0022-538X/87/051398-09$02.00/0
CopyrightC 1987, AmericanSocietyforMicrobiology
Nucleotide
Sequence
and
Transcript Organization
of
a
Region of the
Vaccinia Virus Genome Which Encodes
a
Constitutively Expressed
Gene
Required for DNA
Replicationt
NANCY A. ROSEMAN ANDDENNIS E. HRUBY*
CenterforGene Research, Department of Microbiology, OregonState University, Corvallis, Oregon 97331-3804 Received17November1986/Accepted30January 1987
Avaccinia virus (VV) generequiredfor DNAreplicationhasbeenmappedtothe left side of the 16-kilobase (kb) WHindIII D DNAfragment by marker rescue of a DNA-
temperature-sensitive
mutant, tsU7, using clonedfragments of the viral genome. Theregionof W DNAcontainingthetsl7 locus(3.6 kb)wassequenced. Thisnucleotide sequence contains onecompleteopenreadingframe(ORF)
andtwoincompleteORFsreading from left to right. Analysis of this region at early times revealed thattranscription
from the incomplete upstream ORF terminatescoincidentally with the complete ORFencoding thets17 gene product, which is directly downstream. Thepredictedproteinsencodedbythisregioncorrelate well with polypeptidesmapped by in vitro translation ofhybrid-selected earlymRNA. Thenucleotide sequences ofa 1.3-kbBglll fragment derivedfrom ts17 and from twots17revertants werealsodetermined,and thenatureof thetsU7 mutation was identified.S1 nucleaseprotection studies were carried out to determine the 5' and 3' ends of thetranscriptsand to examine the kinetics ofexpression of the ts17 geneduring viralinfection. Thets17transcriptis present at bothearly and late times postinfection, indicating thatthisgene isconstitutively expressed. Surprisingly, the transcriptional start throughout infection occurs at the proposed late regulatory element TAA, which immediately precedes theputative initiation codon ATG. Althoughthe biological activityof the tsl7-encoded polypeptide was not identified, it was noted that in tsl7-infected cells, expression of a nonlinked VV immediate-early gene(thymidine kinase) was deregulated atthenonpermissivetemperature. This result may indicate that thets17gene product isfunctionally requiredat anearlystep oftheWreplicative cycle.Vaccinia virus (VV) is a large, DNA-containing animal virus which replicates in the cytoplasm of susceptible host cells. VV is an attractive system for the study of gene regulation in higher organisms owing to the temporally coordinatedexpression of itsapproximately 200 genes (25). At the onset ofinfection, immediate-early viral RNAs are transcribed by the virally encoded and packaged RNA polymerase (26). Aprotein product of this temporal class is thenrequiredfor the expression of the delayed-earlygenes, astheirexpression is blocked by protein synthesis inhibitors such as cycloheximide. Approximately 2 h postinfection (p.i.), viral DNA replication is initiated. Concomitant with DNA replication is repression of the expression of most early genes and the initiation of late gene transcription. Thus, theexpression ofmost VV genes can be classified as immediate early, delayed early, or late. However, there is alsoanadditional constitutivetemporal class of genes which appear to be expressed at both early and late times during infection astypified by the well-studied 7.5-kilodalton gene (5)andother VV genes (41).
The precise mechanisms involved in the modulation of VV geneexpression are unknown, although they are believed to operate at the transcriptional, posttranscriptional, transla-tional, and posttranslational levels. Through the identifica-tionand analysis of a number of genes from specific temporal classes it should be possible to identify the regulatory elements involvedin VV gene expression. Sequence
analy-*Corresponding author.
tOregon Agricultural Experiment Station Technical Paper no. 8050.
sesof various regions oftheviralgenome(2, 11,15, 27, 28, 33, 38, 39, 40, 43, 44) have identified many open reading frames (ORFs) whichcorrelate well with earlier transcrip-tionalandtranslationalmapping studies (1, 3, 4). Upstream cis-acting regulatorysequences have been identified for both earlyand late genes, buttheyappear to havelittle obvious homology topreviously identified procaryoticoreucaryotic promoters (2, 10, 19, 37,44).
Inthisstudywehavemappedaviralgenebelongingto the constitutive temporalclassby marker rescue of a tempera-ture-sensitive(ts)DNA- mutant, tsl7(6, 7). The tsl7 locus was mapped within theHindIlI Dfragment, and the nucle-otidesequenceof thisregionwasdeterminedfrom wild-type (wt)viralDNA. Whilethis work was in progress the nucle-otide sequence of the entire HindIll D fragment was re-ported (27; nomenclature for ORFs is in relation to the reported sequence and accordingto the recently proposed system for VV genes [32]). Here, weindependentlyconfirm aportion ofthereportedsequence and show that this region istranscriptionallyactive in vivo. Comparison of sequences derivedfromDNA isolated from tsl7 virus, from two tsl7 revertants
(tsl7r),
andfrom wt virus identified the mutation responsible for the temperature sensitivity of tsl7. Also, analysisof the sequence inconjunction with transcriptional andtranslationalstudiesof this region showed that transcrip-tionofD4, which lies directlyupstream of D5 (to whichtsl7 maps), terminatescoincidentally with the D5 transcript.Thetranscriptionalkinetics of thetsl7gene was examined by S1 nuclease protection studies of steady-state mRNA levels. The results showed that thetsl7gene is a member of theconstitutive temporal class of viral genes. The putative 1398
on November 10, 2019 by guest
http://jvi.asm.org/
VACCINIA VIRUS GENE REQUIRED FOR DNA REPLICATION 1399
C NMK F E 0 1 G L J H D A B
16Kb
LD
At
cE D B- A G4+F4I4H4 C
,~~~~~~~~~~~~~%
%B A' a C
F
IKb
lKb
I
lKb
FIG. 1. Diagrammatic representation of the HindIll restrictionmapof the VVgenome.Restriction fragments generated by digestion of
the 16-kilobase-pairHindIIIDfragment withBamHI (l)andEcoRI (t) areshown. Restriction fragments generated by digestion of the 3.3-kbEcoRI B fragmentbyBglII (A)arealso shown.The 7.0-kbfragment designated clone pSW1 is shown(v).Those framents used in the markerrescuestudyareindicated inthetextandin Table 1. Fragments abletorescuetsl7totemperatureinsensitivity areindicated (*).
promoter of the tsl7 gene was compared with those of
previously reported VV genes.
MATERIALS ANDMETHODS
Cells and virus. BSC40 cells were grown in monolayer
culturesof Eagle minimumessential medium (Flow Labora-tories) supplemented with 10% heat-inactivated fetal calf
serum, 2 mM L-glutamine, and 50 ,ug of gentamicin sulfate
perml. VV(strain WR) is designated aswtin these experi-ments. Revertants of tsl7 weregeneratedby repeated
pas-sagesof tsl7 virusat40°C. tsl7revertantswerethen plaque
purifiedat40°C.
Marker rescue. Cloned wt VV DNA fragments were
purified and sterilized by sequential extractions with phenol andether, followed by ethanol precipitation. From1to2 ,ug ofviral DNAwassuspended in HEPES
(N-2-hydroxyethyl-piperazine-N'-2-ethanesulfonic acid)-buffered saline and coprecipitated with salmonspermDNAtoafinal concentra-tion of 20 ,ug/ml using calcium phosphate (9). Confluent 60-mm plates of BSC40 cells were infected with tsl7 at a
multiplicity of infection of 0.1 and incubatedat31°C for 3 h. Themediumwasremoved, the monolayerwaswashed with
Eagle minimum essential medium, and 15% glycerol was
addedfor 40s.Themonolayerwas then washed twice with
Eagle minimum essential medium, and theprecipitated DNA
waspipettedontothecells. Plateswereincubatedat40°C for 3 hand then washed, and minimum essential medium plus 10%fetal calfserum wasadded. Plateswereincubated for 48
h at 40°C, the infected cells were harvested, and titers of
infectious virus weredeterminedat31 and40°C.
Cloning and sequencing.Aclone of theHindIll Dfragment ofVVDNAwasoriginallyobtainedfrom B.Moss(National
Institutes of Health). Subfragments of viral DNA were
clonedintopUCvectorsby standard methodology(22). M13 phage vectors were used for sequencing by the dideoxy-nucleotide chain termination method (34). Sequences were
analyzed on an IBM personal computer using Microgenie software obtained from Beckman Instruments, Inc. (29). Chemical sequencing was performed essentially as
previ-ously described (23). Enzymes and reagents were obtained
from Bethesda Research Laboratories, Boehringer Mann-heim, P-L Biochemicals, and Aldrich Chemical Co., Inc. Radioisotopes were obtained from New England Nuclear
Corp.
Northern analysis. Two micrograms of polyadenylated viral RNA (isolated essentially asdescribed [42]) was elec-trophoresedinaformaldehyde-agarose gelasdescribed(18).
After transfer to nitrocellulose, specific transcripts were
detected by using the indicated nick-translated viral DNA fragments asprobes.
Hybrid selection. A 20-,ug sample of recombinant plasmid DNA wasdigestedtocompletion withanappropriate restric-tion endonuclease and bound to five-3-mm-square nitrocel-lulose filters. Twenty micrograms ofpolyadenylated RNA isolated from VV-infected cellsin the presence of 100 ,ugof cycloheximide per ml was usedin hybridizationreactions as described (1). Eluted mRNA was translatedin vitro inrabbit reticulocyte lysates which contained 5 ,uCi of
L-[35S]methi-onine (NewEngland Nuclear Corp.; 1,085 Ci/mmol)(14).
Si
nucleasemapping. 5'- or3'-end-labeledDNAfragments were mixed with20 ,ugofviralRNA in a total volume of 30 ,lI containing 80% formamide-40 mM PIPES [piperazine-N,N'-bis(2-ethanesulfonic acid)](pH6.4)-0.4MNaCl-1mM EDTA. Thesampleswereheatedto90°Cfor5minandthen incubatedat42°C for4h. Hybridization reactionswerethen plungedintoice, and 0.3 mlof ice-cold0.28 MNaCl-0.05M sodium acetate(pH
4.6)-4.5 mMZnSO4-400
U of nuclease S1 per ml wasadded. After 5 min onice,the sampleswere incubated at25°C for 1 h and then extracted with phenol-chloroform and ether and ethanol precipitated. Protected fragmentswere analyzed on sequencing gels.Thymidine kinase assay.Duplicatesetsof60-mmdishes of Ltk- cellswereinfectedat amultiplicity of20PFUpercell with wt VV or the indicated ts mutants. After a 30-min adsorption at 25°C, prewarmed (31 or 40°C) medium was
added. One set of plates was incubated at the permissive temperature of 31°C, and the other was kept at the
nonpermissive
temperature of40°C. At 2-h intervals, cyto-plasmic extracts wereprepared
and frozen. Extracts weresubsequently
assayed forthymidine
kinaseactivity
(13).TABLE 1. Titers of progeny of markerrescuebysubclonesof VVHindIlI fragmentDI
Titerattemp:
ViralDNAfragment
31°C 40°C
HindIlI D 1.7 x 105 1.9 x 105
pSW1 1.2 x 103 0
BamHIA 6.5 x 104 1.2 x 105
EcoRIB 2.6 x 105 2.8 x 105
BglIIA 8.0 x 104 1.6x 104
BgIII
B 8.3 x 103 0BgIIIC 2.4 x 104 0
SS 4.2 x 103 0
aViral DNAwas coprecipitatedwithsalmon sperm DNA(SS)andthen
transfected intotsl7-infectedBSC40cells.
VOL. 61,1987
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.612.62.557.66.183.2] [image:2.612.319.560.602.706.2]AsniSerValThrValSerHisAlaProTyrTlhrIleThrTyrHiisAspAspTrpGluProValMetSerGflnLeuVa1GluPheTyrAsnGluValAlaSerTrpLeuLeuArgAspGl
240
GACGTCGCCTATTCCTGATAAGTTCTTTATACAGTTGAAACAACCGCTTAGAAATAAACGAGTATGTGTGTGCGGTATAGATCCGTATCCGAAAGATGGAACTGGTGTACCGTTCGMATC
uThrSerProIleProAspLysPhePheI1eGlnLeuLysGlnProLeuArgAsnLysArgValCysValCysGlyIleAspProTyrProLysAspGlyThrGlyVal ProPheGluSe 360
ACCAAATTTTACAAAAAAATCAATTAAGGAGATAGCTTCATCTATATCTAGATTAACCGGAGTAATTGATTATMAAGGTTATAACCTTAATATAATAGACGGGGTTATACCCTGGAATTA
rProAsnPheThrLysLysSerIleLysGluIleAlaSe rSer IleSerArgLeuThrGlyValIleAspTyrLysGlyTyrAsnLeuAsnIlelIleAspGlyVal leProTrpAsnTy 480
TTACTTAAGTTGTAAATTAGGAGAAACAAAAAGTCACGCGATCTACTGGGATAAGATTTCCAAGTTACTGCTGCAGCATATAACTAAACACGTTAGTGTTCTTTATTGTTTGGGTAAAAC
rTyrLeuSe rCys LysLeuGlyG luThrLysSerHisAlaIleTyrTrpAspLysIleSerLysLeuLeuLeuGlnliisIleThrLysHlisValSerValLeuTyrCysLeuGlyLysTh 600
AGATTACTCGAATATACGGGCCAAGTTAGAATCCCCGGTAACTACCATAGTCGGATATCATCCAGCGGCTAGAGACCGCCAATTCAGAGMMGATAGATCATTTGAAATTATCAACGTTTT
rAspTyrSerAsnIleArgAlaLysLeuGluSerProValThrThr IleValGlyTyrliisProAlaAlaArgAspArgGlnPheGluLysAspArgSerPheGluI leIleAsnValLe
III1 720
ACTGGMATTAGACAACAAGGCACCTATAAATTGGGCTCAAGGGTTTATTTATTAATGCTTTAGTGAAATTTTMACTTGTGTTCTAAATGGATGCGGCTATTAGAGGTAATGATGTTATCT
uLeuGluLeuAspAsnLysAlaProIleAsnTrpAlaGlnGlyPheIleTyrEnd MetAspAlaAlaIleArgGlyAsnAspValIleP
840
TTGTTCTTAAGACTATAGGTGTCCCGTCAGCGTGCAGACAAATGAAGATCCAAGATTTGTAGAAGCATTTAAATGCGACGAGTTAGAAAGATATATTGAGAATAATCCAGAATGTACAC
heValLeuLysThrIleGlyValProSerAlaCysArgGlnAsnGluAspProArgPheVa1GluAlaPheLysCysAspGluLeuGluArgTyrIleGluAsnAsnProGluCysThrL
960
TATTCGAAAGTCTTAGGGATGAGGAAGCATACTCTATAGTCAGAATTTTCATGGATGTAGATTTAGACGCGTGTCTAGACGAAATAGATTATTTAACGGCTATTCAAGATTTTATTATCG
euPheGluSerLeuArgAspGluG luAlaTyrSerIleValArgIle PheMetAspValAspLeuAspAlaCysLeuAspGluIleAspTyrLeuThrAlaIleGlnAspPheIle IleG 1080
AGGTGTCAAACTGTGTACCTAGATTCGCGTTTACAGAATGCGGCGCCATTCATGAAAATGTAATAAAATCCATGAGATCTAATTTTTCATTGACTAAGTCTACAAATAGAGATAAACAA
luValSerAsnCysValAlaArgPheAlaPheThrG luCysG lyAlaIleH isGluAsnVal1IleLysSerMetArgSerAsnPheSerLeuThrLysSerThrAsnArgAspLysThrS
* * 1200
GTTTTCATATTATCTTTTTAGACACGTATACCACTATGGATACATTGATAGCTATGAAACGAACACTATTAGAATTAAGTAGATCATCTGAAATCCACTAACAAGATCGATAGACACTG
erPheHisIleIlePheLeuAspThrTyrThrThrMe tAspThrLeuIleAl aMetLysArgTlirLeuLeuGluLeuSerArgSerSe rGluAsnProLeuThrArgSerIleAspThrA 1320
CCGTATATAGGAGAAAACAACTCTTCGGGTTGTAGGTACTAGGAAAATCCAAATTGCGACACTATTCATGTAATGCAACCACCGCATGATAATATAGAAGATTACCTATTCACTTACG
laValTyrArgArgLysThrThrLeuArgValValG lyThrArgLysAsnProAsnCysAspThrIleHisValMetGlnProProHlisAspAsnIleGluAspTyrLeuPheThrTyrV 1440
TGGATATGAACAACAATAGTTATTACTTTTCTCTACAACAACGATTGGAGGATTTAGTTCCTGATAAGTTATGGGAACCAGGGTTTATTTCATTCGAAGACGCTATAAAAGAGTTTCAA
alAspMetAsnAsnAsnSerTyrTyrPheSerLeuGlnGlnArgLeuGluAspLeuVal1ProAspLys LeuTrpGluProGlyPheIle SerPheGluAspAlaIleLysArgValSerL
1560
AAATATTCATTAATTCTATAATAAACTTTAATGATCTCGATGAAAATAATTTTACAACGGTACCACTGGTCATAGATTACGTAACACCTTGTGCATTATGTMAAAAACGATCGCATAAAC
ys Ile Phe IleAsnSerIleIleAsnPheAsnAspLeuAspG luAsnAsnPheThrThrVal1ProLeuVal IleAspTyrValThrProCysAlaLeuCys LysLysArgSerHisLysH 1680 ATCCGCATCAACTATCGTTGGAAAMTGGTGCTATTAGAATTTACAAAACTGGTMATCCACATAGTTGTAAAGTTAAAATTGTTCCGTTGGATGGTAATAAMCTGTTTAATATTGCACMAA i sProH isG lnLeuSe rLeuGluAsnGlyAlalIleArgI leTy rLysThrG lyAsnProlliisS erCysLysVal1LyslIleVal1ProLeuAspGlyAsnLysLeuPheAsnIleAlaG lnA
1800 GAATTTTAGACACTAACTCTGTTTTATTAACCCAAGAGGAGACCATATAGTTTGGATTAATAATTCATGGAAATTTAACAGCGAAGAACCCTTGATAACAAAACTAATTTTGTCAATAA rgIleLeuAspThrAsnSerVal1LeuLeuThrGluArgG lyAspHis Il1eValTrpIleAsnAsnSerTrpLysPheAsnSerG luGluProLeuIleThrLysLeuIleLeuSerlIleA 1920
GACATCAACTACCTAAGGAATATTCAAGCGAATTACTCTGTCCAAGAAAACGAAGACTGTAGAAGCTAACATACGAGACATGTTAGTAGATTCAGTAGAGACCGATACCTATCCGGATA
rgHIisGlnLeuProLysGluTyrSerSerGluLeuLeuCysProArgLysArgLysThrValGluAlaAsnIleArgAspMetLeuValAspSerValGluThrAspThrTyrProAspL 2040
AACTTCCGTTTAAAATGGTGTATTGGACCTGGTAGACGGAATGTTTTACTCTGGAGATGATGCTMAAAAATATACGTGTACTGTATCAACCGGATTTAAATTTGACGATACAAAGTTCG
ysLeuProPheLysAsnGlyValLeuAspLeuValAspGlyMetPheTyrSerGlyAspAspAlaLysLysTyrThrCysThrValSerThrGlyPheLysPheAspAspThrLysPheV
2160 TCGAAGACAGTCCAGAAATGGAAGAGTTAATGAATATCATTAACGATATCCAACCATTAACGGATGAAAATAAGAAAMTAGAGAGCTATATGMAAAAACATTATCTAGTTGTTTATGCG
alGluAspSerProGluMetGluGluLeuMetAsnIlel leAsnAspIleGlnProLeuThrAspGluAsnLysLysAsnArgGluLeuTyrCluLysThrLeuSerSerCysLeuCysG 2280 GTGCTACCAAAGGATGTTTAACATTCTTTTTTGGAGAAACTGCAACTGGAAAGTCGACAACCAAACGTTTGTTAAAGTCTGCTATCGGTGACCTGTTTGTTGAGACGGGTCAAACAATTT
lyAlaThrLysGlyCysLeuThrPhePhePheGlyGluThrAlaThrGlyLysSerThrThrLysArgLeuLeuLysSerAlaIleGlyAspLeuPheValGluThrGlyGlnThrIleL
2400 TAACAGATGTATTGGATAAAGGACCTAATCCATTTATCGCTAACATGCATTTGMAAAAATCTGTATTCTGTAGCGAACTACCTGATTTTGCCTGTAGTGCATCAAAGAAATTAGATCTG
euThrAspValLeuAspLysGlyProAsnProPheIleAlaAsnMetHisLeuLysArgSerValPheCysSerGluLeuProAspPheAlaCysSerGlySerLysLysIleArgSerA
2520 ACAATATTAAAAGTTGACACAACCTTGTGTCATTGGAAGACCGTGTTTCTCCAATAAAATTAATAATAGAAACCATGCGACAATCATTATCGATACTAATTACAAACCTGTTTTTGATA
spAsnIleLysLysLeuThrGluProCysValIleGlyArgProCysPheSerAsnLys IleAsnAsnArgAsnHisAlaThrI le IleIleAspThrAsnTyrLysProValPheAspA 2640
GGATAGATAACGCATTAATGAGAAGAATTGCCGTCGTGCGATTCAGMAcACACTTTTCTCAACCTTCTGGTAGAGAGGCTGCTGAAATAATGACGCGTACGATAAAGTCAAACTATTAG rgIleAspAsnAlaLeuMetArgArgI leAlaValValArgPheArgThrHisPheSerGlnProSerGlyArgGluAlaAlaGluAsnAsnAspAlaTyrAspLysValLysLeuLeuA
2760 ACGAGGGGTTAGATGGTAAAATACAAAATAATAGATATAGATTCGCATTTCTATACTTGTTGGTGAAATGGTACAGAAATATCATGTTCCTATTATGAAACTATATCCTACACCCGAAG
spGluGlyLeuAspGlyLysIleGlnAsnAsnArgTyrArgPheAlaPheLeuTyrLeuLeuValLysTrpTyrArgLysTyrHisValProIleMetLysLeuTyrProThrProGluG
2880 AGATTCCTGACTTTGCATTCTATCTCAAATAGGTACTCTGTTAGTATCTAGCTCTGTAAAGCATATTCCATTAATGACGGACCTCTCCAAAAGGGATATATATTGTACGATAATGTGG
luIleProAspPheAlaPheTyrLeuLysIleGlyThrLeuLeuValSerSerSerValLysHisIleProLeuMetThrAspLeuSerLysLysGlyTyrIleLeuTyrAspAsnValV
3000
TCACTCTTCCGTTGACTACTTTCCAACAGAAATATCCAAGTATTTTAATTCTAGACTATTTGGACACGATATAGAGAGCTTCATCAATAGACATAAGAAATTTGCCAATGTTAGTGATG
alThrLeuProLeuThrThrPheGlnGlnLysIleSerLysTyrPheAsnSe rArgLeuPheG lyHisAspIleG luSe rPheIleAsnArgHisLysLys PheAlaAsnValSerAspG
3120
AATATCTGCAATATATATTCATAGAGGATATTTCATCTCCGTAAATATATGCTCATATATTTATAGAAGATATCACATATCTAAATGAATACCGGAATCATAGATTTATTTGATAATCAT
luTyrLeuGlnTyrIlePheIleGluAspIleSerSerProEnd MetAsnThrGlyIleIleAspLeuPheAspAsnHis
3240
GTTGATAGTATACCAACTATATTACCTCATCAGTTAGCTACTCTAGATTATCTAGTTAGAACTATCATAGATGAGAACAGAAGCGTGTTATTGTTCCATATTATGGGATCAGGTAAAACA ValAspSerIle ProThr Ile LeuProH isGlnLeuAl1aThrLeuAspTyrLeuValArgThrIleIl1eAspGluAsnArgSerVal1LeuLeuPheHisIl1eMetGlySerGlyLysThr
3360
ATAATCGCTTTGTTGTTCGCCTTGGTAGCTTCCAGATTTAAAAGGTTTACATTCTAGTGCCTAATATCAACATTTTGAAATTTTTAATTATAATATGGGTGTAGCTATGAACTTGTTT IleIleAlaLeuLeuPheAlaLeuValAlaSerArgPheLysLysValTyrIleLeuValProAsnIleAsnIleLeuLysIlePheAsnTyrAsnMetGlyValAlaMetAsnLeuPhe
3480
AATGACGAATTCATAGCTGAGAATATCTTTATTCATTCCACAACAAGTTTTTATTCTCTTAATTATAACGATAACGTCATTAATTATAACGGATTATCTCGCTACAATAACTCTATTTTT
AsnAEspGl1uPheIleAl aGluAsnI le Phe IleHis SerThrThrSerPheTyrSerLeuAsnTyrAsnAspAsnVal1Il1eAsnTyrAsnGlyLeuSerArgTvrAsnAsnSerIlePhe 3600
ATCGTTGATGAGGCACATAATATCTTTGGGAATAATACTGGAGAACTTATGACCGTGATAAAAATAAAAACAAGATTCCTTTCTACTATTGTCTGGATCTCCCATTACTAACACACCT
IleVa1Asp(;luAlaHisAsnI lePheGlyAsnAsnThrGlvC:luLeuMetThrVal1IleLvsAsnLysAsnLvsIleProPheLeuLeuLeuSerC:1vSerProlleThrAsnThrPro 3670
AATACTCTCGGTCATATTATAGATTTMTGTCCGAAGAGACGATAGATTTTGGTGAGATTATTAGTCGTGGTMG AsnThrLeuGlyHisIleIleAspLeuMetSerGluGluThrIleAspPheGlyGlulleIleSerArgGlyLys
1400
on November 10, 2019 by guest
http://jvi.asm.org/
VACCINIA VIRUS GENE REQUIRED FOR DNA REPLICATION 1401
Ix w ccx co Om cc w co mmc cca: x wx ir I
11 11
[image:4.612.59.305.72.125.2] [image:4.612.329.550.74.182.2]25 kd
I...
90.3 kd
D5
22.2 kd+ D6 5 kb
FIG. 3. Restriction map of theEcoRI B fragment. The complete ORF is shown by asolid line; incomplete ORFs are indicated with a broken line. Predicted molecular weights of the ORFs are indicated along with their numerical designations. Rl, EcoRI; R, RsaI; X, XbaI;P,PstI;E,EcoRV; B,BglII; C,CMaI; K, KpnI; S,SalI;H, Hinfl.
RESULTS
Marker rescue mapping. Previously reported marker res-cuemapping,usingtheHindIlI fragmentsof VV, located the tsl7locuswithinthe 16-kilobase (kb)HindlIl Dfragment (7). Subclones derived by restriction enzyme digests of wt HindIIIDwhichwereusedfor fine mapping ofthets17 locus are shown (Fig. 1). Viral DNA fragments were calcium phosphate precipitated and transfected into BSC40 cells whichhadbeen previously infected withtsl7virus. Aftera 48-h incubation at the nonpermissive temperature of 40°C, theinfectedcells wereharvested,andtitersof viral progeny weredetermined at the permissive temperature of 31°C and at the nonpermissive temperature of 40°C (Table 1). At permissivetemperature,tsl7 replicationwasindependent of the identity of the transfected DNA fragment, whereas at nonpermissive temperature replicationwasdependentupon the homologous recombination of a wt viral DNA fragment which contained the appropriate sequences. Marker rescue usingpSWl,a7.0-kb fragmentderived fromtheright-hand side of HindIII D, resulted in tsl7 remaining temperature sensitive. However, the 8.1-kb BamHIAfragmentfrom the left-hand side ofHindlllDwas able toconfertemperature insensitivity.Themutationwasmappedfurther to an EcoRI subclone ofBamHI A, the3.3-kbEcoRIBfragment.EcoRI B wassubclonedintothreeBglIIfragments, andofthese the 1.3-kbBglIIAfragment contained thesequencesrequiredto confertemperature insensitivityto tsl7.
Nucleotidesequence and identification of the ts17 mutation. Tofacilitate molecularstudies ofthe tsl7gene, the nucleo-tidesequences of the 3,309-base-pairEcoRI Bfragment and of 300 base pairs reading rightward into the neighboring EcoRI Afragment were determinedby the dideoxynucleo-tide chain termination method
(Fig.
2). These sequences were determined by cloning appropriate overlapping subfragments ofviral DNAinto M13 vectors inboth orien-tations so that both strands could be sequenced. By this strategy, over 80% of the sequence of both strands was determined. The sequence reported here is in complete agreementwith thatrecentlyreported
(27).Analysis of ORFs found withinthesequencereportedhere showed tandemly oriented ORFs which read from left to
right (Fig.
3). In accordance with therecently
proposed
nomenclatureforVV genes,thesewere
designated
D4,D5,
and D6. D4 is incomplete in this sequence, with the G at
position 1actually beingthethirdbaseoftheinitiation ATG codon ofD4 (27). Thus, the ORF D4 has the
capacity
toenqode a
25,055-molecular-weight
polypeptide. Thecom-pleteD5 ORF, which spans thesite ofthe tsl7
mutation,
isWT AGT AGA TCA TCT GAAAAT CCA CTA ACAAGA TCG SerArg Ser SerGlu Asn Pro Leu Thr Arg Ser tj 17 AGT AGATCA TTT GAAAAT CCA CTA ACC AGATCG SerArg SerPhe Glu Asn Pro Leu Thr Arg Ser t 17rl AGT AGATCA TCT GAA AAT CCA CTAACC AGA TCG Ser Arg Ser Ser Glu Asn Pro Leu Thr ArgSer
t 17-r2 AGT AGATCk TCT GAAAkT CCA CTA ACCAGA TCG Ser Arg Ser Ser Glu Asn Pro Leu Thr Arg Ser
FIG. 4. Nucleotide sequenceof theBglIIAfragment was deter-mined from viral DNAisolated from wt, tsl7,and twotsl7 rever-tants (ts17r1 and Ws17r2). Shown is the sequence from nucleotide positions 1158 to 1190. The amino acid encoded is also shown. The nucleotide positions atwhich there is a changefrom wt sequences areindicated (*).
2,297 nucleotideslongandcouldspecifyapolypeptide with apredicted molecular weight of 90,367.Theincomplete ORF D6 encodes 22,234 daltons of what is reported to be a 68,362-daltonpolypeptide (27).
Toconfirm the locationofthetsl7gene and to determine thenatureofthemutation giving risetoit, the 1.3-kb BglII A fragment was subcloned from tsl7 viral DNA and the nucleotide sequence wasdetermined. Furthermore, a num-beroftsl7 revertants wereisolatedby repeatedpassagesof tsl7 virus at40°C. TheBgIII Afragments from two rever-tants (tsl7rl and
ts17r2)
were also cloned and sequenced (Fig. 4). A comparison between the wt and tsl7 sequences identified twocloselylinkedmutations. Thefirst isa transi-tion mutatransi-tion ofaCto a T atnucleotide position 1168, which causes an amino acidchange ofaserineto aphenylalanine.
Thesecondmutation isatransversion mutationofanA to a C at nucleotide position 1184; in this case the mutation is silent,and the amino acidbeing encoded remains threonine. Sequences derived fromthe twotsl7 revertants were iden-ticalandshowed both tsl7rl and tsl7r2tobetruerevertants. The nucleotide atposition 1168 had reverted back to a C, returningthe codon to a serine. However, the silent muta-tion remained unchanged, thus indicating that tsl7rl and ts17r2 were tsl7 revertants and not wtVV. Therefore, the sequence data (Fig. 4) and the genetic evidence
(Table
1) demonstrate that atransitionmutationatnucleotideposition
1168 isresponsible for thetsphenotype of tsl7 virus. Hydrophilicity-hydrophobicity analysis. The effect ofthe single amino acid change causedby the transition mutation on the hydropathic index (17, 35) of the
predicted
tsl7 proteinwas determinedbyacomparison ofthehydropathy profiles of tsl7 and wt(Fig.
5). The amino acidchange
clearly causedalocalized
change
inthehydropathy profile
of tsl7ascomparedto wt. Thecodons whichdirectly
precede
andfollowthemutationreflectahigher degree of
hydropho-bicity. The hydropathy
profile
ofthe entire tsl7protein,
aspredicted from wt sequences, was also
analyzed.
The tsl7 proteinappearstohaveaprofile
typical
ofaglobular
protein,
althoughthere isastrong
hydrophobic
regionatthecarboxy
terminusof the
polypeptide (data
notshown).
Hybrid selection andin vitrotranslation. Todeterminethe tra-nslation
products
of thetranscripts
encodedfor withinvv
FIG. 2. Nucleotide sequencereadingrightwardbeginning attheleft-handEcoRIsiteofEcoRIBand
extending
300basepairs
rightward
into the EcoRI Afragment. The RNAstart sites determinedby S1nuclease mappingaremarked with arrowheads. The sites of the tsl7 mutationsareindicated(*).VOL. 61,1987
on November 10, 2019 by guest
http://jvi.asm.org/
155 160 165
AMINOACID POSITION
150 155 160 165 170
AMINO ACID POSITION
FIG. 5. Hydrophobicity profileofwt(left)and tsl7(right)in the regionofthetsl7mutation. Theyaxis is the freeenergyoftransfer ofamino acidsidechains; thexaxis is theaminoacidposition.The
analysis shownhere beginsatcodon 150, which correspondswith nucleotide position 1137. The tsl7 mutation changes the codon at amino acidposition 162.
EcoRI B,polyadenylated immediate-early viral RNA hybrid selected by EcoRI B was translated in vitro in rabbit reticulocyte lysatesin the presenceof[35S]methionine.
La-beled protein products were then electrophoresed on a
denaturing sodium dodecyl sulfate-polyacrylamide gel and autoradiographed (Fig. 6).
The in vitro translation systemprogrammed with mRNAs selected by the EcoRI B fragment yielded two major polypeptides with molecular weights of approximately 25,500 and 88,000. This correlates well with the size of polypeptides thatcanbepredicted from the coding capacity
of D4 and D5, i.e., 25,055 and 90,367 molecular weight, respectively. There does not appear to be a translation product that correlates toD6. This maybe due toD6being of the lateviralgeneclass, and thus its transcript wouldnot bepresentin themRNA population usedin thisexperiment.
(-l EP ET _W- .. .
EcoB pUC
200-
97.4-
68-
43-25.7- m
18.4-
14.3-__rn,
The two minor polypeptides of approximately 22,000 and 45,000 molecular weight are endogenous proteins of the translation system andnotofviralorigin.None of theEcoRI B-derived polypeptides comigrated with any of the major early viral proteins produced either in vivo or in vitro (Fig.6, lanes EPand ET).
Identification of transcripts by Northernanalysis. To iden-tify mRNAs that are transcribed within EcoRI B, polyadenylated immediate-early viral RNA was electropho-resed under denaturing conditions on a formaldehyde-agarose gel and transferred to nitrocellulose(Fig. 7). Nick-translated EcoRI B DNA used as aprobe revealed a major transcript of 2.7 kb and a minor transcript of 3.6 kb. To localize these transcripts more specifically, the BglII subclones of EcoRI B were used asprobes; it was found that both transcripts span the three BglII fragments (data not shown).
To further map these transcripts, the EcoRI D and A fragments, which border EcoRI B, were used as probes. Neither EcoRI D nor A appeared to hybridize to the two transcripts which are coded for within EcoRI B. The 2.6-kb transcriptwhichhybridized to EcoRI D correlated well with Dl and may encode the viral guanyl transferase (24). The 0.57- and 0.98-kb transcripts which map to EcoRI A can possibly be assigned to D7 and D8, respectively. The ab-senceof alarge transcript which can be assigned to D6 may again be due to thisgene's beingexpressedlate in infection. This analysis appears to show that the 2.7- and 3.6-kb transcriptsarecodedforentirely withinEcoRI B. In the case of D4, there does not appear to be a transcript of the appropriate size that would be predicted to encode the 25-kilodaltonpolypeptide whose presence was indicated by the sequencedata andbytheresults of thehybridselection in vitro translation. This result is not due to potential pleiotropiceffects of thedrugblock(cycloheximide)used to isolate abundantearly viral RNA. Onlythe 2.7-and 3.6-kb transcripts were hybridized by EcoRI B when aNorthern analysiswasrepeated
using
viralRNAisolatedat2 hp.i.
in theabsence ofdrug (datanotshown).-88
3 4
-3.6 _2.7
2.6
0 *
_0
PF
__
*.S
.-. ;:,P
...
-25.5
FIG. 6. Hybrid-selected cell-free translation. Polypeptides
syn-thesized in rabbitreticulocyte lysateswere separated by gel elec-trophoresis. Lanes:(-),noRNA added;ET,total early viral RNA; EcoB,VVearlyRNAhybrid-selectedbyEcoRIB;pUC, early viral RNA hybrid-selected by pUC-18. Lane EP, Polypeptides pulse-labeled at 1.5 h p.i. in wt VV-infected cells. Numbers at theleft indicate molecular weights (x103) of protein molecular weight markers.
-.98
[image:5.612.63.302.72.173.2]-.57
FIG. 7. Northern blotanalysis. Twomicrogramsof polyadenyl-ated early viral RNA was electrophoresed on a
formaldehyde-agarose gel and then blotted onto nitrocellulose. Nick-translated plasmids containingfragments ofVV DNAwere usedas probes:
lane1,EcoRID; lane2,EcoRI B;lane3, BglII A;lane4, EcoRI A. Numbersatright indicatecalculated transcript sizes (kb) determined by coelectrophoresis of brome mosaic viral RNAs as molecular
weightmarkers.
1.04-
0.79-0
053-Lz0.28- I
0i0.02--0.23-U
-.049-ts1517
1.04--07
I0
0.53-
0.28--.049
-0.74
-0.74-150
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.612.97.267.432.650.2] [image:5.612.375.505.478.652.2]VACCINIA VIRUS GENE REQUIRED FOR DNA REPLICATION 1403
A B
Si A+G
.., _."P
-4!_:_
A i- _ _ 7;j ;- _m
4;
A
A
C
A-A
C\I
PstI 404 Rsal
150
C Eco B
2 5.5 kd
90.3kd
FIG. 8. (A) S1 nuclease mapping of transcripts encoded by EcoRIB.Labeled DNAfragments were hybridized with early RNA and treated with Si nuclease, and protected hybrids were then analyzed on sequencing gels. Lane 1, Sau3A digest of pUC18; lane 2,full-length404-nucleotide5'-end-labeled probe; lane 3, S1 analy-sisusing 404-nucleotide probe; lane 4,full-length 2,080-nucleotide 5'-end-labeled probe; lane 5, Si analysis using 2,080-nucleotide probe; lane 6, S1 analysis using 3'-end-labeled 3,286-nucleotide probe (the full-length3,286-nucleotide probe does not enter the gel system). Sizes in nucleotides of the probes and protected fragments areindicated. (B)S1nucleasemappingof themRNAstartsite. Lane S1 is the S1 nuclease-resistant fragment (wavy line) from the hybridization of a 5' single end-labeled probe (solid line) and early mRNA. Lane A+G is a Maxam-Gilbert A+G reaction of the indicated probe. The sizes of the probes and the protected fragments areindicated innucleotides. The sites of initiationareindicated with arrowheads within the contextof the sequence. (C) Diagrammatic representation of theorganization of earlymRNAs transcribed in EcoRIB.
Sinucleasemapping. Todeterminetheorganizationof the twoearly transcripts encoded within EcoRI B, S1 nuclease protection studiesweredone. Inthesestudiestheindicated probe washybridized to 20
jig
of totalRNA isolatedfrom infected cells in the presence ofcycloheximide. After S1 digestion, the protected fragmentswereelectrophoresedon adenaturing polyacrylamide
gel (Fig. 8A).To map the 5' endofthe D4transcript, a
2,080-nucleotide
5' single end-labeled XbaI probe was isolated. This
probe
protecteda fragment of
approximately
310 bases,mapping
the5' startofD4 to
approximately
23 nucleotidesupstream of theEcoRI D-Bjunction
and theinitiationATG. The 5'end of D5, which encodes the tsl7 gene, was determined by utilizing a 404-nucleotide 5'single
end-labeled PstI-RsaI fragmentas aprobe.Aprotected
fragmentofapproximately
150 nucleotides maps the transcriptional start of D5 about 690nucleotides downstreamof the EcoRI D-B
junction.
To map the transcriptional stop of the two
transcripts,
DNAfragments which
spanned
theentireEcoRl Bfragment
were3'end labeled and usedasprobes. Theresultsshowed
full-length
protection oftheseprobes,indicating
that there is no early termination oftranscription
within the EcoRI B fragment (data not shown).Therefore,
a3,286-nucleotide
Sall
fragment
which spans the EcoRI B-Ajunction
was 3'end labeled and used as a probe. A protected fragment of 1,350nucleotidesmapped a single 3' endapproximately 200 nucleotides downstream of the EcoRI B-A junction. A secondprotectedfragment(Fig. 8A, lane 6) does not have a 3' end within EcoRI B, since a smaller overlapping probe showed only full-length protection. This protection must arise from a leftward-reading early transcript encoded by EcoRIA.
This analysis identifiedtwo earlytranscriptsreading from lefttoright which areapproximately 3,590 and 2,877 nucle-otides in length. This is consistent with the Northern data and shows that the twotranscripts terminatecoincidentally (Fig.8C).
To facilitate future studies ofthe ts17 gene and its pro-moter, thetranscriptional start site for D5 was specifically mapped. The404-nucleotidePstI-RsaI fragmentwas 5' end labeledat the RsaIsite,gelisolated, andhybridizedto 20 ,ug of viral RNA as described above. After
Si
nuclease diges-tion, protected fragments were electrophoresed alongside theproducts ofanA+Gchemical sequencingreaction of the 404 nucleotide probe (Fig. 8B). Multiple transcription initi-ation sites were mappedatpositions -2 to -5nucleotides, upstreamofthefirst ATG ofD5.Initiationoccurswithinthe sequenceTAA, which lies directlyupstreamoftheputative initiation codon, ATG.Transcript kinetics. Since the sequence element TAAAT has been proposedto be a lateregulatory element and has herebeenshown to occurinagene theexpression of which isrequired early,thekinetics oftranscription initiation ofthe tsl7 gene was determined. This was accomplished by S1 analysis of the steady-statemRNAlevels overtime.
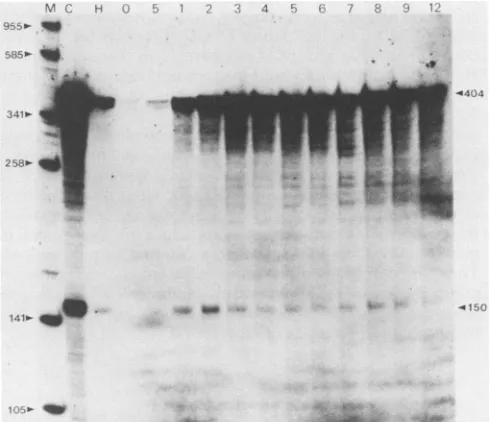
The 5' probe previously described (Fig. 8B) was hybrid-ized to 20 ,ug of total RNA, which was isolated at hourly intervals throughout infection. This analysis also included RNA isolated in the presence of
cycloheximide
and hydroxyurea.Cycloheximide
treatmentallowstranscriptionV1M h :1
q-.. W
C, I lX
341m-28
1410- 40 '
41l0
FIG. 9. Kinetics of transcription initiation. A 5' single end-labeled probe (see Fig. 8B)was hybridized to20 Fgoftotal viral RNAisolated in thepresence ofcycloheximide (C)orhydroxyurea
(H)or attheindicated timespostinfectioninthe absence ofdrugs.
Marker is a Sau3A digest of pUC18 (M). The 404-nucleotide full-length probeisshown, alongwiththe sizeofthe Si nuclease-protectedfragment.
VOL.61, 1987
141- _M
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.612.56.300.72.303.2] [image:6.612.317.562.448.659.2]of
immediate-early
genesonly,
whereas withhydroxyurea
treatment,
delayed-early
genes are also transcribed. Both treatments block late geneexpression.
After Si nuclease
digestion,
aprotected fragment
ofapproximately
150 basepairs
was detected(Fig. 9).
Thiscorresponds
tothe5' startlocatedinFig.
8A and B. Thetsl7gene was first transcribedat 1 h
p.i.
The level oftranscrip-tion initiatranscrip-tion remained stableuntil3h
p.i.,
when itappeared
todecrease. Thetsl7transcript
wasthenpresent for upto12 hp.i.
Since the half-life ofVV mRNA lateininfection has been estimated to beapproximately
1 to 2 h(13, 30),
the presence of tsl7 RNA at late times is mostlikely
due tocontinued
transcription.
Regulation
ofthymidine
kinaseexpression
by W DNA-mutants. Of the threecomplementation
groups which have been determinedto haveaDNA-phenotype,
only
onehasbeen
assigned
abiological function,
thatbeing
the DNApolymerase
gene(16, 36).
Itremains to be shown whether the loci of the other two DNA-complementation
groups encodeproteins
which have abiological activity
thatis anintegral
part ofDNAsynthesis,
orratherplay
anessentialrole
early
in the VVreplicative
cycle
which is notdirectly
related to DNAmetabolism. Itwas therefore of interestto
examine the
regulation
ofthe VVthymidine
kinasegene(tk)
in DNA- mutant-infected Ltk- cells to ascertain at which
point
the orderedexpression
of this genemight
beinter-rupted. Expression
of the VV tk gene is regulated at thetranscriptional, translational,
andposttranslational
levelsby
other VV gene
products
from both theearly
andlate gene classes(12, 13).
The kinetics of tk
expression
in cells infected withwt orwithtsmutantsof the threeDNA-
complementation
groups wasdetermined(Fig. 10).
Inwt-infected cells tk isexpressed
at theonset ofinfection,
is attenuated at 4to 6 hp.i.,
and thenremains stable.The levelofexpression
ishigher
at40°C
than at
31°C,
probably
due to metabolic processesbeing
increasedat the
higher
temperature. ThethreeDNA-com-plementation
groups,5, 21,
and24,
arerepresented by
ts42,tsl7Its24,
andts25, respectively.
Atthepermissive
temper-ature of31°C,
theregulation
of tkexpression
for all threecomplementation
groups iscomparable
to thatseenforwt.However,
at thenonpermissive
temperature of40°C
thereare distinct differences in the
impact
of each locus and its defectivegeneproduct
ontkexpression.
Inthecase of tsl7and
ts24,
whichtogether
comprise complementation
group21,
theexpression
ofthe tk gene is not switched off. ts42,which is the DNA
polymerase
gene, shows kinetics ofexpression
verysimilartothatseenforwtvirus.Inthecaseof
ts25,
theexpression
of tkdoesnotappear tobe induced;the levels of tk
activity
are very close to that found inpurified
viralcores(D.
E.Hruby,
submitted forpublication).Thus,
thederepressed
phenotype
seenforcomplementationgroup 21 is not the direct result of the DNA- phenotype since theother DNA-
complementation
groups exhibitdis-tinctly
different kineticsof tkexpression.DISCUSSION
Transcriptional
mapping
and sequencing studies haveshown VV genes to be organized in a tightly clustered,
tandemly
oriented fashion(8, 20, 21, 27, 28, 40, 45). Here,we
independently
confirm a portion of previously reportedsequence and showthatthepredictedD4and D5 ORFs are
transcriptionally
active. D4istranscribedearly in infection.Whether it is also
transcriptionally
active at later times is unknown. D4 has apredicted
coding capacity for a25,055-molecular-weight
polypeptide. However,
thetranscript
which is
apparently
derived from the D4 gene isapproxi-mately
3.7 kblong,
muchlarger
thanisrequired
toencodea25.5-kilodalton
protein.
In contrast, the 3.0-kbtranscript
associated with D5 is of the
appropriate
size to encode a90,367-molecular-weight
protein product,
identified as the tsl7 gene product. The data inFig.
8 suggest that the D4transcript
terminates at the sameplace
as the downstream D5transcript.
There doesnotappeartobeatranscriptional
stop
signal
proximal
to the 3' end ofD4. In contrast, the proposedtranscriptional
termination sequenceTTTTTAT
(31) appears twice, beginning at nucleotide
positions
3406 and 3475atthe 3' end ofthe D5transcript.
This is within thecoding
sequence of D6. These twosignals
mapapproxi-mately 150and 80 bases upstream of the shared3'terminus ofthe two early transcripts mapped by theS1 analysis.
wt
3-
2-ts17 ts24
3-
2-0
ts25 ts 42
3-
2-2 4 6 8 10 2 4 6 8 10
HOURS POST-INFECTION
FIG. 10. Thymidine kinase activity. Thymidine kinase assays were carried out on duplicate sets of 60-mm dishesof Ltk- cells which wereinfected with the indicated ts mutant. The plates were incubated at either the permissive temperature of 31°C(0) orthe nonpermissive temperature of 40°C (-) and harvested at 2-h intervals.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.612.326.559.250.673.2]VACCINIA VIRUS GENE REQUIRED FOR DNA REPLICATION 1405 Marker rescue mapping and the comparison of sequences
derived from wt, tsl7, and two tsl7 revertant viral stocks have conclusively demonstrated that D5 is the tsl7 locus. This locus encodes a gene product which appears to be functionally required early in the viral replication cycle. Without afunctionaltsl7 gene, theviral replication cycle is blocked prior to DNA replication. Yet, a kinetic analysis shows that the tsl7 locus is transcribed continuously throughout infection, beginningat 1 h p.i., and is therefore a memberof the constitutive temporal class.
The increasing amount of VV sequence data available allowsfor acomparative analysis of putative viralpromoters and thusfacilitatestheidentification ofconsensus sequences which may beregulatory elements. Transcription initiation atD5 hasmultiple start sites which occurbetween2 and 5 nucleotidesupstreamoftheinitiation ATG. Similar findings havebeenreported for both early (5,39, 43) and late (2, 11, 33, 44)VVgenes. In the latter case,transcriptional initiation occurswithinthesequenceTAA,which isdirectlyupstream of theinitiation ATG. It has beenproposed thatthe sequence TAAATisessential for latepromoterfunction(10)andmay actually bepartofasignalsequencefor lategeneexpression (33). However, since the sequence TAAAT serves as the
transcriptional
start site forthe D5 locus atboth early and late timespostinfection,
this indicates that there must be additional factors involved with specifying the temporal expression ofVVgenes.Thisfinding illustratestheflexibility thatmustberequiredtoregulatethecomplexexpression ofa
variety
ofspecific
temporal classes of virally encodedgeneproducts.
The tsl7gene is
comparable
topreviously
sequencedVV genes inthatitis 66%A-Trich,
withthefirst100nucleotides upstream of the 5' start being 71% A-T rich. A close examination ofthe first 100 nucleotides shows an A-T-richoctet at
positions
20and40nucleotidesupstreamofthefirst ATG. A-T octets at thisposition
have been proposed as regulatory elements for early VV genes (38, 39, 43, 44).However,
the octets of D5 are not homologous to theproposed
consensussequenceproposed fortheseregulatory elements. Itappears that the D5 gene has promoter elements which have been associated withboth early and late consen-sus sequences. This makes clear thenecessity
for further sequenceanalysis
inconjunction
with kinetic studies foridentifying temporal
subsets of viral geneexpression.
A directedgenetics approach
will then berequired
to deter-mine which elementsareinvolved inregulating
gene expres-sion. Suchanapproach
iscurrently being
usedtodefinethe elements involved in theexpression
andregulation
of the tsl7gene.ACKNOWLEDGMENTS
We thank R. C. Condit for generously providing his entire collection oftemperature-sensitive mutants to our laboratory. We thank LisaWilson, Walt Hodges, and Chris Franke for excellent technical support. We also thank ScottL. Weinrich for theuseof
timecourseRNAs andforhelpfuldiscussionsthroughoutthiswork.
Thisstudywas supported byaPublic Health Service grant from the National Institutes of Health(AI-21335)andbyagrant fromthe American Cancer Society (MV-194). D.E.H. was supported by a
Research CareerDevelopmentAward(AI-00666)fromtheNational Institutes of Health.
LITERATURECITED
1. Belle Isle, H., S. Venkatesan, and B. Moss. 1981. Cell-free translation ofearly late mRNAs selected by hybridization to
cloned DNAfragments derived from the left 14 millionto 72
million daltons of the vaccinia virus genome. Virology 112:306-317.
2. Bertholet, C., R. DrilHien, andR. Wittek. 1985. One hundred basepairs of5'flanking sequence of a vaccinia virus late gene are sufficient to temporally regulate late transcription. Proc. Natl. Acad. Sci.USA 82:2096-2100.
3. Cabrera, C.V.,M. Esteban, R.McCarron, W. T. McAllister, andJ.A.Holowzak. 1978. Vaccinia virustranscription: hybrid-ization of mRNA to restrictionfragments of vaccinia DNA. Virology 86:102-114.
4. Chipchase, M.,F.Schwendimann,and R.Wyler. 1980.Amapof thelateproteins of vaccinia virus. Virology 105:261-264. 5. Cochran, M. A., C. Puckett, and B. Moss. 1985. In vitro
mutagenesis of the promoter region foravaccinia virus gene: evidence for tandemearly and late regulatory signals. J. Virol. 54:30-37.
6. Condit,R.C.,and A.Motyczka.1981. Isolation andpreliminary characterization oftemperature-sensitive mutants of vaccinia virus.Virology 113:224-241.
7. Condit, R. C., A. Motyczka, and G. Spizz. 1983. Isolation, characterization, and physical mapping of temperature-sensitive mutantsof vaccinia virus. Virology 128:429-443.
8. Cooper, J. A.,R.Wittek,and B. Moss. 1981. Extensionof the transcriptional and translational map of the left end of the vaccinia virus genometo21kilobase pairs. J. Virol. 39:733-745. 9. Graham,F. L.,and A.J.vander Eb.1973. Transformation of ratcellsbyDNAof human adenovirus 5. J. Virol. 54:536-539. 10. Hanggi, M., W. Bannwartb, and H. G. Stunnenberg. 1986. Conserved TAAAT motif in vaccinia virus late promoters: overlapping TATA box and site of transcription initiation. EMBO J.5:1071-1076.
11. Hirt, P.,G. Hiller, and R.Wittek.1986. Localization andfine structureofavaccinia virus geneencodinganenvelopeantigen. J. Virol. 58:757-764.
12. Hruby,D. E.1985.Inhibition of vaccinia virusthymidinekinase bythe distalproductsofitsownmetabolicpathway.VirusRes. 2:151-156.
13. Hruby,D.E.,and L. A.Ball. 1981.Control ofexpressionof the vaccinia virusthymidinekinase gene. J. Virol. 40:456-464. 14. Hruby,D.E.,and L. A.Ball. 1982.Mappingand identification
of the vaccinia virus thymidine kinase gene. J. Virol. 43: 403-409.
15. Hruby, D. E.,R. A.Maki,D. B. Miller, and L. A.Ball. 1983. Finestructureanalysisandnucleotide sequences of the vaccinia virus thymidine kinase gene. Proc. Natl. Acad. Sci. USA 80:3411-3415.
16. Jones,E.V.,and B. Moss. 1984.Mappingof thevaccinia virus DNA polymerasegenebymarkerrescue andcell-free transla-tion ofselected RNA. J. Virol. 49:72-77.
17. Lawson, E.,A.Sadler,D.Harmatz,D.Brandau,R.Micanovic, R.MacElroy,and C. R.Middaught.1984.Asimple
experimen-tal model for hydrophobic interactions in proteins. J. Biol. Chem. 259:2910-2912.
18. Lehrach, H.,D.Diamond, J.M.Wozney,and H. Boedtker. 1977. RNA molecular weight determinations by gel electrophoresis underdenaturingconditions,acriticalreexamination. Biochem-istry16:4743-4752.
19. Mackett,M.,G. L.Smith,and B. Moss.1984. General method forproductionandselection of infectious vacciniavirus recom-binantsexpressing foreigngenes. J.Virol. 49:857-864. 20. Mahr, A., and B. E. Roberts. 1984. Organization of sixearly
transcripts synthesized from a vaccinia virus EcoRI DNA fragment.J. Virol. 49:497-509.
21. Mahr, A.,andB.E.Roberts. 1984.Arrangementof late RNAs transcribedfroma7.1-kilobase EcoRI vacciniaDNAfragment. J. Virol. 49:510-520.
22. Maniatis, T.,E. F.Fritsch,andJ. Sambrook. 1982. Molecular cloning:alaboratory manual. ColdSpringHarborLaboratory, ColdSpring Harbor,N.Y.
23. Maxam, A. M., and W. Gilbert. 1977. A new method for sequencingDNA. Proc. Natl. Acad. Sci. USA 74:560-564. 24. Morgan, J. R.,L. K.Cohen, and B. E. Roberts. 1984.
Identifi-cation of theDNAsequencesencodingthelarge subunit of the VOL.61, 1987
on November 10, 2019 by guest
http://jvi.asm.org/
mRNA-capping enzyme of vaccinia virus. J. Virol. 52:206-214.
25. Moss, B. 1985. Replication ofpoxviruses, p. 685-703. In B. N. Fields(ed.),Virology. RavenPress, New York.
26. Munyon, W., E. Paoletti, and J. T. Grace, Jr. 1967. RNA polymerase activity in purified infectious vaccinia virus. Proc. Natl.Acad.Sci. USA8:2280-2287.
27. Niles, E. G., R. C. Condit, P. Caro, K. Davidson, L. Matusick, andJ. Seto. 1986. Nucleotide sequenceandgenetic map of the 16-kb vacciniavirus Hind IIIDfragment. Virology 153:96-112. 28. Plucienniczak, A., E. Shroeder, C. Zettlmeissi, and R.E. Streeck. 1985.Nucleotide sequence ofacluster ofearly and late genes in a conserved segment of the vaccinia virus genome. Nucleic Acids Res. 13:985-998.
29. Queen, C., and L. J. Korn. 1984. Acomprehensive sequence analysis program for the IBM personal computer. Nucleic Acids Res.12:581-599.
30. Rice, A. P., and B. E. Roberts. 1983. Anovel mechanism of viral induced inhibition of host cell protein synthesis: vaccinia virus accelerates thelabilization of mouse L-cell mRNAs. J. Virol. 47:529-539.
31. Rohrmann, G., L. Yuen, and B. Moss. 1986. Transcription of vaccinia virus early genes by enzymes isolated from vaccinia virions terminates downstream of a regulatory sequence. Cell 46:1029-1035.
32. Rosel, J., P.L. Earl, J. Weir, and B. Moss. 1986. Conserved TAAATG sequence at the transcriptional and translational initiationsites of vaccinia virus late genes deduced by structural andfunctional analysis of the HindIII Hgenomefragment. J. Virol.60:436-449.
33. Rosel, J., andB. Moss. 1985. Transcriptional andtranslational mapping and nucleotide sequence analysis of a vaccinia virus geneencoding the precursor of the major core polypeptide 4b. J. Virol. 56:830-838.
34. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA
sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. USA74:5463-5467.
35. Taylor, W.R.1986. The classification of amino acid conserva-tion. J. Theor. Biol. 119:219-234.
36. Traktman, P.,P.Sridhar,R. C.Condit,and B. E.Roberts. 1984. Transcriptional mappingof theDNApolymerasegene of vac-cinia virus. J. Virol. 49:125-131.
37. Vassef,A.,M.Mars,A.Dru,A.Plucienniczak, R.Streeck,and G. Beaud. 1985. Isolation of cis-acting vaccinia virus DNA fragments promoting the expression of herpes simplex virus thymidinekinasebyrecombinant virus.J. Virol. 55:163-172. 38. Venkatesan, S.,B. M.Baroudy,and B. Moss. 1981. Distinctive
nucleotide sequences adjacenttomultiple initiation and termi-nationsites ofanearly vaccinia virus gene. Cell 25:805-813. 39. Venkatesan,S., A. Gershowitz, and B. Moss. 1982. Complete
nucleotide sequences of adjacent early vaccinia virus genes located within the inverted terminal repetition. J. Virol. 44: 637-646.
40. Weinrich, S. L., and D. E. Hruby. 1986. Atandemly-oriented late gene cluster within the vaccinia virus genome. Nucleic AcidsRes. 14:3003-3016.
41. Weinrich, S. L., and D. E. Hruby.1987. Noncoordinate regula-tion ofavaccinia viruslate gene cluster. J. Virol.61:639-645. 42. Weinrich, S. L., E. G. Niles,andD. E.Hruby. 1985.
Transcrip-tional and translaTranscrip-tional analysis of the vaccinia virus late gene L65. J. Virol. 55:450457.
43. Weir, J. P., and B. Moss. 1983. Nucleotide sequence of the vacciniavirusthymidine kinase gene and the nature of sponta-neousframeshift mutations. J.Virol. 46:530-537.
44. Weir, J. P., and B. Moss. 1984. Regulation ofexpression and nucleotide sequence of a late vaccinia virus gene. J. Virol. 51:662-669.
45. Wittek, R., B. Richner, and G. Hiller. 1984. Mapping of the genescoding for thetwomajor vaccinia viruscorepolypeptides. Nucleic Acids Res. 12:4835-4848.