilibrium centrifugation.
Velocity sedimentation in sucrose. Samples (0.1 to0.2ml)werelayeredonpreformed 5 to 25% sucrose gradients which contained 1 M NaCl, 1 mM EDTA, and 10 mM Tris-hydrochloride, pH 7.9. Separations werecarried out in the SpincoSW56 rotor for 90 min at 50,000 rpm and 20 C. Gradients were fractionated fromthe bottom and aliquots were assayed for trichlo-roacetic acid-insoluble radioactivity.
BND-cellulose chromatography. Benzoyl-naphthoyl-DEAE-cellulose (BND-cellulose) which had been purchased from Serva, Germany, was washed with ether as described by Sedat et al. (17). DNA in 0.3 M NaCl, 1 mM EDTA, and 0.01 M Tris-hydrochloride (pH 7.9) was loaded on columns (2
by 1.5cm). Double-stranded DNA was eluted with a buffer which contained 1MNaCl, 1 mM EDTA, and 10 mM Tris-hydrochloride (pH 7.9), and DNA con-taining singlestrandswaseluted with 2% caffeine,1M NaCl, 1 mM EDTA, and 10 mM Tris-hydrochloride (pH 7.9). The column was finally washed withabuffer containing 0.1 M NaOH, 0.9 M NaCl, and 1 mM EDTAtoeluate any remaining DNA. Fractions with double-stranded DNA were pooled and the density wasadjustedby addition of solid CsCl before equilib-riumgradient centrifugation.
RESULTS
Location of termination sites for
replica-tion ofAd2 DNA. Completed DNAmolecules,
which have been pulse labeled during a time
period shorter than the time needed for one
complete round ofreplication, willbe
preferen-tially labeled at the terminationsites for
repli-cation. With the use of restriction endonu-cleases it is possible to isolate specific DNA fragments and thereby determine the specific
radioactivity in different segments of a given
DNA molecule. This experimental approach has been utilized to map the origin and termi-nation pointfor replication on simian virus 40
andpolyoma DNA (2, 3).
We have pulse labeled cells 14 h after infec-tion for5,10, and60min. Thetotalintracellular
DNA was extracted as described above and
the DNA was fractionated
by chromatography
on BND-cellulose. Since intermediates in
adenovirus replication contain
single-stranded
branches, this procedureshouldefficiently
sep-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
ADENOVIRUS DNA REPLICATION
arate replicating viral DNA molecules from
mature DNA molecules. Fractions which were
eluted with 2% caffeine from BND-cellulose
columnscontained, asexpected,predominantly
DNA with a higher buoyant density than
ma-ture viral DNA (not shown). About 20% of the
labeled DNA after a 5-min pulse was eluted
with 1 M NaCl and these fractions were
col-lected and purified by equilibrium
centrifuga-tion to remove contaminating cellular DNA
(Fig. 1). In some experiments the DNA was
further purified by velocity sedimentationon a
sucrose gradienttoeliminate traces of
replicat-ing or degraded molecules. Purified mature
DNA was precipitated with ethanol and
di-gested with EndoR-EcoRI after addition of 32P-labeled DNA which had been extracted
from purified virus. Fragments were separated
on composite 1.1% polyacrylamide, 0.7%
aga-rose gels which were stained with ethidium
bromide. Bands wereeluted electrophoretically
and counted after precipitation with
trichloro-acetic acid. The ratio between 8H- and
1P-labeled DNA was determined for all six
frag-ments and the results are illustrated inFig. 2.
Corrections were made for differences in base
composition between individual fragments by
comparing the
'H/32P
ratios for mixtures ofDNA uniformly labeled with ['HIthymidine and [3"P ]phosphate.Aclear increase in the ratio wasobserved towards theright handendofthe
genome when DNA was analyzed after pulse
labelingfor 5and 10 min. This suggests thatone
terminus for replication is located at the right
hand end of Ad2 DNA. Possible termination
points in the left hand half of the Ad2 DNA
could not be resolved after cleavage with
En-doR.EcoRI since the large EcoRI-A fragment
which corresponds to 59% of Ad2 DNA is
located inthispart of the molecule(11).
There-fore, pulse-labeled, mature moleculeswerealso
cleaved with EndoR.HpaI after addition of
32P-labeled DNA. This enzyme cuts Ad2 DNA intosevenfragments, four of which arelocated
within fragmentEcoRI-A(6, 11).The fragments
wereseparatedoncomposite 1.1%
polyacrylam-ide, 0.7% agarose gels. After staining with
ethidium bromide, the fragments were eluted
andanalyzedasdescribed aboveand the results
aresummarized inFig.3. An increasing
3H/3"P
rato was observed also towards the left hand
endoftheAd2 DNAwhencompleted molecules
were analyzed which had beenpulse labeled for
' a.
(M CN
100 0
800
r-.E
600
-oo
3 0
0
X0 200
1 5 10 15 20 25 30
Fraction number
400-c
E
In
-W
0
0 0
0
0
0.
(N0
300-31
200±2
100+
10
0
O
f|_t~~~~~~~~~~~
1k
B -n
-10 15 20 25 30
Fraction number
35 40
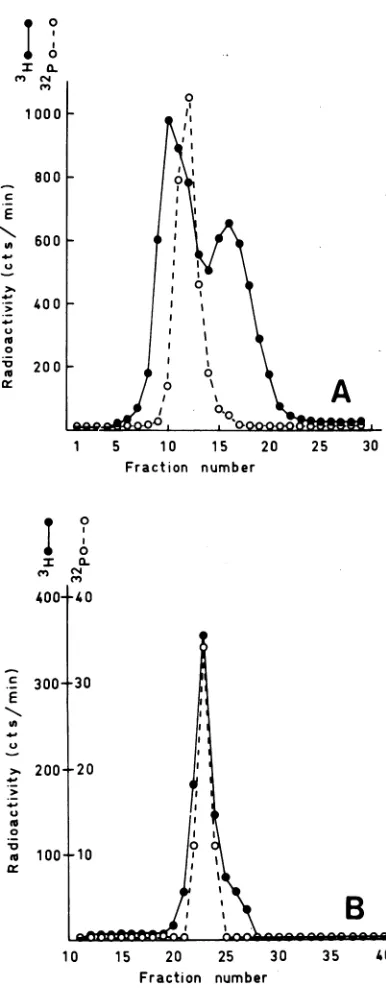
14hafterinfection. Thetotal intracellularDNA was extracted asdescribedandfractionated by BND-cel-lulosechromatography.Centrifugationwasperformed
at 20C and 40,000 rpmfor 42h in theSpinco Ti50 rotor. The density increases towards the left. (A)
Intracellular DNA before fractionation by BND-cel-lulose chromatography. (B) Double-stranded DNA elutedfrom BND-cellulosewithabuffercontaining1 M NaCl, 1 mM EDTA, 0.01 M Tris-hydrochloride (pH 7.9).Symbols:0,intracellularDNA;0, "2Pviral DNA marker.
FIG. 1. CsClequilibriumgradient centrifugation of intracellular DNA before and after fractionation by
BND-cellulose chromatography. Cells were pulse la-beledfor 10minwith [3H]thymidine(120MCi/mI)at
761
VOL16, 1975
B
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.493.247.440.72.570.2]762 TOLUNAND PETTERSSON
A
a.
CM4
I
CY)
0
2.5
F
2.0 H
1.5
H
1.0
0.2
0.4
[image:4.493.55.451.66.321.2]FRACTION
OF
FIG. 2. Relativeamountsof['H]i
different fragments obtained by c
completed molecules with EndoR- I had been pulse labeled for5(0), 1
(x) was analyzed. "2P-labeled DN
n1/P;{i~,,v&vien"IZQ S N roAcroa
B F
D
E
C
incells
infected with
Ad2. Results from
severalI I I I I laboratories indicate thatreplicatingAd2 DNA
moleculescontainanunusualamountof
single-stranded branches (12, 21, 24). Itwastherefore proposed that adenovirus DNAreplicates by a
strand displacement mechanism. In this study we have designed experiments to determine
whether one specific or both strands are
dis-placed during replication. Since replicating
molecules contain exposed single strands it
should be
possible
to directly performhybridi-zation studies between replicating molecules
and separated strandsofAd2 DNA. Two kinds
/ ofexperiments have been performed.
10
Infected cells were pulse labeled 14 h after0 infection with
[3H
Ithymidine.
The totalintra-0 cellular DNAwasextractedandfractionated
by
o equilibrium centrifugation inCsCl. Most of the
'rx.**X
label banded attheposition of the viral marker____________sDNA and a small peak was observed at the
0.6
0.8
density
ofcellular DNA(Fig.
5). DNA with agreaterbuoyantdensitythan the marker(heavy
Ad
2 DNA
DNA) was also
observed.
Ithymidine label in This DNA has
previously
been found tocleavage of newly contain replicating molecules and its increased
;coRI DNAwhich buoyant density is due to the presence of single
0
(0),
and 60min strands (12). Fractions corresponding to therA extracted from heavy DNA were pooled as indicated in
Fig.
5'"nr 'o fWVDfWmO"#
purw&iea vurwons was usea asareference. inefragmenc
order (11) is shown on top of the figure. Corrections have been made for differences in thymidine content inindividual fragments by determination of the ratio between3H/32Pfor fragments which had been derived
fromDNA uniformly labeled with["2P]phosphateand
[3H]thymidine. The ratios in the figure are normal-ized to fragment RI-A.
5 and 10 min. Thus there seems to be an
additional termination site at theleft hand end
ofthe Ad2 genome.
To confirm these results and to improve the
resolution ofpossible internal termination sites,
mature DNA that had been pulse labeled for
10 min was also cleaved with EndoR.HindIH.
Thisenzyme cuts Ad2DNA in12fragments (R.
Roberts, personal communication), the order of
which is shown in Fig. 4. All fragments except
the smallest(HindIII-L)wereisolated from gels
and the ratio between 'H
pulse-labeled
and32P-labeled reference DNA was determined.
Thetermination sitesattheright and left hand
ends were confirmed but no clear internal
termination sites were observed. A minor in-crease in ratio near the middle of the Ad2
genome was observed but this increase was
always much lessprominant than the maxima
atboth ends. (Fig. 4)
Characterization of single-stranded DNA
E
C
F
A .,
2.5
0~
c,4
\
2.0
I
p_1.5
1.01
0.2
0.4 0.6 0.8FRACTION OF Ad 2 DNA
FIG. 3. Relativeamountsof ('H]thymidinelabel in different fragments obtained by cleavage of newly completed molecules with EndoR -HpaI. The
sym-bols are explained in the legend to Fig. 2. The fragment order (11) is shown on top of the figure. Fragments HpaI-A and HpaI-B were not separated. All values are adjusted for variations in thymidine
content and normalized to a mixture of fragments HpaI-AandHpaI-B.
I I
A
B
D G
I I I
0
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.493.258.447.352.591.2]ADENOVIRUS
G
C
B
I J D
I I I I I I
a.
CN
I
(V)
0
I-A
H
2.5
_
2.0
1.5
1.0
00
0.2
0.4
0.6
0
FRACTION
OF
Ad 2
FIG. 4. Relativeamountsof[3HJthymidi different fragments obtained by cleavage completed molecules with EndoR-Hinc
DNA pulse labeled for 10 min was anal
fragmentorder which is shown on the topo,
was kindly provided by R. Roberts, Cc
HarborLaboratory. Thefragmentsize was
by determination ofthedistributionof32p eachfragmentafter cleavage andseparatic
which wasuniformly labeled with [32P]pho Pettersson unpublished data). Fragment ignated according to their mobility in ag It has been observed that fragments Him -E and fragments HindHI-F and -G r the reverse order in composite 0.7% ag acrylamide gels (R. Roberts, personal cc tion). The ratios in the figure are norn fragmentHindfIU-A.
and dialyzed against TE buffer. He
withoutfurther treatment wasthen i
underannealing conditionswith 31P-l
and L-strand probe DNA as describ
The fraction of the
"!P-labeled
pr(whichbecamedoublestranded after ii
wasdeterminedbychromatography o
ylapatite. Both labeled probes hybric
nearly equal extent in repeated
exj
andabout50% ofthe
8'P-labeled
DNArecovered in the double-stranded fr
gardless whether H- or L-strand DNA for hybridization (Table 1). Saturati
strands was presumably not achieve(
someregions of each strand are expos
lowfrequency and because of self-hyb
between replicating molecules. The re
gest thatboth strands are exposed dui
cationofAd2 DNA.
LE F
K
Anothertype
ofexperimentwasperformedtoconfirm that both the complementary strands
are exposed in adenovirus-infected cells: cells
were pulse labeled for 45 min, 14 h after
infection asdescribed above. The total
intracel-lular DNA was extracted and fractionated by
equilibrium centrifugation in CsCl. The
frac-tions which corresponded to heavy DNA were
pooled as indicated inFig. 5. After removal of
CsCl by chromatographyon Sephadex G50the
DNA wasdegraded bysonication for 30 s inTE
buffer with 0.01 M NaCl. The sonicated DNA
was chromatographed on hydroxylapatite and
fractions which eluted with 0.14 M phosphate
buffer were collected. About 20% ofthe pulse-labeledheavy DNA wasrecovered inthe single-stranded DNA fraction (Table2) whereas
con-trol experiments showed that less than 3% of
duplex adenovirus DNA elutes with 0.14 M
1.8
phosphate buffer after sonication.Single-stranded DNA, labeledwith [H]thymidine and
DNA
isolated by hydroxylapatite chromatographyine labelin fromsonicated heavy DNA, was then incubated
>
of
newly under annealing conditions with an excess oflIII.
nnly unlabeledH- andL-strandDNApreparedfromlyzed.
The virions asdescribed byTibbettsetal. (23)Thefld
Spring
fraction of 3H-labeled DNA which becamedou-estimated
ble stranded after incubation was determinedlabeled in by chromatography on hydroxylapatite. The
onofDNA
)sphate(U. o
s are des-
I
rarose gels. 0.
dIIU-D and
rnigrate in 30.000 {arose-2.2% -700
Dmmunica-nalized to
'n 20.000--500
eavy DNA
-1
incubated - o
abeled H- 300
led above. 10.0
0.0o
obe DNA
'
>ncubation in hydrox-dized to a periments
kcould be
action
re-Lwasused ion of the d because
;ed witha
iridization ,sults
sug-ring
repli-1 5 10 15 20 25 30
Fraction number
FIG. 5. Equilibrium centrifugation in CsCI of
in-tracellular DNA, extracted from cells which were
pulse labeled with [3H]thymidine(50 uCi/ml) for 45 min at 13 h after infection. The initial density was 1.705g/mlandcentrifugation was at 40,000 rpm and 20C for 43 h in a Spinco Ti5O rotor. Fractions containing heavy DNA were pooled as indicated by the bar. The density increases towards the left.
Symbols:
@,
intracellularDNA; 0,"2P-labeled
viral DNA marker.VoL16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.493.49.240.81.308.2] [image:5.493.256.446.391.585.2]764 TOLUN ANDPETTERSSON
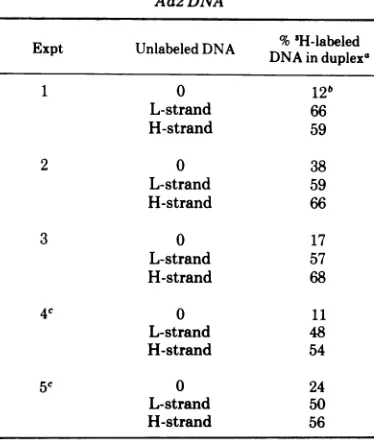
TABLE 1. Hybridization between 'H-labeledheavy
DNA and"2P-labeledcomplement-specific Ad2 DNA
Amount of %
3Piea
%"2P-la-Amountav ofA
leledL- beled H-Expt heavYDNA; strand DNA' strandDNAbin duplex in duplex
1 350 14c 13
1,050 26 24
2 2,500 15 23
6,250 27 34
3d 28,000 41 51
94,500 45 58
aThe amount of 3H label was used toquantitate heavy DNA from differentexperiments. The DNA in all experiments was labeled under identical condi-tionsandshould thus have thesamespecificactivity. Because of different amounts of contaminating com-pleted molecules in preparations ofheavy DNA from different experiments there may, however, be some variations in the ratio between concentration of sin-gle-stranded DNA and radioactivity.
'The probe DNAwasdegraded to anapproximate size of 350nucleotidesby boilingfor20 min in 0.3M NaOHbeforehybridization.
cLess than 4% of theprobeDNA was recovered in
the duplex fraction after incubation without heavy DNA.
dIn this experiment the extracted DNA was sheared bypassage twice througha 25-gaugesyringe needle prior to equilibrium gradient centrifugation to increase the capacity of the gradient.
TABLE 2. Percentage of 'H-labeled sonicated heavy DNA which is eluted with 0.14 Mphosphate buffer
afterchromatographyonhydroxylapatitea
%HeavyDNAeluted
Expt with 0.14 Mphosphate
buffer
1 24
2 20
3 20
4 18
aInfected cellswhich had been pulse labeledfor 45 min wereextracted and the total intracellularDNA
wasfractionatedbyequilibrium gradient
centrifuga-tion. Fractions corresponding to heavy DNA were
pooled and sonicated before passage over
hydroxyl-apatite columns.
results showed that 50to 65% of the 'H-labeled
DNA became double stranded after incubation
with both L- and H-strand DNA (Table 3). A
significant fractionof thesingle-strandedDNA
also self-hybridized when incubated under
an-nealing conditions without addition of
unla-beled single strands of Ad2 DNA (Table 3). This
explainswhymorethan50%ofthelabeled DNA
in most experiments was recovered as duplex
DNA when hybridized with either one ofthe
complementary strands.
Sussenbach and van der Vliet (20) have
shown that single-stranded viral DNA
accumu-latesininfected cells which havebeengrown in
the presence ofhydroxyurea. We have isolated
DNA from infected cells which were treated
with 10' M hydroxyurea at the time of
infec-tion.The cellswerewashed withwarmmedium
18 h after infection and then incubated inthe presenceof4
,Ci
of['H]thymidineperml for 30min.The total intracellular DNAwasextracted
and fractionatedonCsClgradients. The labeled
DNA from hydroxyurea-treated cultures
con-tainedconsiderably more single-stranded DNA
ascomparedwith cultures whichweregrown in absence of the drug (not shown). Heavy DNA
was isolated, asdescribedabove, and degraded
by sonication. Single-stranded DNA, labeled with ['H
]thymidine
wascollected afterchroma-tography on hydroxylapatite and used in
hy-bridizations with unlabeled L- and H-strand
TABLE 3. Hybridization of'H-labeledsinglestrands from heavy DNA with unlabeled L- and H-strand
Ad2 DNA
Expt Unlabeled DNA DNA in
duplexd
1 0 12b
L-strand 66
H-strand 59
2 0 38
L-strand 59
H-strand 66
3 0 17
L-strand 57
H-strand 68
4c 0 11
L-strand 48
H-strand 54
5c 0 24
L-strand 50
H-strand 56
aInexperiments 2 and 3 twodifferent concentra-tions ofunlabeled L- and H-strandswereused which gave identical results.
"Sampleswhichwerenotincubatedunder
anneal-ing conditions contained less than 4% of DNA in duplex.
cDNA was isolated from cells which had been treated withhydroxyurea.
J. VIEROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.493.265.452.376.596.2] [image:6.493.55.249.463.542.2]ADENOVIRUS DNA REPLICATION 765
DNA. The results showed that single strands
whichweregeneratedin the presence of
hydrox-yurea also hybridizedwith both strands of the
viral DNA (Table 3).
DISCUSSION
Experimentsperformed according to the
pro-cedure of Dannaand Nathans (3) reveal
termi-nation sites at both ends of the Ad2 genome.
This finding is inagreement with the results of
Horwitz (9) which showed thatthe two
molecu-lar halves ofAd2 DNA have the samespecific
activity whennewlycompletedDNAmolecules
were analyzed after pulse labeling for short
periods of time. For DNA molecules which are replicatedasproposedbyCairns (1)thiswould
suggest an internal origin ofreplication.
How-ever, available experimental data convincingly
show that the twostrands ofadenovirus DNA
replicate asynchronously (5, 12, 19, 21, 24)
which makes the interpretation of the results
more difficult. Sussenbach and his colleagues
(5, 19, 21) haveproposed that replication of Ad5
DNAalways starts at the AT-rich end(the right
handend). Assuming that Ad2 DNA replicates in an identical way one wouldexpect to find a
terminationsite at the left hand end as we have
demonstrated after cleavage of pulse-labeled
DNA with
EndoR.HpaI
andEndoR.HindM.
From the Sussenbach model several additional
termination sites would be expected from the
so-called complementary strand synthesis. If
displacement synthesis predominantly leads to
duplex daughter molecules and free single
strands, a strong termination site could be expected also at the right hand end if one
assumes that replication only rarely initiates
internally on the displaced single strand.
However, we have found by hybridization
ex-periments that both complementarystrandsof
Ad2 DNA are exposed in cells which contain
replicating molecules; a finding which is in
agreement with the resultsreportedby Lavelle
etal.(10).The resultsare inconflict with those ofSussenbachet al.(19) and would be
compati-ble with a mechanism of replication which
allows initiation to take place at either end of
the adenovirusDNA. It is noteworthy that the
two ends of adenovirus DNA are identical
because of thepresence of aninverted terminal
repetition (7, 26) and the ends may thus serve
as recognition sites for the DNA polymerase.
The discrepancy between our results and those
of Sussenbach et al. (19) are puzzling. One
possibility is that replication during the first
round(s) initiates at one specific end, but later
proceeds according to another scheme which
leads to displacement of both strands. When comparing different studiesonadenovirusDNA
replication it is important toremember thatall
investigations so far, including our own, have
been made under extraordinary conditions,
such as growth in the presence of
bromodeoxy-uridine, infection athigh multiplicities, or
syn-thesis in isolatednuclei. Also, techniquesused
forselective extraction and purification of
repli-cating DNA may preferentially lose molecules
which are ofparticular importance for
replica-tion.Therefore theresults shouldbeinterpreted
with caution until experimental conditions are available which allow studies of replicating molecules under more natural conditions.
We have recently learnedthat E. Winnacker
and co-workers (personal communication) have independently demonstrated termination sites
for Ad 2 DNAreplication near both ends of the
viral DNA.
ACKNOWLEDGMENTS
We wish to expressour gratitude to Lennart Philipson for many helpful discussions and forproviding laboratory facili-ties to ourdisposal. Wearealso indebtedtoR. Roberts at the Cold Spring Harbor Laboratory for making the HindI cleavagemap availabletous prior topublication and to R. Kamen,ICRF laboratory, for a gift of HpaIendonuclease. We thankJ.Rosefor communicatingresultsbefore publication. We areparticularly grateful to E. Winnacker for a generous gift of endonuclease HinduI and for open discussions of unpublishedresults.
Grantsfrom the Swedish MedicalResearch Council and the Swedish Cancer Society supported this study. Aslihan Tolun is a recipient of a guest scholarship from the Swedish Institute.
LITERATURE CITED
1. Cairns, J. 1963. The chromosome of Escherichia coli. ColdSpring HarborSymp.Quant.Biol.28:43-46. 2. Crawford,L.V.,A. K.Robbins,and P.M.Nicklin. 1973.
Location of theorigin and terminus ofreplication in polyomavirusDNA. J.Gen.Virol. 25:133-142. 3. Danna,K.,and D. Nathans. 1972.Bidirectional
replica-tion ofsimian virus 40 DNA. Proc.Natl. Acad. Sci. U.S.A.69:3097-3100.
4. Doerfler, W., and A. K.Kleinschmidt. 1970. Denatura-tion pattern of the DNA of adenovirus type 2 as
determined by electron microscopy. J. Mol. Biol. 50:579-593.
5. Ellens, D.J.,J. S.Sussenbach, and H. S. Jansz. 1974. Studiesonthe mechanism ofreplicationofadenovirus DNA. III. Electron microscopy of replicating DNA. Virology 61:427-442.
6. Gallimore, P. H., P. A. Sharp, andJ.Sambrook. 1974. Viral DNA in transformed cells. II. A study of se-quences of adenovirus DNA in nine lines of trans-formed ratcells usingspecific fragments ofthe viral genome. J. Mol. Biol. 89:49-72.
7. Garon,C. F.,K.W.Barry, and J. Rose. 1972.Aunique formofterminalredundancyinadenovirus DNA mole-cules. Proc. Natl. Acad. Sci. U.S.A. 69:2391-2395. 8. Green,M.,M.Pinia,M R.Kimes, P. C.Wensink,L. A.
MacHattie, and C. A. Thomas, Jr. 1967. Adenovirus DNA. I. Molecular weight and conformation. Proc. Natl.Acad. Sci. U.S.A.65:226-233.
VOL16,1975
on November 10, 2019 by guest
http://jvi.asm.org/
766 TOLUN AND PETTERSSON
9. Horwitz, M. S. 1974. Location of the origin of DNA replicationinadenovirustype2.J. Virol. 13:1046-1054. 10. Lavelle, G., C. Patch, G., Khoury, and J. Rose. 1975. Isolation and partial characterization of single-strandedadenoviral DNA produced duringsynthesis of adenovirustype2DNA. J. Virol.16:775-782.
11. Mulder, C., J. R. Arrand, H. Delius, W. Keller, U. Pettersson, R. J. Roberts, and P. A. Sharp. 1974. Cleavagemapsof DNA fromadenovirustypes2and 5 by restriction endonucleases EcoRI and HpaI. Cold Spring HarborSymp. Quant. Biol. 39:397-400. 12. Pettersson, U. 1973.Some unusual propertiesof
replicat-ing adenovirustype2DNA. J. Mol. Biol. 81:521-527. 13. Pettersson, U., C.Mulder, H. Delius, andA.Sharp.1973.
Cleavage ofadenovirus type 2 DNA into six unique fragments by endonuclease R.RI. Proc. Natl. Acad. Sci. U.S.A. 70:200-204.
14. Pettersson, U., and L. Philipson. 1974. Synthesis of complementary RNAsequencesduringproductive
ade-novirus infection. Proc. Natl. Acad. Sci. U.S.A. 71:4887-4891.
15. Pettersson, U., and J. Sambrook. 1973.Amount of viral DNAinthegenomeof cellstransformed by adenovirus
type2.J. Mol. Biol.73:125-130.
16. Sambrook, J., P. A. Sharp, and W. Keller. 1972. Tran-scription ofsimian virus 40.I.Separationofthe strands
of SV40 DNA and hybridization of the separated
strandstoRNAextracted fromlytically infected and
transformed cells. J.Mol. Biol. 70:57-71.
17. Sedat, J., A. Lyon, and R. L. Sinsheimer.1969. Purifica-tion ofEscherichia colipulse-labeled RNAby
benzoyl-ated DEAE-cellulose chromatography. J. Mol. Biol.
44:415-434.
18. Sharp, P. A., W. Sugden, and J. Sambrook. 1973. Detectionoftworestrictionendonucleaseactivitiesin Haemophilus parainfluenzaeusing analytical
agarose-ethidium bromide electrophoresis. Biochemistry 12:3055-3063.
19. Sussenbach, J.S., D. J.Ellens, and H.S. Jansz. 1973. Studiesonthe mechanismofreplicationofadenovirus DNA. II. Thenatureofsingle-stranded DNAin replica-tive intermediates. J. Virol. 12:1131-1138.
20. Sussenbach,J.S.,and P. C.vanderVliet.1973.Studies onthe mechanism ofreplicationof adenovirus DNA.I.
The effectofhydroxyurea. Virology 54:299-303. 21. Sussenbach,J.S.,P.C.vanderVliet, D. J. Ellens,and
H. S. Jansz. 1972.Linearintermediates in the replica-tion of adenovirus DNA.Nature (London) New Biol. 239:47-49.
22. Tibbetts, C., and U. Pettersson. 1974. Complementary strand-specific sequences from unique fragments of adenovirus type 2 DNA for hybridization-mapping experiments. J. Mol. Biol.88:767-784.
23. Tibbetts, C.,U.Pettersson,K.Johansson,and L.
Philip-son. 1974. Relationship of mRNA from productively
infected cellstothecomplementary strandsof
adenovi-rustype2DNA. J. Virol. 13:370-377.
24. Van der Eb,A. J. 1973.Intermediatesin type 5
adenovi-rusDNAreplication.Virology51:11-23.
25. Van derEb,A.J.,L. W. VanKesteren,and E. F. J. Van
Bruggen. 1969. Structural properties of adenovirus DNA's. Biochim. Biophys. Acta182:530.
26. Wolfson, J., and D. Dressler. 1972. Adenovirus 2 DNA
contains an inverted terminal repetition. Proc. Natl.
Acad. Sci. U.S.A.69:3054-3057.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 2.completeddifferent Relative amounts of ['H]i fragments obtained by molecules with EndoR-](https://thumb-us.123doks.com/thumbv2/123dok_us/1567965.109389/4.493.258.447.352.591/fig-completeddifferent-relative-amounts-fragments-obtained-molecules-endor.webp)