0022-538X/81/030871-05$02.00/0 Vol.37, No.
Nucleotide
Sequence Changes
inPolyoma
ts-a
Mutants:
Correlation
with
Protein
Structure
PRESCOTT L. DEININGER, PATRICIA LAPORTE,ANDTHEODORE FRIEDMANN*
Department ofPediatrics, University of California, San Diego, La Jolla, California92093
The mutations in three polyoma ts-a mutants have been determined. Two
mutants,ts-25and ts-52,have different single-base changesatthesameposition
(2883) in the early region correspondingtoaconservedglycineresiduevery near
the C-terminus of the polyoma large T antigen. Mutant ts-48 hasasingle-base
changeatposition 2341,aswellas asecondchangeatposition 1228, in the region
oflarge Tantigen shared with medium T antigen.
A thorough understanding of the
structure-functionrelationshipof abiologically active
pro-tein requires detailed infonnation of the
enzy-matic and otherbiologicalfunctions, knowledge
of itsprimarystructurethrough amino acid
se-quencing,adescription of secondary and tertiary
structuralfeatures through X-ray
crystallogra-phy and otherphysicalmethods, the
identifica-tion of funcidentifica-tionaldomainsand residuesthrough
characterization ofmutants, and evolutionary
studies of functionally and genetically related
proteins to identify conserved regions of the proteins. Suchanalyses require readysourcesof
purified proteins
andwell-developed genetic
sys-temsin additionto assaysforprotein function.
Within the past several years, the
availability
ofpurifiedsimian virus 40(SV40) largeTantigen
(13,26) and thedescriptionof itsproteinkinase
and ATPaseactivities(11, 27), together withthe
likely availability of other papovavirus tumor
antigens,have
begun
tomakepossible
both anexamination of thestructure-function
relation-ships of thepapovavirustumor(T)
antigens
andan
understanding
of the mechanisms of celltransformation mediatedby these proteins. Of
majorimportance hasbeen the
development
ofmethodsforthe rapid determination of the
nu-cleotidesequence oflarge DNAmolecules (17, 21), by which the nucleotide sequences of the entire genomes ofSV40 (5, 19),
polyoma
virus(2, 8, 24), and BK virus (22) have been
deter-mined. These studies providedvaluable
infor-mation aboutthenature andstructure ofallof
the virus-coded proteins,
including
the Tanti-gens,longbeforepurified proteinsbecame
avail-able. Using nucleotide sequence information
alone,wehave identified
functionally
important regionsofthese molecules fromanidentificationof conserved regions in the viral genomes (7).
Otherworkers have examined the effects of
mu-tations at avarietyofpositionsin thegenomes
onthe function of viral geneproducts.
We wish to present here the nucleotide
se-quenceofpolyomagroupAmutantsthataffect
the structure and expression of the polyoma
large Tantigens and a correlation of the altered
sequences with characteristics of the protein
structure.
MATERIALS AND METHODS
Wild-type strain 3 polyoma is the strain whose complete nucleotide sequence has been described and analyzed in detail in thislaboratory (2, 8). The tem-perature-sensitive (ts) mutants ts-25, ts-48, and ts-52 were obtained from Walter Eckhart, TheSalk Insti-tute, La Jolla, Calif., and have been characterized extensively (3). All three mutants have the polyoma ts-aphenotype; i.e., they failto transform cellsat a nonpermissive temperature, andtheyare also
condi-tionallydefective in the induction oflargeTantigen
and in the inductionof viral DNAsynthesis.Allviral DNAswere prepared after infectionof 3T6cells by
previouslydescribed methods(9).
Preparationof clonedfragments.DNApurified
from themutants wascleaved with MboI and cloned
byligationinto theBamHI site ofplasmidpBR322;
selection ofampicillin-resistant colonies and detection ofappropriate polyomainsertsweredonebythe
hy-bridization method of Grunstein and Hogness (12),
using nick-translated (16) wild-type polyomaHaeIII fragment7as aprobe. Similarly,thePstIfragments
werecloned into the PstI site of pBR322, and inserts were detected as above by selecting tetracycline-re-sistantcolonies andbycolony hybridization with wild-typepolyoma HpaII fragment6.
Full-genome cloneswerepreparedbycleaving mu-tantDNAwith BamHI andligatinginto the BamHI siteofpBR322. Theresulting colonieswerescreened as described above. Plasmid DNA was prepared in SF8 Escherichia coli(4) and purifiedin cesium chlo-ride-ethidium bromidegradients (1, 20). HaeIII and
HpaII fragments were prepared, labeled, and
se-quencedasdescribedpreviously(9). All cloned
mate-rialswerepreparedand used in accordancewith Na-tionalInstitutes of HealthandUniversityof Califor-nia, SanDiego,Biosafety Committee guidelines.
Nucleotide sequencing. Details of thesequence
871
on November 10, 2019 by guest
http://jvi.asm.org/
872 DEININGER, LAPORTE, AND FRIEDMANN determination of wild-type polyoma have been re-portedpreviously (2, 8, 9). The nucleotide sequences of themutant DNAsweredeterminedbythe method of Maxamand Gilbert(17)andincluded theG-specific
methylene blue reaction for G(6)andthepotassium
permanganatereaction for T(C.Houck, personal com-munication). Details offragment preparation,labeling,
andpurification, gel electrophoresisand
autoradiog-raphy,informationstorage, and sequencecomparisons have beenpublishedpreviously from this laboratory (7-9).
RESULTS
Marker rescue experiments from other
labo-ratories havepreviously mapped the mutations
ofseveralgroup mutantsof
polyoma
(18). ts-a waslocalizedto afragment
between theHindIII siteatnucleotideposition1671 andanHhaI site atposition 2331, whereas mutants ts-25, ts-48,and ts-52 have all been localized to the HhaI
fragment spanning the
region
from positions2331 to 2913 bythenumbering convention used
by Friedmannet al. (9). Thesequencechanges
inthemutantsexaminedinthisstudyare
sum-marizedinTable1.Eachof themutantsshowed
onlyasingle mutation within the markerrescue fragment,indicating that the mutations
given
inTable 1 are responsible for the ts
phenotype.
Mutant ts-48is, in
addition,
knowntobemissing theHpaIIcleavage siteatthejunction ofHpaII fragments 7 and 8. Wehave sequenced acrossthis site from theMboI siteatposition1207and havefoundatransition from the
wild-type
Cto a T atposition 1228(W. Eckhart, S. Delbruck,P. Deininger, T. Friedmann, and T. Hunter, Virology, in press).
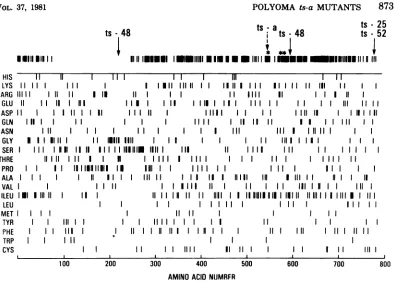
Figure1shows the amino acid distributionof the polyoma largeT antigen and the positions
of themutations determinedinthisstudy. The
positionsatwhichallthreepapovavirus
nucleo-TABLE 1. Nucleotide and amino acid sequence changes in polyoma ts-a mutants
Mutant Nucleo-
Nucleo-
Amino acidchangeand tide no. chne positionchange
ts-25 2883 G- T Gly-.Cys(largeT antigen residue 778)
ts-48 2341 G- T Ser- Illeu(largeT
antigen residue
597)
1228 C-T Thre- Ileu (large T
antigen residue 597)
ts-52 2883 G A Gly-+Ser(large T
antigen residue 778)
tide sequences are identicalinthelargeT
anti-genareindicated at the top of Fig. 1.
DISCUSSION
The ts-amutantscharacterized in thisstudy
were generatedby nitrous acid mutagenesis of
the polyoma genome, making it possible, and
evenlikely, that some of these mutants contain
multiple alterations in large T antigen and other
viralproteins, as in the case of ts-48, in which
wehave found one alterationaffectingonly large
Tantigen, oneaffectingboth thelargeTantigen
and the medium T antigen (see below), and
anotheraltering the VP2,3 terminator (Eckhart
etal., in press). The use of the marker rescue
data and analysis ofwild-type revertants makes
it possible to determine the region containing
the alterations responsible for the phenotypic
changes brought about by an altered large T
antigen (18). Inthe cases of allthree mutants,
there is only a single-base-pair change within
the mapped region, indicating that, for these
mutants, it is asingle amino acid change that
affects the tsphenotype and that other possible
alterations outside the marker rescue region
have little or noeffect onT-antigenfunction.
The mutation in ts-48 that confers the ts-a
ptenotype
is a transversion from aG to a T atposition 2341, resulting in a substitution of an
isoleucine residue for the normally occurring
serine residue inlargeTantigen (Table 1).This
mutation is inoneof themosthighlyconserved
regions of the papovavirus large T antigens, although the wild-type polyoma serineresidue
itself has not been conserved in the two other
papovaviruses (Fig. 1). The serine residue is
locatedamid anumber ofhydrophobicresidues,
mostly leucine and alanine,in a general region
that also contains a number ofthe positively
charged residues lysine and arginine (Fig. 1). Themoreshort-rangeenvironment, however,in
the immediatevicinity of thets-48mutationis
devoid ofchargedresiduesover a stretchof12
amino acids. Thus, the change ofaserineto an
isoleucinemayenrichthisalready hydrophobic region.
The ts-52mutation responsible for the altered
phenotype is a G-to-A transition at position
2883, very near the portion of the sequence
encoding the C-terminus of the T antigen at
position 2919. This region in polyoma virus
shows relatively poor conservation with the
other papovaviruses, but the alteration itself
doesrepresent thechangeof aconserved glycine
(Fig. 1)toserine. Mutant ts-25 contains a G -.
Ttransversion at the same position as the ts-52
mutation, resultingin achange from glycine to
cysteinein themutant protein. These alterations
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.488.50.240.483.645.2]POLYOMA ts-a MUTANTS 873
tS - 48
tSi
tS - 48 tS- 52I
111imin i I miiii0 NI mI IsI I11llEhil I--1 1 fI * I II* I E"l 1 1111111 I1I
HIS II 11n I
LYS 1111 I III
ARG GLU ASP GLN ASN GLY SER THRE PRO ALA VAL ILEU LEU MET TYR 1I
11111 1i I11 1
II 1111 1 11 11111 1111
liil l IIlIl l
I 1 1111 | 11 |I1
lI l il l l l l
1111 1 11 I1
PHE III
TRP I 11
CYS
100
11
I I
II
I mII iiiii
I I1 lI 11 II III 11111 I II liii I III
I 111 I IIII
III I 1111 III11 I I I
l111i 1111 11 11
11 II
1III
I 11 11I I
Hl1 I II 11
III1
mII
1iII I 11 11I II I 11 III I11 III' II I I IIrI
11111 11 III II
11 I
II II 11
I lMIi 11
II I I
III I 111111
II
I IIII1 1111 I I
lI III I I
I I I lI I
II I
I Hll1 III I III II Il
I I II III I I
11 I 11111111i 11 11111 111111 1 11111
II I II I I 11 11 II I I
11 III 11I
11 I l11111 I I I I I 11 III I I III
111 I I
I1I
II III I1l I
200 300 400 500 600 700 800
AMINO ACIDNUMRFR
FIG. 1. Amino acid distributionofthepolyoma largeTantigen. Thearrowsindicate theposition ofthe
mutations ints-25, ts-48,andts-52. Thepositionsthatareconserved inallthreepapovavirus largeTantigens
areindicatedatthe top. Thelargegaprepresents theuniqueportion ofthepolyoma large Tantigen. The
asterisksindicate thepositions ofthesequencechangesinfourSV40tsAmutants(23),andthedashedarrow
indicates thets-apolyomamutantcharacterizedbyThomas etal.(25).Thets-25sequenced byThomas etal.
is mutatedatthesamesiteasthets-25sequenceinourstudy.
occurin therelatively highlynegatively charged
C-terminus of thelarge T antigen and place a
serineresidue withinanegatively charged
clus-ter. The C-terminus of the wild-type large T
antigen contains eight negatively charged
resi-dues andonlyonepositivelycharged residue.
We have detected in ts-48asecondearly
re-gion point mutation outside the markerrescue
region, a C-to-T transition at position 1228
whichcausesachange from threonineto
isoleu-cine in large T antigen atamino acid position
219(Fig. 1), andachangefromprolinetoserine
inmedium Tantigen.This mutation lieswithin
adomainofthepolyoma largeTantigenthatis
unusual in several respects. Most notable are
theverymarkedclusteringofglycineandserine
residues, the large numberofprolineand
thre-onine residues, and the deficiency of charged
residues. This region contains aportionof the
coding regionfor the polyomamedium T
anti-gen,and since there is noequivalent regionin
theBK virusorSV40genomes,itcanbe
consid-ered aninsert, relative to those genomes. The
markerrescue results indicate that neither the
changeinmediumTantigennorthechangein
thelarge T antigen caused by the mutation at
position 1228 isresponsible for the ts-a
pheno-type.However,it ispossible that the medium T
antigen, notrequired for lytic infection of
per-missive cells,or evenlarge T antigen functions
arealtered ina mannernotdetected bythe lytic
infectionassayused for markerrescue.
Theregion of the polyoma medium T antigen
affected by the ts-48 mutationatposition 1228
is within the unusual glutamic acid-, aspartic
acid-, and proline-rich region of the molecule
that characterizes the C-terminal half of medium
T antigen. The mutation also lies within the
deletedregion of d11013 and d123 andnearthe
deletions ofd11014 anddll015, a series of
mu-tantsthat generally reduce the transformation
efficiency or the transformation phenotype in
rat cells (10, 14). We do not know whether
aspectsof the defectincelltransfornationof
ts-48mayberelatedtotheaminoacidsubstitutions
in medium Tantigen.
Analysis of the conserved regionsofthelarge
T antigens of the three papovaviruses reveals
further common andpossibly important
struc-tural features. In the 79 amino acid residue
VOL. 37, 1981
mm1iiimili
I
I
I I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.488.53.447.52.334.2]874 DEININGER, LAPORTE, AND FRIEDMANN
region of thelargeTantigenN-terminaltothe
proximal large Tsplice, none of the 13 amino acid differences between BK virus and SV40
occur atpositions thatareconserved inpolyoma
virus and SV40. The latter conserved residues
thereforeseem toberelatively important. Sim-ilarly,throughouttherestof the
large
Tantigen molecules, most of the amino acid differencesbetween SV40 and BK virus alsooccurat
posi-tionsnotconservedin
polyoma
virus andSV40,
indicatingthat the conserved sitesare
important
forproteinfunction.Of the22aminoacid
differ-ences between BK virus and SV40 atresidues
conserved in polyoma virus and
SV40,
14 are functionally conservative, i.e., Lys -. Arg,Asp-*Glu, hydrophobic-*
hydrophobic.
Thesefindings suggest that most of the amino acid
residues of thelargeTantigens
play
important and similar structure-function roles and are thereforeconserved.Therearetwosmallregions
that may beexceptions,onejust N-terminalto
the ts-48 mutation at nucleotide 1228, where
several ofthe
polyoma-SV40
conserved residuesarealteredin BKvirus,and the otheratthe C-terminal tailin thevicinityof the ts-52mutation.
Thomas and hercolleagueshavedetermined
thenucleotide sequencealterations
responsible
for thets
phenotype
intheprototypical
group Amutant ts-aand ints-25
(25).
Theirts-25showsa change identical to that described
here,
whereasts-acontainsa
change
atposition
2193, correspondingtoamino acid547inFig. 1.The mutations in four tsA mutants ofSV40
havebeen identified
by
Seifetal. (23),and thepositions of those mutations are indicated by asterisks in
Fig.
1. All four fall in part ofthesequence near that ofpolyoma mutants ts-48
and ts-a, as identified
by
Thomas et al. (25),suggesting particular importance for that por-tionofthemolecule.It isof interesttonotethat, whereas the mutationsin ts-48 andts-25 cause
changes to more hydrophobic amino acid resi-dues, the mutation ints-52results in a change fromglycinetoserine,tworesidues withlittleor nodifference in theirhydrophobicity(15).
ACKNOWLEDGMENTS
WethankWilliamFolk forallowing us to see his results beforepublication and Walter Eckhart, Tony Hunter, and Russell Doolittle for their helpful discussions. We also thank H.U.Bernardforthets-52whole-genome clone, Abby Esty for technicalassistance,andDorothyMillerforadministrative help.
Thiswork was supported by Public Health Service grant CA24288 from theNational Cancer Institute, and P.L.D. was
supported byPublic HealthService training grant GM07199 from the National InstitutesofHealth.
LITERATURE CITED
1. Clewell,D.,andD.Helinski.1969.Supercoiled
DNA-proteincomplex in E. coli. Proc. Natl. Acad. Sci. U.S.A. 62:1159-1166.
2. Deininger, P., A. Esty, P. LaPorte, and T. Friedmann. 1979.Nucleotide sequence andgenetic organization of thepolyoma late region: featurescommon tothe poly-omaearly region and SV40. Cell 18:771-779. 3. Eckhart, W. 1974. Properties of temperature sensitive
mutants ofpolyoma virus. ColdSpringHarborSymp. Quant. Biol. 39:37-40.
4. Enea, V., G.Vovis, and M. Zinder. 1975. Genetic studies withheteroduplex DNA of bacteriophage Fl. J. Mol. Biol. 96:495-509.
5. Fiers, W., R.Contreras, G. Haegeman, R. Rogiers, A. Van deVoorde, H. VanHeuverswyn,J.Van Her-reweghe, G. Volckaert, and M. Ysebaert. 1978. Complete nucleotide sequence of SV40 DNA. Nature (London) 273:113-120.
6. Friedmann, T.,and D. Brown.1978.Base-specific re-actionsuseful for DNA sequencing. Nucleic Acids Res. 5:615-622.
7.Friedmann, T., R. Doolittle, and G. Walter. 1978. Aminoacid sequencehomology between polyoma and SV40 tumor antigens deduced from nucleotide se-quences. Nature(London) 274:291-293.
8. Friedmann, T.,A.Esty,P.LaPorte,and P.Deininger.
1979.Thenucleotide sequence and genome organization ofthepolyoma early region: extensive nucleotide and amino acidhomologywithSV40. Cell 17:715-724. 9. Friedmann,T.,P.LaPorte, and A. Esty. 1978.
Nucleo-tide sequence studies of polyoma virus. J.Biol. Chem. 253:6561-6567.
10. Griffin, B.,andC. Maddock.1979.Newclasses of viable deletionmutantsin theearlyregion ofpolyoma virus. J.Virol. 31:645-656.
11. Griffin, J., G. Spangler, and D. Livingston. 1979. Protein kinaseactivity associated with simian virus 40 Tantigen. Proc. Natl. Acad. Sci.U.S.A. 76:2610-2614. 12. Grunstein, M.,and D.Hogness.1975.Colony
hybridi-zation:amethod for theisolationofcloned DNAs that contain aspecificgene.Proc. Natl. Acad.Sci. U.S.A. 72:3961-3965.
13. Kriegler,M.,J.Griffin,and D.Livingston.1978. Phe-notypiccomplementation of the SV40 TS A mutant defect in viral DNA synthesisfollowingmicroinjection ofSV40antigen.Cell 14:983-994.
14. Magnusson,G., and P.Berg. 1979.Construction and analysis of viable deletionmutantsofpolyoma virus. J. Virol. 32:523-529.
15. Manavalan, P., and P. Ponnuswamy. 1978. Hydropho-bic character of amino acid residues inglobular proteins. Nature(London)275:673-674.
16. Maniatis, T., A.Jeffrey,and D. Kleid. 1975. Nucleotide sequence of the rightward operator of phageA.Proc. Natl. Acad. Sci. U.S.A. 72:1184-1188.
17. Maxam,A.,and W.Gilbert. 1977.A newmethod for sequencing DNA. Proc. Natl. Acad. Sci. U.S.A. 74:560-564.
18. Miller,L.,and M.Fried. 1976.Construction of the ge-neticmapofthepolyoma genome. J. Virol. 18:824-832. 19. Reddy, V., B.Timmappaya, R. Dhar, K. Subraman-ian, B.Zein,J.Pan,J.Celina, and S. Weissman. 1978. The genome of SV40. Science 200:494-502. 20. Redloff, R., W. Bauer, and J. Vinograd. 1967. A
dye-buoyant density method for the detection and isolation ofclosedcircular duplex DNA. Proc. Natl. Acad. Sci. U.S.A.57:1514-1521.
21. Sanger,F., S.Nicklen, and A. Coulson. 1977. DNA sequencing with chain terminating inhibitors. Proc. Natl.Acad.Sci. U.S.A. 74:5463-5467.
22. Seif, I., G. Khoury, and R. Dhar. 1979. The genome of human papovavirus BKV.Cell18:963-977.
23. Seif, I., G. Khoury,and R.Dhar. 1980. A rapid
enzy-J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
matic DNAsequencing technique. Nucleic Acids Res. 8:2225-2240.
24. Soeda, E., J. Arrand,N.Smolar,J. Walsh,and B.
Griffin.1980.Coding potentialandregulatorysignals of thepolyomavirus genome. Nature (London) 283: 445-453.
25. Thomas, T., P.Vollmer, and W. R. Folk. 1980.
Nucleo-tidesequencechanges in polyoma virus A gene mutants. J.Virol. 37:1094-1098.
26. Tjian, R. 1978. Thebindingsite onSV40 DNA for a T antigen-related protein. Cell 13:165-179.
27. ITian,R., and A.Robbins. 1979.Enzymatic activities associated withpurified simian virus 40 T antigen re-lated protein. Proc. Natl. Acad.Sci. U.S.A. 76:610-614.