Copyright ©1985, AmericanSociety for Microbiology
Partial
Nucleotide Sequence of Rous Sarcoma
Virus-29
Provides
Evidence that the Original Rous Sarcoma Virus Was
Replication Defective
ANINDYADUTTA, LU-HAI WANG, TERUKO HANAFUSA, AND HIDESABURO HANAFUSA*
The Rockefeller University, New York, New York 10021
Received28February 1985/Accepted 13 May 1985
Rous sarcoma virus-29 (RSV-29) is the strain ofRSV that has the least number of passages beyond its isolationfrom chickentumorno. 1amongallcurrentstrains ofRSV.Biologicalcharacterization indicated that it was replication defective. RNA analysis of nonproducer clones of RSV-29-infected chicken embryonic fibroblastsshowed thepresenceofasubgenomicmessageof 2.6 kilobases containingsrcandagenomicRNA of 7.7 kilobases that containsgag,pol, and src, butnotenv. Thesrc-containing EcoRIfragmentofRSV-29
proviralDNAwasmolecularly cloned. Sequenceanalysisoftheregionsflankingsrcrevealed that theenvgene
wascompletely deletedinRSV-29andthatthesequence acrossthe deletionwasexactly thesame astheBryan high-titer strain of RSV.Thesequenceimmediately 3'tosrcinRSV-29wascloselyrelatedtothatofthePrague strainof RSV.Thefactthat the strain ofRSV whichhas theminimalnumber ofpassagesbeyond itsisolation isreplication defectivesupports thehypothesis ofLernerandHanafusa (J. Virol. 49:549-556, 1984) that the original RSV is a defective transforming virus. This defective transforming virus is postulated to be the precursortoother defectiveRSVs like theBryan high-titer strainand tonondefective RSVs like the Prague strain. Theparticularclone ofRSV-29 thatwestudiedalso hadashort stretchofsequenceduplicationatthe 3' end ofthepolgene,whichwaspresumably created byan errorofreversetranscription.
In most acutely transforming retroviruses, acquisition of anoncogenefrom host genomicsequences byahelper virus hasresulted in the deletion ofone ormore genesessential for replication. Asaresult,these virusesareusually replication defective (11, 17, 24). Thecommonstrainsof Roussarcoma virus (RSV), Schmidt-Ruppin (SR-RSV) (22) and Prague (PR-RSV) (12), are anomalous in that they contain an
oncogene andyetpreserveall thegenesforreplication (34). However, the Bryan high-titer strain of RSV (BH-RSV)has been found to lack the envgene, rendering the virus repli-cation defective (4, 8, 21). BH-RSV could have been a deletion variant derived from nondefectiveRSV. However, the possibility that BH-RSV arose by an independent transduction ofc-src by a nondefective helper virus could
not be ruled out. Sequence analysis ofmolecularly cloned BH-RSV proviral DNA hasdefined theexact nature ofthis deletion. The DNA sequence has also revealed extensive
homology between thesequencefromthe3' end ofsrctothe long terminal repeat in BH-RSV and the corresponding
sequence at the 3' end ofa helper virus, Rous-associated virus 2 (RAV-2). This observation suggested the steps by whichrecombination betweenthehelpervirus and the3' end ofc-src message produced the3' end of BH-RSV (1, 13).
RSV-29 is avariantofRSVobtained fromastock of Rous chickentumorno. 1thatwasdried andstored since 1929. In 1963, one ampoule of this dried tumor was suspended in bufferandinjectedinto chickens. Extracts of thetumorsthat developed contained viable tumorigenic virus, which was called RSV-29 (26).
We found that RSV-29 is also replication defective. We therefore decided to examine RSV-29 for the nature of its defectivenessandfor thestructureofits3' endtodefine its
* Correspondingauthor.
relationship to other strains of RSV and possibly to refine
our views regarding the putative recombinations that took
place between the helper virus and the genomic c-src se-quences togive rise tothe RSVs.
MATERIALSANDMETHODS
Cellsand viruses.Chickenembryofibroblasts(CEF)were from gs-chf-embryos (SPAFAS, Inc., Norwich, Conn.) and were grown before and after transformation in Scherer medium as described previously (7), but the 1:1 mixture of HamF-10 andDulbecco modified Eaglemediumorminimal essential medium were also used in some experiments.
RSV-29, provided by R. M. Dougherty, is only two tissue culturepassagesbeyondachickentumorinduced withadried preparation storedby W. J. Purdyin 1929(26).
Cloning of RSV-29-infected chicken cells. A secondary culture of CEFwasinfected withRSV-29 and itshelpervirus athighvirusdilution(multiplicityof infection of10-4to10-5
focus-forming units per cell). After 5 h, the infected cells weretrypsinized andplatedoutat adensityof106 cellsper
100-mm dish and overlaid with minimal essential medium containing0.36%agar.Individualcolonieswerepickedafter 14 days of incubation and were cultured separately on a feeder layer of uninfected CEF. Clones of cells that were morphologically transformed but did not produce focus-forming virus were superinfectedwith helper virusRAV-2. Clones that consistently produced infectiousfocus-forming virus on superinfection with RAV-2 but that did not do so without helper virus were designated nonproducer (NP) clones and were used for studies on viral RNA. Proviral DNAwas preparedfrom NP clone 1.2.
Preparation of DNAprobes. Viral DNA probes (see Fig. 1B) were prepared from the following viral DNA plasmid clones: pSRA-2, which contains the entire SR-RSV-A (SF)
genome (3); pFC3, which contains the 2.5-kilobase-pair
728
on November 10, 2019 by guest
http://jvi.asm.org/
(kbp)EcoRI DNAfragment of SR-RSV-A (NY)containing
the 5' region ofgag (obtained from F. Cross); pLTR-L6,
which contains the PvuII-to-EcoRI DNA fragment of the SR-RSV-B U3 region (obtained from T. Takeya); and
pTT107, which contains the 3.1-kbp EcoRI fragment of SR-RSV-A (NY) containing the env-src region (31). DNA
fragments representing defined regions ofthe viral genome were prepared bydigestion oftheaboveplasmidDNAs with appropriate restriction enzymes followed by agarose
gel
electrophoresis and electroelution. The purified viral DNA
fragmentswere labeled with [32P]dCTP by nick translation (14).
Viral RNA analysis. Totalpoly(A)-containingRNAswere
prepared from two tofour 10-cm dishes of RSV-29-infected CEF accordingto theprocedures described
previously (33).
Portions (5 to 10 ,ug) of each poly(A)+ RNA
preparation
were used for analysis by agarose gel electrophoresis, Northernblotting,andhybridization with variousviral DNA
probes (Fig. 1). RNase T1 resistant
oligonucleotide
finger-printing hasbeen describedpreviously (32).
Molecularcloning of RSV-29proviralDNAfragment. Total cellular DNA was extracted from RSV-29 clone 1.2 as
described previously (25) and was
digested
tocompletion
with was e tonit
fragn
A
7-7E
2-6ki
PROS E
FIC nonpr RSV-virus, izatio Probe SR-R thesr are ir SR-R tovir Ba, I
Hincl exten
genor
bottoi
ol
> 1
A u, a: B
5-0kb- -
-_.kb-._-_...
3 2
kb-FIG. 2. (A) Southern
hybridization
oftotal cellular DNA from SR-RSV- and RSV-29-infected CEF. Theprobe
used is showninpanelB. (B)Size of
src-containing
EcoRIfragment expected
fromanondefective RSV
(SR-RSV
orPR-RSV)
andfromadefective RSV withadeletion ofenv(BH-RSVorRSV-29).Thesizes of theEcoRlfragmentsareindicatedinkb belowthe
figures. Enzymes:
R, EcoRI;H,
HindlIl;
K, KpnI.an excess ofEcoRI. A fraction ofthe
digested
DNA the src gene of SR-RSV-A(NY)
andhybridized
with a.lectrophoresed througha0.8%agarose
gel,
transferred5.0-kbp
EcoRIfragment
of RSV-29proviral
DNA(Fig.
2).
trocellulose, and
probed
with the0.8-kilobase(kb)
PvuII The rest of thedigested
DNAwasfractionated under iden-nentofpTT107. Thisfragment
containedthe 3' third of tical conditionswithappropriate
molecularweight markers,
and the
5.0-kbp
DNAfragment
wasrecoveredby
electroelu-tion. This DNA was cloned into lambdagtWES,
and therecombinant
plaques
were screenedby
hybridization
withthesame0.8-kbPvuII
fragment
ofpTT107
describedabove. p 'Ppp n pn p n P p n p p n P One of the positive recombinants was purified by three rounds of plaque purification, and the 5-kbp src-containing EcoRIfragment
was subcloned into the EcoRI site oflb-e
3#
3 3| ' -'w
pBR322
to constructplasmid pR29.
Allprocedures
used IWw were essentially as described previously (14).
:b- * j " _ - Restriction
mapping.
pR29
wasdigested
withsingle
andmultiple
restriction enzymes, and the DNAfragments
were fractionated by electrophoresis through agarose oracryla-E 2 3 4 5 b. 7 B ,ol midegels and stained with ethidium bromide. Themap was
confirmed
by partial digestion
of32P-end-labeled DNA withappropriate
restriction enzymes.LTR g° Fe e^: ,~ re ~2 DNA sequencing. The KpnI-to-NcoI fragment measuring 300 base
pairs (bp)
onthe 5'sideofsrcwassubcloned fromR 53 8 pR i -A 83 pR29 into replicative forms of
M13mpl8
andM13mpl9
+
^____
j
4
w.
between theKpnI
andHinclI
sites. The450-bp
PstI-to-EcoRI
fragment
onthe3' side ofsrc was subclonedbetween the PstI and EcoRI sites of theM13mp8
andM13mp9
Pj;SAT!VE
P.JA
GENDM
RSV-29 RNA4S _NA,
IC
IN NP_
CLIRNONS
E _ F .Oreplicative
forms.
The
recombinant
plasmids
were
used
to
SUB
SGNB-EiM.,-" transform Escherichia
coli
JM101,
whichwasplated
on YT26Kb-s->j
<-~--
--$~XE- - - ---agar
(16)
containing
isopropylthiogalactoside
anddibromo-i.
1dichloro-indolylgalactoside.
Whiteplaques
were screened oducing(n)
and producing(p)
clones of CEFinfected
with for the presence of insertsby gelelectrophoresis analysisof*29 alone and a mixture of RSV-29 and RSV-29-associated the size of phage DNA from culture supernatants (16). respectively,wereanalyzed byNorthernblottingandhybrid- One recombinant virus of each type was selected and on with nick-translated probes indicated below the lanes. (B)
sequenced by
thedideoxy
method(20)
with both[32P]dATP
-s used for theNorthernanalysis. SR-A, Representationof the and
[35S]dATP (New England
NuclearCorp., Boston,
.SV-A genome.The arrow indicates thesplice acceptorsite for Mass.). The products of the sequencing reactions were
,cmessage. IC, Intercistronic regionof RSV. Theprobesused fractionated on 8 and 5% polyacrylamide sequencing gels in ndicated by lines below the corresponding segments of the buffer consisting of 50 mM Tris base, 50 mM boric acid, and
aSV-A
genome.A+ belowaprobe indicatesthat ithybridizes
2 mM disodium EDTA. Each fragment was sequenced at -alRNA;a- indicatesnohybridization. Enzymes: R,EcoRI, leasttwice on either strand.5amHI;
K,KpnI;
X,XhoI;
S, Sall; N, NcoI; A, AccI; Hthe
Dn
eneswerand.
[I;Bg,BglI;P,PvuII. The hatched box indicatestheminimum The DNA sequences were analyzed with the computer
Itof thedeletion thatwouldexplain the results. Theputative programsin theSequence
Analysis Package designed
bythe mic and subgenomic RNAs from RSV-29 are shown at the BiomathematicsComputation Laboratory
of theUniversity
m. of
California,
SanFrancisco.on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.318.560.74.211.2]RESULTS
Defectiveness of RSV-29. Intheearly 1960s, Simons found thatRSV-29isreplication defective (P. J. Simons, personal communication). Toconfirm the defectiveness of thevirus, CEFinfected withhighdilutions(multiplicityof infection of 10-5)of the initial RSV-29 stock wereplatedin softagarfor the isolation ofcolonies. About 70% of the isolatedcolonies, which originally did not produce transforming virus, re-leasedhigh titers of transforming virus upon superinfection
withhelper viruses (RAV-1 orRAV-2). By the interference assaywith avianleukosis virusof knownsubgroups (6, 34),
we found that the original RSV-29 stock is of subgroup A. Since the virus appears tobedefective intheenvgene(see
below),this subgroup specificity representsthatofahelper
virusassociated with theoriginal stock of RSV-29.
Analysis of RSV-29 RNAs. The total poly(A)-containing RNAs obtained from RSV-29-infected NP clones and the clone doubly infected with both RSV-29 and RSV-29-associated virus were analyzed. Representative results of oneNPclone andoneproducercloneareshown inFig. 1A. The NP clones contained two species of RSV-29-specific RNAs, measuring 7.7 and 2.6 kb, detectable by several probes. The doubly infected producer clones contained an additional 2.8-kb species ofRNA. The 7.7-kb RNA ismost likely thegenomic RNA, and the2.6-kb RNA is thespliced src mRNA ofRSV-29. The producer clone also contained the helper viral genomic RNA that could not be separated from the 7.7-kbRSV-29genomicRNA but thatwas revealed by hybridization with probes derived from env and the intercistronicregionbetween envand srcof the SR-RSV-A
genome. The 2.8-kb RNA appeared to be the RSV-29-associated virus env mRNA. Neither the 7.7-kb nor the
2.6-kb RNA ofRSV-29hybridizedwithprobesderivedfrom env and intercistronic regions. The result of the hybridiza-tion ofRSV-29 RNAs withvarious SR-RSV-A-derived DNA probes is summarized in Fig. 1B. Sinceprobes 5, 6, and 7 (Fig. 1B)failedtohybridizetoRSV-29genomicRNA while allotherprobes did,it is concluded that theRSV-29genome
containsadeletion in theenvregion. The 5'boundaryof the deletion islocated between the BamHI and XhoIsites,since thegenomicRNAhybridizedtoprobe4.The3'boundaryis locatedbetween the AccI site and thespliceacceptorsite for src mRNA(position -76 from the initiation codon ofsrc), because probe 8 hybridized with the 7.7-kb genomic RNA and with the 2.6-kb mRNA. There was no evidence of deletions in theotherreplicative genesorinthesrcgene.
The 32P-labeled RNAs from cells infected with RSV-29 and its helper virus were also analyzed by RNase
T1-resistantoligonucleotide fingerprinting. Thepatternsof both viralRNAs, except for thesrc-and env-specific oligonucle-otides, superimposed (data not shown). The pattern ofthe non-src oligonucleotides is different from those of several
strains of helper viruses originally isolated from BH-RSV (RAV-1, RAV-2, RAV-3, andRAV-7).
Molecular cloning ofa 5-kbp pol-src fragmentof RSV-29.
Whendigestedwith EcoRI,the RSV-29proviral DNAgave a 5-kbp fragmentthat hybridizedwith aprobefromthesrc gene(Fig. 2). These datasuggestthat thegenomicstructure ofRSV-29 is similarto that ofBH-RSV (13) inwhich the
EcoRIsites ingag and U3arepreservedbut the site in env is lost. This 5-kbp fragmentwaspreparedfrom CEF trans-formed clonally with RSV-29, molecularly cloned into
lambdaphagevector, and then subcloned intopBR322. Restriction site mapping ofpR29. The restriction map of
the EcoRI fragment of RSV-29 is presented and compared
with those of PR-RSV-C and BH-RSV (Fig. 3). In thepol
gene, the map of RSV-29 is the same as that of PR-RSV with twoexceptions: (i) an extra PvuII site is present near the 3' end of pol which is absent in PR-RSV, SR-RSV, and BH-RSV, and (ii) a BamHI site at the 3' end ofpolthatis present in PR-RSV but is absent in RSV-29. BH-RSV also lacks thisBamHI site. In the src gene, the restriction maps of PR-RSV andRSV-29 are similarexcept for the absence of one BglII and two BalI sites in RSV-29. None of the restriction siteslocatedinthe env geneofPR-RSV (23) were found in RSV-29. Furthermore, the restriction fragments
bridgingacrosstheputativedeletion in pR29, e.g., the NcoI
1.2-kbp fragmentand theBalI1.3-kbp fragment, comigrated on electrophoresis with the corresponding fragments from
pBH-beta (13), suggestingthat the sizes of the deletions in the viruses RSV-29and BH-RSV are very similar.
pol-src junctionin RSV-29. Tostudy the deletion in detail, the300-bp KpnI-NcoI fragment of pR29 was cloned into M13 phage vectors in both orientations and sequenced. The sequence from the KpnI site inpolto the Ncol site at the
initiation codonof src ispresentedinFig. 4A.
The sequence across the deletion of env is exactly the same as in BH-RSV (Fig. 4B). Following the termination codon ofpol (asdefined inPR-RSV-C) are 6nucleotidesthat arealsopresent inPR-RSV-C,andimmediately3' tothis are the 91 nucleotides preceding the initiation codon of src in
BH,PR-C,andSR-AstrainsofRSV,aswell asin recovered avian sarcoma virus 1441and c-src(13, 23, 28, 29).
As shown inFig.4A anddiscussed below(seeFig. 7), the RSV-29 clone we studied has amutation in the 3' endofpol.
It has 13 basesreplacing2basesatthe samepositionin the sequenceof PR-RSV(23)whichwouldresultinaframeshift in the
carboxy-terminal
partof the polprotein, replacingthe last 28 amino acids of the PR-RSV pol protein with 43 different amino acids. Thenewreading frame is terminatedby
anopal termination codonat-61relativetotheinitiationcodon ofsrc.
3'end ofRSV-29.The
450-bp fragment
fromthe PstI siteRpoI PR-C I.rI Ps HH
Hp R BH-RSV
I-4.A
A K gp85
I
Xb Bo tBg Ba X B
BlN P
N
+4+ + +
+ (s +
II IIIII,,,I, ,I ++4+ + +4* 4. +
+ 4 +
gp37 src p R
R|P |N
Ps,).sl
lt
B i B H Ps BINBI BgBp Pss
R
4.ll 11 ---- IIII.IA---4.-4r--.
_ + _ _ _- __6 A++& __ v +.+ _ +
+^16
R
,, ,, oil II I I
__ -_
+ -+ __-++ + _+
[image:3.612.312.562.468.603.2]0.2Kb inRSV-29
ProvirolDNA
FIG. 3. Restrictionmapof RSV-29proviralDNAcomparedwith PR-RSV-C(PR-C) (23)andBH-RSV(13) betweentheEcoRI site in
gag and theEcoRI site inU3. A +or- signindicates whetherthe site waspresentorabsentin BH-RSV and RSV-29. Adotted line marking an enzyme site indicates a site present in BH-RSV or
RSV-29 which is absentinPR-RSV. Wheretworestrictionenzyme
sites cannot be resolved in the scale of this diagram, they are
indicated inaverticalarrayatthesamesiteonthe PR-RSV-Cmap;
thisarrayis thesamefor the BH-RSV andRSV-29maps.Enzymes: R,EcoRI;Ps,PstI;H,HindIII; Hp,HpaI;Xb, XbaI; Ba, BamHI; Bl, BalI; N, Ncol; A, AccI; Bg,Bglll;X,XhoI;P, PvuII;B,BgI; K, KpnI. A indicates that PstI sites were not determined for BH-RSV(13).
RSV-29
I
on November 10, 2019 by guest
http://jvi.asm.org/
[A)
KpnI
KpI- 10 20 30
GGTACCCTCT CGAAAAGTrA AACCCACAT
70 80 90
TGAGCCGAG CCTClTG CAGCATITC
I
env splice acc
40 50 60
CACCCAAAAG GATCACGTGA CTAAGAAAGA
100 110 120
TGATTGGATA CCCAGXGAGA AACCCCCAGC pol mutation
130 140 150 160 170 180
AACAAGCAAG AAOGACTCCA GCGAGAAACC GCCAGCAACA ACCAAGAAAG ACCCGGAGAA -91
src
190 200 210 220
GACACCCITT CTCCCAACGA GACTTAATrA TAlTGTCTGT
iT---r---pol Aenv stop in RSV-29 PR-C
230 240
CTCC1~CG AGCIAGCA
src pol
splice acc stop RSV-29
250 260 270 280 290 300
ACTCTCCIGG TGGCCTCGCG TACCACI=i GCCAGGCGGT AGCTGGACA GTGCAGCCCGA start
src
1309 CCACCATGG
NcoI
[B]
RSV- 29 BH-RSV PR(pol) PR(src)
PR-C src R29
pol src spl pol
stop -91 acc stop
__ I_ _
GAAGACACCCI CCA T[
******************** ******C**AT*A*IGtIC
*TAG*CTA**************************A*********
10/15
src start RSV-29 'IXnXX TCCTCC ACI;4CCCCACGCCGTASCTOCCACACTGCACCCGACCACCATCG BH-RSV *****A***************************************-******************* PR(src) *********************************************-*******************
[C]
PR-C src
pol v-src splice
stop -91 acc
RSV-29 CGAGATTAAITATATI77CTGTGT GCMCAGGAGCGAGCTIGAC
PR(pol) *C**AT*A*7UG7=***G *7CAGGA
c-src TCTGACACC C*GTC***CTCG*** **************** 12/19
FIG. 4. (A)DNAsequenceof309-bp KpnI-NcoIfragment of RSV-29.Thepolmutationrefersto 13 basesinthepol geneofRSV-29 which replaced2bases in thesameposition in PR-RSV-Candisdiscussed in detail in the legendtoFig.7.(B)RSV-29sequencefromtheend of poltotheNcoIsitecompared withthatof BH-RSV andwiththe sequence of PR-RSV pol[PR(pol)] and thatof PR-RSV from 104bases
upstreamfromthe initiation codon ofsrc[PR(src)].The box shows aputative recombination point inanondefective RSVthatcouldgenerate
the deletion inRSV-29orBH-RSV. (C) Sequence of RSV-29acrossthedeletion ofenvcomparedwith that ofPR(pol) (seeabove) and the regionupstreamofc-src(c-src).The box shows anotherrecombination pointbetween the 3' endof poland the region upstreamofc-src that
would produceadefective virus like RSV-29.Inpanels B andC,a*indicatesthe same nucleotide as RSV-29, and a-indicates a nucleotide thatispresentinRSV-29 butabsentin the othersequences.
atthe 3' endofsrcto the EcoRIsite in U3 wascloned into
M13mp8andmp9vectorsandsequenced(Fig. 5A). There is
extensivehomologybetweenPR-RSV-C (23) and RSV-29 in theregions designatedasF3(30),FlA,
FiB,
FlD(13),PPT,and U3 (Fig. 5B). FlC is interrupted in PR-RSV-C by F2 sequences. In contrast, the
FiC
of RSV-29 is uninterrupted andhomologoustothatof SR-RSV-A (13, 28).The 3' end of the RSV-29 src gene also contains the
38-nucleotide sequence that is presumably derived from the chickenc-srclocus 1 kb downstream from the c-src
termi-nationcodon, called c-src' (30). It also contains box P(Fig. SA), which has been hypothesized to be involved in the recombination betweenc-srcand c-src' (Fig. 6), and boxQ,
which may have been involved in the recombination be-tweenhelpervirus(RAV-2)andc-src' sequencestogenerate the 3' endofBH-RSV (13)(Fig. 6).
on November 10, 2019 by guest
http://jvi.asm.org/
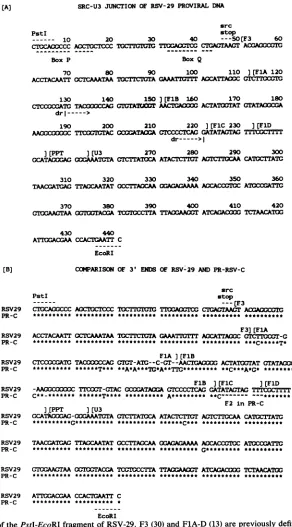
[image:4.612.134.475.74.560.2](A) SRC-U3 JUNCTION OF RSV-29 PROVIRAL DNA src
PstI stop
10 20 30 40 ---50[F3 60
CTOCAOCCCACCTOCTCCC TOCTI'TCT TVCGTC CTOACTAAGT ACAOOCGC
Box P BoxQ
70 80 90 100 110 J(FlA 120
ACCTACAATT CCTCAAATAA ¶IXCTTC'LA GATI:lT ACCATrAOGCCtIV
130 140 150][FlB 160 170 180
CTXXCATG TACCACGTC'ItTrA T AACTCA ACTATrTATGTATAOXA
drl--->
190 200 210 220 ][FlC 230 ][FlD AAOOCOtC TrCACO-OCATAIAGTCCCCTCAC GATATACTAC TITTCCTTTT
dr--->1
](PPT ][U3 270 280 290 300
GCATACAGOCAAATMTA CTCTATICA ATACTCTI1TACC CATOCTATC
310 320 330 340 350 360
TAACGATGIA TTACCAATAT CCTC A AAAA ACCACCG4
ATCCU&TTC
370 380 390 400 410 420
rWAACTAAOC:ACGAT =CCTTATTAAA0T ATCAGACOC TCTAACATOG
430 440
ATTGGACGAA CCACTCAATT C EcoRI
[B] COMARISON OF 3' ENDS OF RSV-29 ANDPR-RSV-C src
PstI stop
__ _ _ _ _ - --tF3
RSV29 CTGCA00CCC AOCTOCTCCC TGCT1-TUG TlAi CTGACTAACT ACGAOOCGTC
PR-C ********** ********** ********** **
F3]JFlA RSV29 ACCTACAATT OCTCAAATAAl rA CAATICiTO AOCATTrAGC CTCTiCCGr-G
PR-C ********** ********** ********** ********** **********
FlA] (FlB
RSV29 CTCCCCCATC TACXXXCCAG
GTGT-ATC--C-GT-_-AACTlAG=O
ACTAT00TATGTATACCCGA PR-C ********** ******T*** **A*A***TC*A**TTG********* **C***A*G* *********TFlB ][F1C ][FlD
RSV29 -AAM0c0C TTOCT-GTAC
GcXXATAOCA
GTCCCCTCACGATATAGTAG TCTlr PR-C C**-******* ******T**** ********** **C- --F2 in PR-C
]tPPT ][U3
RSV29
GCATtkXMAG-CCGAAATCTA
CTCTIATCCAATACTCTTGT
ACTCTI'CAA
CATCTTATCPR-C **********C*** ********** **********
RSV29 TACGATCGA TIACCAATATGCCTIAOCAA
OGAGACAAAA
AGCACcCTOCAT1CTTG
PR-C G****** **********
RSV29 CXAAAAAOCG=IACGATC A TTACAAT
ATCAGACOOC
TCTAACATOCPR-C ********** ********** ********** ******
RSV29 ATITGACGAA CCACTGAATT C
PR-C ********** **********
EcoRI
FIG. 5. (A)Sequence ofthePstI-EcoRI fragment of RSV-29. F3 (30) and F1A-D(13) arepreviously defined segmentsofthe sequences
flanking src in RSV. dr refersto the limits of the direct repeatflanking srcinSR-RSV and PR-RSVas originallydefined bySchwartzetal. (23). BoxesPandQarehypotheticalpoints of recombination required to generate the 3' endof v-src (13;legendtoFig. 6). (B)Comparison of3' ends ofRSV-29 andPR-RSV-C (PR-C). A* indicates anucleotide that is present inRSV-29, anda - inagivensequenceindicatesa
base that is absent in that sequence but present in the other sequence. A seriesof -indicates where PR-RSV-C hasacquired anF2(13) sequence.
DISCUSSION andHanafusa (13), thispol-src
junction
could beformed asaresultofrecombination betweenthehelpervirusandc-src
Deletion of env. DNA sequence data confirmed the con- usinghomologous sequences existingdownstreamfrom
pol
clusionfrom RNA analysisthat RSV-29 containsadeletion in PR-RSV-C(sequencesofpoland its
neighboring
regions of the entire env gene. The deletion involves exactly the of helper viruses have not been determined, but they are samesequences as inBH-RSV (13).Asdiscussedby Lerner probably similar to those ofPR-RSV-C) and upstream ofon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.156.448.72.599.2]Termination codonfor
c-src C-SRC'
54 1 14 17 1
11 5
V-SRC 47
14 17 115
Termination
codon for v-src
3' END OFHELPER VIRUS
3' END OF RSV
FIG. 6. Model for the generation ofthe 3' end of v-src (12).
Boxes P and Q are putative recombination points between c-src,
c-src' (30) and 3' end of helpervirus. The lengths of the relevant
regions in number of nucleotides are indicated. Recombination 1
probably took place duringreversetranscription. Recombination 2
could have happened by a rearrangement at the c-src locus, by
aberrant processing ofahelper virus-c-src hybrid RNA, orby an errorinreversetranscription.
c-srcinthegenome ofchickens (Fig. 4C). Such a recombi-nation wouldcause adeletion of the entireenvsequenceand produceareplication-defective virus. Alternatively, similar homologous sequences existing downstream from pol and upstream from v-src within the PR-RSV-C genome could also be utilizedtocausetheenvdeletionin thenondefective virus (Fig. 4B). Thus, as with BH-RSV, analysis of the sequence across the deletion of env fails to distinguish whether the deletion occurred during the generation of
sarcoma virus inchicken tumor no. 1 or occurred second-arily inanoriginally nondefective PR-RSV-C-like virus.
The exact homologies in the deletion in BH-RSV and RSV-29 suggest that thesetwoviruseshavedescended from
a commonprogenitorvirusand that the differences in their3' sequences might have arisen in BH-RSV by recombination withhelper viruses during subsequentpassages(seebelow). The fact that a strain of RSV with the least number of
passages beyond chicken tumor no. 1 has deleted its env
A RSV-29 #1. 2 PR-RSV-C
lBlr/2][ Xr H A2
R
gene and istherefore replication defective suggests that the
original isolate of RSVwasalso replication defective. This viewis supported by the observations of Hagino-Yamagishi et al. (5), whofound that two independently isolated avian sarcomaviruses(Si and S2), obtained by inoculating chick-ens with avian lymphoid leukosis virus, contained src and were replication defective. This implies that src, like other oncogenes, canbe transduced into avian lymphoidleukosis
virus toform a replication-defective transforming retrovirus. Furthermore, while replication-defective mutants generated
from nondefective virus have been selected by isolating
single colonies (10, 15), it is difficultto explain how a pure stockofadefective virus (e.g., RSV-29 and BH-RSV) could be established bypassageof nondefective virus inchickens
in which defective viruses do not appear to have any selective advantage over their nondefective counterparts.
Taking all these arguments into consideration, we propose that, like all other transforming retroviruses, RSV was
originally isolated as a replication-defective virus, and that the commonly studied replication-competent strains have been generated during subsequent recombinations with helper virus.
Mutation in pol. The discovery ofthe 13 nucleotides in RSV-29 substituting for 2 nucleotides in pol (as compared
with PR-RSV-C [23] and BH-RSV [M. Sudol and H.
Hanafusa, unpublisheddata]) is interesting. We donotknow whether the mutation and theresultingmissensetranslation
arepeculiartotheparticularNPclonethatwassequencedor arepresentinallclonesof RSV-29. It would be interestingto study thefunctionalandstructuralproperties ofthepol gene product in this clone. On the basis of the findings of Hippenmeyer and Grandgenett (9), the mutation in this
domain is expected to leave the DNA polymerase and
RNase-H functionsofthe enzyme intact.
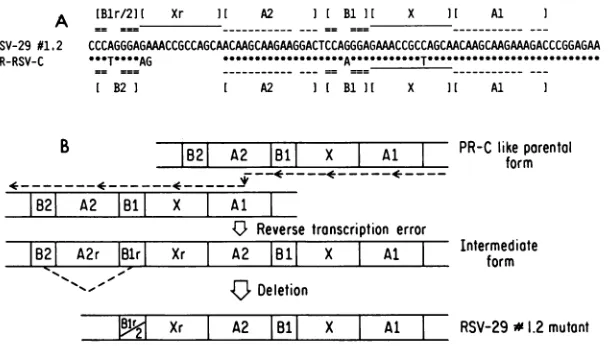
Apossible mechanism forthegenerationof this mutation ispresentedinFig.7, basedsolelyonsequencecomparisons
andidentificationof direct repeats. Severalprevious
hypoth-esespostulated involvement of direct repeatsin the genera-tionoftransformation-defective deletion mutantsfrom RSV
(2, 18, 19). Since PR-RSV and BH-RSV (M. Sudoland H.
Hanafusa, unpublished data)arehomologousin sequence in thepol gene, it is reasonableto assumethat the
PR-RSV-C-like structure,containingthedirectrepeatsAl and A2,isthe
parental virus. After having reverse transcribed
A1-X-B1-I A1-X-B1-I B1I I X II Al
== =_=
CCCAGGGAGAAACCGCCAGCAACAAGCAAGAAGGACTCCAGGGAGAMCCGCCAGCAACAAGCAAGAAAGACCCGGAGAA
.*T**AG*--T----AG
---A---
--T---
---I B2] I A2 I[ B1 ] X ] Al I
DD_r p;
IB21 A2 B181I X Al 1fKL lKform
B21 A2 1131
I
X Al4U-
Reverse transcription errorB21
A2rlBir
I
XrI
A2|B11
XI
Al IIntermediate
form~Deletion
renaui
p
~2 Xr I A2 |18| X Al I RSV-29#1.2mutontFIG. 7. MutationinpolinRSV-29no.1.2.(A)Comparison of the 3' ends ofpol in RSV-29no.1.2and inPR-RSV-C(22). A*indicates thesamebaseas RSV-29,andablankspaceindicates the absence of that base in PR-RSV-C.Al,A2 andBi, B2arepairs of directrepeats presentinthePR-RSV-Csequence.(B)Diagram oftheerrorinreversetranscription and the deletion usingthesedirectrepeatsthat would generatethemutationsequencedin RSV-29no. 1.2.
C-SRC
14
zI,I
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.59.299.75.239.2] [image:6.612.159.463.530.703.2]RAV-O
KI_-SR-RSV
DJ-E
BH-RSV
D_-E
GAG
POL
liz'
IZZHID
ENV
GAG
POL
ENVSRC
4
I
I
MH2
GAG POL
SRC1
I
RAV-2
GAG
POL
SRC
RSV-29
m
I
PR-C Minus F-2
FIG. 8. Comparison of the 3' ends of several avian retroviruses to show the regions across which certain pairs of viruses are related. The
arrowed box ineachviruscorrespondsto box Q of Fig. 5 and 6.
A2, the enzyme could have fallen off the template and then reattached, possibly to a second template, at
Al
because of thehomology betweenAl and A2. This would generate theintermediate form shown in the figure, with repeats of
X-Bl-A2 (Xr-Blr-A2r). A subsequent deletion, using the
direct repeats Blr and B2, could generate the variant we have identified.
3' endof RSV-29. Comparison of the sequence atthe 3'
end of RSV-29 reveals analmost exact homology between RSV-29 and PR-RSV-C in this region, suggesting that the two viruses are closely related (Fig. 5B). In line with the
discussion above concerning the deletion in env, this might mean that PR-RSV-C is derived from RSV-29and that the
insertion of F2 in theFlCsequenceof PR-RSV-C occurred at a later date. However, an alternative possibility is sug-gestedby thereportedextensive homology of3' sequences in
SR-RSV-A
andMH2
leukemia virus (27). The fact thatpairs of viruses like MH2 and SR-RSV-A, which certainly have had
different
origins and different passage histories,share common 3' sequences, suggests that in the course of
passage in
chickens,
theseviruseshavebeen associatedwith a common pool of helper viruses with which they often exchange their3' sequences inblock. Superimposed on thisv-src
PR-RSV-C
RSV-29 SR-RSV-A BH-RSV RAV-2 Y73 MH2 UR2
c-src'
G¶EGIGGAGGTCGCTGAG TAAGTACGAGGC
__________________
-G-TGCCGCGA---A---A --GTA-ACTT-T
A-A-A---A A-A-A---A--- A--- -A-A---GA-CG
IOB
--GTA-ACTT-T --GTA-GCTT---ACA -GGTA-GCTT-- ATCTGCAC-TC-FIG. 9. Ahighly homologous sequencepresentatthe 3' ends of several avianretroviruses andinc-src'.The box labeledv-srcrefers tothe 3' endofthesrcgenein the RSVs. A-denotes thesamebase
atthatpositionasin PR-RSV-C.BoxQis thesame asinFig.5 and 6.
arepoint mutations, deletions, andinsertions which do not
obscuretherelatedness of the 3' sequences. Thehomology
between RSV-29 and PR-RSV-Cfoundinthisstudy couldbe
analogousandmight merelyreflect the fact that these viruses haveacquiredtheir 3' sequencesby similar exchangeswith a commonhelper virus. Thishypothesiswould also propose that F2 wasinsertedinto PR-RSV-C at alater date.
The relationships discussed aboveand that between
BH-RSV andRAV-2(1, 13),areshowninFig. 8. Acomparison of these 3' sequences revealed a highly homologous short
stretch in all these viruses(Fig.9). Thishomologousregion
is present at the 3' end of the src sequence in theRSVs, from
box Q to the termination codon (Fig. 9). The homology betweenhelpervirus sequence and c-src' inthisregionwas
postulatedtobe responsible for the recombinationatthe 3'
endrequiredtogenerate RSV inchickentumor no. 1(1, 13) (Fig.6and9). Itisinterestingtoobserve that this sequence
isrelativelywellpreservednotonlyintheRSVs, but alsoin RAV-2 (1), MH2 (27), Y73 (11), and UR2 (17) in their 3'
noncodingregions, suggesting thatthis sequenceispresent in severalhelperviruses that have notbeenassociated with
anyofthe RSVs and so its presencecouldhot beanartifact
created by exchange of3' sequences between helpervirus andsrc-containingRSV(Fig. 9).This is inconcordance with our hypothesis, because it affirms that box Q could have been present in the original helper virus that gave rise to RSV inchickentumor no. 1.
ACKNOWLEDGMENTS
Wethank H.Iba forhelpful discussions,andS.Sugano,R.Jove,
and F. Crossforreadingthe paper. Wearealsograteful toR. M.
Doughertyforprovidingastock of RSV-29and to P. J. Simon for communicating earlierresults.
This work was supported by Public Health Service grants
CA14935andCA18213toH.H. and CA29339toL.-H.W.fromthe National CancerInstitute. L.-H.W. isarecipientof PublicHealth
Service Research Career Development Award CA00574from the
National CancerInstitute. A.D. was supportedin partbya grant from R. J.Reynolds Industries,Inc.
LITERATURECITED
1. Bizub, D., R. A. Katz, and A. M. Skalka. 1984. Nucleotide
sequence of noncoding regions in Rous-associated virus-2:
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.181.465.67.267.2] [image:7.612.73.291.564.671.2]comparisons delineateconserved regions important in replica-tion andoncogenesis. J. Virol. 49:557-565.
2. Coffin, J. M. 1979. Structure, replication and recombination of retrovirus genomes: some unifying hypotheses. J. Gen. Virol. 42:1-26.
3. DeLorbe, W. J., P. A. Luciw, H. M. Goodman, H. E. Varmus,
andJ. M. Bishop.1980.Molecularcloning and characterization of avian sarcoma virus circular DNA molecules. J. Virol. 36:50-61.
4. Duesberg, P., S. Kawai,L.-H.Wang,P. K.Vogt,H. M.Murphy,
and H. Hanafusa. 1975.RNAofreplication defective strains of Rous sarcoma virus. Proc. Natl. Acad. Sci. U.S.A. 72:1569-1573.
5. Hagino-Yamagishi, K., S. Ikawa, S. Kawai, H. Hihara, T.
Yamamoto, and K. Toyoshima. 1984. Characterization oftwo
strains of avian sarcoma virus isolated from avian lymphatic leukosisvirus-induced sarcomas.Virology 137:266-275.
6. Hanafusa, H. 1965. Analysis of the defectiveness of Rous sarcomavirus. III.Determining influence ofanewhelper virus
on the host range and susceptibility to interference of RSV. Virology 25:248-255.
7. Hanafusa,H.1969. Rapid transformation of cells by RSV.Proc.
Natl. Acad.Sci. U.S.A. 63:318-325.
8. Hanafusa, H.,T.Hanafusa,and H. Rubin.1963. The defective-ness of Rous sarcoma virus. Proc. Natl. Acad. Sci. U.S.A. 49:572-580.
9. Hippenmeyer,P.J.,and D. P.Grandgenett. 1984.Requirement of the avian retrovirus pp32 DNAbinding protein domain for replication. Virology 137:358-370.
10. Kawai, S.,and H.Hanafusa. 1973.Isolation of defectivemutant
of avian sarcoma virus. Proc. Natl. Acad. Sci. U.S.A. 70:3493-3497.
11. Kitamura, N., A. Kitamura,K. Toyoshima, Y. Hirayama, and M. Yoshida. 1982. Avian sarcoma virus Y73 genome sequence andstructural similarity of its transforming geneproducttothat of Rous sarcomavirus. Nature(London) 297:205-208. 12. Klement, V., and J. Svoboda. 1963. Induction of tumors in
Syrian hamsters bytwovariants of Roussarcomavirus. Folia Biol. (Prague) 9:181-186.
13. Lerner, T. L., and H. Hanafusa. 1984. DNA Sequence of the Bryan high-titer strain of Rous sarcoma virus: extent of env
deletion andpossible genealogical relationship with other viral strains. J. Virol. 49:549-556.
14. Maniatis, T., E. F.Fritsch, and J. Sambrook. 1982. Molecular cloning: alaboratory manual. ColdSpring Harbor Laboratory, ColdSpring Harbor, N.Y.
15. Martin, G. S.,K.Radke,S.Hughes,N.Quintrell, J.M.Bishop,
and H. E.Varmus. 1979. Mutantsof Rous sarcoma virus with extensive deletionsof the viral genome. Virology 96:530-546.
16. Messing, J. 1983. New M13 vectors for cloning. Methods Enzymol. 101:20-78.
17. Neckameyer, W. S., and L.-H. Wang. 1985. Nucleotide se-quence of avian sarcoma virus UR2 and comparison of its transforming gene with other members of the tyrosine protein kinase oncogenefamily. J. Virol. 53:879-884.
18. Omer, C.A., K. Pogue-Geile,R. Guntaka, K. A.Staskus, and
A.J. Faras. 1983. Involvement ofdirectly repeated sequences in the generation of deletions of the avian sarcoma virus src gene. J.Virol.47:380-382.
19. Parvin, J. D., and L.-H. Wang. 1984. Mechanisms for the
generation of src-deletion mutants and recovered sarcoma vi-ruses:identification of viral sequences involved in src deletions and in recombination with c-src sequences. Virology 138:236-245.
20. Sanger, F., S. Nicklen, and A.R.Coulson. 1977. DNA sequenc-ing with chain-terminatsequenc-ing inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74:5463-5467.
21. Scheele, C. M., and H. Hanafusa. 1971. Proteins of helper-dependentRSV. Virology 45:401-410.
22. Schmidt-Ruppin, K. H. 1964. Heterotransplantation of Rous sarcoma and Rous sarcoma virus to mammals. Oncologia 17:247-272.
23. Schwartz, D.E., R.Tizard, and W. Gilbert. 1983. Nucleotide sequenceof Rous sarcomavirus. Cell 32:853-869.
24. Shibuya, M., and H. Hanafusa. 1982. Nucleotide sequence of Fujinami sarcoma virus: evolutionary relationship of its
trans-forming gene with transtrans-forming genes of othersarcomaviruses. Cell 30:787-795.
25. Shibuya, M., H. Hanafusa, and P. C. Balduzzi. 1982. Cellular sequencesrelatedtothreenew oncgenesof aviansarcomavirus (fps, yes, and ros) and their expression in normal and
trans-formed cells. J. Virol.42:143-152.
26. Simons,P.J., andR.M.Dougherty. 1963.Antigenic character-istics of three variants ofRous sarcomavirus.J. Natl. Cancer
Inst.31:1275-1283.
27. Sutrave, P., H. W. Jansen, K. Bister, and U.R. Rapp. 1984. 3'-Terminal region of avian carcinoma virus MH2 shares
se-quenceelements with aviansaracomavirusesY73and SR-A. J. Virol. 52:703-705.
28. Takeya, T., R. A. Feldman, and H. Hanafusa. 1982. DNA
sequence of the viral and cellular src gene of chickens. I. Complete nucleotide sequence ofanEcoRI fragment of
recov-ered aviansarcomavirus which codesfor gp37 andpp60rc.J. Virol.44:1-11.
29. Takeya, T.,and H.Hanafusa.1982.DNAsequence of the viral and cellular src gene of chickens. II. Comparison of the src
genesoftwostrains of aviansarcomavirus and of the cellular homolog. J. Virol. 44:12-18.
30. Takeya, T.,andH. Hanafusa. 1983. Structure and sequence of the cellular gene homologous to the RSV src gene and the mechanism for generating the transforming virus. Cell 32:881-890.
31. Takeya, T., H. Hanafusa, R. P. Junghans, G. Ju, and A. M.
Skalka. 1981. Comparison between the viraltransforminggene (src) of recovered aviansarcomavirus andits cellularhomolog. Mol.Cell. Biol. 1:1024-1037.
32. Wang, L.-H.,P. Duesberg, K. Beemon, and P.K. Vogt. 1975. Mapping RNase
T,-resistant
oligonucleotides of avian tumorvirus RNAs: sarcoma-specific oligonucleotides are near the poly(A) end and oligonucleotides common to sarcoma and transformation-defective viruses are at the poly(A) end. J.
Virol. 16:1051-1070.
33. Wang,L.-H., R.Feldman,M. Shibuya, H. Hanafusa,M. F. D.
Notter,andP.C.Balduzzi.1981.Genetic structure, transforming sequence, and gene product of avian sarcoma virus URL. J.
Virol.40:258-267.
34. Weiss,R.1982. Experimental biology and assay of retroviruses in RNA tumor viruses. Cold Spring Harbor Monogr. Ser. 1OC:209-260.