Copyright © 1993, AmericanSocietyforMicrobiology
The NF-KB
Binding Site Is Necessary
for
Efficient
Replication
of Simian
Immunodeficiency
Virus of
Macaques
in
Primary Macrophages
but Not
in
T Cells In
Vitro
ROBERT E.BELLAS,1 NANCY HOPKINS,'ANDYENLI2t*
CenterforCancerResearch, Massachusetts InstituteofTechnology, Cambridge, Massachusetts 02139,1and
Department ofMicrobiology and MolecularGenetics, NewEnglandRegionalPrimate Research Center, HarvardMedicalSchool, Southborough, Massachusetts017722
Received7 October1992/Accepted 16February 1993
We demonstrate here that the nuclear factor-KB(NF-KB) bindingsite inthe simianimmunodeficiencyvirus
(SIVmac) longterminal repeat is essential for efficient virusreplicationinprimaryalveolarmacrophagesbut
dispensable for efficientreplication in primaryTcells. Mutation of the NF-KB site doesnot seriously impair
replication ofaT-cell-tropic SlVmac239or amacrophagetropicSIVmacEm* inperipheralbloodlymphocytes orestablishedCD4+celllines; however,mutation ofthe NF-KB siteprevents efficient SlVmacEm*replication
in primary alveolar macrophages. These data suggest that efficient replication in primary macrophages requiresbothenvelopeandlongterminal repeat determinants.
Simianimmunodeficiencyvirus (SIV)isaprimate lentivi-rus closely related to the human immunodeficiency virus
(HIV) (3, 7)withrespect tosequence,genomicorganization, tropism for CD4+ cells, capacity to establish persistent infections of the host, and, for some viruses, ability to inducean immunodeficiency disease. Rhesus macaques
ex-perimentally inoculated with the molecularly cloned SIV-mac239developwithin0.5to2yearsasyndrome character-ized by AIDS-like features, including immunosuppression, opportunistic infection, CD4+ T-celldepletion, emaciation,
andencephalopathy (4, 14, 27). SIV in the rhesusmonkeyis
quitepossibly the best animal model for AIDS research. Isolates of SIV and HIVvarydramaticallyintheirabilities to infect primary macrophages (2, 4, 8, 15, 28). Because
infectionofmacrophagesmaybeimportantinallowingthese viruses to establish long-term persistent infections of the hostand alsomaybeinvolved in certain disease
manifesta-tions,inparticularin centralnervoussystemdisorders(fora
review, seereference 9),there has been interest in
identify-ingsequencesthat affectreplicationinmacrophages. Recent
studies havedemonstrated thatenvelope sequencesplayan
importantrole inmacrophage tropism (12, 24, 31);however,
no evidence has suggested thatlong terminal repeat (LTR)
sequences can differentially affect replication in
macro-phages versusTcells.
Allisolates of HIV andSIV of whichwe areawarepossess in their LTRs at least one perfectly conserved copy of a
bindingsite for the nuclearfactor-KB(NF-KcB). Many
exper-iments have demonstrated that the NF-KB site is important
for inducible expression of HIV type 1 (HIV-1) LTR-re-porter constructs in transient-expression assays in T cells
(22, 25). However, Ross et al. (29) and Leonardet al. (16)
found that deletion of the NF-KB sites hada modestor no effectonHIV-1replicationinperipheral blood lymphocytes
(PBLs). This result is rather surprising in light of the high degreeofconservationoftheNF-KBbinding site. To
deter-* Correspondingauthor.
tPresentaddress:DepartmentofMicrobiologyandImmunology, UniversityofMaryland atBaltimore, 655 WestBaltimore Street, Baltimore,MD21201-1559.
mine the role of the NF-KB site in SIV replicationinboth T cells and macrophages, we constructed an SIV possessing LTRs lacking the SIV NF-KcB site and examined itsgrowth
properties.
MATERUILSANDMETHODS
Cell culture. MT-2, MT-4, C8166, and CEMx174 cells
were maintained in RPMI 1640medium supplemented with 10% heat-inactivated fetal bovine serum, 2 mM glutamine,
and50 U of penicillin and 50jigof streptomycinperml.The
cellswere maintained by twice weekly dilution to 5 x 105
cellsperml. Rhesusmacaque(Macacamulatta) PBLswere
obtained by banding in sodium diatrizoate-Ficoll mediumas described elsewhere(14).PBLsweretreated for48hwith1
,ug ofphytohemagglutinin permlofthemediumusedfor the establishedcelllines. After 48h,thePBLswerewashedand
resuspendedin culturemediumcontaining 10% interleukin-2 (Pharmacia)at aconcentration of 106 cellsperml. Cultures of alveolar macrophageswereobtained by bronchoalveolar
lavage from rhesus macaques as described elsewhere (4)
with RPMI 1640 medium supplemented with glutamine and antibioticsasdescribedaboveand10% HIV-negative human serum(GIBCO)at5x 105cellsperml in24-or48-well plates
(Falcon).
Generation of virus.Plasmid DNAsforthe 5'and3'halves
of the SIVmac genome were digested with the restriction enzyme SphI. These two halveswere then ligated withT4 DNA ligase and transfected with DEAE-dextran as de-scribed elsewhere (14) onto CEMx174 cells. Virus-contain-ingsupernatantswerecollected, clarified bylow-speed
cen-trifugation (500 x g), filtered through 0.45-,um-pore-size membranes, aliquoted, and stored at -80°C. Virus was
quantitatedby a p275a5 antigen capture assaywith a
com-mercially available enzyme-linked immunosorbent assay
(ELISA)kit(Coulter, Hialeah, Fla.).
Site-specific mutagenesis. Generation of site-specific muta-tion of both the 5' and3'LTRs ofSIVmac239 nefopen(239) andSIVmacEm* nefopen (Em*) was achieved by using a
four-primer polymerase chain reaction (PCR) protocol as described elsewhere (12). The final product ofthe second-2908
on November 9, 2019 by guest
http://jvi.asm.org/
into a BamHI- and HincII-digested Bluescript pBS KS-plasmid (Stratagene). One resulting LTR subclone was then used to regenerate the 5' half bysubcloning anNdeI-HincII
fragment into anNdeI-HincII-digested parental genomic 5' half; the 3' halfwasgenerated by subcloning anNdeI-HincII and EcoRI fragment in the polylinker into an NdeI (partially digested)-EcoRI 3' end of 239. To incorporate this LTR into the 3' half ofEm*, the SacI-EcoRI fragment of the 239CTC was ligated intoan SacI-EcoRI-digestedEm*.
Preparation of nuclear extracts. Nuclear extracts were
prepared by a modification of a procedure developed by Christine Jamieson (13a). A total of 5 x 107 cells were
pelleted by low-speed centrifugation (500 xg) and washed threetimes in ice-cold phosphate-buffered saline. The cellu-lar pellet was then gently resuspended in 0.5 ml of 0.5% Nonidet P-40-10 mM Tris hydrochloride (pH 7.6)-i mM EDTA, and the suspensionwas held at 4°C for 5 min. The nucleiwere collectedbycentrifugation (500 xg) and
resus-pended in 100 ,ul of buffer C (17). The nuclei were then vortexed vigorously for 2 min. The extracts were then
centrifugedat12,000 xgfor 5 minat4°C.The supernatants werethenaliquoted and stored fornolonger than 3 weeksat
-800C.
Electrophoretic mobility shift assays. Electrophoretic
mo-bility shift assayswereperformedessentiallyasdescribedby Manleyet al. (17)with Tris-borate asthe bufferingsystem. Complementary oligonucleotides were end labeled with T4 polynucleotide kinase and
[y-32P]ATP,
annealed, and sepa-rated from unincorporated label bypolyacrylamide gel elec-trophoresis. Binding reactions(25-,ulfinalvolume) employed10,000 cpm of probe, 5 ,ug of nuclear extract, 100 ng of
poly(dI)-poly(dC), and 100 ng of sonicated, denatured salmon spermDNA. Unlabeledcompetitor oligonucleotides
were present at 100-fold molar excess. The complexes
formed werevisualized by electrophoresis through
nonde-naturing polyacrylamide gels and autoradiography of the driedgels. The sequences of the sense strand of the
oligo-nucleotides usedwere as follows: NF-KB, GCAGGGACTT
TCCACAA;CTC,
GCACTCAC'TTTCCACAA;
AA,GCAGGGACTTlTAAACAA; andSpl, ACAAGGGGATGTTACG
GGGAGGTACGGGGAGGTACGGGGAGGAGCC.
Virus infections. All infectionsemployed 4-ngequivalents
of virus stock that were prepared simultaneously for any
giveninfection under conditionsoptimumfor virus infectiv-ity. CEMx174 cellsweretransfected withproviralDNAand
passagedevery3days.At the time of maximumcytopathic effect, the cells were washed and resuspended in fresh medium.Twenty-fourhourslater, the supernatantfluidwas harvested. Under these conditions, the infectivity of the
stocks, independentoftheNF-KBmutation,is 50 50% tissue culture infectious doses per ng ofp279ag protein astitrated
on CEMx174 cells. For the established cell
lines,
4 ng ofp279agcoreantigenequivalentswasadsorbedto2x 106cells in 1 ml of tissue culture medium (RPMI 1640 plus 10%
heat-inactivated fetal bovineserum) for 2 h. The cellswere then washed three times with phosphate-buffered saline to removeunadsorbed virus andwerecultured. The cellswere
splittwo orthreetimesweeklyto a densityof5 x 105cells per ml. The amountofvirusinthecell-free supernatantwas
determinedbyantigen-capture ELISA (Coulter). For infec-tion of PBLs, rhesus macaque (M.
mulatta)
PBLs were isolated bybanding in sodiumdiatrizoate-Ficoll medium as describedpreviously (14), stimulated for 48 h with 1 ,ug ofphytohemagglutinin perml,washed, and infected with4ng of p27 equivalents of virus for 2 h. The cells were then
Q c* c
competitor ° (5:P
fi
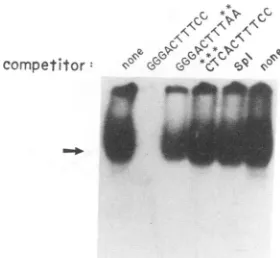
-FIG. 1. AGGG-to-CTCmutation lacks theabilitytocompete for nuclear factors thatnornallybind the 239 NF-KB site.An electro-phoretic mobilityshift assayemployingadouble-stranded
oligonu-cleotidecorrespondingtothe SIVNF-KB siteasprobe
(GGGACTT
TCC) to detect factors found in T cells (C8166 extracts) wasperformed as described in Materials and Methods. The indicated unlabeled oligonucleotide competitors were used at 100 molar
excess. Asterisks indicate positions at which the oligonucleotide
differs fromthewild-type239 NF-KB site.Spl, anoligonucleotide
correspondingtothe SIVSplbindingsites. Thearrowindicates the positionofmigration of the protein-probecomplex. Free,
uncom-plexed probeisnotshown.
washed three times and cultured in the presence of
purified
humaninterleukin-2
(10%)
(Pharmacia).
The cellsweresplit
to 106cells per ml twiceweekly. Macrophage
cultureswere infectedby
incubation of the cells with4-ng
equivalents
of virusovernight,
followedby
washing
withphosphate-buff-ered saline and
culturing
in the medium asdescribed above for cell culture.RESULTS
We
employed
afour-primer
PCR-based in vitro mutagen-esisprocedure
to generate an LTR subclone which con-tained aGGiGACTITCC-to-CTCACTITTCC
(CTC)
triple-point
mutation in theNF-KBbinding
site. Thismutation has been showntoabrogate
binding
ofNF-KB-relatedproteins
to the HIV-1 LTR(6)
and showednoability
at100-fold molar excess tocompete for factors found in T cells thatnormally
bind the
wild-type
SIV NF-KB site(Fig.
1).
After DNAsequencing
of the entireregion
amplified by
PCRconfirmed thatnoundesiredchanges
had occurredduring
themutagen-esis
procedure,
viruswasgenerated
aspreviously
described(14)
into CEMx174 cells to generate theT-cell-tropic
239lacking
an NF-KB site(239CTC)
or themacrophagetropic
virus
lacking
thissite,
Em*CTC.We
compared
the 239CTC mutant with 239(wild-type)
viruswith respecttotheir abilitiestogrow inthree
CD4+
cell lines known to supportreplication
of 239.Figure
2 demon-strates that 239 and the 239CTCmutant show no dramatic differences inpeak
virusproduction
inCEMx174, MT-2,
orMT-4 cell
lines;
239CTC virusproduction lags
slightly
be-hindthat of 239 in MT-2 cellsbutnotin CEMx174orMT-4 cells. This
lag
wasobservedinthreeseparateinfectionsand isconsistent with results observedby
Rossetal.(29)
thatincertain T-cell types the HIVNF-KB site may
play
arole inreplication.
239CTCvirus,
likethewild-type 239,
leadstoalytic
infection characterizedby
syncytium
andgiant
cell formation(data
notshown). Therefore,
theNF-KBsite ofthe--O.
on November 9, 2019 by guest
http://jvi.asm.org/
[image:2.612.370.510.79.208.2]a
4-
3-
2-
1-
0-10 20 30
4i
3-
2-
1-
0-l
10 20 30 10 20 30
days post-infection
days
post-infection
days
post-infection
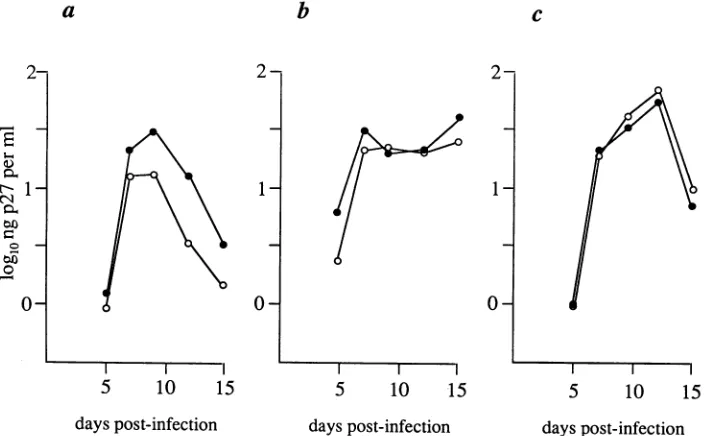
FIG. 2. The NF-KB site has modest effectsongrowthofSIVin three establishedCD4'cell lines. Virus in the cell-free supernatantwas assayedbyantigen-captureELISA(Coulter). (a)CEMx174; (b) MT-4; (c)MT-2.Opencircles,239CTC(noNF-KBsite); closedcircles,239 (one NF-KB site[wild type]).
T-cell-tropic 239 appears to play a modest (ifany) role in viral replication in these established cell lines.
Wedetermined theeffect of the SIVNF-KcBsite inprimary
cultures of rhesus macaque PBLs(Fig. 3). PBLsfrom three different rhesus macaques were stimulated for 2 dayswith
phytohemagglutinin, infected with either the 239CTCorthe 239virus, and then cultured in the presence ofinterleukin-2. Nosignificant difference between 239 and239CTCregarding
theirgrowth inPBLs was seen.Therefore, in agreement with the resultsseenforHIV-1deleted of its NF-KBsites
(16, 29),
anSIVlackinganNF-KBsitereplicatesquitewell in
primary
CD4+ Tcells.a
b
We considered thepossibilitythat the lack ofaneffect of the NF-KBsite incertain T cells is duetorapidreversion of the sequence to thewild type. However, this seemshighly unlikelybecause (i)we usedatriple-point mutation, which might beexpectedtobesomewhat difficultto reverteasily; (ii)thephenotypesof the virusasshown in the present paper
are highly reproducible and independent of the particular
DNA stock orvirus stockused; and (iii) the lack of
differ-encebetween thewild-typeandmutantswas seen
indepen-dently ofmultiplicityofinfection. Inexperimentsnotshown,
wehave observednodifferencesingrowth kinetics between the wild-type and NF-KB mutant virus in CEMx174 cells
C 2
5 10 15
2-
0-5 10 15 5 10 15
days post-infection days post-infection dayspost-infection
FIG. 3. The NF-KB site hasnoeffectongrowthof SIV ininterleukin-2-stimulatedrhesusmacaquePBLs.(a,b,andc)Growthcurves on
PBLsfrom individual macaques. Opencircles, 239CTC(noNF-KBsite); closedcircles, 239(oneNF-KBsite [wildtype]).
b
c
4- 3.-E 3
0-to
X._
0
2-E
a)
0~
Cl.
to02
-
on November 9, 2019 by guest
http://jvi.asm.org/
[image:3.612.126.477.76.287.2] [image:3.612.127.481.482.700.2]a
0
.17AA-,
j.
;
2-- 1
-N
04
to
0
0-
2-
1-
0-10 20 30
days post-infection
FIG. 4. The NF-KB site is important
primary rhesusmacaque primary alveolar
Growth curves on macrophages from indi circles,Em*CTC (no NF-KB site); closedc
site[wild type]).
ti
3 gr
a e.IIS$@@ @ ...
/L.
- ', a
a & J A _
_ .1*. r I
a - j
,
!- v
I~-~~~~~~G r;ta
10 20 30
days
post-infection
for growth of Em* in -macrophages. (a and b)
vidual macaques. Open
ircles,Em*(oneNF-KB --'
-3,~~~~~~~~~~~~~~~~~,C
when infections were initiated with400, 40, 4, or 0.4 ngof
p27"ag. If the mutants were reverting, one would perhaps
expectthatatalowmultiplicityofinfectiononemightsee a
longer lagbefore themutantstartedtogrowwell; however, we neverobserved anydifferences, suggesting that thelack of the NF-KB mutation in T cells seems to be almost certainlyareflection of the inputgenotype.
Recent evidence suggests that cells of the monocyte-macrophage lineage may serve as areservoir for virus and
playanimportant roleinallowing the virustopersist in vivo (11, 22, 30). Mori etal. recently showed that Em*, avirus
which differs from 239 by only nine amino acid changes in
theenvelopegene,will efficiently replicate inprimary
mac-rophages (21).Weintroduced the CTCmutantLTR into this macrophagetropic Em*, creating Em*CTC, and we
com-pared Em* andEm*CTCwithrespecttogrowthin primary rhesus macaque alveolar macrophages and T cells. As a
control,wealso tested thegrowthof both viruses in Tcells,
since Em* grows in these as well. Em*CTC replicates
extremely poorlyinmacrophages,withpeaklevels from5to 10% of those of Em* (Fig. 4). In an experiment involving
macrophages plated at a higher density, Em*CTC core
antigen levelswere less than 1% ofthat produced byEm*
(datanotshown).
Thereare someconditionsunder which HIVcanreplicate without being released into the medium (32). In this case,
however, there is cell death andsyncytium formation.
Fig-ure 5 documents that in addition to the lack of virus
production by p279ag assay, an independent criterion, i.e.,
cytopathic effects induced by the virus, seems to suggest that the virus withoutNF-KB replicates less. Em* leads to giant cell formation and cell lysis, while Em*CTC has no detectable cytopathic effect in macrophages (Fig. 5). This
Em*CTCvirus isfullyreplicationcompetent, because
com-parison of itsgrowthwith Em* in CEMx174orMT-4 cells
(data not shown) or primary PBLs (Fig. 6) reveals no or modest differences in growth capacity. Therefore, the SIV
e m e &!* ° t e ' -c'
C'L,'. .~ *. A a,.
II~~44~4 4V14 4 41'
[image:4.612.323.552.71.549.2]0,1.0 0 :4 42 0 ,
FIG. 5. EmCT is less cyoati thnE*imcohg
&4'
priary macophge i4vito. ~ '
4 i a
This~
is,0th~
first~~p
deosrto of a.poetefetoha 0~~~~ ,a0
NFB
site
fo via elcato npi ycls htti 4t'~~ ~ ~~~~4e.fcisi m a i
*~~~~~~~~~~~~~~~~jak.-0.
[image:4.612.61.298.76.289.2]0 ~ ~ ~ ,a 44e
FIG. 5.EmiCTC is less cytopathic than Em* in macrophage
cultures. Cultureswerephotographedwithalow-power objective15
daysafterinfection withEmn* (a), Emn4CTC (b),or novirus(c).
NF-KB site appears to be
required
forefficientreplication
inprimary
macrophages
invitro.DISCUSSION
This is the first demonstration ofa potent effect of the NF-KB site for viral
replication
inprimary
cells. That this effect is inmacrophages
isintriguing
in thelight
ofrecentfindings.
First ofall, fornonprimate
lentiviruses, theability
to infect and
replicate
well inmacrophages
is considered crucial to theability
to establish along-term
persistent
infection(11, 23).
Inaddition,
Schuitemakeret al.(30)
haveI
24
on November 9, 2019 by guest
http://jvi.asm.org/
a
b
c
2-
1-
0-
2-
1-0
-5 10 15 5 10 15
1
5 10 15
days post-infection days post-infection days post-infection
FIG. 6. The NF-KB site isnotessential for efficientreplicationof Em* in PBLs.(a, b,andc)Growthcurvesof Em*orEm*CTC in PBLs from individualmacaques. Open circles, Em*CTC; closedcircles, Em*.
recently demonstrated that during the long-term persistent stageof HIVinfection, macrophagetropicvariants
predom-inate. Moreover, viruses isolated from PBLs during this stage replicate well in macrophages also, suggesting that
they have recently entered the T-cell population from a
monocyte reservoir. Only in the late stages of disease do
T-cell-tropicvariantscometopredominate.Schuitemakeret al. suggestthat macrophages andmonocytes serve asvirus reservoirs and mediatepersistence. Iftheir model is correct,
then sequences that influenced replication inmacrophages
would beexpectedtobeveryhighly conserved, asis indeed the NF-KBbinding site.
Much evidence supportsarole forspecificLTRsequences
in HIV-1replication inmonocytes-macrophages. For
exam-ple, the latently HIV-1-infected Ul monocytic cell line
responds to NF-KB inducers by greatly increasing viral
production (5, 10); the monocytic cell line THP-1 can
de-velop restricted expression of HIV-1 and under these cir-cumstances contains anuclear inhibitor of NF-KB activity; in this system, the transfection of an NF-KB expression
vectorovercomesthe restriction and increases virus produc-tion (19); and lipopolysaccharide, an NF-KB inducer, in-creases HIV replication in monocytes (26). In addition,
HIV-1 infection ofU937monocyticcells leadstoanincrease inNF-KBactivity, perhaps byaction of the viralproteaseon aninactive NF-KBprecursor (1). Finally, although
conflict-ing reportsexist, the HIVenvelope proteinmay be ableto stimulatemonocyticcells toproduce tumor necrosis factor
alpha,which is known toactivate NF-rB (18, 20).
The results of studies mapping determinants of
macro-phage tropism (13, 24, 31) have led to the suggestion that envelope sequences are the primary determinant and are sufficient forefficientreplicationinmacrophages. Moreover,
no evidence hasbeen seen fora role of LTRsequences in
macrophage tropism. However, in the studies described above, the NF-KB site in both the T-cell-tropic and the macrophagetropic parents waswildtype; thus, a contribu-tion of the NF-KB site to macrophage tropism would not have been detected in these experiments. In addition, the
experimentsdescribedbyLeonardetal.(16)andbyRosset al. (29) studying the effect of the NF-KB site on HIV-1
growth used a clone (NL-43) that does not replicate in
primary macrophages; therefore, these investigators were
notableto determine the contribution of theNF-KB siteto HIV-1 growth in primary macrophages. In light of data
suggestinganimportantrole formacrophagetropicviruses in
HIV and SIVpersistence, it is temptingto speculate thatin vivo selective pressure for maintenance of the NF-KB site may be a reflection of its importance for replication in
macrophages.
It is quite possible, however, that in addition the NF-KB
site doesplayarole inreplicationin Tcellsinvivoand that the in vitro conditionsemployedhere donotreflect that role. Thestateof activation andgrowthfactorspresent, etc.,may
bevery different between PBLs in culture and in vivo; in
addition, the way in which virus spreads and grows in in vitro culturemaybeverydissimilartowhathappensin vivo.
Indeed,the viruslacking NF-KB did showareproducible lag in kineticscomparedwith that of thewild-typevirus in MT-2 cells. This result is consistent with results obtainedbyRoss et al. (29), who noticed thatwith respect to virus growth kinetics,the HIV-1 NF-KB site had differenteffects, depend-ing on the particular T-cell lines employed. One other
possibilityisthat the NF-KB site isimportantin reactivation of latent virus in Tcells,andgrowthcurves asperformedin thepresent studyandbyothers (16, 29) do notreflect that
activity.
However,the fact thatwehaveemployedthepathogenic molecular clone 239 in the present studies will allow us to experimentallyinfectrhesusmacaquestodetermine the role of NF-KB in viral persistence and pathogenicity. One pre-diction of the resultsreportedherewouldbethatin rhesus macaques infected with the 239CTC NF-KB mutant virus,
SIV willreplicate onlyintheT-cellcompartment; an
inabil-itytoefficiently replicateinmacrophages mightpreventviral persistence. Careful monitoring of virus and viral DNA present in PBLs and macrophages at different times after
infection would allowtesting of this prediction.
1.
0
0
o
O
on November 9, 2019 by guest
http://jvi.asm.org/
[image:5.612.129.488.75.298.2]Wethank both RobertBeckerfortaking the photographs used in Fig. 5 and Kent Hunter. In addition, we thank K. Mori for graciously providing us with the Em*clone prior topublication. We express our gratitude to the Department of Primate Medicine, in particular to Prabhat Sehgal at the NewEngland Regional Primate Centerfor obtaining blood and lavagefluidsfrom the macaques.
R.E.B. is a recipient of a National Institutes Fellowship. Y.L. gratefully acknowledges grants A125108 and CA50146 from the National Institutes ofHealth.
REFERENCES
1. Bachelerie, F., J. Alcami, F. Arenzana-Seisdedos, and J. L. Virelizier. 1991. HIVenhancer activity perpetuated by NFKB induction on infection ofmonocytes. Nature (London) 350:709-712.
2. Banapour, B., M. L. Marthas, R. J. Munn, and P. A. Luciw. 1991. In vitro macrophage tropismof pathogenic and nonpatho-genic molecular clones of simianimmunodeficiency virus (SIV-mac). Virology 183:12-19.
3. Desrosiers, R. C. 1990. The simian immunodeficiency viruses. Annu. Rev. Immun.8:557-578.
4. Desrosiers, R. C., A. Hansen-Moosa, K. Mori, D. P. Bouvier, N.W. King, M. D.Daniel, and D. J. Ringler. 1991. Macrophage-tropic variants of SIV are associates with specific AIDS-related lesions but are not essential for thedevelopment of AIDS. Am. J. Pathol. 139:29-35.
5. Folks, T. M., J. Justement, A. Kinter, C. A. Dinarello, and A. S. Fauci. 1987. Cytokine-induced expression of HIV-1in a chron-ically infected promonocyte cell line. Science238:800-804. 6. Franza, B. R., Jr., S. F. Josephs, M. Z. Gilman, W. Ryan, and
B.Clarkson. 1987. Characterization of cellular proteins recog-nizing the HIV enhancer using a microscale DNA affinity precipitation assay. Nature (London) 330:391-395.
7. Gardner, M. B., and P. A. Luciw. 1989. Animal models of AIDS. FASEB J. 3:2593-2606.
8. Gartner, S., P. Markovits, D. M. Markovitz, M. H. Kaplan, R. C. Gallo, and M. Popovic. 1986. The role of mononuclear phagocytes inHTLV-IIIILAVinfection. Science233:215-219. 9. Gendelman, H. E., J. M. Orenstein, L. M. Baca, B. Weiser, H.
Burger, D. C. Kalter, and M. S.Meltzer. 1989. The macrophage in the persistence and pathogenesis of HIV infection. AIDS 3:475-495.
10. Griffin, G. E., K. Leung, T. M. Folks, S. Kunkel, and G. J. Nabel. 1989.Activation of HIV gene expression during mono-cyte differentiation by induction of NF-KB. Nature (London) 339:70-73.
11. Haase, A. T. 1986. Pathogenesis of lentivirus infections. Nature (London)322:130-136.
12. Ho, S. N., H. D. Hunt, R. M. Horton, J. K. Pullen, and L. R. Pease. 1989. Site-directed mutagenesis by overlap extension usingthepolymerase chain reaction. Gene 77:51-59.
13. Hwang, S. S., T. J. Boyle, H. K.Lyerly,and B. R.Cullen. 1991. Identification of the envelope V3 loop as the primary determi-nantof cell tropism in HIV-1. Science 239:71-74.
13a.Jamieson, C. Personalcommunication.
14. Kestler,H., T.Kodama, D. J. Ringler, M. Marthas, N. Pedersen, A. Lackner, D.Redier, P. Sehgal, M. Daniel, N. King, and R. Desrosiers. 1990. Induction of AIDS in rhesus monkeys by molecularly cloned simian immunodeficiency virus. Science 248:1109-1112.
15. Koyanagi, Y., S. Miles, R. T. Mitsuyasu, J. E. Merrill, H. V. Vinters, and I. S. Y. Chen. 1987. Dual infection of the central nervoussystem by AIDSviruses with distinct cellular tropisms. Science 236:819-822.
16. Leonard,J., C.Parrott,A. J.Buckler-White, W. Turner, E. K. Ross, M. A. Martin, and A. B. Rabson. 1989. The NF-KB binding
repeat are not required for virus infectivity. J. Virol. 63:4919-4924.
17. Manley, N. R., M. A. O'Connell, P. A. Sharp, and N. Hopkins. 1989. Nuclear factors that bind to the enhancer region of nondefective Friend murine leukemia virus. J. Virol. 63:4210-4223.
18. Merrill, J. E., Y. Koyanagi, and I. S.Y. Chen. 1989. Interleu-kin-1 and tumor necrosis factora can be induced from mono-nuclear phagocytes by human immunodeficiency virus type 1 binding to the CD4 receptor. J. Virol. 63:4404-4408.
19. Mikovits, J., A. Raziuddin, M. Gonda, M. Ruta, N. C.Lohrey, H.-F. Kung, and F. W. Ruscetti. 1990. Negative regulation of human immune deficiency virus replication in monocytes: dis-tinctions between restricted and latent expression in THP-1 cells. J. Exp. Med. 171:1705-1720.
20. Molina, J.-M., D. T. Scadden, C. Amirault, A. Woon, E. Vannier, C. A. Dinarello, and J. E. Groopman. 1990. Human immunodeficiency virus does not induce interleukin-1, interleu-kin-6, or tumor necrosis factor in mononuclear cells. J. Virol. 64:2901-2906.
21. Mori, K., D. J. Ringler, T. Kodama, and R. C. Desrosiers. 1992. Complex determinants ofmacrophage tropism in env of simian immunodeficiency virus. J. Virol. 66:2067-2075.
22. Nabel, G., and D. Baltimore. 1987. An inducible factor activates expression of human immunodeficiency virus in T-cells. Nature (London) 326:711-713.
23. Narayan,O., and J. E. Clements. 1989. Biology and pathogen-esis of lentiviruses. J. Gen. Virol. 70:1617-1639.
24. O'Brien, W. A., Y. Koyagi, A. Namazie, J.-Q. Zhao, A. Diagne, K. Idler, J. A. Zack, and I. S. Y. Chen. 1991. HIV-1 tropism for mononuclear phagocytes can be determined by regions ofgpl20
outside theCD4-binding domain. Nature (London) 348:69-73. 25. Osborn, L., S. Kunkel, and G. J. Nabel. 1989. Tumor necrosis
factor alpha and interleukin-1 stimulate the human immunode-ficiency virus enhancer by activation of the nuclear factor KB. Proc. Natl. Acad. Sci. USA 86:2336-2340.
26. Pomerantz, R. J., M. B.Feinberg,D. Trono, and D. Baltimore. 1990. Lipopolysaccharide is a potent monocyte/macrophage-specific stimulator of human immunodeficiency virus type 1 expression. J. Exp. Med. 172:253-261.
27. Regier, D. A., and R. C. Desrosiers. 1990. The complete nucle-otide sequence of a pathogenic molecular clone of simian immunodeficiency virus. AIDS Res. Hum. Retroviruses 6:1221-1231.
28. Ringler, D. J., M. S. Wyand, D. G. Walsh, J. J. MacKey, P. K. Sehgal, M. D. Daniel, R. C. Desrosiers, and N. W. King. 1989. The productive infection of alveolar macrophages by simian immunodeficiency virus. J. Med. Primatol. 18:217-226. 29. Ross, E. K., A. J. Buckler-White, A. B. Rabson, G. Englund, and
M. A. Martin. 1991. Contribution of NF-KB and Spl binding motifs to the replicative capacity of human immunodeficiency virus type 1: distinctpatterns of viral growth are determined by T-celltypes. J. Virol. 65:4350-4358.
30. Schuitemaker, H., M. Koot, N. A. Kootstra, M. W. Dercksen, R. E. Y. deGoede, R. P. van Steenwik, J. M. A. Lange, J. K. M. E. Schattenkerk, F. Miedema, and M. Tersmette. 1992. Biological phenotypeof human immunodeficiency virus type 1 clones atdifferentstages of infection: progression of disease is associated with a shift from monocytotropic to T-cell-tropic viruspopulations. J. Virol.66:1354-1360.
31. Shioda, T., J. A. Levy, and C. Cheng-Mayer. 1991. Macrophage and T cell-line tropisms of HIV-1 are determined by specific regions of the envelope gpl20gene. Nature (London) 349:167-169.
32. Strebel, K., D.Daughterty,K. Clouse, D. Cohen, T. Folks, and M. A. Martin. 1987. The HIV "A" (sor) gene product is essentialforvirus infectivity. Nature (London) 328:728-730.