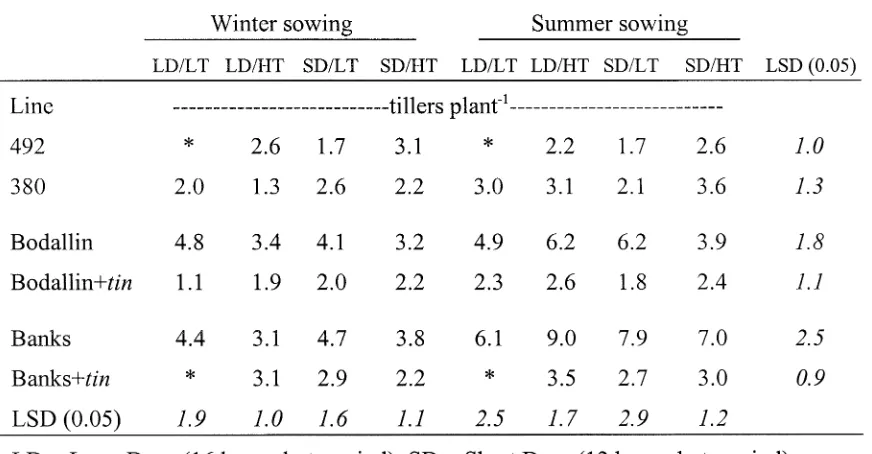
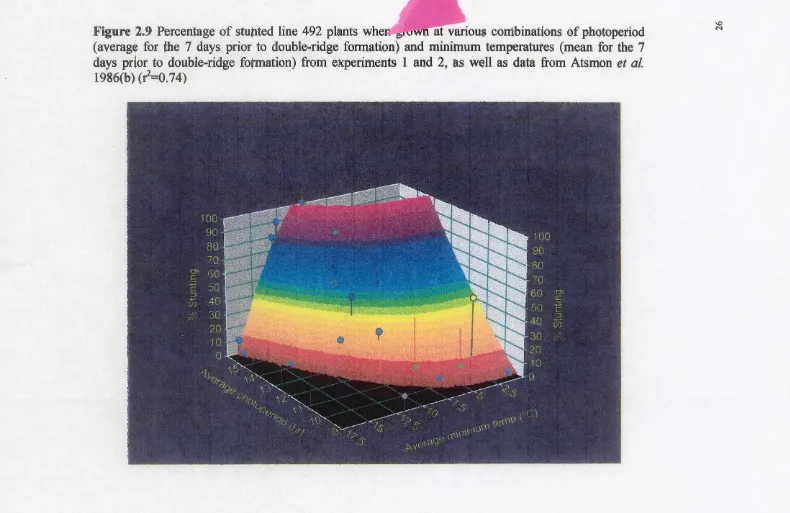
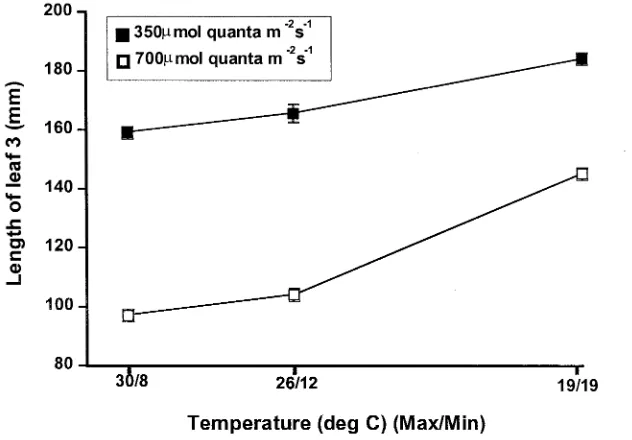
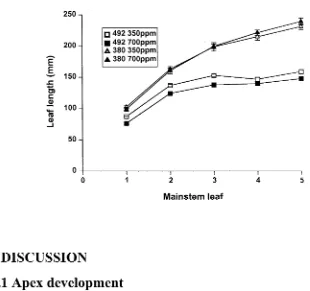
Evaluation of a tiller inhibition (tin) gene in wheat
Full text
Figure
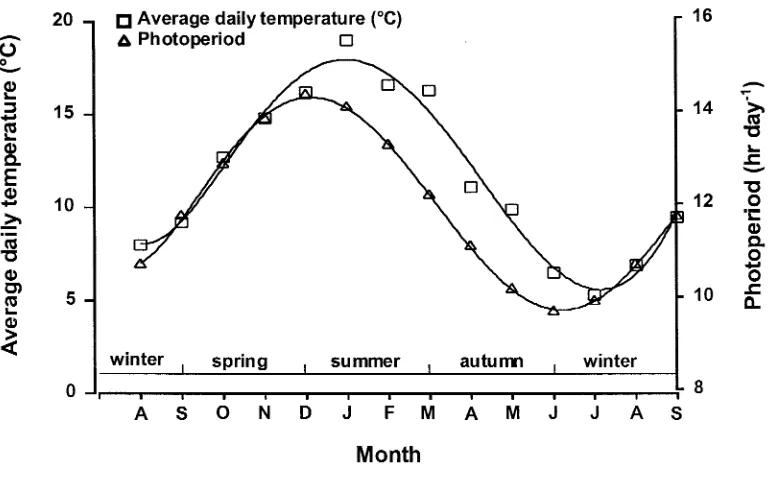
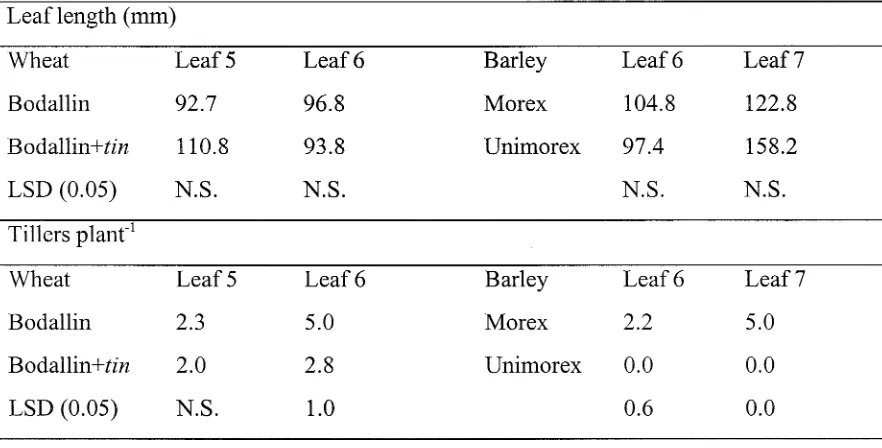
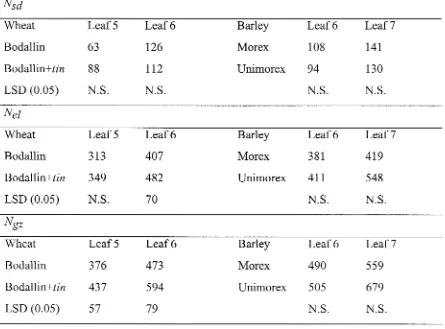
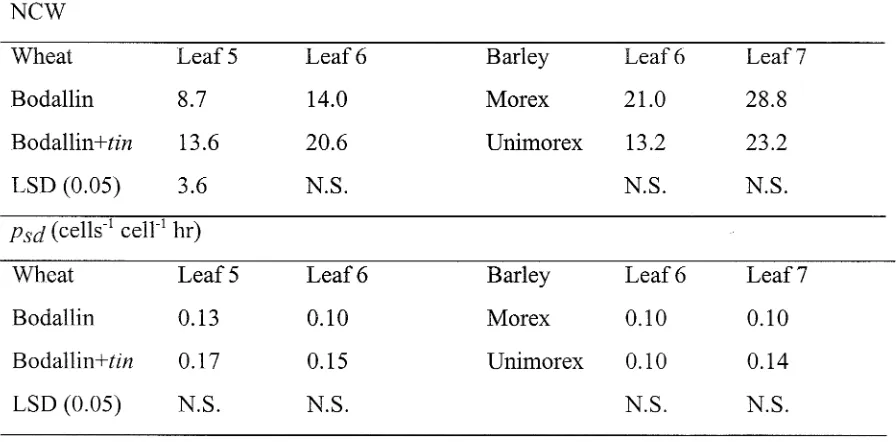
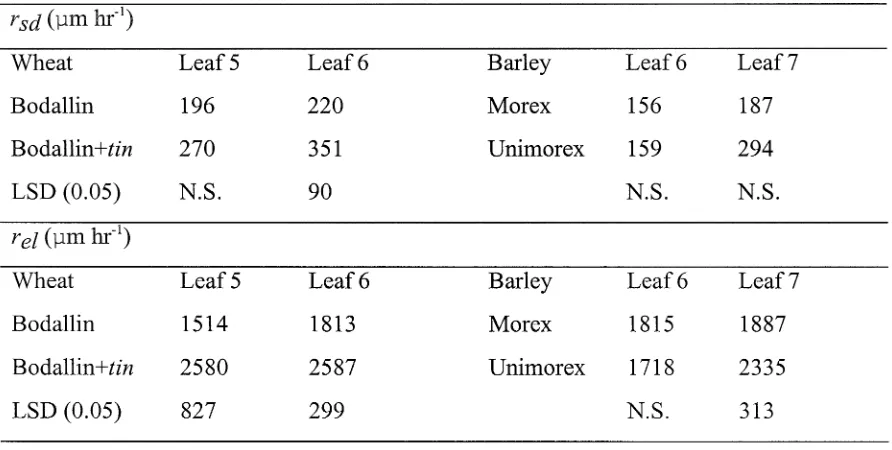
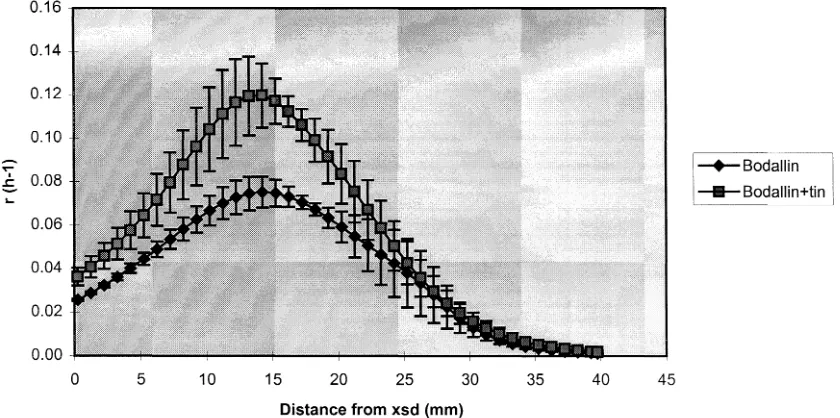
Related documents
In this paper, a collocation method based on RBFs for numerical solution of three-dimensional linear Fredholm integral equations of the second kind on the cubic domains is
Methods: This is a cross-sectional study of undergraduate medical students on self-rated measures of empathy collected at entrance and at the conclusion of the medical degree, and
Conclusions: Compared with non-Xp11.2 translocation RCCs, the Xp11.2 translocation RCCs seemingly showed a higher tumor grade and pathologic stage and have similar
Background: Famine early warning systems clearly identified the risk of famine in South Central Somalia in 2010 – 2011 but timely action to prevent the onset of famine was not
study, for the model group, we detected significantly increased HIF-2 α levels
While over-the-counter drugs are the most commonly over- used headache medications in primary care, accounting for approximately 60% of overused medication, second- ary
Annual product review is an evaluation conducted annually to assess the quality standard of each drug product with a view to verify the consistency of existing process and to check
The matrix type patches were prepared using polymers such as HPC, Hydroxyl Propyl Methyl Cellulose (HPMC E15CPS, HPMC E50CPS) by solvent casting method and to study the effect
