0022-538X/89/062657-10$02.00/0
Copyright © 1989, American
Society
forMicrobiologyMultiple
EBNA1-Binding
Sites
Are
Required
To Form
anEBNA1-Dependent Enhancer
and
To
Activate
aMinimal
Replicative
Origin
within
oriP of
Epstein-Barr Virus
DOREEN A. WYSOKENSKI ANDJOHN L. YATES*
Departmentof Human Genetics, Roswell Park Memorial Instituite, Buffalo, New York 14263
Received 1 December1988/Accepted 8March 1989
EBNAl activatesthe EBV plasmid maintenance sequenceoriP by bindingtoitstwoessential regions. One
region isafamily of 30-base-pair (bp)repeatsandis activated by EBNA1to actas atranscriptionalenhancer. The other region contains a 65-bp dyad symmetry and lacks enhancer function. To explore the functional
differencesbetween thetworegions,wedeterminedoriPactivitiesasfunctions of thenumberof30-bprepeats
andcompared them withactivities determined when tandem copies ofthe dyadsymmetryregionwereusedto
replace the 30-bp repeats.Three conclusions have been drawn. (i) Activation of the 30-bprepeatsby EBNA1
toenhancetranscriptionortopermit plasmid maintenance isahighly cooperativeprocessinvolvingatleastsix or seven30-bprepeatsforfullactivity. (ii) Tandemcopiesofthe dyadsymmetryregion cooperativelyenhance
transcription butarelesseffectivethan 30-bprepeatsprovidingasimilar number ofEBNAl-bindingsites.(iii)
Tandemcopies ofthe dyadsymmetryregion alone cooperatively activate replication, suggesting thatthe region
contains theactual origin ofreplication. We alsoreport thatwhile rodent-derived cell lines do not support
replication of EBV-derived plasmids they do permit EBNAl-dependent enhancer activity. EBV plasmid
replication thus requires the interaction of EBNA1 or oriP with a host factor that is not required for
enhancement of transcription.
The170-kilobase-pair (kbp)genomeof the human
herpes-virusEpstein-Barr virus (EBV) is maintained extrachromo-somallyinlatentlyinfected cells (24)as acovalently closed,
circularplasmid (18)thatisreplicated duringSphase ofthe cellcycle (13). The EBV genomeencodes two components
known to function in plasmid maintenance: an 1,800-bp
cis-acting region, oriP,thatsupportsthestablemaintenance ofrecombinant plasmids carrying it (37), and the nuclear antigen, EBNA1, that is required for oriP to function in permissive cells (19, 38). Since EBNA1 is the only EBV-encoded protein requiredforplasmid maintenance, plasmid replication is presumedto be performed largely by thecell DNAreplication machinery, directed by EBNA1 to initiate replicationwithinoriP(see reference 21forareview).
Two salient characteristics of oriP and its activation by EBNA1 that are common features of the control of DNA replication in both procaryotes and eucaryotes are the
involvement of multiple initiator protein-DNA interactions (8)and oftranscriptional control elements (5). oriP is
com-posedoftwoessentialregions(31), eachcontaining multiple EBNA1-binding sites(28) (Fig. 1A). One essential regionis composed ofapproximately 20 tandem copies of a 30-bp sequence, each copy containing an EBNA1-binding site.
This family of 30-bp repeats acts as a transcriptional
en-hancer inastrictlyEBNA1-dependent manner(30, 31).The other essential component oforiP, located almost 1,000 bp from the 30-bp repeats, contains two symmetrically posi-tionedEBNA1-binding sites withina 65-bp dyadsymmetry
and two additional EBNA1-binding sites flanking the
sym-metrical sequence. This region, referred to as the dyad symmetry region, lacks significant enhancer activity. By analogy to known replicative origins, particularly those of
simian virus 40 and polyoma virus, for which bidirectional
DNA synthesis begins at a region ofdyad symmetry
con-*Correspondingauthor.
taining symmetrically positioned binding sites for the
initia-torprotein large Tantigen, it was suggested that the dyad symmetry region oforiPcontains theactual origin of repli-cation(31). The 30-bp repeatscan thus be viewed as
acti-vating this origin from a distance, perhaps through an
enhancerlike mechanism. The 30-bp repeatsand the region of dyad symmetry of oriP, in fact, exhibit the spatial independencecharacteristicofenhancer-promoter combina-tions;whilebothcomponentsoforiPmustbepresentonthe sameDNAmoleculetopermitDNAreplication,the relative orientationofthetwocomponents and thedistance between them, when varied fromzerotoseveralthousand basepairs,
areof littleimportance (31;D. Reisman, Ph.D. dissertation, University ofWisconsin, Madison, 1986).
Wesoughttoascertain howmultiple EBNA1-bindingsites in the two components of oriP are involved in apparently
distinct functions. First, the minimal numbers of 30-bp
repeats needed for full enhanceractivity and for oriP repli-cation were determined. Next, to determine whether the
functional differences between thetworegionsoforiP result
from the different numbers ofEBNA1-binding sites in the
two regions or from functional differences between these
binding sites and theirneighboring sequences, tandem
cop-iesof thedyadsymmetryregionweretested fortheactivities
of the 30-bp repeats. The results imply that efficient
en-hancerfunction and activation of oriPreplication minimally involve the cooperation ofseven, or possibly six, EBNA1
dimers. EBNA1-binding sites within the 30-bprepeats were
found to activate replication and transcription more
effi-ciently than those within the dyad symmetry region, yet tandem copies ofthe dyad symmetry regionexhibited
lim-ited EBNA1-dependent enhancer activity and could
repli-catein the absence ofthe30-bprepeats. The resultsprovide experimental evidence that the dyad symmetry region
con-tains thereplicative originof oriP and thatthe 30-bp repeats
2657
on November 10, 2019 by guest
http://jvi.asm.org/
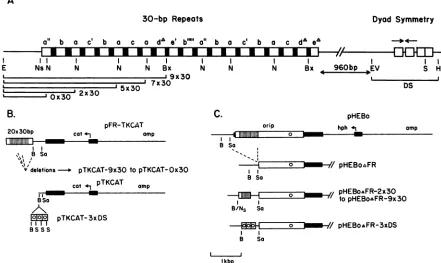
A
30-bp Repeats Dyod Symmetry
ao b a c'b a c a da 'hb""' " b a c' b a c doeA
III I I I
E Ns N N N N Bx N N N Bx 4 960bp EV S H
a . a% -'
0x30 2x30
B.
- 7x30
5x30
pFR-TKCAT
20x30bp cat 5, amp
I B So '0' II
vdeletions-- pTKCAT-9x30topTKCAT-Ox30
cat pTKCAT amp
I
BSo IAI1
5I1010 pTKCAT-3xDS
BS S S
C.
orip
B So
pHEBo
hph *a
I
pHEBo&FR
I I
B So
Ito=
pHEBo&FR-2x30
B/Ns So
_ 7/pHEBo&FR-3xDS
B So
Ikbp
FIG. 1. Structure of oriPof EBV strain B95-8(1)andplasmidsusedtostudyitscomponents.Restriction endonuclease sites areshown for BamHI(B), BstXI (Bx), EcoRI (E),EcoRV(EV), HpaI (H),NdeI(N),NsiI(Ns), Sall (Sa),andSmaI(S).(A)Structureof oriP.Segments ofthefamily of 30-bprepeatsandthedyadsymmetryregion (DS)thatweresubclonedfor functional tests(BandC)areindicatedbybrackets.
Open boxes in the30-bprepeats and within thedyad symmetry region representthe18-bp, symmetricEBNA1recognition sequences(28). Closedboxesrepresentthe12-bpspacersthat alternate withEBNA1-bindingsitestoformthe30-bp repeats. Convergingarrowsindicatethe
65-bpdyadsymmetry.For the30-bprepeats,theletterarepresents theconsensus18-bp symmetricsequence;b,c,and drepresentsequences
that differfrom theconsensus sequence atoneposition; anderepresentsasequencethat differsat twopositions. Each superscript prime symbolrepresents onebasechange fromtheconsensus sequence inthe 12-bpspacer, whileeachsuperscriptdeltarepresentsashortened
spacer.(B and C) Plasmids usedin thisstudy,shown linearizedatthepBR322-derivedEcoRI site.Thin linesrepresentE.coli-derivedDNA
(amp, ampicillin resistancegene;hph,hygromycinBphosphotransferasegene;cat,CATgene).Closedboxesrepresentthepromoter(-197
to+56[arrow]) and 3-flankingsequences(626 bp from SmaItoPvuII)of the HSV type 1 TKgene. Openboxesrepresent oriPDNA,with
vertical linesindicating the 30-bprepeatsandacircleindicating the dyadsymmetryregion.
supportits activation throughacooperativeaction of
multi-ple EBNA1 molecules.
MATERIALS ANDMETHODS
Cells and media. The EBV-positive Burkitt's lymphoma cell lineRajiandtheEBV-negativeBurkitt's lymphomacell lineWilsonwere growninRPMI 1640 medium supplemented
with 7% fetal bovine serum plus 7%Nu-serum
(Collabora-tiveResearch, Inc.). LMTK- cellsweregrowninminimum
essential medium alpha with the same serum supplement.
Wilson(p174) is a derivative of Wilson carrying multiple
copies ofthe oriP-based, hygromycin B resistance plasmid p174, whichexpressesEBNA1 (38).Wilson(p174) cellswere
maintained in medium containing 300 FLg of hygromycin B
per ml, except for a few generations before and during
chloramphenicol acetyltransferase (CAT) gene expression
studies.
Recombinantplasmids. Plasmids were constructed by
us-ing standard methods (20). pFR-TKCAT carries the family of 30-bp repeats on a fragment of approximately 900 bp
extendingfrom an EcoRI site to aBaI31 deletion endpoint
positioned between the EcoRI and BamHI sites of pBR322-derived DNA and located 2.1 kbp 3' or 2.5 kbp 5' to the thymidine kinase (TK) promoter (Fig. 1B). It was
con-structedfrompABal4(37) by replacing theneogene,located
between the promoter and polyadenylation signals of the herpessimplexvirus TKgene,with the bacterialCATgene.
pTKCATwasgeneratedfrompFR-TKCATby replacingthe
DNA between an EBV SphI site and the SphI site in
pBR322-derivedDNA withaBamHI linker, leavingonly 15 bpof EBV DNA between theEcoRI andBamHIsites (Fig. 1B). pTKCAT-9x30 was generated by deleting DNA
be-tweenthe BstXI sitenearthecenterof the30-bprepeatsand the flanking BamHI site ofpFR-TKCAT. pTKCAT-9x30 through pTKCAT-Ox30 were constructed by partially di-gesting pFR-TKCATwith NdeI,followedby digestion with BamHI, repair with the Klenow large fragment of DNA polymerase I, ligation, and a final digestion with BstXI to remove unwantedproducts of NdeI partialdigestion.
The dyad symmetry region of oriP was excised from
pHEBo (34) as a 139-bp fragment by using endonucleases
EcoRV andHpaIandwassubcloned inpUC12attheHindll site.Next,aBglIIlinkerwasintroducedintotheneighboring
PstI site of the pUC12 polylinker, generating p400. The 159-bp BamHI-to-BglII fragment containing the dyad sym-metryregionand20bpof linker DNAwasexcisedfrom p400 andinserted atthe BamHI siteofpTKCATin one,two, or
three tandemcopies, all in theorientation shown inFig. 1B (indicated by the SmaI site), generating pTKCAT-lxDS through pTKCAT-3xDS.
DS
amp
..V..
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.97.538.78.341.2]pHEBoA&FR was created by deleting DNA between the unique Sall site of pHEBo and an NcoI site immediately flanking the 30-bp repeats, regenerating the Sall site (Fig.
1C).
The shortened 30-bp repeat arrays and tandem copies of the dyad symmetry region were excised from the pTKCAT vector withNsiI
plusSalJ
and with EcoRI plusSalI,
respectively, and inserted into pHEBoAFR between its filled
BamHI
site and itsSall site.p294, obtained from Bill Sugden, is a 10.4-kbp, oriP-carrying plasmid derived from pHEBo that expresses the
EBNA1
gene from the immediate early promoter of human cytomegalovirus.Transient expression and replication assays. DNA was introduced into 6 x 106to 1 x
107
Raji, Wilson, or Wilson (p174) cells by electroporation with a ZA 1000 instrument (Promega) having a 0.5-cm-wide chamber of 0.5-ml volume at 1,500 V (3,000 V/cm) with 5pug
of DNA per ml of RPMI 1640 medium on ice. (Calculated resistance xcapacitance,
approximately 1 ms.)
MouseLMTK-cells were transfected with 1.5
jig
of CAT expression plasmid plus 5p,g
ofEBNA1
expression plasmid per 6-cm dish by using the calcium phosphate method (37) and were shocked 5 h later with 20% glycerol for 2min.After 24 h, the cells were washed with phosphate-buffered saline containing 1 mM EDTA, trypsinized, and split into two 10-cm dishes so that one dish could be harvested for CAT and the other for DNA, as described below. Cells were harvested after 72 h in culture, lysed by three freeze-thaw cycles, and assayed for CAT levels as previously described (12), with the following modifications. Reaction mixtures contained 10 nmol ofchloramphenicol
at a specific activity of5 mCi per mmol in a volume of 70,ul.
Acetylation rates were linear with time up to 7 nmol of acetylation (70%) or for at least 4 h and were linear with respect to added cell extract up to 100,ug
of protein. For experiments with Raji cells, 50 to 100,ug
of protein, measured by the Lowry method, was assayed for 25min
to 3 h, as appropriate for the CAT activities present.To measure transient plasmid replication, 3 x
107
to 6 x107
cells were harvested 4 to 5 days after electroporation and washed in phosphate-buffered saline containing 1 mM EDTA. The washing step removed over 95% of the unrepli-cated, DpnI-sensitive DNA that is associated with cells after electroporation or calcium phosphate-mediated transfection. Cells were suspended in 4 ml of buffer (20 mM Tris, 10 mM EDTA [pH 8.0]) and lysed by rapid addition of sodium dodecyl sulfate to a final concentration of 0.6%. Lysates were gently mixed with 1 ml of5 M NaCl, chilled at4°C
overnight, and centrifuged at 12,000 x g for 1 h. Low-molecular-mass DNA was isolated from the resulting Hirt supernatants (15) by deproteination with phenol followed by precipitation with isopropanol. After an additional phenol treatment and ethanol precipitation, DNA from approxi-mately
107
cells was digested with 5 U ofSalIplus 4 U ofDpnI
for 4 h. Samples were electrophoresed in0.7% agarose gels and then alkaline transferred (29) from the gels to Zetaprobe membranes (Bio-Rad Laboratories). Plasmid DNA was detected by hybridization with random-prime-labeled (9) pHyg (50 ng; 2 x107
to 5 x107
dpm) under the conditions of Church and Gilbert (4). pHyg is the precursor of pHEBo (Fig.1C) and lacks EBV sequences (34).Plasmid
maintenance assays. Three days after electropora-tion, Raji cells were plated onto 24-well culture dishes in medium containing 300,ug
of hygromycin B (Calbiochem-Behring) per ml. For each electroporation, cells were plated in 6 to 12 wells of 24-well culture dishes at densities of105,
104, 103,
or102 cells perwell. Cells werefed atleast onceper week by replacing part of the mediumin each well. Wells with proliferatingdrug-resistant cells were scored after3 to 4 weeks in culture. The frequency of outgrowth of drug-resistant cloneswas then calculated byassumingaPoisson distribution involving the fraction of negative wells at the dilution for eachthatgavesomepositive andsomenegative wells.DNAs from Hirt supernatants ofexpanded hygromycin B-resistant clones or pools of clones were analyzed on Southern blots of agarosegelsasdescribed above. Plasmids wererecoveredfromHirtsupernatantsbytransformation of Escherichia
coli
as described previously (37).RESULTS
The experiments weredesignedtodetermine
(i)
the activ-ities of the 30-bp repeats of oriP asfunctions of the number oftandem30-bpcopiesand(ii) whether tandemcopies
ofthe dyad symmetryregionoforiPcontaining similar numbersof
EBNA1-binding sites function similarly. As measures of 30-bp repeat activities, bothEBNA1-dependent
enhance-ment oftranscription and cis-complementation ofthedyad
symmetry region oforiP to allow EBNA1-dependent DNA replication weredetermined.
Enhancer activity as function of numberof
EBNAl-binding
sites. Deletions were introduced into the
family
of30-bp
repeatsbyusingthe plasmidpFR-TKCAT, which carriesall 20 copiesof30-bprepeats oforiPandthebacterial CATgene expressed from the rather weak TK promoter of
herpes
simplex virus. Expression of the CAT gene from the TK promoterisenhancedup to200-fold in
EBNAl-positive
cells by the presence of the 30-bp repeats onpFR-TKCAT
(30).
Thenumber of30-bp repeats
carrying
intactEBNA1-binding
sites onpFR-TKCAT was reducedfromthefull20
copies
to9, 7, 5, 2, and 0 copies (Fig. 1A and B).
Next,
thedyad
symmetry region of oriP, defined here as a
139-bp
DNA segment containing the 65-bpdyad
symmetry with its twoEBNAl-binding sites and the two additional
flanking
EBNAl-binding sites, was inserted into
pTKCAT
in one, two, or three tandem copies (Fig. 1A andB).
pTKCAT
is identical to pFR-TKCAT except that it lacks the30-bp
repeats. The enhancer activities of these constructs were
then compared by using
electroporation
to introduce the plasmids into Raji cells and bymeasuring
CAT levels 72 h later.BecauseColEl-relatedplasmids(such
aspBR322)
candimerize during propagation in Escherichia
coli
and the 30-bprepeatscan enhancetranscription
fromlong distances,
we took care that only plasmid
preparations
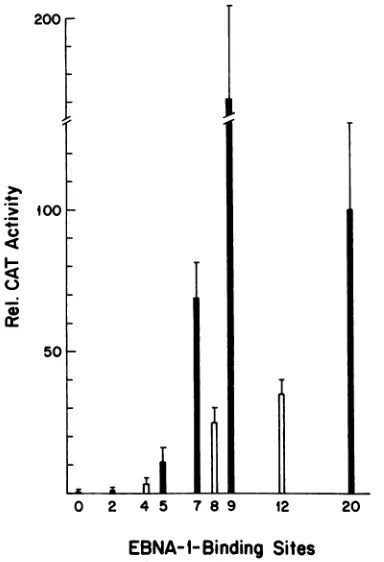
with less than 5%dimeric molecules were usedforenhancer assays.The relative CAT activities are shown in
Fig.
2. Nine intact 30-bp repeats werefully
active,
enhancing
CAT expression 70% more than didall 2030-bp
repeats, while 7 copies were two-thirds asactive asall 20. This isconsistent with the observation by Reismanand
Sugden
of fullen-hancer activity with 12 30-bp repeats
(30).
With five intact 30-bp repeats,activity
dropped
to11%;
two30-bp
repeats
gave only 1%of fullactivity,
raising
CATexpression
toonly
twice the unenhanced level. These results
suggest
that aminimum of six or seven
30-bp
repeats arerequired
for efficient enhanceractivityand thatactivity
drops
rapidly
asthe numberof30-bp repeats are reduced below this. Because there are sequence differences among
30-bp
repeats, we do not know whetherindividual
repeats
within thesegments tested contributeequally
to enhanceractivity.
It is conceivable, for
example,
that30-bp
repeats
6 and 7on November 10, 2019 by guest
http://jvi.asm.org/
200r
> too
O
50-0 2 4 5 7 8 9 12 20
EBNA-1-Binding
Sites
FIG. 2. Enhanceractivitywithvarious numbers of30-bprepeats
(closedbars) or copies of thedyad symmetry region(open bars).
Tests were performed in Raji cells as described in Materials and Methods. Mean CAT activities per microgram of extract were obtained fromfourelectroporations. Relative valuesareindicated
along with the number-corrected standard errors. The activity obtained withpFR-TKCAT,set at100%,wastypically2.5 nmol of
acetylationperhper100 ,ugofprotein.Theunenhanced CAT level
obtained with pTKCAT, typically 0.012 nmol/h per 100 p.g of
protein,wasthreetofivetimes thebackground valueobtained when an extract of mock-electroporated cells was tested. The values shownhavebeen corrected forbackground.
from the leftarealone responsible formostof theenhancer activity of the seven leftmost repeats rather than acting cooperatively with the other repeats. However, consider-ationof thesequencedifferencesamongtheserepeatsmakes this seem very unlikely. In Fig. 1A each 30-bp repeat is depicted as an open box representing the 18-bp symmetric
region containing the EBNA1 recognition sequence and a
closedboxrepresenting the 12-bpspacersequence. Repeats
1, 6,and 8containtheconsensussymmetric region (Fig. 1A,
a), while repeats 2, 4, 5, and 7 differ from the consensus sequenceinthisregion by only 1 bp (b and c).Thusrepeats
6 and 7are identical to repeats 3 and 4 exceptfora single
changeinthespacerofrepeat4(denoted by '). Similarly,we
do not knowwhether the EBNA1-binding sites in the dyad
symmetryregioncontribute equallytoenhanceractivity. We
note,however, thatmostofthenonlinearincrease in activity
was observed when going from four to eight binding sites from thedyad symmetryregion or whengoing from five to seven ornine 30-bprepeats,implyingthatsimilar numbersof
binding sites fromtworegionsarerequired for cooperativity.
While one copy of the dyad symmetry region had very
littleenhancer activity(30, 31)(Fig. 2),twoorthree tandem
copies exhibited significant activity, being 25 and 35% as
activeastheintactfamily of 30-bprepeats,respectively.The increase in enhancer activity from3.5%withone copyof the
TABLE 1.
EBNA1-dependent
enhancer activity in human andmousecellsCATactivityafor cell line:
LMTK- cotransfected Testplasmid Wilson Wilson(p174) with:
(EBNA1-) (EBNA1+) p392 p294
(EBNAl-) (EBNA1+)
pTKCAT 0.17 0.2 1.0 1.0
pFR-TKCAT 0.11 42 1.9 13
pTKCAT-3xDS 0.14 7.4 1.4 13
a Valuesareaveragesofduplicatedeterminations andareexpressedasthe percentageofchloramphenicol(total,10nmol)acetylated per hourof
reac-tion. Assayed were 100 pLg ofprotein extracted from approximately 10'
WilsonorWilson(p174)cellsand1.5,ug of protein extracted from approxi-mately 5 x 103LMTK- cells.
dyad symmetry region to 25% with two copies clearly exceededalinearincrease,asobserved for enhanceractivity
asa function of low numbers of30-bp repeats (Fig. 2). As with the family of 30-bp repeats (30, 31), the enhancer activity of three tandem copies of the dyad symmetryregion wasEBNA1dependent; itwasobserved inEBNAl-positive Wilson(p174) cells butnotinEBNA1-negative Wilson cells (Table 1).
As it will be shown below, tandem copies of the dyad symmetry region allow DNA replication in Raji cells. It is therefore possible that some ofthe increase in CAT gene expression observed withtwoorthree tandem copies of the dyadsymmetryregionwasduetoanincrease in the number oftranscriptionally active plasmids because ofreplication. We can roughly estimate the extent of replication if we
assume thatoriP-carrying plasmids replicatean average of once percelldivision cycle during the transient expression assay, asthey dowhentheyhave beenstably introducedby genetic selection. Recent experiments indicate that this is thecase with EBV-derived plasmids introducedintohuman 293 cells(J. L. Yates, unpublished results). Following elec-troporation, the surviving Rajicells appear tolagfor24 hand then to undergo two population doublings before being harvested 72 h after electroporation. If an oriP-carrying plasmid is assumed to replicate twice under these condi-tions, then pTKCAT carrying three copies of the dyad symmetryregion that replicates 40%asefficientlyasdoes a plasmid carrying oriP (see Fig. 4) would be expected to increase in number by less than twofold. In addition, as shownbelow, the enhanceractivity of tandemcopies of the dyad symmetry region or of the 30-bp repeats can be demonstrated by using rodent cell lines, which are nonper-missivefororiP-specificreplication.
In conclusion, a sufficient number of EBNA1-binding sites, 8 or 12, from the dyad symmetry region can act to enhance transcription, although not as efficiently in the human cell lines tested as a similar number of EBNA1-binding sites in thecontextofthe 30-bp repeats. Because the 30-bp repeats and the region of dyad symmetry have no common sequences other than the EBNA1-binding sites, and because these DNA segmentshave no enhancer activity in the absence of EBNA1, we conclude that multiple EBNA1-bindingsites,minimallysix or seven, are necessary and sufficient forenhancer activity.
EBNA1-dependent enhancer activity in cells nonpermissive forEBNA1-specific replication. Totesttheenhanceractivity of tandem copiesof thedyadsymmetryregionin the absence of DNA replication, we took advantage of the fact that
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.612.84.276.72.353.2]rodent cell lines are nonpermissive for oriP-specific replica-tion but support the EBNA1-dependentenhancer activity of the 30-bp repeats. pTKCAT, pFR-TKCAT, or
pTKCAT-3xDS
DNA was cotransfected into mouse LMTK- cells along with a plasmid that expresses EBNA1 from the cyto-megalovirus immediate early promoter or with a control plasmid lacking theEBNA1
gene. Three tandem copies of the dyad symmetry region enhanced CAT gene expression in anEBNA1-dependent
manner to the same extent as did the family of 30-bp repeats (Table 1).The cotransfected plasmids p294, which expresses the
EBNA1
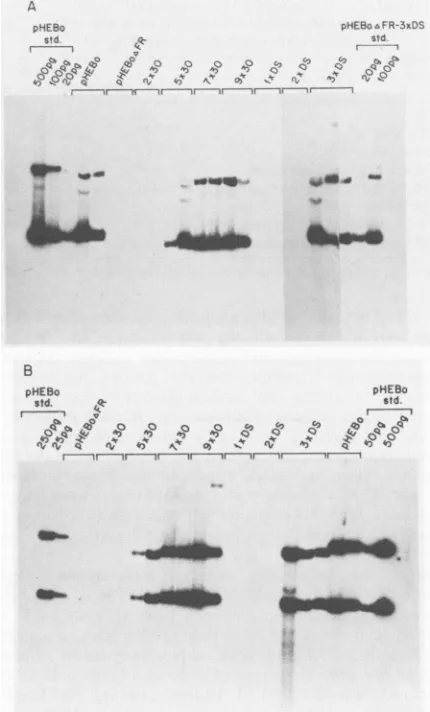
gene, and p392, which lacks the EBNA1 gene, both carry oriP. To test for replication of these DNAs in the transfected LMTK- cells, low-molecular-mass DNA was isolated 72 h after transfection froma portion of the LMTK-cells that had also been transfected with pTKCAT-3xDS. The DNA was digested withBamHIto linearize the plasmids and withDpnI
to degrade unreplicated DNA. (DpnI cuts only DNA containing methylated adenines on both strands of its recognition sequence, GATC. This modification is present on plasmids propagated in dam' E. coli; it is lost if the plasmids are replicated after being introduced into mam-malian cells.) Results of Southern analysis of the DNA are shown in Fig. 3. The same low levels of DpnI-resistant plasmids were detected whether the cotransfecting plasmid carried the EBNA1 gene (p294; Fig. 3, lanes 1 and 2) or lacked it (p392; lanes 3 and 4). In similar studies of transient replication with several human-derived cell lines, EBNA1 stimulated replication of oriP-carrying plasmids 10- to 50-fold (36; Yates, unpublished results). To verify that theDpnI cut DNA to completion, an E. coli-prepared (dam-methy-lated) plasmid DNA, pHEBo, intermediate in size between the two introduced plasmids, was mixed with a portion of the DNA isolated from the transfected cells and then digested and analyzed in parallel (Fig. 3, lane co.); noDpnI-resistant pHEBo DNA was detected. The low-level, apparently non-specific replication of transiently introduced plasmids is not peculiar to LMTK- cells; we have consistently observed it with each of several mammalian cell lines tested, including human 143 cells (36) and Raji cells (see below).We conclude that any EBNA1 oriP-specific replication that may occur in LMTK- cells is less efficient than the EBNA1-independent, apparently template-nonspecific con-version of input DNA to a DpnI-resistant form and is therefore undetectable. This is consistent with the inability of EBV-derived plasmids to be maintained as plasmids under selection in any of several tested cell lines derived from rodents (38). Yet theEBNA1-dependent enhancer activity of the 30-bp repeats is readily observed in LMTK- cells. Transactivation of the 30-bp repeats by EBNA1 and the inability of EBNA1 to activate oriP-specific replication in other mouse-derived cell lines has been independently ob-served (J. Mecsas and B. Sugden, personal communication). Multiple
EBNAI-binding
sites activate replication of dyad symmetry region. Both the 30-bp repeats and the dyad symmetry region oforiP must be present on the same DNA molecule to allow efficient EBNA1-dependent DNA replica-tion (19, 31). The shortened arrays of the 30-bprepeats were tested for the ability to cis-complement the dyad symmetry region of oriP by substituting them for the 30-bp repeats of theoriP-containing hygromycinB resistance vector pHEBo. The family of repeats was deleted from pHEBo, generating pHEBoAFR,to which was added the shortened 30-bprepeat arrays (Fig.1C). The tandem copies of the dyad symmetry region were also moved topHEBoAFRinorder totestthem for the ability to functionally replace the 30-bp repeatswithinFIG. 3. Resultsof assays for replication ofplasmids transiently introduced into LMTK-cells.Seventy-two hours aftertransfection, low-molecular-mass DNAs were isolated from portions (4 x 106 cells) of the transfected cells used in CAT expression assays, digested with BamHI (to linearizetheplasmids)and withDpnI, and electrophoresed on 0.7% agarose gels. An autoradiograph from a
Southern analysisisshown. Lanes 1 and 2, Samplesfromduplicate transfections with pTKCAT-3xDS plus p294 (EBNA1+); lanes 3 and 4. samples from duplicate transfections with pTKCAT-3xDS plus p392 (EBNAl-). Arrowheads mark the positions ofthe full-length, linear, DpnI-resistant plasmids: pTKCAT-3xDS, 4.7 kbp; p392, 8.4 kbp; and p294, 10.4 kbp. For lane co., 5 ng of dam-methylated pHEBo(7.2 kbp) wasmixed withequalportions ofDNA
from samples represented inlanes 1 and 2 andanalyzed inparallel. Some of the p294 DNA molecules carried a spontaneous, 700-bp deletion of the IR3 repeated sequences within the EBNA1 gene.
Such deletionshave nodeleteriouseffectsonEBNA1 functions(36,
38).
oriP. The plasmids were tested for the ability to replicate transiently following electroporation of Raji cells and to be maintained as plasmids in drug-selected cells.
To measure transient DNA replication, low-molecular-mass DNA was isolated from Raji cells 96 h
following
electroporation, digested with Salltolinearizethe
plasmids,
and analyzed for the presence ofDpnI-resistant molecules. Low-level replication of the vectorspTKCATandpHyg
wasobserved (Fig. 4B). No DpnI-resistant DNA could be de-tectedwhen dam-methylated plasmid DNAwasaddedtothe DNA extracted from mock-transfected cells
(Fig.
4,
lanes Co). TheoriP-carrying plasmid pHEBo replicated at a level 50 times more efficient than thebackground,
nonspecific
level. At most, pHEBoAFR carrying all of oriP except the 30-bp repeats replicated at a level twice that ofbackground (Fig. 4B).Replication activity as a function of the number of
30-bp
repeats washighly nonlinear(Fig. 4A). Twocopies of30-bp
repeats stimulated replication to onlytwice the
background
level, at most, while five 30-bp repeats supported transient replicationabouthalf asefficientlyasdid all 20copies.
Seven 30-bp repeats were fully effective. The steeprise inreplica-tion activity as afunction of the number of
30-bp
repeats is thus similar to the enhancer activity curve but is shifted toon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.380.478.77.316.2]A
A .:o c^0 t t
7
10 3.5 54 137 5-6 27 70 100
i--.
so
B
0,6
4 *)
l: q , ,
2.2 6.1 4.7 12 42 25 2.7 3.2 28 70
FIG. 4. Assays for transient replication of plasmids in Raji cells.
DNAs were isolated from cells 96 h (A) or120 h(B) after
electro-poration, digested with Salland Dpnl, and subjected to Southern
analysis. Hybridization signals representing Dpnl-resistant,
full-length plasmidswerequantified 'bylaserdensitometryof
autoradio-graphs exposed for appropriate periods without intensifying
screens. Relative values, averaged forduplicates, are indicated at
thetops of theblots,withpHEBogivenavalue of100inpanelAand
pHEBoAFR-3XDSgiven avalue of 70 inpanel B. FortwoDNAs,
duplicatetransfectionswerenotavailable foranalysis. For lanesCo.
1 ng ofaplasmid DNA was added to DNAextracted from
mock-electroporatedcells and analyzed inparallel.
fewer copies of30-bp repeats, with five copies supporting
justover50% ofreplication activity.Thereasonfor thismay
be that in the replication assay, a vectorcarrying the dyad
symmetry region (with its four EBNAl-binding sites) is by
necessity used in addition to the 30-bp repeats, and these distant sites are likely to cooperate.
Twoorthree tandemcopies of thedyadsymmetry region
activatedreplicationin theabsence of the30-bprepeats(Fig.
4). While addition ofonecopy of the
dyad
symmetryregion
topHEBoAFR
inplace
of the30-bp
repeats stimulatedreplication
to a levelonly
about twice that ofbackground,
addition oftwo orthree tandem
copies
of thedyad
symmetry regionincreased replication to 30 and70%,
respectively, of the level obtained with oriP intact. To determine whether tandemcopies
of thedyad
symmetryregion
wouldsupport
replicationin the absence of any other oriP sequences, these constructs were tested for thecapacity
toreplicate
when carriedby pTKCAT.
Two or three tandemcopies
of thedyad
symmetryregion supported replication
ofpTKCAT
about half as
efficiently
asthey supported replication
ofpHEBoAFR
whenplaced adjacent
to the remainder of oriP-960bp
ofintervening,
nonessentialDNA,
followedby
the
dyad
symmetryregion
in its normallocation.Multiple
copies
of thedyad
symmetryregion
actcooper-atively
topermit replication
rather thanacting independently
(Fig.
4). Ifindividualcopies
of thedyad
symmetryregion
actedindependently
when present on aplasmid
inmultiple
copies,
theprobability
of such aplasmid being replicated
would be
approximately
thesumofthe lowprobabilities
that eachdyad
symmetry copy,and thevectorsequences,would bereplicated.
Forexample,
withpHEBoAFR-3xDS,
which carries atotal of fourcopies
of thedyad
symmetryregion,
each copy and the vectorwouldcontribute about 0.025 and 0.02 unit ofreplication activity, respectively (relative tothe
valueof 1.0 forpHEBo),to
yield
avalue of0.12.Instead,
avalueof0.70 wasobserved
(Fig.
4).Theratherefficient
replication
obtained withtandem cop-ies of thedyad
symmetryregion
suggests that this140-bp
segment contains the actual
origin
ofreplication
of oriP. Activation ofthisorigin
appears torequire multiple (five
toseven)
EBNA1-binding
sites in cis, either from thedyad
symmetry
region
itself whenrepeated
tandemly
orfromthe30-bp
repeats. As with enhanceractivity,
this activation process isaccomplished
moreeffectively by
the30-bp
re-peatsthan
by
asimilar numberofEBNA1-binding
sites from thedyad
symmetryregion
(Fig. 4).
Numberof30-bprepeats
required
forplasmid maintenance;
inefficient functional substitution by tandem copies ofdyad symmetry region. The
oriP-carrying, hygromycin
Bresis-tance
plasmid
pHEBocanbeefficiently
introduced intoRaji
cells,
where it is carried under selection as astable,
multi-copyplasmid (34).
Removalof the30-bp
repeatsoforiPfrom pHEBo decreased thefrequency
at whichhygromycin
B-resistantRaji
clonescould be obtainedby
almost1,000-fold,
from 1 to 2% of
Raji
cells electroporated with pHEBo to0.005% of cells
electroporated
with pHEBoLFR, and pre-vented the plasmids from being maintainedextrachromo-somally (Table
2andFig.
5).Adding
back230-bprepeatsto pHEBoAFRdid notchangeitsproperties,whileaddingback7, 9,
orall 2030-bp
repeatsfully
restored efficientplasmid
maintenance. As with the tests for enhancer activity and transient replication, an intermediate result was obtained when five 30-bp repeats were tested for plasmid
mainte-nance. Although pHEBoAFR carrying five 30-bp repeats
wasabletoconferhygromycinBresistancetoRajicells with
only
1% of the normalefficiency,
thedrug-resistant
clones carried theplasmids
asextrachromosomal, circular, unrear-ranged molecules (Table 2 and Fig. 5). In two clonesanalyzed,
1 and 6 molecules ofpHEBol\FR-5
x30 were recovered per cell, compared with 12 to 20 copies per cell recoveredwithplasmids carrying fullyfunctionalconstructs oforiP(Table
2).Addingone or twotandem copies of the dyad symmetry
region
topHEBoAFR
hadnoapparenteffects.Threetandem'g,sc.,. "Y',,O 0 (i Ili CO
A..-Q * 4z) 0 'Q"
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.94.278.81.528.2]TABLE 2. Plasmidmaintenancein Raji cells under selection as a
functionof the number of 30-bp repeats
Frequencyof No. ofplasmid Plasmid hygromycin molecules recovered
resistance" percell"
pHEBo 1 x 10-2 20, 12
pHEBo/FR 4 x 10-5 0, 0
pHEBozFR-2x30 2 x 10-5 0, 0
pHEBoAFR-5x30 2 x 10-4 1, 6
pHEBoAFR-7x30 2 x 10-2 12, 15
pHEBoAFR-9x30 2 x 10-2 20, 12
pHEBoAFR-20x30 2 x 10-2 ND"
pHEBoAFR-lxDS 2 x 10-5 0, 0
pHEBoAFR-2xDS 4x 10-5 0, 0
pHEBoAFR-3xDS 3 x 10-4 20, 5, 7, 5
aFrequencies were determined by limiting dilution in hygromycin
B-containing medium as described in Materials and Methods. Values are
averaged frequenciesfor duplicateelectroporations,whichgenerally differed
fromeach other by afactorof 2 or less.
6Values wereobtainedby Southern analysisof DNA insupernatantsof
Hirtextracts(Fig.3) andrepresent individual hygromycin B-resistantclones, exceptforthelasttwo values forpHEBoAFR-3xDS, whichrepresentspools
of100to 200clones.Limit ofdetection:0.05molecule recoveredper cell. C ND, Notdone.
A
pHEBo
sthd 45
r3-3-0,0,cQ-,00Q0ve0- ;
B pHEEo
std. 45.
N 0
'5,0, e0 e 0
0 q k1- Ai A2a2'2oR Z
pHEBoAFR-3xDS
std
1- '' I ---I I
pHEBo std.
C, Co CoXe
'21 A21 q-lI~ O "
copies ofthe dyad symmetry region restored plasmid main-tenance to about 1% of normal efficiency, similar to the result obtained with five 30-bp repeats. Most hygromycin B-resistant clones obtained with this construct, pHEBoAFR-3xDS, carried fewer copies of the plasmid per cell than resultedwithpHEBo. However, one clone carried about 20 copies ofthe plasmidpercell (Table 2 and Fig. 5).
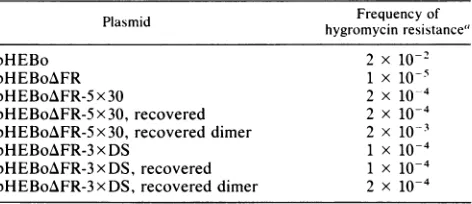
Properties of pHEBoAFR-5x30 and pHEBoAFR-3xDS maintained as plasmids in Raji cells. Because hygromycin-resistant clones are selected to carrypHEBoAFR-5x30and pHEBoAFR-3xDS at low frequencies, it seemed possible that genetically altered forms of these DNAs had been selected. To test this possibility, Hirt extracts from two hygromycin B-resistant clones carrying pHEBoAFR-5x30 and from four different populations carrying pHEBoAFR-3xDS were used to reintroduce the plasmids into E. coli. Mostlymonomericclones but alsoafewdimeric cloneswere obtained fromeach. In every case, analysiswith restriction enzymesthat would have revealed differencesas smallas30 bp in theoriPregion showed the plasmids tobe identicalto theoriginal constructs. Next, therecovered plasmids, both monomeric anddimeric, were testedfor efficiency ofstable introduction into Raji cells. The recoveredplasmids (mono-mers) were stably introduced into Raji cells at frequencies notsignificantlydifferent from thefrequencies obtainedwith the original plasmidDNAs and about 1% of thefrequency observed with pHEBo(Table 3).
The dimer of
pHEBoAFR-3
xDS did not produce signifi-cantly more hygromycin B-resistant clones than did the monomer, while the dimer ofpHEBo/FR-5x30 yielded 10 timesasmanydrug-resistant clonesasdiditsmonomer. We didnotdeterminewhether the increasedefficiency observed with thedimerofpHEBoAFR-5x30resultedfrom itsbeing
a dimerorfrom amutation in one of its monomericunits. In any event, thedimeric forms of theseplasmidswereaminor fraction of the plasmid molecules maintained in Raji cells selectedto carry the originalplasmids (Fig.
5A).
From these results we conclude that the stable mainte-nanceof these DNAsas replicating plasmidsdoesnot arise throughthe selectionofmutantforms. Itmightalsobenoted thatthefrequencywith whichthese
plasmids
are selectedtobe stably maintainedin Raji cells, 1%relativeto
pHEBo,
is..p mI_
FIG. 5. Southern analysis of Hirt supernatants of Raji cells selectedtocarry theindicatedplasmids.DNAobtainedfrom 3x 106 cells was analyzed uncut(A) or wascut with EcoRI (B). Forthe
standards,20 pgcorrespondsto oneplasmid molecule recoveredper
cell.ForuncutDNAs,the bandseentomigrateaheadoftherelaxed
circularform in the standards and samples represents the
super-coiled dimeric form. The band seen in one sample ofundigested
DNAfromcellscarrying pHEBoAFR-2x30islocatedattheposition ofundigested, residual cell DNA in the Hirt supernatants. DNA from oneclone carryingpHEBoAFR-9x30 contains alarger,
rear-rangedbutnonintegrated formof theplasmidatless thanonecopy percellinadditiontomultiplecopiesof theintact,originalplasmid.
PartofpanelAisfromablotofasimilargelbecauseofdistortionin thisregion oftheoriginalgel. SizesofEcoRIdigestionproductsare
4,500bp (pHEBo), 2,300bp(allplasmids),and 400bp(notseen).In
one sample of pHEBoAFR-3xDS, the partial digestion product (2,300plus400bp) ispresent.
more than 100-fold greater than theobserved mutation rate
of the TK gene carried on an oriP-based vectorand intro-duced intoan EBV-transformed
lymphoblastoid
cell lineby
electroporation (7).Given the rather efficient
replication
ofpHEBoAFR-5
x30 andpHEBoAFR-3xDS
in the first few celldivisions follow-ing electroporation, their lowefficiency
ofbeing
stably
maintained-1%comparedwithpHEBo-may
seemsurpris-ing. We suggest two
explanations. First,
efficientplasmid
maintenance of oriP-based vectors such as
pHEBo
under selection is likelytodepend
onthe enhanceractivity
ofthe 30-bp repeats in addition to the efficientreplication
of oriPon November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.54.295.94.220.2]TABLE 3. Frequencieswith whichpHEBozFR-5x30
andpHEBo/FR-3xDS isolatedfrom Rajicells
canbestably reintroduced intoRajicells
Plasmid Frequencyof
hygromycinresistance'
pHEBo 2 x 10-2
pHEBoA\FR 1 x 10-5
pHEBoAFR-5x30 2x 10-4
pHEBoLFR-5x30, recovered 2x 10-4
pHEBoAFR-5x30, recovered dimer 2x10-x
pHEBoAFR-3xDS 1 x 10-4
pHEBoAFR-3xDS, recovered 1 x 10-4 pHEBoAFR-3xDS,recovered dimer 2 x 10-4
"Averagevaluesfor duplicatedeterminations (see Table 1.footnote).
(19, 31). Thus the weak enhancer activities of 5X30 and 3XDS may lead to insufficient expression of the hph gene from the TK promoter and resultin lowcloning efficiencies inhygromycin B-containing medium. Second, the molecular requirements for transient replication may be less than what is required for plasmid maintenance. If each molecule of pHEBoAFRcarrying 5X30 or 3xDS were tocontinue rep-licatingindefinitely, witha50 to70%probability ofdoubling in each S phase, we would expectto obtain manycolonies that would grow slowly under selection and then lose the plasmid rapidly when grown without selection, as is ob-served with "weak" ars elements in Saccharomyces cere-v'isiae (14, 17). This was not observed.
Rather, we found that although Raji clones carrying pHEBoAFR-5x30 and pHEBoAFR-3xDS wereestablished at low frequencies, the plasmids were maintained in the selected clones rather effectively. The clones carrying pHEBoAFR-3xDS grew noticeably slower under selection thandid the clonescarrying pHEBo (doubling in 40to45 h compared with 26 to 35 h). Clones carrying pHEBoAFR-5x30 grew as well as did clones carrying pHEBo under selection, despite having fewer plasmid copies per cell. pHEBoAFR-3xDS was as stable as pHEBo in cells grown in theabsence of selection, being lost at the rate of 4 to 5% per generation. pHEBoAFR-5x30 was somewhat less stable, beinglostat 5to 10% per generation. It is thus conceivable thatestablishment of these two plasmids as stably replicating DNAswithadequate expressionof the hph gene involves an infrequent (1%) epigenetic event.
DISCUSSION
Two main conclusions emerged from the investigations into the functional differences between the two essential regions of oriP of EBV. First, because tandem copies of the dyad symmetry region replicate rather efficiently in the absence of any other EBV sequences in cis, this 139-bp DNA segment is very likely to contain the actual origin of DNAsynthesis. Mapping the origin by more direct, physical means is necessary to verify this assignment. Second, acti-vation of this postulated origin appears to require the pres-ence of multiple
EBNA1-binding
sites on the same DNA molecule, provided naturally by the family of 30-bp repeats oforiPorprovidedartificially and with less effectiveness by tandem copies of the dyad symmetry region itself. The resultsimply that a minimum of sevenEBNA1-binding
sites in as many 30-bp repeats (or possibly six, since six were not tested) are required to activateoriP
efficiently through a cooperative process.Because oriP contains 20 30-bp repeats with functional EBNA1-binding sites, at least in strain B95-8, whose
se-quence is known, it was somewhat surprising tofind that 7
30-bp
repeats support full oriPactivity
and 9 repeatsgive
maximal enhanceractivity. Interestingly, oriPof theclosely related virus herpesvirus papio (26) contains only 10 EBNA1-binding sites in itsfamily oftandem repeats and 4
EBNA1-binding
sites in itsdyad
symmetry region(J.
Pa-gano, D. Loeb,and R. Pesano, personal communication). Inaddition,
sequence differences among members of the EBV 30-bp repeats suggest that a duplication event of the 30-bp repeats(orageneconversion of 10membersbytheother10) occurred in the recent evolutionary pastofstrain B95-8 (1) (Fig. 1A).Enhancer functionofEBNA1 and30-bp repeats. Originally it was hypothesized that a role of the 30-bp repeats in plasmid maintenance could be that of relocalizing the plas-mid to the reforming nucleus at the end of mitosis or of targeting the plasmid to a nuclear subcompartment where DNA replication occurs (31). Although there is no real evidence for such a function of the 30-bp repeats, it is conceivable that the function exists and is at least part of what is measured as enhancer activity in the CAT gene transientexpressionassaysdescribed here. In any event, the 30-bp repeats constitute an EBNA1-dependent enhanceras defined operationally. Previous observations that the mea-sured enhanceractivity of the 30-bp repeats depends on the particular promoter and on the cell line tested argue for a rather direct effect ofthe30-bprepeatsontranscription (30, 31), since all cell lines used in the studies supported main-tenanceoforiP-carrying plasmids and therefore were ableto localize the introduced DNAs to the nucleus. In addition, the 30-bp repeats are likely to play an important role in the control of EBV gene expressionduring latency. A promoter located 2,200 bp from oriP which directs synthesis of the EBNA family of spliced mRNAs in some latently infected cell lines (2) has been shown todepend on EBNA1 and the 30-bp repeats for its activity (35). In the present studies, the demonstration of EBNA1-dependent enhancer activity by tandem copies of the dyad symmetry region implies that multiple EBNA-1 binding sites are necessary and sufficient for this activity, since the 30-bp repeats and the dyad symmetry region share no other sequence.
It is not known to what extent, if any, oriP replication depends on the 30-bp repeat enhancer function per se. However, the requirements for EBNA1-binding sites for enhancerfunction parallel the requirements for activation of replication: seven to nine 30-bp repeats are required for full activity; tandem copies of the dyad symmetry region en-hance transcription, but less effectively than the 30-bp repeats; and the nonlinear dependence of activity on the number of EBNA1-binding sites suggests cooperativity. These similarities suggest that transcriptional enhancement and activation of replication by the 30-bp repeats share certain steps. A recent deletion analysis of the EBNA1 gene supports this notion (36). Deletions within the EBNA1 open reading frame that did not alter DNA binding were found to reduce or eliminate enhancement and transient replication functions similarly or to affect neither, implying that the enhancer-activatingproperty of EBNA1 is an essential com-ponent ofreplication activation by EBNAl.
Host-specific factor required forEBNAI-dependent replica-tionbut not for activation of transcription. In light of several recent experiments demonstrating that activators of tran-scription can function in cells of the most distantly related eucaryotic organisms (27), the finding that EBNA1 can activate the 30-bp repeat enhancer in mouse cells is not surprising. The fact that EBV plasmid replication does not
on November 10, 2019 by guest
http://jvi.asm.org/
cross species lines from cells of primates to cells of rodents indicates that activation of DNA replication requires an activity ofEBNA1(or of oriP) in addition to what is needed for enhancer activation, such as an interaction with a host protein involved in DNA replication. The results suggest thatEBNA1 may interact with separate cell factorsinvolved in the control of transcription and of DNA replication.
Like EBV plasmid replication, simian virus 40 DNA replication occurs in monkey or human cells but not in rodent cells. The host range for simian virus 40 replication has been explained by the ability of simian virus 40 Tantigen to bind to DNA polymerase-alpha or an associated factor from human or monkey cells but not from mouse cells (23, 33). It should be possible to test for a comparableassociation of EBNA1 with DNA polymerase-alpha from permissive cells.
Cooperativity of EBNA1. The cooperative actions of EBNA1 molecules that lead to activation of transcription and DNA replication could occur, presumably, at the stage of DNA binding or subsequent to DNA binding. Recent studies in vitro withEBNA1 partially purified from Raji cells revealed very little cooperativity in the binding ofEBNA1 to the 30-bp repeats (16). Similarly, studies with the DNA-binding domain of EBNA1, a carboxyl-terminal fragment synthesized in E. coli, failed to show cooperativity of binding to adjacent sites, although binding to a single site was very concentration dependent, suggesting multimeriza-tion of theEBNA1 fragment (22). The studies of Jones et al. indicated that the 30-bp repeat sequence has an intrinsic affinity forEBNA1 that is severalfold higher than the affin-ities of the binding sites within the dyad symmetry region (16). Features of the 30-bp repeats that allow them to act cooperatively to enhance transcription and to activate repli-cation more efficiently than tandem copies of the dyad symmetry region could be this greater binding affinity for EBNA1, a more favorable spacing of the binding sites, or structural features that could impart DNA flexibility, such as the A+T-rich (85%) spacer that alternates with the 18-bp EBNA1 recognition sequence.
The lack of a high degree ofcooperativity in binding of EBNA1 to the 30-bp repeats in vitro leads us to speculate that the cooperativity of seven or more EBNA1 dimers observed for the functions of the 30-bp repeats in vivo reflects events that occur subsequent to DNA binding. Since EBNA1 is likely to activate DNA replication and transcrip-tion through an interactranscrip-tion with cellular factors, it ispossible that it is the binding of EBNA1 tothese factors, rather than to DNA, that is highly cooperative in cells. A requirement for cooperative action of multiple DNA-bound transcription-activating proteins in directing transcription initiation by polymeraseII has been inferred from several studies and is presumably an importantfeature in preventingsequestration of the transcription machinery by transcription activators bound at nonspecific sites (10, 25, 27).
It is also possible that a multi-EBNA1 protein complex, formed through binding to multiple DNA sites, isrequiredto destabilize the DNA double helix in leading to initiation of DNA replication, analogous to the actions of the multipro-tein complexes of E. coli dnaA protein at oriC and of bacteriophage AO protein at A ori (3, 6, 11, 32).
ACKNOWLEDGMENTS
We thank Sarah Camiolo for performing some of the plasmid
constructions, plasmid purifications, and CAT assays; Lynne Ma-quat, Tim Middleton, and Bill Sugdenforcomments on the
manu-script; several colleagues at Roswell Park Memorial Institute for
discussions and support; and Nancy Framefor preparingthe manu-script.
Thiswork was supported by aPublic HealthServicegrantfrom
the National Cancer Institute.
LITERATURECITED
1. Baer, R., A. T. Bankier, M. D. Biggin, P. L.Deininger, P. J.
Farrell, T. J.Gibson, G.Hatfull,G. S.Hudson,S. C.Satchwell,
C. Sequin, P. S. Tuffnell, and B. G. Barrell. 1984. DNA
se-quenceandexpression of theB95-8Epstein-Barr virusgenome. Nature(London) 310:207-211.
2. Bodescot, M., M. Perricaudet, and P. J. Farrell. 1987. A promoter for the highly spliced EBNA family of RNAs of Epstein-Barrvirus. J.Virol.61:3424-3430.
3. Bramhill, D., and A. Kornberg. 1988.DuplexopeningbydnaA
protein at novel sequences in initiation of replication at the
origin ofthe E. colichromosome. Cell52:743-755.
4. Church, G. M., and W. Gilbert. 1984. Genomic sequencing.
Proc. NatI. Acad. Sci. USA81:1991-1995.
5. DePamphilis, M. L. 1988. Transcriptional elements as compo-nentsofeucaryoticorigin ofDNAreplication.Cell52:635-638.
6. Dodson, M., J. Roberts, R. McMacken, and H. Echols. 1985.
Specializednucleoproteinstructures attheoriginofreplication ofbacteriophage X: complexes with AO, XP, andEscherichia coliDnaBproteins. Proc. Natl. Acad. Sci. USA 82:4678-4682.
7. Drinkwater, N. R., and D. K. Klinedinst. 1986. Chemically
inducedmutagenesisinashuttlevectorwithalow-background
mutantfrequency. Proc.Natl. Acad. Sci. USA 83:3402-3406. 8. Echols, H. 1986. Multiple DNA-protein interactionsgoverning
high-precisionDNA transactions. Science233:1050-1056.
9. Feinberg, A. P., and' B. Vogelstein. 1983. A technique for
radiolabeling DNA restriction endonuclease fragments to high
specificactivity. Anal. Biochem. 132:6-13.
10. Fromental, C., K. Masamoto, H.Nonayama, and P.Chambon. 1988. Cooperativity and hierarchical levelsoffunctional orga-nization in theSV40enhancer. Cell 54:943-953.
11. Fuller, R. S., B. E.Funnell, and A.Kornberg. 1984. The dnaA
proteincomplexwith the E.coli chromosomalreplicationorigin (oriC)and othersites. Cell38:889-900.
12. Gorman, C. M., G. T. Merlino, M. C. Willingham, I. Pastan,
and B.H.Howard. 1982.The Rous sarcomaviruslongterminal repeat is a strong promoter when introduced into a
variety
of eukaryotic cells by DNA-mediated transfection. Proc. Natl.Acad. Sci. USA79:6777-6781.
13. Hampar,B., A. Tanaka, M.Nonayama, andJ.G.Derge. 1974.
Replicationof the residentrepressedEpstein-Barrvirus genome
duringtheearly Sphase(S-1 period) ofnonproducer
Raji
cells. Proc. Natl. Acad. Sci. USA71:631-635.14. Heiter,P., C.Mann,M.Snyder,and R. W. Davis. 1985. Mitotic
stability of yeast chromosomes: a colony color assay that measures nondisjunction and chromosome loss. Cell 40:381-392.
15. Hirt, B. 1967. Selective extraction of polyoma DNA from
infected mousecell cultures.J. Mol. Biol. 26:365-369. 16. Jones, C. H., S. D.Hayward, and D. R. Rawlins. 1989.
Interac-tion oflymphocyte-derived Epstein-Barr virus nuclear
antigen
(EBNA-1) with its DNA-binding sites. J. Virol. 63:101-110. 17. Koshland, D., J. C. Kernt, and L. A. Hartwell. 1985. Geneticanalysisof the mitotic transmission of minichromosomes. Cell 40:393-403.
18. Lindahl, T., A. Adams, G. Bjursell, G. W. Bornkamm, C.
Kaschaka-Dierich,and U.Jehn.1976. Covalentlyclosed circular
duplex DNA of Epstein-Barr virus in a human
lymphoid
cell line.J. Mol. Biol. 102:511-530.19. Lupton, S.,and A.J.Levine.1985. Mapping
genetic
elementsof Epstein-Barrvirus thatfacilitate extrachromosomalpersistence
of Epstein-Barr virus derived plasmids in human cells. Mol. Cell. Biol. 5:2533-2542.
20. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecular
cloning:alaboratory manual. Cold Spring Harbor Laboratory,
ColdSpring Harbor, N.Y.
21. Mecsas, J., and B. Sugden. 1987.
Replication
ofplasmids
de-rived from bovinepapilloma virustype1andEpstein-Barr
viruson November 10, 2019 by guest
http://jvi.asm.org/
in cells inculture. Annu. Rev. CellBiol. 3:87-108.
22. Milman,G., and E.S.Hwang. 1987. Epstein-Barrvirus nuclear
antigen forms a complex that binds with high concentration
dependence toasingle DNA-bindingsite. J. Virol. 61:465-471.
23. Murakami, Y., C. R. Wobbe, L.Wiessbach,F. B.Dean, and J.
Hurwitz. 1986. Role of DNA polymerase-xandDNAprimasein
simian virus 40 DNA replicationiniitro. Proc.Natl. Acad. Sci. USA74:1605-1609.
24. Nonayama, M., and J.S. Pagano. 1972. Separation of Epstein-Barr virus DNA from large chromosomal DNA in non-virus-producing cells. Nature(London) New Biol. 238:169-171. 25. Ondek, B., L. Gloss, and W. Herr. 1988. The SV40 enhancer
contains two distinct levels of organization. Nature (London) 333:41-45.
26. Pesano, R. L., and J. S. Pagano. 1986. Herpesvirus papio containsaplasmid origin of replication thatactsincis interspe-cies withanEpstein-Barr virus tlrns-acting function. J. Virol.
3:1159-1162.
27. Ptashne, M. 1988. How eukaryotic transcriptional activators work. Nature (London) 335:683-689.
28. Rawlins, D., G. Milman, S. D. Hayward, and G. S. Hayward.
1985.Sequence specific DNA binding of the Epstein-Barr virus nuclearantigen (EBNA) toclustered sites in the plasmid
main-tenance region. Cell 42:859-868.
29. Reed,K. C., and D. A. Mann.1985.Rapid transfer ofDNAfrom agarose gels to nylon membranes. Nucleic Acids Res. 13: 7207-7221.
30. Reisman, D., and B. Sugden. 1986. tranls-Activation of an
Epstein-Barr viraltranscriptional enhancer by the Epstein-Barr
viral nuclearantigen 1. Mol. Cell. Biol. 6:3838-3846.
31. Reisman, D.,J.L.Yates, and B. Sugden. 1985. A putative origin ofreplication of plasmids derived from Epstein-Barr virus is composed of two cis-acting components. Mol. Cell. Biol. 5: 1822-1832.
32. Schnos, M., K. Zahn, R. B. Inman, and F. R. Blattner. 1988. Initiation protein induced helixdestabilizationattheXorigin:a
prepriming step in DNA replication. Cell 52:385-395.
33. Smale, S. T., and R. Tjian. 1986. T-antigen-DNA polymerase-alpha complex implicated in simian virus 40 DNA replication. Mol. Cell. Biol. 6:4077-4087.
34. Sugden, B., K. Marsh, and J. L. Yates. 1985. A vector that replicates as a plasmid and can be efficiently selected in B-lymphoblasts transformed by Epstein-Barr virus. Mol. Cell. Biol. 5:410-413.
35. Sugden, B., and N. Warren. 1989. A promoter ofEpstein-Barr virus which can function during latent infection can be
Ir-ans-activatedby EBNA1,aviralproteinrequired forDNA replica-tion duringlatentinfection.J. Virol. 63:2644-2649.
36. Yates, J. L., and S. M. Camiolo. 1988. Dissection of DNA
replication and enhancer activation function of Epstein-Barr virus nuclear antigen 1. Cancer Cells 6:197-205.
37. Yates, J. L., N. Warren, D. Reisman, and B. Sugden. 1984. A
cis-acting element from the Epstein-Barr viral genome that
permits stable replication of recombinant plasmids in latently infected cells. Proc. Natl. Acad. Sci. USA 81:3806-3810. 38. Yates, J. L., N. Warren, and B. Sugden. 1985. Stable replication
ofplasmidsderived fromEpstein-Barr virus in various mamma-lian cells. Nature (London) 313:812-815.