0022-538X/83/090487-08$02.00/0
Copyright © 1983, AmericanSocietyforMicrobiology
Nonviable
Mutants
of
Simian
Virus 40 with Deletions Near the
3' End of Gene
A
Define a Function for
Large
T
Antigen
Required
After Onset of
Viral DNA
Replication
JOANNE TORNOWANDCHARLES N. COLE*
DepartmentofHumanGenetics, Yale University Schoolof Medicine,NewHaven, Connecticut 06510 Received 3 February 1983/Accepted 7 June 1983
Deletionmutantsof simian virus40(SV40)withlesionsatthe three DdeI sites
nearthe 3' endof the early region were constructed. Mutantswith deletionsat
0.203 and 0.219mapunits (mu) which didnotchangethelargeTantigen reading
framewereviable. This extendsslightly theupstreamboundary for the location of viablemutantswithdeletions in the 3' end of theAgene. Mutantswith frameshift deletions at 0.193 and 0.219 mu were nonviable. These are the first nonviable
mutantswith deletions in thisportion of theAgene.Noneof the three nonviable
mutants with deletions at 0.219 mu produced progeny viral DNA. These three
mutantsall used the alternate readingframe located in thisportion of the SV40
early region. Themutant witha deletionat0.193mu, dlA2459, waspositivefor viral DNA replication and was defective foradenovirus helper function. All of thesemutationswerelocated in theportionof theSV40largeTantigenwhich has
nohomologytothepolyomaTantigens.These resultsindicate that thisportionof largeTantigen isrequiredforsomelate stepin theviralgrowth cycle andsuggest
thatadenovirushelperfunction isrequiredforproductiveinfectionbySV40.
Papovavirus large T antigens are multifunc-tional during both productive and transforming
infections. Muchofourknowledge about simian
virus 40 (SV40) largeTantigen derives from the
study of mutants with lesions in the A gene,
which encodes largeT(13). These include both
temperature-sensitive(7, 43) and
non-condition-ally defective deletion (10, 25) mutants. These
studies indicate that large T is responsible for
theinduction of viral DNA synthesis(7, 43), the
stimulation of host DNA replication (20), regula-tion of the level of early mRNAs (3, 36, 45),
induction, and, in some cases, maintenance of
late mRNA synthesis (1, 12), induction of in-creased levels of host cellenzymesinvolved in nucleic acidmetabolism(23),activation of silent
rDNAgenes(40),provision of adenovirus helper
function (9, 19, 34),aswellasinitiation, and, in
most cases, maintenance of the transformed
stateinnonpermissive cells (21, 22, 44).
Comparison of the DNA sequences of SV40
(17, 35) and polyoma (18, 39) indicates that
substantial regions ofhomology exist between
the two genomes. Each genome also contains
unique regions. A sequence of approximately 500base pairs (bp)in themiddle ofthe polyoma
t Present address: Department of Biological Chemistry, CaliforniaCollege ofMedicine, UniversityofCalifornia,
Ir-vine, CA 92717.
tPresent address:DepartmentofBiochemistry,Dartmouth MedicalSchool, Hanover,NH 03756.
earlyregion encodes the unique portion of
mid-dle T antigen and, in another reading frame, a
portion of large T. Thisregion has no
counter-part in SV40. SV40 hasasequenceof approxi-mately 400 bp, between mappositions 0.25 and 0.174 mapunit (mu), that hasnocounterpart in polyoma. This region encodes the C-terminus of SV40largeTantigen. Two properties have been
assignedtothis portion of the SV40genome:(i)
adenovirus helper function, requiring no more
than the C-terminal 37 amino acids oflarge T
(16, 33), and (ii) the62-base small SV40-associ-ated RNA, which is complementary to early
mRNA near 0.21 mu(2, 27). This portion ofthe
SV40early region also containsanopenreading frameof 99 codons which beginsat0.219 mu and
endsat0.169mu.Thetermination codon for this alternate readingframe occurs beyondthe
nor-mal termination codon for large T antigen, but
upstreamfrom the 3' end of the early mRNAs.
Several SV40 mutants contain deletions near
the 3' endof theAgene(see Fig. 1; 10, 15, 31,
32,33). All ofthese mutants areviable andhave
lesions distal tomap position 0.217 that do not
changethe largeTantigen reading frame. One,
d1265 (Cole), has a 39-bp deletion (47) which
results in thereplacementofthe six C-terminal
amino acid residues with four encoded
down-stream from the normal termination codon for
large T (14). Although some of these mutants have slightly slower rates of DNA replication
487
on November 10, 2019 by guest
http://jvi.asm.org/
(33), both the lytic and the transforming
infec-tioncyclesproceed normally. Much oftheSV40
A gene distal to map position 0.217 has been
deleted inone oranother of thesemutants. Since
many of these viable deletion mutants fail to
produce the SV40-associated RNA (2; M. Pol-vino-Bodnar and C. N. Cole, unpublished data), it cannot be essential for productive infection.
Although some of the mutants exhibit reduced
levels of adenovirus helper function (9), none
are absolutely defective for this function. No
essential functionprovided bythisportion ofthe
Agenehas been described.
In thisreport,wedescribe thepreparationand
characterizationofadditionalmutantswith
dele-tions in thisportion ofthe SV40Agene. These
mutants were prepared as recombinant DNA
molecules. The method used allowed the
isola-tion ofmutantswithsmall deletions(3to22bp),
incontrasttothelargerdeletions(20to1,000 bp)
isolated inpreviousstudies(10, 25, 33).
Wefound thatmutantswithdeletionsat0.203
and 0.219 mu, which did not alter the large T
antigen reading frame, were viable. This result
extends slightly the upstream limits for viable
deletionmutants. Mutants with frameshift
dele-tionsat0.193and0.219mudid not formplaques.
The nonviable mutant with a lesion at 0.193,
dlA2459, was positivefor viral DNA synthesis,
whereas no viral DNA was produced in cells
infected with nonviable mutants with deletions
at0.219mu. Inaddition, dlA2459wasdefective
for adenovirus helper function. Therefore, the
C-terminus ofSV40 largeTantigen is essential
forproductiveinfection andprovidessome
func-tion requiredafter the onset ofviralDNA
repli-cation. This late function ofSV40 largeT
anti-gen may be related to adenovirus helper function.
MATERIALS ANDMETHODS
Cells,cellculture, and viruses.ThegrowthofCV-1 andCV-lp cells has been described previously (33). All cellsweremaintainedinDulbecco modifiedEagle mediumcontaining5%fetalbovine serum. Our
wild-type strainofSV40wasthesmallplaquestrain
orig-inally characterized byTakemoto etal. (42). Mutant d1BC865wasoriginallydescribedbyCarbonetal.(6) and hasadeletionattheEcoRIsite. Mutant d1A1209 (10) lacks 329 bp (Polvino-BodnarandCole, unpub-lisheddata)between 0.65 and 0.587 mu. Viable dele-tionmutantsdl1263 andd11265,lacking 33 bpat0.20
muand 39bpat0.18mu,respectively,wereoriginally
describedbyColeetal.(10);viablemutantviral DNAs
werepreparedasdescribedpreviously (10).
Preparationofmutants. EcoRI-cleavedSV40DNA
wasinserted into the EcoRI site ofpBR322.ThisDNA wasdigestedwithDdeIendonuclease in thepresence
of180,ugofethidium bromideperml(30, 38)at37°C for 45min.Under theseempiricallydetermined condi-tions, mostmolecules werecleavedonce. ThisDNA
wasextracted withphenol, precipitated withethanol, suspendedinS1buffer(280mMNaCl,50 mM sodium
acetate [pH4.6], 1 mMZnCl2), and digested withSi
nuclease, using conditions which led to the removal of asmall number of nucleotides from each end (5). After electrophoresis in a 1.0% agarose gel in Tris-borate-EDTA buffer (89 mM Tris-hydroxide, 89 mM boric acid, 2.5 mM EDTA [pH 8.3]), the largest class of linears was isolated by electroelution in E buffer (40 mMTris-hydroxide, 12.5 mM sodium acetate, 2.5 mM EDTA). These were ligated at a concentration of 5 ,ug/ml, using T4 DNA ligase, and were used to trans-fect Escherichia coli HB 101 (26). Although pBR322 contains eightDdeI sites, most of these occur within thegenesfor resistance to ampicillin and tetracycline or within the pBR322 origin of replication. Since transfected bacteria were plated on agar containing both ampicillin (20 ,ug/ml) and tetracycline (12.5
,ug/ml),mostof the deletion mutants isolated mapped
within the SV40 portion of the recombinant. Minily-sates were prepared by the method ofBirnboim and Doly (4) and were screened for the presence of mu-tantswith small deletions by digestion separately with
HindlIl andDdeI.Plasmid DNA was prepared by the
method of Clewell and Helinski (8) and was purified by equilibrium density centrifugation in gradients of cesi-umchloride-containing ethidium bromide (300 ,ug/ml). DNA tobe usedfor sequence analysis was rebanded onceinacesium chloride-ethidium bromide gradient. DNA of SV40 mutants dIBC865 anddIA1209 was digested withBamHIand was inserted into theBamHl site of pBR322. These plasmids were designated pdlBC865 andpd1A1209,respectively.
Plaque assays. The titers of wild-type SV40 and viable deletion mutant virus stocks were determined
by plaque assay on monolayers of CV-lp cells as
previously described (29). All recombinant mutant
DNAs wereprepared for complementation analysis by
digestionwithEcoRIor BamHI (toseparate viral and
bacterial plasmid sequences) and were ligated at low concentration (5 ,ug/ml) with T4 DNA ligase. Gel electrophoresiswasused to insure that both digestion and subsequent ligation were complete. Genetic com-plementation analysis, using viral DNAs, was per-formed as described previously (29). Some mutants were testedfor temperature sensitivity by incubation of transfected CV-lp cells at 32, 37, and 41°C.
Analysis ofDNA replication. DNA replication was
analyzed bytransfection of cultures of CV-1 cells,in
60-mmdishes, as describedpreviously (33), but with thefollowing modifications. Mutant viral DNAs were separated fromplasmid sequencesasdescribed above. Each60-mmdish received 0.2mlofasolution contain-ing Tris-buffered saline(TS;25 mMTris-chloride [pH
7.5]-137 mM NaCl-5 mM KCI-0.6 mM
Na2HPO4-0.05mM
MgClz-0.7
mMCaC12),DNA(1jig/ml),
and DEAE-dextran(Pharmacia Fine Chemicals, Inc., Pis-cataway, N.J.;500jig/ml) for45min.Cellswerethen washedoncewithTS and were incubated in Dulbecco modified Eagle medium containing 100 ,uM chloro-quine phosphate (Sigma Chemical Co., St. Louis, Mo.) and2% fetal bovineserumfor4h(G. Magnus-son, personal communication). This treatment in-creased the percentage of cellsexpressingTantigento 50 to 60%, as measured by immunofluorescence (J.Tornow, unpublished data). After chloroquine
treat-ment,this mediumwasreplaced with medium lacking chloroquine andcontaining2% fetal bovine serum.
Analysisof Tantigens.Tantigenswerelabeled with
[355]methionineand wereanalyzed by
on November 10, 2019 by guest
http://jvi.asm.org/
489 tation with anti-T serum, followed by electrophoresis
aspreviously described (33).
Chemicals andenzymes. Restriction endonucleases were purchased from either P-L Biochemicals, Inc., Milwaukee, Wis., or New England Biolabs, Beverly, Mass. DNA polymerase I and the Klenow fragment of DNApolymerase I were obtained from New England Biolabs; T4 DNA ligase was from Collaborative Re-search, Inc., Waltham, Mass., or New England Bio-labs; calf intestinal alkaline phosphatase andS1 nucle-ase wereobtained from Boehringer Mannheim Corp., New York, N.Y. Proteinase K was from Beckman Instruments, Inc., Fullerton, Calif. or Bethesda Re-search Laboratories, Bethesda, Md. [cx-32P]dNTPs (800Ci/mmol) and[35S]methioninewere obtained from Radiochemical Centre, Amersham, England. Chloro-quine phosphate was purchased from Sigma. Cesium chloride was obtained from Gallard Schlessinger, CarlePlace, N.Y.
RESULTS
Deletion mutants with lesions in the
C-termi-nalportion oflarge Tantigenhave beenisolated
previously in this(10, 33) and other (15, 31, 32)
laboratories. The location of the deletions in
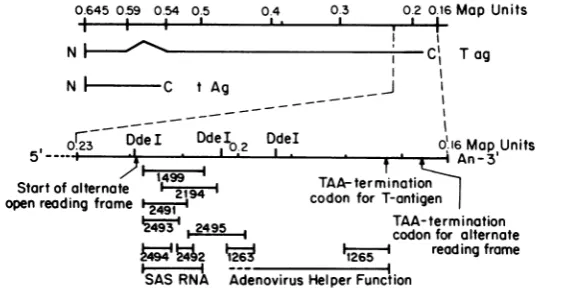
these mutants is shown in Fig. 1. They include
d11263, dl1265, d12491 through d12495, d12194,
and theviable (temperature-independent)
rever-tantoftsA1499, d11499(N. Bouck, A. Pater, C.
Chang, and G. diMayorca, personal
communi-cation). All ofthese mutants contain deletions which aremultiples of3 bp, ranging from 15 to
81bp, leaving the largeTantigen reading frame intact distalto thedeletion. Withthe exception
ofnucleotides 2,721 to2,797, all of the SV40A gene distal to nucleotide 2915 (the 5' boundary of the deletion in d11499(31; numberingsystem as defined by Tooze [46]) has been deleted in
one oranotherof these viable mutants.
As partofaproject to prepare a setofSV40
0.645 0.59 0.54 0.5
2 a
N
N I C t Ag
C.
Dj52
de I DdeIo DdeI 0.lA1499
Start of alternate 2 TAA-termination openreading frame 12491 I codon for T-antigen
_24931
2495 TAA-terrI codonfor
I92
SIl-6
rea [image:3.492.253.449.77.214.2]SASRNA AdenovirusHelperFunction
TABLE 1. Propertiesofmutantswithdeletionsin
the 3' end of theA gene
MuatDdel
siteDeletionNuloie
Mutant deleted size
Ndcletede
Viability
no.
(mu) (bp) d2413 0.219 22 2,925-2,946
-2414 0.219 10 2,927-2,936
-2415 0.219 10 2,928-2,937
-2423 0.219 6-9 +
2431 0.219 3-6 +
2440 0.219 3-6 +
2429 0.203 6-12 +
2451 0.203 6-9 +
2459 0.193 14 2,785-2,798
-a Sizes of deletions of nonviable mutants were de-termined by DNA sequence analysis. The deletion sizes of viablemutants wereestimated from restriction endonucleaseanalysis (datanotshown).
b Nucleotide sequences of themutants were deter-mined by the method of Maxam and Gilbert (28).
Nucleotidenumbering followed thenumberingsystem of Tooze (46).
cViability is definedasthe abilitytoformplaques onmonolayers ofCV-lpcells.
mutants with small deletions throughout the
SV40 early region, we prepared mutants with
small deletions atDdeI cleavage sites, mostof
whicharelocated within theearlyregion.Inthis
report, we describe the properties of nine
mu-tants with deletions at the three DdeI sites
(0.193, 0.203, and 0.219 mu) in the C-terminal
portionof the SV40genome. The nine mutants
are listed in Table 1. The characterization of
mutantswithdeletionsatother DdeIsites will be
described elsewhere (J.TornowandC.N.Cole,
Proc. Natl. Acad. Sci. U.S.A.,in press).
CV-lp cells were transfected with mutant
DNAs,using plaqueassayconditions. Both
mu-0.4 0.3 0.2 0.16 Map Units
I.. I
Tag
16 Map Units An-3'
nination alternate idingframe
FIG. 1. Map oftheearly region of theSV40 genome. Map coordinates are expressed as the fractional length ofwild-type SV40, where 0.00 is the map position of the singleEcoRIsite. The 3' end of early mRNA is shown enlarged in the lower part of the figure. Beneath this are shown the positions of previously described viable deletion mutants (10, 15, 31-33), as well as the regions in which the SV40-associated small RNA (2) and adenovirus helper function (9, 16) map.
VOL.47, 1983
I--r-cl
I
-i
I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.492.117.398.493.638.2]TABLE 2. Genetic complementation analysis of nonviablemutantsa
DdeIsite Complementing mutant
Mutant no. deleted (PFU4Lg)
(mu) dIA1209 dIBC865
2413 0.219 0 5 x 104
2414 0.219 0 4 x 103b
2415 0.219 0 8 x 104
2459 0.193 0 5 x 104
a Monolayers of CV-lp cells were exposed to dilu-tions of mutant DNA and 0.01 ,ug of complementing mutantDNA.
bThis value is lower than values obtained with other mutants because the dl2414 DNA was not ligated before infection of the CV-lp cells.
tants with deletions at 0.203 mu (d12429 and
d12451) and three of the six with the deletionsat
0.219mu wereviable(d12423, d12431, dl2440).In
general, plaquesappearedatthe sametime and
enlarged at the same rate as wild-type SV40 plaques. These mutantsextend slightly the
up-stream boundary for viable deletion mutants in
the A gene of SV40. The sole isolate with a
deletionat0.193 mu(d12459)and theother three
mutants with deletions at 0.219 mu (d12413,
d12414, and d12415)were notviable. Plaques did
not appear when dl2459-transfected cells were
incubatedat32, 37,or41°C, indicating that this
mutant was nonviable rather than temperature
sensitive. These represent the first nonviable
mutantsisolated thathavelesions in thispartof
theSV40genome.
The DNA sequences of all four nonviable
mutants wereanalyzed.The resultsareshown in
Table 1. Mutant d12459 lacked 14 bp, which
shouldcauseashift in thereadingframe oflarge
Tantigen. The deletions in d12413, d12414, and d12415were22, 10, and10bp, respectively, and were overlapping. Theselesions shouldcause a
shift into the alternate readingframe of99
co-dons located between 0.219and 0.168 mu. The DdeIsiteat0.219mu(CTAAG)containsthelast
termination codon (TAA) upstream from this
alternate reading frame. The approximate sizes
of the deletions in the viable mutants,
deter-minedfrom restriction endonucleasedigestions,
arelisted in Table1.
Genetic complementation analysis. All four
nonviable mutants weretested bygenetic
com-plementation analyses, using dlBC865 and
dlA1209. MutantdlA12O9 contains a deletion of
329 bpbetween map positions0.587 and 0.656,
including the cap site for early mRNAs, the
initiation codon for both large and small T
antigens, and the donor splice site for large T
mRNA(10,33; C.ColeandM.Polvino-Bodnar,
unpublished data). MutantdlBC865 has a small
deletion at the EcoRI site (6) and produces a
small amino-terminalfragment of VP1 (C. Cole and S. P. Goff, unpublished data). All four
mu-tants formed plaqueswithdlBC865butnotwith
dlA1209 (Table 2). Therefore, all four belonged
tothe Acomplementationgroup.
Large T antigens produced by the deletion
mutants. Cultures ofCV-1 cells, transfected by
each mutant or wild-type SV40 DNA, were
labeled with [35S]methionine for1 h, 30h after infection. Large T antigens were examined by
immunoprecipitation, using hamster antitumor
(anti-T) serum and Formalin-fixed protein
A-bearing Staphylococcus aureus, followed by electrophoresis in sodium dodecyl sulfate
(SDS)-polyacrylamide gels. Mutant and
wild-type largeT antigens are shown in Fig. 2. The
threeviable mutantswithdeletions at0.219 mu
produced large T antigens (Fig. 2, lanes 7to 9) indistinguishable in electrophoretic mobility from wild-type T antigen (lane 1). The large T
antigen produced in cells transfected by dl2429
(lane 10) migrated slightly more rapidly than wild-type large T (lane 11). The large T antigen
produced in cellstransfected bydl2459 (lane 3)
migrated morerapidlythanwild-type, in agree-mentwithareading frame shift and termination
of translation at 0.189 mu, just downstream from
the deletion at 0.193 mu. The large T antigens
_,C ._
a 1
FIG. 2. Autoradiogram of [35S]methionine-labeled Tantigens immunoprecipitated from cytoplasmic ex-tracts. CV-1 cells (in35-mm dishes) were transfected withwild-type or mutant DNAs. Cultures were labeled for1hbeginning 30hafter infection, using75 ,uCiof
[35S]methioninein 0.25 mlof medium lacking
methio-nine. Extracts were immunoprecipitated, alkylated, and subjected to electrophoresis on 7 to 20%
SDS-polyacrylamide lineargradient gels. Gels werefixed,
fluorographed, dried, and exposed to Kodak XAR-5 film withanintensifying screen for 3 days at -70°C. The positions of the large and smallT antigens are shown. Lanes: (1 and 11)wild-type SV40; (2) mock-infected; (3) 2459; (4) 2413; (5) 2414; (6) 2415; (7) 2423; (8) 2431; (9) 2440; (10) 2429. Lanes 10 and11arefrom the sameautoradiogram and werealigned with each otherby matching the positions forsmallt.
r-'2 L' ;: >:;':- 9G
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.492.278.423.384.517.2]produced by mutants dl2413 (lane 4), dl2414 (lane 5), and d12415 (lane 6) should be slightly
larger than wild-type large T due to the use of
the alternate reading frame distal to 0.219 mu.
The T antigens produced by these mutants
mi-grated slightly morerapidlythanwild-type large
Tantigen.
DNAreplication inmutant-infectedcells. CV-1
cells were transfected with either wild-type or
nonviable mutant DNAs (dlA2413 or dlA2459)
and were analyzedfor the presence ofprogeny
DNA molecules (Fig. 3). Proteinase K-treated
Hirt supernatants were extracted with phenol, precipitated with ethanol, and digested with MboI. Plasmid, butnotprogeny, DNAcontains methylated adenosine residues in the MboI
rec-ognition and cleavage sequence, 5'-GATC-3',
and is resistant to digestion by MboI. After
digestion, DNAwas subjected to
electrophore-sis inagarosegels,transferredtonitrocellulose,
and hybridized with radiolabeled SV40 DNA.
Progeny viral DNA was synthesized in cells infected by d12459 (Fig. 3C), but not in cells
infected bydIA2413, a nonviable mutantwith a
deletionat0.219mu(Fig.3B). About one-fourth
as much progeny DNA was formed in
dl2459-infected cells asinwild-type SV40-infectedcells (Fig. 3A). No progeny DNA was detected in cells transfected with dlA2415, another
nonvia-ble mutantwithadeletionat0.219mu(datanot
shown). We conclude that mutants with dele-tionsat0.219 mu wereblocked atthe initiation of viral DNA replication, the first step in the
productive infection cycle which is known to
require large T(43). Mutant dlA2459, however, was blockedat alater stage.
Effect of mutantcoinfectiononadenovirus late
protein synthesis. The ability of dlA2459to
en-hance thelevelsof late adenoviruspolypeptides
synthesized in CV-1 cells was examined by
transfecting cells with wild-type SV40 or
dele-tionmutantsand superinfecting them 48 h later
withadenovirus type 2. [35S]methionine-labeled
whole cell extracts were prepared 72 h after
superinfection and analyzed by
SDS-polyacryl-amide gel electrophoresis (Fig. 4). Cells
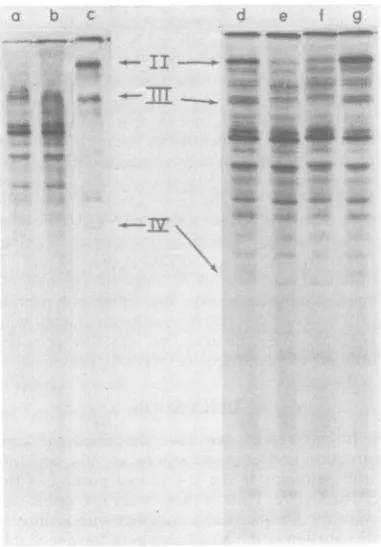
coin-fected bywild-type SV40 and adenovirus (lane
d) had substantially higher levels of adenovirus polypeptidesII, III, and IV thandidCV-1 cells
infectedonly by adenovirus (lane b). The level
of polypeptide IIincellscoinfectedby dlA2459
was slightly lower than that in cells coinfected
by d11265 (lanef) and much lowerthanin cells coinfected with either wild-type SV40 (lane d)or
d11263 (lane g). The level of adenovirus helper function ofd11263 and d11265 isonly 30 and6% of that ofwild-type SV40, respectively,as
mea-sured by plaque assays (9). Fiber polypeptide
(IV)was notdetectable in cells coinfected with
dlA2459. We conclude that dIA2459 provides
little or noadenovirus helper function.
DISCUSSION
In this report, we have described the
con-struction and characterization of nine mutants
with deletions in the C-terminal portion of the
SV40Agene.Fourofthese were not viable and
were the first nonviable mutants with lesions in
this portion oftheSV40genome. The five viable
deletion mutants isolated, with lesions at 0.203
and0.219 mu, were similar to deletion mutants
which we have described previously (10, 33).
Theisolation of viablemutants with deletions at
0.219 muextends slightly the upstream bound-ary for viable deletion mutants. We have
con-structedsix mutants with deletions at 0.243 mu
(Tornow and Cole, unpublished data), and one
of these had an in-phase deletion. All were
nonviable, suggesting that the boundary for the
location of viable deletion mutants lies
some-where between0.219 and 0.243 mu. To date, all
viablemutants with lesions in this portion of the Agene havein-phase deletions.
UrO
o 2 4 5; <
abw--r
_*
-cr,
5-C>r
ur; 0 1 2 3 4 5
:..,
cr c
^ ^ 2 3 4 5 -2
B
FIG. 3. Autoradiograms ofSouthernblots(41),showingDNAreplication inculturesof CV-1 cellstransfected withwild-type(A),dIA2413(B), ordIA2459(C)DNA.Cultureswereharvested after 0, 1,2, 3, 4, and5days. The marker lanes contained20 ngofwild-type SV40DNAdigested with MboI.
47,
'i.
OA
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.492.116.383.520.641.2]Ii-) C
S R
FIG. 4. Effect of dIA2459preinfection on adenovi-rus lateprotein synthesis. Cultures of CV-C monkey
orA549 humancells, in 35-mm dishes,were transfect-edwithwild-typeormutantDNAs(100ngperplatein
avolumeof 0.2 mlof TS containing 500,ug of
DEAE-dextran per ml) or mock-infected and subsequently exposed to 100 ,uM chloroquine diphosphate as de-scribed inthetext.Whenrecombinant DNAplasmids
were used, the viral DNA was separated from the
plasmidsequences asdescribedinthetext.After48h, thecultureswereinfected with adenovirustype2(2x
107 PFU per plate, approximately 10 to20 PFU per
cell). After an additional 48 h, the cultures were
washedoncewith TS, incubated for 30minwith 1 ml ofDMEMlacking methionineperplate, and thenwere
incubated for 2 h with (per plate) 0.3 ml ofDMEM lacking methionine and containing 80 ,uCi/ml of [35S]methionine and 2%dialyzed fetal bovine serum.
Thecultureswerewashed twicewithTS,andextracts
were prepared by the addition of 0.2 ml of sample buffer (1% SDS-1.4 M 2-mercaptoethanol-0.25 M EDTA-15%glycerol-0.1% bromphenol blue).The
ex-tractswereboiledfor 10min, andequal quantities of acid-precipitable radioactivitywereloadedonto17.5% SDS-polyacrylamide gels,asdescribedby Klessigand Anderson (24). The gels were fixed, dried, and
ex-posedtoKodakXAR-5 film for18h. Lanes:(a)CV-C cells, infectedwithadenovirus; (b) CV-C cells, mock-infected; (c) A549 cells, infectedwith adenovirus; (d) CV-Ccells,preinfected with wild-type SV40; (e) CV-C cells, preinfectedwith dlA2459;(f)CV-C cells, prein-fected with d11265; (g) CV-C cells, preinfected with d11263. The positions of adenovirus polypeptides II, III,and IV areshown.
The large T antigens encoded by dIA2413,
dIA2414, and dlA2415 (Fig. 5) should exist as a fusion product between the N-terminal 629
ami-noacids of large T and the residues encoded by
the alternate readingframe at the 3' end of the
early region (Fig. 5), resulting in a polypeptide
slightly larger than wild-type large T antigen. The polypeptides detected (Fig. 2) migrated
more rapidly than large T. There are two
possi-ble explanations for this observation. (i) This new C-terminal domain could cause a major
change in the physical properties oflarge T, so
that the mutant large T migrates more rapidly
than wild-type large T. (ii) This domain of the
mutantlarge Tantigens may be unstable,
result-inginproteolysis. SV40 largeTmigrates with an
apparent molecular weight of90 kilodaltons in
ourSDS-polyacrylamide gels,whereasitsactual size, determined from the DNA sequence, is
81.6kilodaltons (17, 35). It has been shown that
deletions in this portion of large T cause it to
migrate more rapidly than would be expected
from thesizeofthedeletion,suggesting that this
portion of large T contributes to its aberrantly
slow electrophoretic migration (10, 33).
There-fore, the mutant large T antigens would be expectedtomigrateneartheobserved positions
in the absence ofproteolysis.
Mutants dlA2413 and dlA2415, with deletions
at 0.219 mu, were defective for replication of
viral DNA. Mutant dIA2459, with a lesion at
0.193 mu, was not viable but was positive for
viral DNA replication. Therefore, dIA2459was
blocked at a later step ininfectionthan were all
previously described nonviable mutants with deletions in the A gene (10, 25; Cole,
unpub-lisheddata) and most tsA mutants (7, 21, 22, 43).
Amino acids encoded distally to 0.193 mu are
notrequired forthe viral replication function of
large T antigen. We cannot determine whether
amino acids encoded distally to 0.219 mu are
required for replication, because the large T
antigens of dlA2413, dIA2414, anddIA2415 con-tain at their C-termini approximately 95 amino acids encoded by the alternate reading frame.
The loss of the replication function of large T
mayresult from this substitution. The
substitut-ed residues differ substantially in both charge
andhydrophobicity from the wild type(Fig. 5).
The determination of the importance for viral
DNA replication of residues encoded between
0.219and 0.193muwillrequiretheisolation and
analysisof SV40mutantswhichproduce large T
antigens truncated at different sites between
0.219 and 0.193 mu.
In dlA2459-infected cells, DNA replication
occurred, although only one-fourth as much
progeny DNA was synthesized (Fig. 3). No
plaques were produced when CV-lp cellswere
transfected with dlA2459 DNA. In these proper-.,I
=
!R
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.492.51.242.82.356.2]A. 630
SV4.LeuArg AsnSerAspAsp Asp Asp Glu Asp
iR
Gln Glu Asn Ala Asp dlA2459... l650
Lys Asn Glu Asp Gly Gly Glu Lys Asn Met Glu Asp-,-,r"Gly His GluThr Gly
B.
670
leAspSe.r.Gin Se.rGin Gly
,S;e
Phe Gln Ala Pro Gln Se,SerGinete Vol Val CysSer-COOH690
His Asp His Asn GinPro Tyr His Ile Cys Arg Gly Phe Thr Cys PheLys Lys
706
Pro ProrE Pro Pro Pro Glu Pro Glu Thr -COOH
Glu Thr Val Met Met Met Met Lys Thr Ala Arg Lys Met Leu Ile Lys Met LysMetVal Gly Arg Arg Thr Trp Lys Thr Gln Ala Met Lys Gin Ala Leu Ile His Ser Pro Lys Ala His PheArg Pro Leu Ser Pro His Ser Leu Phe Met Ile Ile Ile Ser His Thr Thr Thr PheVal Glu Val Leu Leu Ala LeuLysAsn Leu Pro His Leu ProLeu AsnLeuLysHis Lys MetAsn AlaIle Val Val Val Asn Leu PheIle Ala Ala Tyr Asn Gly Tyr Lys-COOH
FIG. 5. (A) Amino acid sequence of the carboxy terminus of large T antigen. Amino acid residues are numberedaccordingtoTooze(46).Phosphorylatedresiduesareboxed;residues which may bephosphorylated
areenclosedby a dotted box (37). Ser676,Ser677, andSer679arephosphorylated inasmallfraction oflargeT molecules present in the infected cell(G. Walter, personalcommunication).Thecarboxyterminus of thelargeT
antigen produced bydlA2459isalsoshown. TheopenboxrepresentsthatportionofdIA2459largeTidentical to
wild-type antigen. (B) Amino acid sequence of thealternateopen readingframe.
ties, this mutantresembles tsA1642 (11),which
has a single base alteration at 0.31 mu. The
lesions in tsA1642 and dlA2459 affect different functions of large T, since tsA1642 is able to
complementdlA2459 forplaque formation
(Tor-nowand Cole, in press).
Adenovirus helper function is the only known property oflarge T associated with the portion of the polypeptide that is absent from dIA2459 largeT.Since dlA2459largeTlacks the residues associated with adenovirus helper function, it
was not surprisingtodiscoverthatdlA2459had
littleornoabilitytoenhancethesynthesisofthe
lateadenoviruspolypeptides (Fig.4).Therefore,
ourresults suggest thatadenovirus helper func-tion isrequired foraproductive SV40 infection. The large T antigen produced by dlA2459 also lacksat least one site of phosphorylation (Fig. 5).
MutantdlA2459 defines an additional T
anti-gen function that is required for a productive infection. Although SV40 and polyoma largerT
antigens show substantialhomology,each
poly-peptide has regions which have no homology
with the other large T antigen. The entire
C-terminal one-sixth ofSV40large T antigen is not
homologous toanyportion ofpolyoma large T.
The studies described in this report show that
thefunction defectivein dlA2459requires some
of thisportion oflargeT.
ACKNOWLEDGMENTS
We thankGeorge Santangelo and Maryellen
Polvino-Bod-narfor usefuldiscussions,suggestions,andcritical reading of
the manuscript. We also thank Kathy Miceli for excellent technical assistance.
This work was supportedby grant PCM80-21805 from the National Science Foundation. J.T. wassupported by a Public Health Service predoctoral training grant from the National Institutes of Health.
LITERATURECITED
1. Alwine, J. C., and G. Khoury. 1980. Effect of a tsA mutation on simian virus 40 late gene expression: varia-tions between host celllines. J. Virol. 33:920-925. 2. Alwine, J. C., and G. Khoury. 1980. Simian virus
40-associated small RNA: mapping on the simian virus 40 genome and characterization of its synthesis. J. Virol. 36:701-708.
3. Alwine, J. C., S. I. Reed, and G. R. Stark. 1977. Charac-terization of theautoregulation of simian virus 40 gene A. J.Virol. 24:22-27.
4. Birnboim, H. D., and J. Doly. 1979. A rapid alkaline extraction procedure forscreeningrecombinant plasmid DNA.Nucleic Acids Res. 7:1513-1523.
5.Bogenhagen, D. F., S.Sakonju,and D. D. Brown. 1980. A control region in the center of the5S RNA gene directs specificinitiationoftranscription. II. The 3' border of the region. Cell 19:27-35.
6. Carbon, J.,T. E.Shenk,and P.Berg. 1975. Biochemical procedureforproduction of small deletions in simian virus
40 DNA. Proc. Natl.Acad.Sci.U.S.A. 72:1392-13%.
7. Chou, J. Y.,and R.G.Martin. 1974.Complementation analysis of simian virus 40 mutants. J. Virol. 13:1101-1109.
8. Cleweil, D. B., and D. R. Helinski. 1970. A membrane filtertechnique for the detection of complementary DNA. Biochemistry 9:4428-4440.
9. Cole,C.N., L.V.Crawford, andP.Berg. 1979. Simian virus 40mutants with deletions at the 3' end of the early region are defective in adenovirus helper function. J.
Virol.30:683-691.
10. Cole,C. N., T.Landers,S. P.Goff,S.Manteuil-Brutlag,
and P. Berg.1977.Physicalandgenetic characterization VOL. 47,1983
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.492.93.402.67.272.2]of deletion mutants of simian virus 40 constructed in vitro. J.Virol. 24:277-294.
11. Cosman, D. J., and M. J. Tevethia. 1981. Characterization ofatemperature-sensitive, DNA-positive, nontransform-ing mutans of Simian Virus 40. Virology 112:605-624. 12. Cowan, K., P. Tegtmeyer, and D. D. Anthony. 1973.
Relationship of replication and transcription of simian virus 40 DNA. Proc. Natl. Acad. Sci. U.S.A. 70:1927-1930.
13. Crawford, L. V., C. N. Cole, A. E.Smith, E. Paucha, P. Tegtmeyer, K.Rundeli,and P.Berg. 1978. The organiza-tion and expression of the early genes of simian virus 40. Proc. Natl. Acad. Sci. U.S.A. 75:117-121.
14. Denhardt, D. T., and L. V.Crawford. 1980. Simian virus 40T-antigen: identification of tryptic peptides in the C-terminal region and definition of the reading frame. J. Virol. 34:315-329.
15. Feunteun, J., G. Carmichael, J. C. Nicolas, and M.Kress. 1981.Mutant carrying deletions in the two simian virus 40 early genes. J. Virol. 40:625-634.
16. Fey, G., J. B. Lewis, T. Grodzicker, and A. Bothwell. 1979.Characterization of a fused protein specified by the adenovirus type 2-simian virus 40 hybrid Ad2+ND1 dp2. J.Virol. 30:201-217.
17. Fiers, W., R. Contreras, G.Haegeman, R.Rogiers, A. van
deVoorde, H. van Heuverswyn, J. van Herreweghe, G. Volckaert, and M. Ysebaert. 1978. Complete nucleotide sequenceof SV40 DNA. Nature (London) 273:113-120. 18. Friedmann, T., A. Esty, P. LaPorte, and P. Deininger.
1979. Thenucleotide sequence and genome organization of the polyoma early region: extensive nucleotide and amino acid homology with SV-40. Cell 17:715-724. 19. Grodzicker, T., J. B. Lewis, and C. W. Anderson. 1976.
Conditional lethal mutants of adenovirus type 2-simian virus 40hybrids. II. Ad2+ND1 host-range mutants that synthesize fragments of the Ad2+ND1 30K protein. J. Virol.19:559-571.
20. Hatanaka, M., and R. Dulbecco. 1966.Induction of DNA synthesis by SV40. Proc. Natl. Acad. Sci. U.S.A. 56:736-743.
21. Kimura, G., and R. Dulbecco. 1973. A temperature-sensitive mutant of simian virus 40affecting transforming ability. Virology 52:529-534.
22. Kimura, G., and A. Itagaki. 1975.Initiation and mainte-nanceofcell transformation by simian virus 40: a viral geneticproperty. Proc. Natl. Acad. Sci.U.S.A. 72:673-677.
23. Kit, S.1968.Viral-induced enzymes and viral carcinogen-esis. Adv.Cancer Res. 11:73-221.
24. Klessig, D. F., and C. W. Anderson. 1975. Block to
multiplicationof adenovirus serotype 2 in monkey cells. J. Virol.16:1650-1668.
25. Lai, C.-J. and D. Nathans. 1974. Deletion mutants of simian virus 40generated byenzymaticexcision of DNA segmentsfrom the viral genome. J. Mol. Biol. 89:179-193. 26.Mandel, M., and A. Higa.1970.Calcium-dependent
bacte-riophageDNAinteraction. J. Mol.Biol.53:159-162.
27. Mark,D.R., andP.Berg.1979. Athirdsplice site in SV40 early mRNA. Cold Spring Harbor Symp. Quant. Biol.
44:55-62.
28. Maxam, A., and W. S. Gilbert. 1980. Sequencing end-labelled DNA with base specific chemical cleavages. MethodsEnzymol.65:499-560.
29. Mertz, J. E., and P. Berg. 1974.Defective SV40 genomes: isolation and growth of individual clones. Virology.
62:112-124.
30. Parker, R. C., R. M. Watson, and J. Vinograd. 1977. Mapping of closed circular DNAs be cleavage with re-striction endonucleases and callibration by agarose gel electrophoresis. Proc. Natl. Acad. Sci. U.S.A. 74:851-855.
31. Pintel, D., N. Bouck, and G. diMayorca. 1981.Separation oflyticandtransformingfunctions of the simian virus 40A
region: two mutants which are temperaturesensitive for. lytic functions have opposite effects on transformation. J. Virol.38:518-528.
32. Pintel, D., N. Bouck, G. diMayorca, B.Thimnappaya,B.
Swerdlow, andT.Shenk.1979. SV40mutanttsA1499 is head-sensitive forlyticgrowth but generates cold-sensi-tive rat cell transformants. Cold Spring Harbor Symp. Quant. Biol. 44:305-309.
33. Polvino-Bodnar, M., and C. N. Cole. 1982.Construction andcharacterization of viable deletion mutants of simian virus 40 lacking sequences near the 3' end of the early region. J. Virol. 43:489-502.
34. Rabson, A. S., G. T.O'Conor, I.K.Berezesky,and F.J. Paul. 1964.Enhancement ofadenovirusgrowthin African greenmonkeykidney cell culturesby SV40. Proc. Soc. Exp. Biol. Med. 116:187-190.
35. Reddy, V. B., B.Thimmappaya, R. Dhar, K.N.
Subra-manian, B. S.Zain,J. Pan,P. K. Ghosh,M. L. Celma,
andS.M.Weissman. 1978. The genomeof simian virus 40.Science 200:494-502.
36. Reed,S. I., G. R. Stark, and J. C.Alwine.1976. Autoregu-lation of simian virus 40 geneAby Tantigen.Proc. Natl. Acad. Sci. U.S.A.73:3083-3087.
37. Scheidtmann, K.-H., B. Echle, and G. Walter. 1982.
Simian virus 40 large T antigen is phosphorylated at
multiple sitesclustered in two separateregions.J.Virol. 44:116-133.
38. Shortle, D., andD.Nathans.1978. Localmutagenesis:a
method forgenerating viral mutantsinpreselectedregions of the genome. Proc. Natl. Acad. Sci. U.S.A. 75:2170-2174.
39. Soeda, E., J. R. Arrand, N. Smolar, J.E.Walsh,andB. E. Griffin. 1980.Codingpotentialandregulatory signalsof thepolyomavirus genome. Nature (London) 283:445-453. 40. Soprano,K.J.,V.G. Dev, C.M.Croce,andR.Baserga. 1979.Reactivationof silent rRNA genesbysimian virus 40in human-mousehybridcells. Proc. Natl. Acad. Sci. U.S.A. 76:3885-3889.
41. Southern, E. M. 1975. Detectionofspecific sequences among DNAfragments separatedby gelelectrophoresis. J. Mol.Biol. 98:503-517.
42. Takemoto, K. K., R. L. Kirschstein,andK.Habel. 1966. Mutantsof simian virus 40differinginplaquesize,
onco-genicity,and heatsensitivity.J.Bacteriol. 92:990-994. 43. Tegtmeyer, P. 1972.Simian virus 40deoxyribonucleicacid
synthesis: the viralreplicon.J.Virol. 10:591-598. 44. Tegtmeyer, P.1975. Function ofsimian virus40geneAin
transforminginfection.J.Virol. 15:613-618.
45. Tegtmeyer, P.,M.Schwartz, J.K.Collins,andK. Run-deli. 1975. Regulation of tumor antigen synthesis by simian virus40geneA.J. Virol.16:168-178.
46. Tooze, J. 1981. DNAtumorviruses. ColdSpringHarbor Laboratory, ColdSpring Harbor, N.Y.
47. VanHeuverswyn, H., C.Cole,P.Berg,andW.Fiers. 1979. Nucleotide sequence analysis of two simian virus 40
mutantswithdeletions in theregioncodingfor the carbox-yl terminus of theTantigen.J.Virol.30:936-941.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 2. antigens Autoradiogram of [35S]methionine-labeled immunoprecipitated from cytoplasmic ex-](https://thumb-us.123doks.com/thumbv2/123dok_us/1443687.96778/4.492.278.423.384.517/fig-antigens-autoradiogram-s-methionine-labeled-immunoprecipitated-cytoplasmic.webp)