A clinical cytogenetical study of the horse : a thesis presented in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Veterinary Science at Massey University
239
0
0
Full text
(2) i. A CLINICAL CYTOGENETICAL STUDY OF THE HORSE. A thesis presented in partial fulfilment of the requirements for the degree of Doctor of Philosophy in Veterinary Science at Massey University. Murray Garfield Blue B. V. Se. ( Massey ). 1976.
(3) ii ABSTRACT. Chromosome preparations were derived from short-term cultures of blood lymphocytes and from fibroblast-type cell cultures of subc�taneous fascia obtained from normal horses.. 44. clinically. The quality of mitotic cells compared. favourably with similar studies in other species.. Some. characteristic morphological features were found which aid in the recognition of certain chromosomes, but the identification of many homologous pairs by conventional techniques was difficult.. The normal karyotype of the horse ( 2n= 64). was characterized following Giemsa staining and C-banding. The latter procedure was shown to be a valuable aid for the identification of specific chromosomes, in particular the sex chromosomes.. After centromere staining, the X chromo-. some of the horse. lS. characterized by the presence of. prominent C-bands on its long arms, while the Y chromosome appears as an intensely-stained block. Analysis of mitotic chromosomes prepared from ten phenotypically normal but infertile mares revealed, in one animal, the consistent lack of one sex chromosome in the four tissues studied.. Centromere staining and sex chromatin. studies confirmed that this mare had the karyotype 63XO. Examination of the genitalia of this XO mare disclosed the presence of apparently normal. reproductive organs except. that macroscopic follicles were absent from both ovaries. Histologically, a number of presumptive atretic Graafian follicles and a discrete area of apparently functional luteal tissue were discovered in the right ovary. ��. The sig-. nificance of these findings are discussed and compared with.
(4) iii the features of the analogous syndromes associated with the XO anomaly in other species. It is suggested that disorders of the sex chromosomes may occur commonly in horses and may contribute significantly towards the low r ertility of the thoroughbred mare. Attention is drawn to the important practical applications of cyto genetic studies in the investigation of infertility in broodmares, and in the certification for sale of young horses intended for use as breeding stock. Cytogenetic studies of spontaneous abortions in women have established that a large proportion of pregnancy wastage in humans is due to chromosome aberrations.. The counterparts. of many of the chromosome errors initially described in humans have now been reported among domestic animals.. In. particular, the XO anomaly seems remarkably common among infertile mares.. These findings have indicated that chromo. somal abnormalities may play a significant role in the aetiology of spontaneous abortion and embryonic loss in domestic animals.. To investigate an hypothesis that chromo. some anomalies are an important cause of prenatal loss in the mare, an attempt was made to analyze, cytogenetically, a ser1es of equine abortions.. F rom a range of tissues. obtained from 26 aborted foals, cell cultures were prepared but failed to grow, and chromosome analysis was therefore not possible for any of these specimens.. It seems that the. equine foetus is retained in the uterus of the mare for a considerable period after the foetus has died and usually consists of partially-autolyzed non-vital tissues at the time of its expulsion. Consequently, a study was made of the metaphase.
(5) iv chromosomes prepared from 22 equine embryos after their surgical removal from mares' uteri. found in each specimen.. A normal karyotype was. Although inconclusive on their own,. the results of this study form an important contribution towards cumulatiye studies of embryonic loss in the horse. The current findings are discussed in the light of similar studies of induced abortions in women and embryonic loss in animals, and the potential for further investigation. in. this. field is emphasized. From the limited number of cytogenetic studies in domestic animals, it appears that chromosome analyses, particularly with the aid of more refined techniques such as centromere staining, may be of considerable value in investi gations of infertility and embryonic loss in the horse..
(6) V. ACKNOWLEDGEMENTS. This thesis is the record of a research project involving three years of study and research within the faculty of Veterinary S6ience at Massey University, Palmerston North, New Zealand.. In the course of this work. I have had the cooperation of a large number of people, for which I am most grateful. I am especially indebted to my supervisor, Professor A.N. Bruere, for providing me with the opportunity to undertake this study.. His continual guidance and enthus. iasm is greatly appreciated. F or the availability of the technical facilities required for this work and for assistance in their use, my thanks are due to the Heads and personnel of the following departments of Massey University:. Veterinary Clinical. Sciences, Veterinary Pathology and Public Health, and the Library.. Although it is not possible to personally acknow. ledge all of the people who assisted with this study, special Dr. Helen Chapman. mention must be made of the following: for her constant and invaluable advice; with the preparation of photographs;. Mr. Torn Law for aid. Messrs. Carol Black,. Cedric Barnett, Linley Fray, Bert de Cleene and Priscilla Jaine for their tireless technical assistance;. and. Mr. Mervyn Birtles for his patient tuition with the histolo gical preparations. Great enthusiasm for this study was shown by Mr. Harry Dewes; behalf.. I with to thank him for his efforts on my. I am also grateful to Professor E. D. F ieldon and. Dr. B.E. Goulden who helped in proof-reading of the thesis,.
(7) vi and to Mr. Martin Copland for his eternal comments and criticism. Finally, I must acknowledge the Mackenzie Education Foundation and in particular Mr. R.A. Mackenzie, who provided the financial grant to support this research project..
(8) vii TABLE OF CONTENTS P age PART. I. PAR T. II. PRELUDE. 2'1. MAT ER IA LS A ND METHODS. 29. II. '1 . -. PRELIMINARY ST UDY OF ABORTED FOETUSES II. 2.. 30. METHOD FOR OBTAINING EQUINE EMBRYOS. 30. i). Exper imental an imals. 30. Pregnancy diagnosis. 3'1. i i) -. II. 3.. i i i) Anaesthes ia. 3'1. iv). 32. Surgery. IN V ITRO CULTURE OF BLOOD LEUCOCYTES. 34. i). Hi story. 34. i i). Culture of cells. 34. i i i) Metaphase arrest with colch i c ine iv). 35. Treatment with hypoton i c flu id. 35. v). Fixation of cells. 36. v i). Sta ining techniques. 37. a) General procedure for m itotic c ells b) C entromere sta in ing II. 4.. 37 37. FIBROBLAST CULTUR E. 39. i). Introduction. 39·. i i). Prep aration of cell monolayer from t i s sue explant s. 40.
(9) v iii Page a) Preparation of tis sue. 40. explants b) Meth od for o btaining plasma. 41. c) Explant attachment for p rlmary culture. 41. d) Development of cell monolayer. 42. iii) Tryp s in disaggregation technique f o r cell culture. 44. a) Treatment o f embryo s. 44. b) Primary culture. 45. iv). Replen ishment o f gro wth medium. 46. v). A s ses sment of cell growth. 47. v i). Preparatio n o f cultures for subculture or harvest. 49. a) Subcultures. 49. b) Harvest of f ibroblast cultures. II . 5 .. v ii) Storage of cells. 50. SEX C HROMATIN STUDIES. 51. i). Sex chromatin of nerve tis sue. ii). Sex chromatin o f p olymorpho nuclear neutrophil s. II. 6 .. 51. 51. A NALYSIS OF RESULTS. 52. i). M icro s c opy. 52. Photographic techniques. 52. ii). ·. iii) C onstruction of karyotypes PART III. 50. 53. GESTATION A ND A BORTION IN THE MARE. 54. III . 1.. 55. LITERATURE REVIEW.
(10) ix Page i). Normal gestation. i i). Cau ses of abortion in the mare. PAR T IV. -55. 59. III . 2.. RESULTS FROM A BORTED FOETUSES. 66. III . 3.. DISCUSSION. 70. CYTOGE NETIC STUDIES IN THE HORSE IV . 1 .. 72. THE KARYOTYPE OF THE HORSE. 73. i). 73. L iterature review a) Chromo s omes of the hor se, Equus caballu s. 73. b) Chromo s omes of the donkey , Equu s asinu s i i). 78. Results and Discu s s i o n. 78. a) Chromo s ome number. 78. b) Chromo s ome morphology. 86. c) Centromere stain ing of horse chromo somes IV . 2.. 96. CYTOGENETIC STUDIES OF INFERTILE HORSE S. 99. i). 99. Literature review. a) Ster ility of equine hybrids 99 b) The chromo s omes of intersex h o rses. 101. c) Sex chromo s ome anomalies in infertile mares ii). Results:. Studies o n XO mare. 103 107. a) H i story. 110. b) Chromo s ome studies. 110. c) Examinati o n of reproductive tract. 116.
(11) X. d) Sex chromatin studies iii) Dis cus sion Origin of. xo. '140 anomaly. F requency of XO anomaly Effect of. xo. '1 40 '1 4'1. anomaly on. '146. phenotype Effect of. '1 30. xo. anomaly o n the. reproductive o rgans. '1 48. Inactivation o f X chromos omes. 149. Triple-X females. '1 5 '1. Fertility and the lack of o ne X chromo some Part V. '1 5 2. CHROMOSOME STUDIES IN ABOR TION A ND EMBRYONIC LOSS. '1 5 6. V. '1 .. '1 5 7. L ITERATURE R EVIEW i). C hromo somes and human abo rtion. '1 5 7. a) Incidence of abo rtion. '1 5 7. b) Chromo s ome studies in spo ntaneous abortions. '1 57. c) Types of c hromo s ome anomalies in abo rtions. '1 60. d) Tris omy and mono s omy in spontaneous abortions e) Polyploidy. '1 63 '1 64. ... f) Mis cellaneous chromo s ome anomalies in spontaneous abo rtions. '1 68.
(12) xi Page g ) Parental chromo s omes and sp ontaneo u s abo rtion h ) Polymorphisms. 1 68 1 69. i ) Chromo s ome anomalies in induced abo rtions ii ). iii ). Chromo s omes and embryo nic death in animals. 1 71. The chicken. 1 71. The rabbit. 1 71. Domestic animals. 172. Aetiology o f chromo s ome dis order s in abortions. V. 2 .. RESULTS AND DISCUSSION i) ii ). 1 69. 1 74 1 76. Clinical details of equine embryo s. 1 76. Chromo some studies. 1 81. Pr oblems with cell culture s. 1 84. Mo saicism. 1 85. iii ). Po s sibility o f a sire effect. iv ). Chromo some anomalies and embryonic lo s s in the horse. v). 1 86. 1 87. The aetiology of chromo some dis o rders in relation to embryonic lo s s in the horse Drugs. 1 91 1 91. Heritable chromo s o me aberrations. 1 91. Maternal age. 1 92. Aging o f gametes. 1 93.
(13) xii Page v i) Conclus ion. 1 95. APPENDICES. 1 97. BIBLIOGRAPHY. 21 1.
(14) x i ii LIST OF TA BLE S Page. T able III. 1 .. Factors assoc iated with abo rtion in the. 60. mare . III. 2 . IV. 1 .. IV. 2 .. Autopsy details of abo rtuses . � Establishment of the chromosome numbers. 68. of the horse, donkey and mule .. 76. Culture methods used for o btaining ho rse karyotyp e .. IV. 3 .. D istr ibuti o n of chromosome counts from horses with no rmal karyotypes .. IV. 4 .. 77. 82. Distr ibut i o n of chromosome c ounts from ho rses with no rmal karyotypes and at least. 30 cells c ounted . IV. 5 .. Spec ies c o mpar ison of aneuploidy from embryo c ell counts .. IV. 6 .. 1 36. Posit i o n o f the sex c hromatin body within the nuc leus .. IV. 1 2.. 1 13. Sex chromatin in nerve cells of a normal stalli o n, normal mare and XO mare .. IV. 1 1 .. 1 09. C hromosome counts of metaphase spreads from XO mare .. IV. 1 0 .. 1 08. C hromosome c ounts fro m infertile mares shown to have normal karyotypes .. IV. 9 .. 1 06. Clini c al details of infert ile mares referred for chromosome analysis .. IV. 8 .. 84. Clinical details o f mares with sex chromo some disorders .. IV. 7 .. 83. 1 36. C o mparison of sex chromosome disorders among i nfertile mares and women .. 1 42.
(15) xiv Page. Table IV . 13 .. Spec ies character i st i c s a s s o c iated with sex chromo s ome dis orders .. V.. 1.. Chromo s o me anomalies among human spontan eous abo rt i o ns .. V.. 2.. 1 50. 1 60. Summary of chromo some studies in human. ;. spontaneous abo rti ons.. 1 62. V.. 3.. Chromo s ome studies in induced abo rtions .. 1 70. V.. 4.. Clini cal details of equine embryo s .. 1 78. V.. 5.. Chromo s ome studies of equine embryo s .. 1 82.
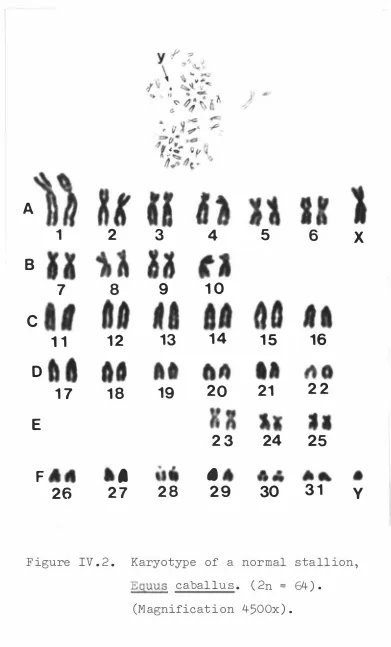
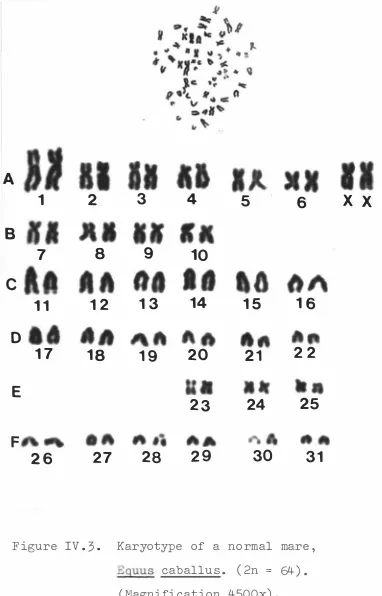
(16) XV. LIST OF F IGURES Page PART I I .. 1.. Surgical removal of an e quine embryo from the ut erus of a mare .. 2.. F low diagram of leuco cyt e cu lture t e chnique from horse b lood .. 3.. 43. F ibrob last c e l l culture which has f o rmed a comp l e t e mono layer .. 5.. 38. Primary c e l l culture showing out growth of c e l l s from a t i s sue exp lant .. 4.. 33. 48. Mono layer c e l l culture aft er 16 hours expo sure to c o lchic ine .. 48. 69. PART I I I .. 1.. G e st at ional age s of abortu se s .. PART IV .. 1.. Karyotype of a t etrap l o id c e l l in a c e ll culture from an e quine embryo .. 85. 2.. Karyotype of a normal stal l i on.. 88. 3.. Karyotype of a normal mare.. 89. 4.. Karyotype o f a normal stall ion showing characterist ic s e condary c onstrict ions on acroc entric chromo s ome s .. 5.. 91. Pre sumed s e condary c onstrict ions on the s hort arms of a large met ac e ntric chromosome .. 6.. 92. Pre sumptive chromo somal translo c at ion invo lving a large met acentric chromo s ome from a normal mare .. 7.. 94. Karyotype of a c e l l from an e qu ine embryo showing pre sumpt ive premature c hromat id · separat ion and a s s o c iat ion o f t wo homo logous met acentric chromo s o me s .. 95.
(17) xvi Page 8.. Karyotype from a normal stall ion aft e r c entromere st aining .. 9.. 97. Charact eris t i c app earance of the sex chromo s ome s of the horse after c e ntromere staining .. �0.. 98. Phenotype o f thoroughbred mare shown t o have XO anomaly .. �� -. Karyo type of mare shown to have XO anomaly .. �2.. ��4. Karyotype of 63XO mare aft er centromere st aining .. ��5. �3 -. Int ernal genit alia of. xo. mare .. �4.. Int ernal ge n it alia of. xo. mare ; vagina ,. cervix and ut erus op ened . �5.. Left ovary o f XO mare .. �6.. H i s t o logical s e ct ion o f endom etrium from XO mare .. � 7-. ��7. ��8. � 2�. Pre sump t ive at ret ic Graaf ian f o l l i c l e s ( a and b ) and lut eal t i s sue ( c ) in the right ovary o f the XO mare .. �8.. � 22. Pre sumpt ive Graafian f o l l i c le s in the right ovary of the XO mare .. � 22. �9.. Wall o f pre sump t ive Graaf ian f o llic le� 23. 20 .. Atre t i c or degenerat ing Graaf ian f o l l ic l e .. 2� .. D i s c r e t e are a of lut e a l t i s sue in the right ovary of the XO mare .. 22 .. � 24. � 27. Typ ic a l hist o l ogical appe aranc e of funct ional lut eal t i s sue from right ovary o f XO mare .. � 27.
(18) xvi i Page 23 .. Charact erist ic act ive lut e a l endoc r ine c e lls .. 24 .. 1 29. Sex chromat in body ad jacent to the nuc l e o lus in a neurone from a normal mare .. 25 .. 1 37. Sex chromat in body free in the nuc l e op lasm of a neurone from a normal mare .. 26 .. 1 38. Sex chromat in body adj ac ent t o the nuc l e ar membrane in a neurone from a normal mare .. 27 .. 1 38. Charact eri s t i c nu c l e ar app endage ( " drumst ick " ) on a po lymorphonuc l e ar neutrophil from a normal mare .. 28 .. 1 39. Nuc l e ar app endage on a po lymo rpho nuc l e ar neutrophil from a nonua l s t a l l ion .. PART V .. 1.. Po s s ib l e me chani sms for the origin of t r ip lo idy .. 2.. 1 80. E qu ine embryo with we l l deve loped amnion and umbi l ical art eries .. 6.. 1 79. N o rmal horse embryo with membrane s removed .. 5.. 1 79. Normal horse embryo showing remnant of yo l k sac and the s inu s t e rminal i s .. 4.. 1 67. Norm a l horse embryo ( E . cabal lus ) with in intact all ant o chori o n .. 3.. 1 39. 1 80. E qu ine twin embryo o f fewer than 20 d ay s ge s t at ion .. 1 83.
(19) xvi ii LIST OF APPENDICES App endix I.. Page Paras ite c ontro l o f experiment al animal s .. II .. · Drugs used for anae sthe sia and surgery .. III .. Mat erial s re quired for blood leuco cyt e. 1 98 1 98. culture .. 1 99. a). Preparat ion o f culture .. 1 99. b). Harve st ing of cultures .. 200. c). To mount slide s .. 200. IV .. Preparat ion o f G iemsa stain .. 201. V.. Procedure for cent romere staining .. 201. VI .. Preparat ion o f glas sware .. 202. VII .. Mat erial s re quired for fibroblast culture :. V II I .. Plasma c lot emb e dding. t e chn ique .. 202. a). To obt ain t is su e from l ive horse s .. 202. b). Growth medium .. 202. c). To prepare tric ine buffer .. 203. d). To prepare pla sma c lot .. 203. e). A . T . V . ( Ant ib io t i c -tryp s in-versene solut ion ) .. 203. f). PBS . ( Pho sphat e -buffered sal ine ) .. 204. g). General e quipment .. 204. Mat erials re quired for f ibroblast culture:. Tryp sin disaggregat io n. t echnique .. 205. a). Transport medium .. 205. b). Tryp s in s o lut ion .. 205. c). A nt ib iot ic s o lutio n .. 205. d). Co lchicine .. 205. e). Long t erm storage of c e l l s .. 205.
(20) XlX. AJ2J2endix. Page f). IX .. X.. 206. General equipment .. Analysis of re sult s .. 206 ... Examinat ion of s lide s .. b). Deve lopment o f f ilm .. 206. c). Deve l opment of print s .. 206. Chromo s o me c ount s of hors e s prep ared from leuc o cyt e cultures .. XI .. 207. Chromo some c ount s of hors e s prep ared - by f ibroblast culture s of t i s sue e xp lant s .. XII .. 206. a). 208. Chromo some c ount s prepared by leucocyt e culture f rom hors e s with normal karyo type s .. C as e s with fewer than 30 c e l l s. are exc luded . XIII .. 209. Chrom o s ome count s from fibroblast c e l l culture s , exc luding c a s e s with f eweE than 30 c e l l s c ount e d . a). 210. Chrom o s ome c ount s from hors e s with norma l karyotyp e s prepared by c e l l culture s o f t i s sue exp lant s .. b). 210. Chromo some c ount s f rom e quine embryo s with normal karyotype s .. 210.
(21) 20. PART I. PRELUDE.
(22) 21 PART I. PRELUDE. Spont aneous ab ort ion h as b e e n defined as th e natural t erminat ion of a pre gnancy b efore th e f o etus 1s viab l e ( Mo r ison , 1 970 ) .. In th e h o r s e th i s inc lude s f o al s born. b efore 290 days of ge st at ion (Ros sdal e , 1 965 ;. P latt , 1 973a) .. E ve n t oday , l ittle i s known o f th e cau s e s o f prena t al l o s s i n dome st i c animals ( Hanley , 1 961 ) alth ough it 1 s r e spons ib l e f o r a serious l o s s o f p o t ent ial product ivit y . A prenat al de ath rat e of about 40% h a s b e en e st ab l i sh e d for th e rat , rabb it and ferret ( C orner , 1 923 ) and s imilar value s of 35 -40% and 25% are quot ed for th e pig ( Smith and Marlowe , 1 97 1 ) and cow ( Hanley , 1 961 ) re sp e ct ive ly . I n th e se spe c ie s prenat al lo s s a c c ount s for at least one th ird of th e t otal c onc eptions and th e actual f igure may b e e ve n great er wh en h ith erto undet e c t ed e arly embryonic lo s s e s are inc lude d .. From s l aughter ob servat ions , Qu inl ivan ,. M art in , T aylor and C airney ( 1 966 ) report e d a prenat al lo s s o f 23% in sh e ep , with th e gre at e st mortality o c curring during th e f irst 30 days of pre gnancy . Th e mare apparent ly h a s th e lowe st reproduct ive rat e o f a l l th e dome stic. animals and reproduct ive effic iency , as m e a sured by live foal p e rcent age , is lower in th oroughbreds and standardbreds than in oth er breeds managed l e s s int en s iv e ly (Burkh ardt , 1 948 ;. Hut ton and M e ach am , 1 968) .. In th e Unit ed St at e s , Bergin ( 1 970 ) re c orded a l ive f o a l perc e nt age in thoroughbreds o f 55 . 3% b etwe en 1 950 and 1 95 6 .. By 1 970 th i s f i gure h ad fa llen t o 50% .. We ath erby' s. s t at ist i c a·l dat a repo rt e d a 50% l ive f o al rat e f or th e.
(23) 22 Unit ed Kingdom and E ire in 1 973 , wh ile in Austral ia , O sb orne ( 1 975 ) def ined nat ional foaling rat e s of 49% in 1 950 and 47% in 1 97 3, and not ed an increas ing dispar ity b etwe en c on c ept ion and foaling rat e s in rec ent years .. In New. Z e aland , D ewe s ( 1 973 ) found th at a foal ing perc e ntage of 67% in 1 903 h ad fallen t o 49% in 1 971 .. An even lower. f o a l ing r at e of only 45% was recorded by van N ie kerk ( 1 965a ) i n South Africa between 1 962 and 1 965 . O sb orne ( 1 975 ) suggested th at th e gravity of th e s ituat ion may h ave b e en exaggerat ed as th e s e e st imat e s of f ert ility were ba s e d on th e numb er of th oroughbred mare s . r e g i st ered in th e stud-bo oks of th e various countries rath e r th an o n th e numb er o f mare s actual ly serve d .. More ob j e c t ive. dat a c an be obt ained from surveys of mat ing group s . A thr e e ye ar survey of two large th oroughbred studs 1n New Z ea l and disc lo sed that of all mare s mat e d , only 54% produc ed a viable foal (Ell iott , C allagh an and Smith , 1 97 1 ) . A s imilar f igure was released by the New Z e aland Th orough b r e d Breeders' A s s o c iat ion ( Anon . , 1 970 ) f o r th e number o f l ive f o a l s re sult ing from mat ings by e ight commercially p opular s t a l l ions in 1 966 .. In a group of 85 2 th oroughbred. mare s , Laing and Le e ch ( 1 975 ) found th at of th e 77 . 9% wh ich c onc e ive d , 69 . 7% produced a viabl e f o al .. Pro duct ive mare s. again const ituted only 5 1% o f th e total numb er o f mare s surveyed .. In addit ion , the s e f igure s att ach e d ma jor sig. nif icanc e to th e h igh inc idenc e ( 30 . 3% ) of prenat al lo s s , of wh ich only a small proport ion was manif e st e d as c l inical abort ion . E st imat e s of total prenat al l o s s in th e horse h ave b e en confus e d due t o problems of measuring th e conc ept ion rate .. In mo st stud i e s conc ept ion rat e has b e e n det ermined.
(24) 23 by r e ctal palpat ion b et ween 35 and 45 days aft er serv i c e ( Su l l ivan , Turner , S e l f , Gutt eridge and Bartlett , 1 975 ) . Much controversy e x i s t s ove r th e role that manual pregnancy t e st ing may play in embryonic lo s s .. V o s s , P ickett and Back. ( 1 97 3 ) e st ablish e d a de leterious effect on conc ept ion rat e o f rectal palpat ion during o e strus , and O sb orne ( 1 975 ) inc riminat ed rectal p alpat ion during e arly pregnancy as a c au s e of embryonic l o s s .. Oth er workers (Allen , 1 974a ;. I rwin , 1 975 ) have f ound early manual pregnancy t e s t ing t o h ave no effect o n th e inc idenc e o f fo etal de ath . Under current systems of int ens ive stud management th e total c oncept ion rat e o f thoroughbred mare s is prob ably b e t ween 65% and 70% ( M cG e e , 1 970 ;. Laing and L e e ch , 1 975 ) .. Sul l ivan et al . , ( 1 975 ) rec orded a f irst s e rvice c onc ept ion r at e of 43% , a cumu l at ive pre gnancy rate o f 77% and a preg nancy loss of 1 2% aft e r 45 days .. F igure s released by th e. N e w Z e aland Racing C onferenc e reveal an annual abort ion rat e o f 1 3% , s imilar t o that report ed from oth e r studie s ( Day , 1 95 7 ;. Matassino , 1 962 ;. McGe e , 1 970 ) , and a ?0% inc idence. of so -called 'barren' mare s .. A proportion of th e lat t e r h ad. almo st c ert ainly undergone embryonic lo ss . Th e most rel i ab l e method o f determining total pre nat al l o s s is by p o s t -mort em examinat ion of mar e s at various int erval s aft er s ervic e .. In 1 920 , a survey in Scot land. ( C o rner , 1 923 ) of 28 , 000 mat ings in C lyde sdale s and 3 , 640 m at ings in th oroughbred hor s e s revealed prenat al death r at e s of 48% and 5 9% r e spect ively .. A comp arab le re sult was. o bt ained by O sborne ( 1 975 ) f o llowing th e insp e ct ion o f 5 1 98 ut eri .. Sh e det e c t e d p o s it ive evidenc e of abort ion in 5 0 . 3%. o f 765 ut eri wh ich sho wed e videnc e o f pregnancy .. In th e. s tudy by O sborne ( 1 975 ) , a proport ion of th e l o s s was attri-.
(25) 24 but e d to incre ased stre s s factors , namely unsymp ath etic h andl ing , long dist ance transport and inade quat e fo· o d and wat er . F rom a study of prenat al l o s s in th e p ig , Corner ( 1 923 ) discovered an init ial lo s s of 20% of ova before th e f ourth week of pregnancy .. Hanley ( 1 961 ) report ed that 3 3%. o f p ig embryo s die during th e first h alf of g e s t at ion . Dufour and F ahmy ( 1 975 ) cons idered embryonic mort ality t o b e th e most import ant f act or det ermining l itter s iz e in d iff erent breeds of pig s .. Th ey found that 1 9% of ova. re l e ased from th e ovary were l o st during th e f irst 23 day s o f pregnancy , wh ile l o s s e s aft er th is stage were very few . I n h is inve st igat ion of rep e at -breeder cows , def ined as tho s e cows not conc e iving to f irst service , C a s ida ( 1 95 3 ) f ound th at 59 . 4% of pre sumab ly normal embryos were lost during th e first 34 days of ge station .. Obvious ly early. e mbryoni c death was a major limit ing f actor in th e reprod u c t ive p erformance of these animals . F rom cumulat ive studies in humans , R o c k ( 1 940) con c luded that a� abo rt ion rate of 25% was a c onservat ive e st imat e , and it was later sh own (Hert ig , R o c k , Adams and M enkin , 1 959) that th e gre at e st l o s s of ova o ccurred dur ing pre imp l ant at ion s t ages of pre gnancy .. Foll owing th e examin. a t ion of t en abnormal human conceptu s e s obt ained by hyst er e ct omy , Hert ig , R o ck and Adams ( 1 95 6 ) doubted wh e th er th e more abnormal conceptu s e s would h ave deve loped t o a stage s uf f i c i e nt to cau s e c l inical evidenc e of pregnancy .. Com. b in ing th e fre quency of c l inical abo rt ions ( 1 5% ) with a 3 0% lo s s of pre imp lant at ion stage s , C arr ( 1 970a) c alculat ed that prenatal de ath acc ount ed for 45% of all c oncept ions.
(26) 25 in wome n . A s i n oth er sp e c ie s , detected abort ions in th e h orse repre sent only a small f ract ion of total prenat al lo ss , mo st of wh ich o c curs dur ing e arly st age s of pre gnancy ( Moberg , 1 975 ) .. The ext e�� o f th e prob lem of e arly embryonic lo s s. in mare s was emph as iz ed by Du P l e s s is' ( 1 964) d i s c overy th at 3 8% of mare s wh ich did not return to o e strus aft er s e rvice were found to be barren wh en examined for pregnancy aft er 40 days . · In ye arl ing mare s Mitch e l l and A llen ( 1 975 ) report e d a h igh inc idenc e ( 46% ) of e arly pregnancy l o s s e ve nly d istribut e d between 60 and 140 days ;. l o s s e s earli e r. th an 60 days of g e s t at ion were unknown but lo s s e s lat er th an 1 40 day s were very f e w . Th e c ause s o f embryonic l o s s in dome st ic animals h ave remained large ly unexplained . h a s oft en b e en emp ir i c a l . f ert ile c ows :. C onsequent ly , treatment. In 1 953 C a s ida wrot e of sub. "M any different treatment s are att empt ed in. g e tt ing rep eat bre e ders to conc e ive .. Almo st any treatment. app e ars t o do some good , but none appe ars t o do a gre at de al of good . " Inve st igat ion of th e aet io logy of embryonic lo s s in th e h o r s e has b e e n d ominat e d by the f ie lds of inf e c t ion and e ndo crino logy .. Th i s emphasis is now b e ing que st ioned .. I nf e ct ious agent s were shown t o b e of l itt l e or no imp ortanc e in th e aet io logy of e arly f o etal lo s s in yearling mare s ( M it ch e l l and Allen , 1 975 ) .. A lth ough inf e c t ion o f. th e foetus in ut e ro may b e a s ignif i c ant c ause of prenat al mort ality in l at e r month s of pregnancy , P l att ( 1 973b , 1 975 ) c o n c luded that inf e ct ion was prob ably s e ldom th e primary c au s e o f abort ion .. T o exp lain th e divergence b etween c o n-. c ept ion and f o al ing rat e s , Osborne ( 1 975 ) drew att ent ion.
(27) 26 t o th e increa s ing stre s s e s of mod ern int ens ive stud manage ment .. Other factors incriminated in prena t al loss h ave. inc luded hormone malfunction , immaturity and inade quat e nutr it ion . Much of th e enthus iasm for various tre atment s , p ar t ic ularly hormone th erapy , h as derived from c l inical exp e rienc e s .. In th i s c ont ext it has b e en shown ( M alp as ,. 1 93 8 ) th at 62% of women wh o h ad abort ed twice c arried th e th ird baby to term with out sp e c if ic treatment .. Gan j am ,. Kenney and F lickinger ( 1 975 a ) h ave que st ioned th e effic acy of e xogenous proge st erone in the tre atment of ab ort ion and c o n c lude d that " th e wh o l e prob lem of h ab itual abort ion and it s tre atment in mare s needs re-evaluat ion . " F or more than 50 years it h as b e en known that many abo rt ed conceptu s e s were structurally abnormal .. Environ. ment al f actors were b e l ieved to be primarily re sponsib l e for congenit al defe c t s in th e develop ing embryo unt il abnormal it ies we re d i s c overed in pre imp lant at ion stage s of p ig embryo s ( C orner , 1 923 ) . f inding Corner ( 1 923 ) wrote :. In a s s o c iat ion with th i s " it se ems that in th e p ig. th e r e i s litt le e vide nc e th at faulty imp lantat ion is a c au s e o f e mbryonic mort ality and abnormality , but much to ind i c at e th at embryo s may b e c ome abnormal in sp it e of a ut erine e nv ironment wh ich is by all th e crit eria at our present c ommand , anatomic ally and funct ional ly normal . ". Anomal i e s. of deve lopment were pre s ent at a l l st ag e s o f pregnancy and h e also ob served de f e ct ive and normal embryos in th e same ut e rus .. Corner ( 1 923 ) attribut e d at le ast part of th e. e mb ryonic mortality t o leth al factors with in th e germ c e ll s and embryo s .. Similarly , abnormalit i e s in e arly monkey.
(28) 27 e mb ryo s were thought to have originat ed from c onst itut ional de f e ct s of the embryo it s e lf ( Corner and Bart e lme z , 1 953) . Hert" ig and Rock ( 1 949) studied 28 e arly human con c eptuse s obt aine d by hyst erectomy and dec ided that 25% of them had been de s t ined to abort . t r ium was normal .. In all c as e s the endome. In a sub se quent s tudy ( H ert ig et al . , 1 95 6). �4 zygot e s of fewer than 1 7 days g e s t at ion we re re covered from the ut eri of women of proven f ert ility .. T en conceptu s e s. were abnormal , o f which four would probab ly never have i mp lant ed . The ide a that chromo some disorders might be. a. s ign i. f ic ant c ause of spont ane ous abort ion in humans re ceived supp ort with the f inding of two chi ldren with lethal chromo some const itut ions (Edwards , Harnden , C ame ron , Cro sse and W o lff , 1 960 ;. P at au , Smith , Therman , Inhorn and Wagner ,. 1 960) and short ly aft er , two inst anc e s of foetal death were found to be a s s o c iat ed with triplo idy of the fo etus ( Penro s e and De lhanty , 1 961 ;. De lhanty , E l l i s and Rowley , 1 961) .. The f irst report s of chromo s o me studie s of spont an e ou s abort ions in women reve aled a high inc idence of c hromo s ome aberrat ions ( C arr , 1 963 B enirs chke , 1 963) .. C lendenin and. M any sub s e quent studies have e st�b l i shed. the inc idenc e of chromosome anomal ie s in spontane ous abor t ions at approximat ely 36% ( C arr , 1 970a) , a frequency r emarkab ly s imilar to that found in t en day p ig blast o cy s t s ( McF e e ly , 1 967) . A lmo st 1% of liveborn infant s have a major chromo s omal abnormality (Friedrich and Nie l sen , 1 973 ;. Hamert on ,. C anning , R ay and Smith; 1 975 ) but this repre sent s only a small proport ion of all conceptuse s with chromo some.
(29) 28 abnormal it i e s .. The inc idenc e of chromo some anomal i e s f ound. in conc eptu s e s de cre as e s progr e s s ive ly from conc ept ion unt il t e rm ( C arr , 1 970a ;. Mach in , 1 97�) .. A variety o f chromo s ome aberrat ions are now known t o be assoc iat ed with embryonic death and pre nat al l o s s in th e pig (McF e e ly , 1 967 ) and cat ( B enirs ch ke , Edwards and Low , 1 97� ) and anomal i e s of th e sex chromo some s h ave b e en found �. in infert il e mare s .. Studie s of h ab itual abort ion in women. h ave sugge sted th at p arental chromo some anomalie s may p l ay a role in th e aet io logy of re current abort ion and one c a s e o f a chromo somal trans locat ion in a b o ar h as b e en l inke d · with embryonic de ath ( A ke s s on and Henricson , 1 972) . Thus human studie s h ave revealed th e importanc e of chromo s omal errors in infert i l ity and abort ion and nume rous , example s of chromo s o me anoma l i e s in the dome s t i c animal s are now known .. Due t o th e limit e d amount of r e s earch undert aken. in th is f i e ld , th e s ignif icance of ch romo some d i sorders in embryonic l o s s h a s not yet b e e n e st ab l i sh e d for th e dom e st ic animals .. Howeve r , th e pre l iminary report s out l ined above. h ave demonstrat e d remarkab l e s imilarit ies to th e s ituat ion in man ..
(30) 29. PART II. MATERIALS AND METHODS.
(31) 30 PART II. MATER IALS AND METHODS II . � .. PRELIMINARY STUDY OF ABORTED FOETUSES -.. A pre liminary study of spont ane ously abort ed e quine f o etu s e s was undert aken on c as e s re f e rred by f ie l d vet erinarians .. Twenty- s ix abortus e s with g e st at ional age s. b etwe e n f ive month s and t erm were invo lved in th i s study . Wh en th e abortus was availab l e a rout ine autopsy was p e rforme d , supp lement ed by b ac t eri o l ogical and h istological inve s t igat ion of body organs , inc luding th e sp le e n , brain , lung , lymph node s , p lacent a , kidney , l iver and adrenal g lands . F rom th e foetus , a s amp le of subcut ane ous f as c ia was c o lle c t e d into a universal bottle c o ntaining HBSS (Hanks ' B alanced Salt So lut i on ;. Hanks , � 948 ) , and c e l l culture s. were prepared ac cording t o th e meth od des cribed by Hyman ( '1 968 ) . II. 2 . i). METHOD FOR OBTAINING EQUINE EMBRYOS. Exper iment al animals A group of � 2 mare s and one st allion was graz ed on a. 3 0 acre gras sed p addock.from O ctober unt il March .. During. th e int e rv ening s ix month s th ey were confined t o a thre e acre f ie ld and fed h ay .. E ach hors e was dre nch ed regu l arly. with 60g E qui z o le and �5g N e guvon at two month ly interval s . T reatment f or l i c e in wint er and spr ing invo lved dust ing e ach h o r s e twice at �4 day int erva l s with rot enone p o wder (App endix . I ) . Due to graz ing m·anagement o b s e rvat ion of th e mat ing.
(32) 31 group during th e bre eding s e as on was infre quent and it was not p o s s ib l e to obt ain ac c·u rat e mat ing dat e s for individual mare s .. D i agno s i s of pregnancy·therefore re lie d on re ct al. p alp at ion . ii). Pregnancy d�agno s i s D iagno sis of e arly pre gnancy b y rectal palpat ion. invo lved th e det e ct ion of ch ange s in tone and th ickne s s of th e wall s o f the ut erine h orns and palpat ion of an embryonic mas s .. V an N ieke rk ' s ( 1 965 a , 1 965 c ) c laim th at it is. " p o s sib l e to make a diagno s i s of pregnancy at 14 days and a def init e diagno s i s o f pregnancy 1 6 t o 1 9 days aft er ovu l at ion" was disputed by B ain ( 1 967a) wh o was unab le t o give a nega t ive and often even a p o s it ive diagno s i s o f pre gnancy at f ewer th an 25 day s .. Bain ( 1 967a) considered 30 days after. ovu lat i o n th e ear l i e s t pract ical t ime t o pres ent mare s for pre gnancy diagno s i s . Th e reproduct ive tract of e ach mare was examined p e r re ctum a t int ervals of one we ek through out th e breeding s e ason .. P art icu l ar att ent ion was p a id t o th e s t at e of the. ovarie s and ut eru s .. In view of Bain ' s ( 1 967a) c onclus ion ,. mare s were not pre s ent ed for surgery unl e s s a pregnancy of at least 30 days durat ion h ad been e st ab l i sh e d .. A p o s it ive. diagno s is of pre gnancy was made at approximat e ly 30 day s g e s t at io n on the bas i s of ut erine t one and th e presenc e o f an emb ryonic swe l l ing .. Th i s was c onf irmed immediat e ly prior. to surgery . iii). Anae sth e s i a E a ch mare was premedic ated with a tranquilliz ing drug. and th iopent one s odium was used to induce anae sth e s i a with in a p added stall .. Th is was f o llowe d by intub at ion , and m a ln-.
(33) 32 t e nanc e of anae sth e s ia with oxygen and h aloth ane supp lied through. �. semi- c l o s e d t o and fro anae sthet ic apparatus. inco rporat ing a soda lime c annister to remove c arbon dioxide . iv). Surgery Th e patient was p o s it ioned on th e adjustab le oper. at ing tab l e in dors a l re cumb ancy with th e h ind legs pas s iv e ly ext e nded , and th e operat ion s it e was rout ine ly prepared f o r surgery . An inc is ion 1 5 c m in length was made through th e l ine a alb a in th e ventral midline betwe en th e umbilicus and th e mammary gland .. Th e ut eru s wa s locat e d and th e pregnant. h orn ext erioriz ed .. Unt il th e uterine inc i s ion was comp l e t e d. the embryo was t emp orarily exc luded from th e s it e o f inc i s ion i n th e ut e r ine h orn b y manual pre s sure from out s ide th e ut erus .. St ill e n c l o s e d with in it s int act ch orio. allant oic membrane the embryo was expr e s s e d through th e inc i s ion and trans f e rred to a sterile glas s cont ainer ( F igure I I . 1 ) . St andard surg ical t e chnique s were e mployed to c l o s e th e w-ound .. Separat e c l o sure o f th e perit oneum was cons id. ered unnec e s sary and it was u sually inc luded in a s ingl e layer of suture s with th e l in e a alb a . Tet anus ant it oxin was administ e red to th e patient aft e r each operat ion but no addit ional postoperat ive tre at ment was given .. Th e pat ient was returned t o th e mat ing. -group thr e e days aft er surgery and th e skin suture s were removed a week- lat e r ..
(34) 33. F igure I I . 1 .. Surgical remo va l o f an equine embryo from the uterus o f a mare ..
(35) 34 II. 3 . i). IN VITRO CULTURE OF BLOOD LEUCOCYTES. Hist o ry In 1 960 , Moorh e ad , N o we l l , M e l lman , B at ipps and. Hungerf ord introduced a pro c e dure for th e prep arat ion o f human leuco cyt e s for chromo s ome analy s is .. Th i s involved th e. sho rt t erm c e l l culture of 1 0 ml of venous b l o o d .. Modif i. cat ions o f th e basic t e chnique (Edwards and Y oung , 1 961 ; Edwards , 1 962 ;. F r� land , 1 962 ) were de s igned t o separat e. out th e l euc ocyt e s and redu c e th e vo lume of b l o o d required , and Arakaki and Sparkes ( 1 963 ) obt ained sat isfact ory chromo some preparations from blood vo lume s a s smal l a s 0 . 05 ml . 'Th e h orse lends it se lf t o leuco cyte culture more re adily th an do th e other dome s t ic animals due t o the rap id spontane ous separation of leu c o cyt e s and erythro cytes wh en e quine b l o o d is allowed to st and .. Payne , E l l sworth and. DeGroot ( 1 968 ) u t ilised th i s p roperty in th e d e ve lopment of a s imp le t e chnique for th e sh o rt- t e rm culture of e quine lymph o cyt e s .. In th is study l eu co cyt e culture s t o det erm ine. th e mit o t i c karyotype of l ive h o rs e s were pre p ared by a modif i c at io n of th e latter pro c edure .. The c omp lete leuc o. cyt e culture t e chnique used f o r th e h orse i s sho wn in th e flow diagram , F igure II . 2 .. Details of th e mat erials used. are given in Appendix I I I . ii). Culture o f c e l l s F our 1 0 ml blood s amp l e s were c o lle c t ed asept ic a l ly. from th e jugular ve in int o an h epariniz ed vacut ainer . F o r t ranspo rt t o th e laborat ory th e vacut ainer w a s packed with i c e ins ide an insulat ed flask . E a ch s amp l e was allowed t o st and vert ic ally f or 20 m inut e s at room t emp erature ... Th e e rythro cyt e s s ediment ed.
(36) 35 quickly wh ile th e leucocyt e s lagged b eh ind and formed a " buffy c o at " .. Two ml of th e l eucocyt e-rich p lasma were. transfe rred to a universal container to wh ich was added s ix ml of medium 1 99 , 0 . 2 ml phyt oh aemagglut inin and suff i c i ent s odium b ic arbonat e ( 1 0% ) to ad just th e pH t o neutrality . E ach culture was incub ated at 37° C for 48 to 72 h ours . iii). M e t aphase arre st with co lch i c ine A lt e rat ions in th e pH of th e culture were visib l e as. a ch ange in colour due t o th e indic ator , ph e no l red , wh ich was inc luded in th e growth media . �. Rap id metab o lism of c e ll s. re sult e d in a f a l l in pH and a colour ch ange from red t o ye llow .. Culture s judged t o contain act ive ly dividing c e ll s. were t r e at e d with co lch ic ine aft er 4 8 hours incubat ion wh ile colch i c ine was not inoculat ed int o s lower growing cultur e s unt il 72 h ours of incubat ion h ad e l ap s e d .. A f t e r th e. addit ion of c o l ch ic ine ( 0 . 1 �g in 0 . 1 ml) t o e ach cultur e , incub at ion was c ont inued for a furth er 1 6 h ours . D ifferent animals and t issu e s vary in th e ir re sp onse to co lch ic ine , and th e durat ion of treatment and conc ent rat ion of co lchic ine must vary according to exp erienc e with individual t i s su e s .. Exc e s s ive contract ion o f chromo some s ·. h as b e e n noted with colch i c ine tre atment at h igh conc ent rat ions ( Hyman , 1 968 ) and for prolonged periods ( Moorhe ad e t al . , 1 960 ) .. In th is study sat i sf actory re sult s were. ach ieved with c o l chicine tre atment over a c on s iderab l e int erval but a t very dilut e conc e ntrat ions . iv). T r e atment with hyPo t onic fluid Hyp otonic fluid tre atment of c e ll suspensions to. spre ad met apha s e chromo some s was introduced b y H su ( 1 95 2 ) . Th e di s covery o f th is effect was the result o f an acc ident.
(37) 36 wh e reby culture s were wash e d in hypot onic Tyrode ' s solution instead of isotonic saline b efore f ixat ion .. M any solut ions. are now known to produc e a c omparab l e effect ( M akino and Nish imura , 1 952 ;. H su and P omerat , 1 953 ;. Hyman , 1 968 ) .. Rupture o� c e l l membrane s may be a .cons e quenc e of . pro longed e xp o sure of c e ll s t o hypot onic so lut ion and th ere is a t e ndency f o r chromo some out line s to be come b lurred ( T j io and Le van , 1 956) .. Th e a im is to abbreviat e-hypot onic. treatment t o a m inimum in · th e attempt to induc e chromo some sc attering with out th e unf avourabl e effect s on th e chromo some surface . Th e auth o r u s e d a 0 . 075 mo lar solut ion of potas s ium ch loride ( Hungerf ord , 1 965 ) f o r hypot onic tre atment . Ade quat e spread and defin it ion of met aph ase chromo some s were obt ained after incub at ing c e l l s in hyp o t onic solut ion for 20 minut e s . v). F ixat ion of c e l l s Th e qual ity o f met aph a s e p l at e s is inf luenced b y the. method and durat ion of f ixat ion .. Rothfel s and Siminovit ch. ( 1 958) found th at c lump ing of chromo some s could b e reduc ed by the s l ow addit ion of f ixat ive and delayed breaking up of th e c e l l depo s it .. Th is c an a l so b e achieved by th e f ixat ion. of c e l l s suspend e d in c ons ider ab l e amount s of hypotonic f luid ( Axe lrad and McCul lough , 1 95 8 ) .. Ford and H amert on. ( 1 956) report e d that th e qual ity of f ixat ion was sl ight ly improved by us ing ch illed f ixat ive . In th e pre sent study f ixat ion of c e l l s f o r 60 minut e s a t 4° C produced good qua l ity met aph ase chromo some s . Ce l l s in susp ens ion we re f ixed undisturb e d for 60 minut e s at 4° C in a fre sh ly prep ar e d mixture of methyl alcoh ol and glac ial ac et ic ac id ( 3 : 1 ) .. Spreading of.
(38) 37 chromo s ome s was fac ilitated by dropping th e c e l l suspens ion from a h e igh t of 20 cm on to c le an s l ide s covered by an even f ilm of ch illed distilled wat e r .. Slide s were th en allowed. t o air dry b efore e ach s l ide was labe lled with a serial number , c a s e numb er and th e y e ar . vi). St a ining t e chniques a) General procedure for mitot ic c e l l s P re p ared dry slide s were dippe d in ab s o lut e alcoh o l. and st ained with 2% Giemsa f o r 10 minut e s , rin s e d under t ap wat er and allowed t o air dry ( Appendix IV ) . P rior t o staining , a pre cursory examinat ion of th e s l ide s was made with a ph ase c ont rast micr o s c o p e .. Th o s e. s l ide s c ons idered suitable for chromo some analy s i s were st aine d , s o aked in xylol for f ive m inut e s , and a coverslip was aff ixed us ing DeP ex . b ) C e ntromere staining S in c e th e init ial ob s e rvat ions of th e diff erent ial st aining of th e c entromeric h e t erochromatin of mitot ic chromo s ome s from mice aft er treatment with sodium hydroxide and G ie m s a stain ( P ardue and Gall , 1 970 ;. Yunis , Ro ldan ,. Y asmineh and Le e , 1 971 ) , c e nt romeric st aining a s a means of chromo s ome ident ification h as been re cogni z ed in several spe c i e s inc luding man ( Ch e n and Ruddl e , 1 97 1 ) , th e mou s e ( H su , C ooper , Mace and Brinkley , 1 97 1 ) , o x ( H ansen , 1 973 ) and th e g o at and she ep (Evans , Buckland and Sumner , 1 973 ; B ruere , Z artman and Chapman , 1 974) . The meth od de scribed by Ch andley and F le t cher ( 1 97 3 ) was u s e d for c ent romeric s t aining of mit ot ic ch romo some s in th i s study .. Th i s procedure was in turn b a s ed on that of. Sumner ( 1 972) with th e variat ion th at incub at ion with Ba( OH ) 2 l ast e d 1 0 minut e s rather th an 10 s e c onds ( App e ndix V ) ..
(39) 38 F I G U R E 11. 2 .. F LOW D I AG RAM O F LE UCOCYT E CU LT U R E T EC H N I Q U E F R OM HO RSE B LOOD.. Hepari nized whole b lood ( 1 Oml). Cel l culture media Medium 1 99. �. Stand vertically at room temperature for 1 5-30 minutes. Phytohaemagglutin. +. Leucocyte- rich plasma withdrawn. ~. 6 ml. 2ml. "'-..I/�. Sod ium bicarbonate ( 1 0%) Na H C03. U N I V E RSAL. 37° for Aqua-Colch in 1 O.l�g 37°C for. �. t �. 72 hours Total i ncubation time 88 hours. Universal jars in i ncubator.. 1 6 hours. Transfer cu ltures to centrifuge tubes. �. Spin 5 minutes at 1 000 r. p. m. Discard supernatant. .------. �. Cel l deposit. � Resuspend cells in 1 Oml hypoton ic - ( 0. 075M ) KCI.. I ncubate 20 minutes at 37°C. �. Centrifuge 5 mi nutes at 1 000 r.p.m.. •. Discard su pernatant. �· Cel l depos it. .. Stand 60 minutes at 4°C. �. ...... Suspend. �----.._. .....---. Centrifuge 5 minutes at 1 000 r.p.m. Discard 4-" supernatant Cel ls d ropped from p ipette onto ch i l led wet.,_ sl ides. Air-dried.. -. �. Cel l deposit. ________ .. t. Resuspend in few drops fresh fixative. cells in chil led, freshly prepared fixative.
(40) 39 II . 4. i). F IBROBLAST CULTURE. Int r o duct ion M any methods of c e l l culture h ave b e en used for th e. prepar at ion of metaphase chromo some s from mammal ian c e l l s . The c e l l culture t e chnique s commonly used invo lve ( 1 ) th e c ulture of t i s sue explant s with th e aid of adh erent s , and ( 2 ) m6nolayer c e l l cultur e s employing en zym e s for c e l l d i sp e r s i on . A lth ough enzymic disaggregat ion for small explant s of s o lid t is su e h as been des cribed ( Puck , Cie c iura and R ob inson , 1 95 8 ) , th i s t e chnique is often inade quate f o r f ibrous t is sue s and small b iop s ie s .. For th e se t is su e s th e explant. culture methods using adh erent s are effect ive . S inc e th e introduct ion of th e c e llu l o s e sponge t o h o ld e xp lant s against th e growing surface ( Leighton , 1 95 1 ) , a wide range of adh e rent s performing a sim ilar t ask h as been d es c r ib e d .. H su and Ke llogg ( 1 960 ) u s e d a c e l loph ane sh eet. to h o l d t i s sue exp lants against th e growing s�rf ace , wh ile B asrur , B asrur and G ilman ( 1 963 ) d e sc ribed an exp l ant cul t ure t e chnique allowing chromo s ome ch arac t e r i z ation of c e lls growing on covers lip s .. Prob ably th e mo st p opular meth o d. of f ix ing t is sue fragment s t o th e gr owing surfa c e has b e en th e u s e o f a p l asma clot ( Harnden , 1 960 ) .. C e l l s from th e. primary exp lant may be dispersed with tryp s in and subcultured , o r th ey may b e studied dire ct ly .. Th e pro c e dure used in th is. study was adap t e d from the p lasma c lot emb edding t e chnique de s c r ib e d by Hyman ( 1 968 ) . Th e mono layer t e ch nique of c e l l culture develope d by Evans and E arle ( 1 947) has proved very pop ul ar and it s use h as b e e n d e scrib e d by a large numb er of worker s ( R inaldini ,.
(41) 40 1 954 ;. H inz and Syverton , 1 95 9 ) .. The c e l l s are dispers ed. from living t is sue s by the action of enzyme s , mo st commonly tryp s in , o ft e n supplement ed by me chanical agit at ion .. A l. though mo st suit able for the c e l l culture o f embryonic t i s sue s ( Mo s c ona� 1 952) , the culture of tryp s in-dispersed c e l l s a s a monolayer has b e en suc c e s sfully app l ied to a wide r ange of t i s sue s and ext ende d e ven t o p l ac ent al t i s sue (Thiede , 1 960 ) and trophoblast ( Valent i , 1 965 ) .. A s imilar. pro c e dure was followed by the author for the mono layer cell culture of e quine embryonic t is sue s . A ll c e l l culture pro c e dure s r e quire c ompl e t e steri l ity and a h igh degre e of cleanline s s of apparatus and glassware . Det a i l s of the material s and so lut ions used are given in Appe nd ic e s V I I and VIII . ii). P r eparat ion of cell mono layer from t i s sue explant s M e t aphase spreads of c e lls from solid t is sues were. obt ained f ollowing c e ll culture by a p lasma c l ot embedding t e chnique ( Hyman , 1 968 ) .. This pro c e dure was employed for. t is su e s ample s from abort ed e quine f o etus e s and living adult hors e s . a ) Preparat ion of t is sue exp lant s P rior t o samp ling , the b iop sy s ite was g iven a rout ine surgical preparat i o n .. A s amp l e of sub cut ane ous. f a s c i a was exc ised from the neck of a standing horse with the aid of a l o c al anae sthe t i c (Xy l o caine 2% ) .. This was. inf i lt r at e d in the form of an L-b l o c k t o exc lude toxic eff e ct s of the local anae sthe t ic s o lut ion from the t is sue samp l e .. A s imilar procedure but without l o c al anae sthet ic. was f o l lowe d for the sampl ing of t i s su e s f rom non-living spe c imens .. T i s sues sele c t ed for culture included mus c l e ,.
(42) 41. t endon , c art ilage , l iver , lung , mucous memb r ane , amnionic membrane , tunic a albuginea and s kin . A t is sue sample one cub ic c ent imetre in s iz e w� s c o l le c t ed int o a 25 ml universal b ottle c o nt a ining ten ml of HBSS .. Under a laminar flow c ab inet the t i s sue was. divided int o fragment s about 1 mm s quare .. The s e were trans. f erred to a universal c ont ainer , t en ml o f growth medium were added , and the culture place d in an incub at or at 3 7° C . A f t er two days incub at ion the float ing explant s were rep l a c e d from the universal int o 50 ml culture f lasks . F ourt e e n exp l ant s , each t o be emb edded bene ath a p lasma clot , were e quidist ant ly p o s it ioned on the floor o f e ach f l a s k . At l e a s t 6 cultures were prepared for each spe c imen. b) Method for obt aining plasma F r e sh p lasma was obt ained from a normal donor or , when c o nvenient , from the p at ient under inve st igat ion . B lood was c o l l e cted asept ically into a st e r i l e vacut ainer c ont aining thr e e ml of A l s ever ' s s o lut ion , c entrifuged at 2000 r . p . m . for ten minut e s , and the calc ium-free plasma withdrawn int o a sterile b i j ou bott le .. P la sma was be st. used immediat e ly but in s ome case s was stored at 4vc for as long as t wo days . c ) Explant at t achment for primary c ulture The e qu ine plasma was re c onst itut ed by mixing 0 . 2 ml of s t e r i l e c a l c ium chloride s o lut ion (2% C aC l 2 ) with one ml of the prepared p lasma s o lut ion. approximat e ly f ive minute s .. C lott ing b egan aft e r. One drop of re c onst itut ed. plasma was superimposed over e ach explant and the exc e s s removed with a P asteur p ipett e , l e aving the t is sue fragment surrounde d by a thin f i lm of p lasma .. F irm c lot format ion.
(43) 42 was enhanced by incubat ing the culture flasks f or 1 5 minut e s at 37° C before growth medium was added . Explant s in primary culture s remain att ached t o the f lask while c e l lular growth radiat e s outwards (F igure I I . 3 ) . F ibrob last - l ike c e ll s mo st commonly pro�if erat e from explant s �. obt ained from almo st any o rgan ( Puck , Cie c iura and Rob inson , 1 95 8 ) . Before the out growth of l iving cells c o mmenc e s , pro t e o lyt ic dig e st ion of the c lot by enzymic action within the exp l ant may re sult in the exp lant bre aking loose from the c lot (Hyman , 1 968 ) .. The p eriod of incub at ion with. exp lant s float ing in growth medium appeared to reduce e nzyme act ivity and the prob lem was overc ome . d) Deve lopment of c e l l mono l ayer When ade quat e c e ll growth around the eA� lant s had - -. deve loped , the primary culture was rinsed witb two ml of pho sphat e buff ered saline ( PB S ) and incubat e d at 3 7° C f or f ive minut e s with one ml o f 0 . 05% trypsin s o l ut ion (ATV ) . ... Tryp s in act s on the int erf ibrillary substance ( Day , 1 949 ) and the plasma c lot , dis lodg ing b oth growing c e l l s and exp l ant s . After c e ll det achment was complete , the remaining tryp s in was inact ivated by the addit ion of one ml of f o etal bovine serum t o e ach culture f lask .. Growth medium was. rep lenished and the culture incubat ed for s ix hours b e f ore the media cont a ining c e llular debris and f lo at ing explant s was a sp irat ed and replaced with fresh media .. Living c e ll s. remain att ached t o the f l o o r of the flask and mult ip ly t o f orm a mono l�yer ..
(44) 43. F igure II . 3 .. Primary c e ll culture showing outgrowth of ce l l s from a t i s sue exp lant . (Magnif ication 5x) ..
(45) 4-4iii). Tryps in d i s aggregat ion te chnique £or c e l l cu lture a ) Treatment of embryo s E qu ine embry o s aged between 35 and 65 day s of ge st a. t ion were remove d asept ically from the ut eri of pregnant mares and transp ort ed separat e ly to the lab orat o ry in 500 ml screw-t op j ars c ont aining a sterile transport medium (Appendix V I I I a ) .. The int act embryo was pho t ographe d and. a measure o f crown-rump length as s e s s ed with a f l exib l e rule .. Due t o varying crit eria of measurement , it was. difficult t o obt ain a c ons ist ent st andard for aging horse embryo s .. The method employed was comparab le to the 11 cur'ved. crown-rump length " used by Joubert ( 1 956) , t aken as 11 the gre at e st distanc e from the t ip of the forehead along the dorsum to the t a i l head" .. The age of the embryo was then. c alculat ed from known standards (B ergin , G ier , Frey , and M arion , 1 967 ;. E vans and Sack , 1 973 ) .. Under s t e r i le condit ions the embryo was s eparat e d from i t s membrane s , transferred to a petri d i s h cont aining two ml of an ant ib iotic solut ion (A pp endix - V I I I c ) and choppe d as f in e ly as p o s s ib l e with cro ssed s c alpe l b l ade s . The re sult ing- mas s of cub e s of edge less than t wo mm float ing in ant ib iot ic solut ion was transferr e d t o a 1 25 ml f l ask and immer s e d in tryp s in . Problems o f c e llular c o ntaminat ion were c o nsidered to be minimal b e c ause e quine embryo s were obt ained within their int act p lac ental membranes b e f ore imp l ant at ion was advanc e d .. For c e ll culture only the embryo it s el f was u s e d. and t h e surrounding membrane s were discarde d ..
(46) 45 b ) Primary culture F ifty ml of 0 . 25% t ryp s in solut ion were introduced to the f lask and the mixture agit ated with a magnet ic st irrer . Aft e r 2 0 minut es agitation at 3 7° C the sup ernat ant was f ilt ere d through a st ainle s s s t e e l mesh int o s e ve ral univer sal cont ainers f o r centrifugat ion.. O c cas ionally c e ll y i e ld. was insuff ic ient at this st age and a further 50 ml of tryp s in s o lut ion were added and the flask content s agit ated again for 20 minut e s .. In some c a s e s this pro c e s s was repeated. several t ime s . F o llowing centrifugat ion the supernat ant was remov�d and the c e ll s re suspended in t en ml of foetal bovine s e rum , transfe rred to a graduat ed c entr ifuge tube and spun at 1 500 r . p . m . f o r f ive minut e s . The c e ll c oncentrat ion was e st imat ed from packed c e ll vo lume in the graduat ed c entr ifuge tube .. T o ino culate. primary culture s , c e l ls were re suspended in f o et al bovine serum to a c onc entrat ion of 2% packed cell vo lume , and 0 . 5 ml of the re sult ant suspen s ion introduced int o e ach 50 · m1 culture f lask.. Each culture was fed with f ive ml of growth. medium ( Appendix V I I b ) and incubat ed at 37° C .. A minimum. number of 7 primary culture f lasks were prep ared from e ach spe c 1me n . The l ag p eriod o f growth f ollowing the exp l ant at ion of a suspens ion of c e ll s aft e r treatment with tryp s in ( Levan and B ie s e le , 1 95 8 ) was not evident in this study .. Normal. and abnormal c e l l populat ions apparently have a s imilar init ial growth rat e ( Hyman , 1 968 ) but the p o s s ib ility that chromo s ome anomal i e s may be c aused or conc e aled by eff e c t s of c e l l culture c annot b e ignored ..
(47) 46 iv). R eplenishment of growth medium R e qu irement s of growth media and growth limit ing. f actors have been we ll documented ( Harris , 1 955 ; 1 968 ) .. Hyman ,. Details o f the growth medium employed in this study. are l i s t e d in App endix V I I . H arris ( 1 955 ) report ed that growth rat e is indepe ndent o f serum c oncentrat ion above a minimal conc ent rat ion ( 1 0 -1 5%) . Hsu and Ke llogg ( 1 960 ) recommended 20% serum in growth media for primary culture s and an atm6 sphere of 5% c arb on dioxide t o prevent a rap id rise in pH .. Although high levels of serum. have b e e n f ound t o lead t o incre ased lipid c ont ent of c e lls and ac idif icat ion of media ( Hanks , 1 948 ) ,. a. c oncentrat ion. o f 40% s e rum was found to be suit able f or this study . F ive ml of growth medium was used t o sust ain c e l l growth i n 5 0 m l culture f lasks .. T o remove unatt ached c e lls. and debr i s , the growth medium was renewed s ix hours aft er the ino culat ion of primary and s e condary c e ll culture s . Otherw i s e the growth medium r emained unchanged for as long as one we ek unl e s s there was a marked change in pH as indic at e d by a c o l our change due to the pheno l red c ont ained in the growth medium . A c t ively growing cultures o f embryonic origin provide c arb on d ioxide from c e l l met ab o l i sm and ad justment of cul ture atmo sphere with 5%. co 2. ·. is unn e c e s sary for such t is sue s .. Howeve r , tric ine buffer ( Appendix VI I . c ) was inc luded in the growth medium t o st ab il iz e pH . Disturb anc e s of pH c an c au s e aneup l o idy , as not e d -. by F ord ( 1 97 3 ) , who als o ob s e rved chromo s ome abnormalit i e s i n act ively dividing human f o etal f ibroblast s cultured in an alkal ine medium .. F luctuat ions in pH have b een sugge sted.
(48) 47. ( Ford , 1 973 ;. Ingall s and Shimada , 1 974) as a mechani sm for. induc ing chromo s omal errors in the zygot e: v). A s s e s sment o f c e l l growth An invert ed micro scope was used to as s e s s c e l l growth. in culture f lask i .. A culture was subdivide d when c e l l. mult ip l i c at ion had formed a c omp lete mono layer o n the f loor of the f lask . A l l c e l l s adopt an e longated conf igurat ion when grown for suf f ic ient ly long periods in culture ( Puck , C ie c iura and R ob inson , 1 95 8 ) .. F ibrob lasts in the proc e s s of mit ot ic. divi s io n are charact eriz ed by a change in morpho logy from spindle - shaped to spherical .. Thus the proport ion of divid. ing c e l l s c an b e est imat ed by micro scopic examinat ion t o give an indicat ion o f the t iming and effect o f c o lchic ine treatment ( F igure s I I . 4 and I I . 5 ) .. A culture was c on s id. ered suit ab le f o r c o l chic ine t reatment to arre st mit o t i c division i n met aphase when the c e l l mono layer c overed approxim at e ly 50% of the available surface ..
(49) 48. F igure I I . 4 .. F ibrob last c e l l culture which has formed a comp le t e mono layer . ( M agnif icat ion 5x) .. F igure I I . 5 .. Mono layer c e ll culture aft er 1 6 hours expo sure to c o lchic ine . Not e the large number o f spheric al c e l ls which have been arre sted in metaphase. •.. ( Magnif icat ion 5x) ..
(50) 49 vi ). Preparat ion of culture s for sub culture or harve st M o no layer c e l l culture s growing in 50 ml flasks were. e ither d iv ided into s e condary culture s , froz en for st orage , or treat e d with c o lchic ine and harve sted .. A st andard t e chni. que was u s e d to provide a c e l l su spension . Gro wth medium was asp irated from the culture flask and the mono layer rins ed with two ml of PBS .. Under the. act ion of o ne ml of 0 . 05% t ryp s in s o lut ion ( ATV ) , living f ibrob l as t s be c ame spherical and lo st the ir att achment t o the f l as k .. Depe nding on c e l l type and the age o f the culture ,. det achment was c o mplete aft er f ive t o 15 minut e s at 37° C . · R ap idly growing c e l l s and tho s e arre sted in met aphase by c o l chic ine were dis lodged more e a s ily than o lder c e ll s in s l o w growing culture s .. An inverted microscope wa s used to. che ck that all c e l l s were det ached . Sus c ep t ib i l ity to tryp s in varie s acc ording to c e l l typ e ( Harr i s , 1 95 5 ) but tryp s in is very slow t o damage l ive c e l l s ( M edawar , 1 941 ;. Mo sc ona , 1 95 2 ) .. Tryp s in s e lectiv e ly. dige st s d e ad c e l l s due to change s in p e rmeab i l ity ( R inaldini , 1 954) .. Whether or not the t ryp s in tre atment o f c e ll culture s. result s in an increased frequency of chromo some breaks i s a matter o f disput e ( Levan and B ie s e le , 1 958 ;. H su and K e llogg ,. 1 960 ) . a ) Sub cultur e s When c e ll det achment was c omp let e , surp lus tryp s in was inact ivat e d by the addit i o n o f one ml of f o et al bovine s e rum ( F . B . S . ) to e ach culture f l ask . Sub culture s were prepared by ino culating half of the c e l l suspens ion in e ach culture ( one ml ) into a fresh f lask . T he original f lask and new sub culture were both f ed with f ive ml of growth medium and incub at e d at 3 7° C .. Thi s proc e s s.
(51) 50 c ould b e repeated many t ime s and a maximum of e ight p a s s age s was used for one case ( 1 9B ) in this study . b ) H arve st o f f ibrob last cultur e s C o l chic ine was inoculat ed int o each culture to a f inal c onc ent rat ion of 0 . 02 �g/ml and incubat ion c o nt inued for 1 6 hours b efore harve st .. Each culture was tre at e d with tryp s in. as d e s crib e d previou s ly and transferred to a graduated c entrifuge tube t ogether with the growth medium and PBS washings from the same flas k . Sub s e quent harve st ing procedure s were ident ical t o tho s e u s e d for l eucocyt e culture s . vii). Storage of c e l l s Suspens ions o f c e l l s were st ored at -70° C .. Embryonic. c e l l s not requir e d for the ino culat ion of primary c e l l cul ture s were fro z e n after disaggregat ion and c o nc entrat ion . Primary and s e condary c e l l culture s surp lus t o re qui�ement s were a l s o c onc ent rated and fro z en at -70° C . C e il s de st ined for fro z en storage were suspended in FBS to a packed c e l l concentrat ion of 20% , to which was added 10% by vo lume of dimethyl sulphoxide .. The suspens ion was. dispensed in one ml aliquot s int o glass ampoule s , wrapped in several layers of c otton wo o l , p acked in a plast ic b ag and p lac ed at -70° C for at least 1 2 hours .. The g l a s s amp ou l e s. were then unwrapp ed and stored in a plastic c ont ainer . When re quired to ino culate fresh culture s , an amp oule was thawed rap idly at 37° C , d i lut ed with FBS and inoculat e d int o culture f lasks t o a f inal c onc entrat ion o f 2% p acked c e l l volume ..
(52) 5 '1 II . 5 . i). SEX CHROMATIN STUDIES. S e x chromat in o f nerve t i s sue During the autop s i e s o f thr e e horse s , s e c t ions o f. nerve t is sue were obtained w i thin one hour of de ath from the f o l l owing regions : � ( '1 ). C e rebe l lum .. ( 2). C e r�ical reg i o n of sp inal cord .. (3). T horac ic region of sp inal cord .. ( 4). Lumbar re gion o f sp inal c ord .. (5). Sp inal cord f r o m the region o f the sacral ve rtebrae .. The s e h i s t ological s e c t ions were f ixed in 4% f ormalde hyde for 24 hour s before undergo ing a rout ine emb edding pro c e s s prior t o sect ioning .. Sect ions of thickne s s 7 � were. transferred to s lide s and st a ined with cre syl e cht vio l et ( Co leman and B e l l , 2% ) for f ive s e c o nds .. The gray mat t e r. was examined micros cop ically f o r t h e pre sence of s ex chromat in . Approximat ely '1 00 c e l l s from e ach region of the sp inal c ord were s c ored for the pre senc e or ab s ence of a s ex chromat in body .. F or e ach c e l l examined a sket ch was drawn , showing. the shape of the cell and t he r e lat ive po sitions of the nuc l eus , nuc l e o lu s and s e x chromat in . ii). S e x chromat in o f polymo rphonu c l e ar neutrophils Peripheral blood sme ars from three hor s e s were rout ine ly. prepared , air dried , and st ained with M acNeal ' s st ain . S lide s -. were s c anned under o il immers ion ( 500x magnif i c at ion) and neutrophils s c ored for the p r e sence of nuc lear appendag e s ( " drumst icks " ) ..
(53) 52 II . 6 . i). ANALYSIS OF RESULTS. Micro s c opy A l l s l ide s were s c anned with the low p ower ( 1 0x). ob j e ct iv e .. Suit able metaphase spre ads were s e l e c t e d f o r. examinat ion under ··the 20x ob j e ct ive .. Vernier reading s o f. the r e l at ive p o s it ion of the micr o s c ope st age were re c orded f o r e ach s e l e cted c e l l .. The se lect ion and numb er of meta. phase s analyz ed are import ant factors in a s s e s s ing the b ias against the detect ion of po s s ible mo saic s .. T o avo id s e l e ct ion. of nuc l e i , every met aphase s e lected under low p ower was c ount e d under high p ower . Chromo s ome spre ads of sat isf act ory qua l ity were studied under oil immers ion at 500x or 1 000x magnif ic at ion. The chromo some s in e ach cell were count ed and r e corded , and any abnormalit ies noted .. From e ach case a numb er of c e l l s. were cho s e n t o b e photographed and ka+yotyp e d .. A s im i l ar. pro c e dure was adopt ed with the sex chromat in studie s . The numb er of c e l l s c ounted and analy z e d varied b etwe e n c as e s .. Hyman ( 1 968 ) c ons idered the analysis o f 20. c e l l s s e le cted at random from at least two culture s t o b e suf f ic ient exc ept in the case of mo saicism.. M cCreanor ( 1 974). judged the count ing of 30 - s e l e cted c e l l s suff ic ient t o exc lude mo s aic i sm .. M o s aicism is suspe cted if a cons is t ent abnormality. is f ound in some but not all c e lls . ii). Pho tographic t e chnique s Phot ographs were t aken at 500x magnif i c at ion with a. gree n B al z ers K-4 f ilter placed over the light s ourc e t o redu c e g l are .. The wide f ie ld ( 1 0x) eyep ie c e s were exchanged. for one o rdinary eyepiece ( 1 0x ) and one photographic eyepie ce with a f o cu s s ing frame ..
(54) 53 A c onst ant exp o sure t ime o f one sec o nd was adhered t o · with C op ex-pan rap id f ilm 3 7 A . S . A . (Agfa-G evaert ) .. E ach. f ilm was deve l oped , and enlarged and print e d on I lfobrom ( I l ford , Austral ia Propriet ary Ltd . ) Numb er 2 or 3 p ap e r , depending on wheth� r the negat ive was dark or l ight . further detai l s refer t o App endix IX ) .. ( F or. For the print ing of. ne gat iv e s two s c ale s o f enlargement were u s e d :. 6x and 9x. magnif icat ion . iii ). C onstruct ion of karyotype s . Ademco dry mount ing t i s sue was fixed t o the b ack of. a print with a heated iron .. Individual chrom o s ome s we re. c arefu l ly cut out , p aired , and arranged in group s on a rul e d c ardboard she et .. The l o o s e chromo some s , t ogether with. a photograph of the ent ire c e ll , were pre s sed ont o the c ard board she e t ( 70 -80° C for 1 5 s e cond s ) to comp l e t e the karyo typ e ..
(55) 54. PART I I I. GESTAT ION AND ABORTION I N THE MARE.
(56) 55 PART I I I. GESTAT ION AND ABORT ION IN THE MARE III . 1 . i). LITERATURE REVIEW. Normal Gestat ion The durat ion of pregnancy in the mare i s influenced. by m any f act ors (Trum , 1 95 0 ) and var i e s wide ly around a me an of 3 3 6 . days ( B e rgin , 1 970 ) .. Diffe rent iat ion of the embryo. and organogene s i s are c omp l e t ed during the f irst 30 days of g e stat ion . Individual mare s are mo st irregular with respect t o o e strous behaviour and the t iming o f ovulat ion , but mo st mare s ovulate about. 2�. hours b efore the end of behavioural. o e s t rus ( Hughe s , St abenfe ldt and Evans , 1 975 ) .. During e ach. o e s trous cyc l e many follicl e s re c ommence deve lopment but only a f e w ovulate while the remainder undergo atre sia . Aft e r rupture of the Graaf ian f o l l i c l e int o the ovulat ion f o s s a (W ithersp oon , 1 975 ) , the ovum is transferred to the f a l l op ian tube by the fimbr iated infundibulum .. Transport. o f the ovum to the site of f e rt i l i z at ion , the ampulla , is a c c o mp l i shed by c ilial current s and muscular contract ions unde r the cont r o l of sex hormone s . In mo st mammal s the fe rt i l e life of the e gg is probab ly about. 24. hours , but det eriorat ion commence s aft er a few hours. and m ay have imp ortant eff ect s on sub s e quent deve lopment . Sperm remain viab le in the f emale r eproduct ive t ract for c ons iderab ly longer than the o vum and in mo st specie s sperm must undergo a period of c ap a c it at ion in the f emale genital tract b e fore they are c ap able of f e rt il iz at ion .. Mat ing should. therefore o c cur s light ly before ovu lat ion t o ensure that.
(57) 56 c apacitated sp erm are pre s ent in the fallopian tub e when the ovum is lib erat e d .. However , the ne c e s s ity f o r a p eriod of. sperm c apac it at ion has not been establ ished in the hors e . The entry of a sperm int o the egg s t imulat e s b oth structural and funct ional changes :. mo st import ant are the. init iat ion o f deve lopment of the fert il i z e d o vum and the Suc c e s s ive c l e avage. induct ion o f the po lyspermy block .. divis ions c o nvert the f e rt ilized ovum into a ball of c e l ls , the morula , and ac cumu l at ion of f luid within the moru l a leads t o the format ion of the b last o cyst .. The b last o cyst c o n s i s t s. of a ho l low sphere c o nt aining an inner c e l l m a s s which g iv.e s rise t o the embryo , and the peripheral trophob last which f orms the p lacent a and extra-embryonic membrane s . Tubal t ransport. lS. again achieved by c i l iary act ion. and mus cular p eristal s i s , and the early blasto cyst re ache s the ut erine lumen usua l ly b etween four and s ix days aft er ovulat ion ( O guri and T sut sumi , 1 972) .. In the m B.re , only. fert iliz ed eggs ever re ach the uterus ;. unf ert i l l z e d ova. remain trapp ed at the isthmus of the fallop ian tub e for as long as s even months and may undergo some de gree of p artheno genetic c l e avage b efore degenerat ing ( van N iekerk and Gerneke , 1 966) . Like other mammalian embryo s , the e qu ine embryo has limit ed nutrient re s e rv e s and must rely on ut erine milk for nourishment b e f ore impl ant at ion.. The upt ake of ut erine milk. is enhanced by spe c ialized group s of c e l l s on the trophob last which f o rm a trans ient att achment to the ut erine epithe l ium after about 20 days of g e s t ation ( McLaren , 1 97 2 ) . The e qu ine p lacent a has b e en clas s i f i e d as diffu s e , microc otyledo nary and ep ithe l io chorial ( St eve n and Samu e l , 1 97 5 ) .. S ince there is no ero sion of mat ernal o r f o et al layer s.
(58) 57 it i s diff icult to de c ide exac tly when att achment of the embryo o ccurs .. Z ie t chmann and Kro ll ing ( 1 95 5 ) c onsidered. that the embryo remained unat t ached in the ut erine hor n for up t o five weeks , but van Niekerk ( 1 965 c ) not iced the f irst s igns of allant o iq deve lopment 21 days aft er ovulation . Not unt i l the e ighth we e k do chorionic villi grow int o the fo lds of ut erine wall (Aust in , 1 969) and by the f ourt eenth we ek imp l ant at ion is complet e (McLaren , 1 972) . As the primit ive yo lk- sac plac ent a i s c onvert ed int o a t rue allant o chorionic p lacent a , growth of the allant o is i s acc ompanied by the format ion of endometrial cup s , f irst vis ib l e as a band around the c ircumf erence of the pregnant ut e r ine horn aft er about 40 days of ge station ( C legg , B o da and C o l e , 1 954) .. Sp e c ialised chorionic girdle c e ll s invade. the endometrium on the 3 6th day (Moor , A llen and Hamilt on , 1 975 ) t o produce the endometrial cup s re spons ib le for the s e cret i on o f pregnant mare ' s serum gonadotrophin ( PMSG) . The pre c is e func t ion o f the endometrial cup s has remained ob s cure , but they are the sour c e of high leve ls of gdnado trophin in the mare ' s b l o od between 40 and 1 30 day s of pregnancy ( Allen , 1 969) .. Use i s made of this in the haemag-. glut inat ion inhib it ion a s s ay for pregnancy diagno s i s . R e c e nt stud i e s have indicat ed that the primary corpus lut eum doe s not. regre s s comp let e ly unt il mid-ge stat ion ( Squire s , Garcia. and G inther , 1 974) and ovar ian act ivity during pregnancy i s . .. mo s t marked before day 40 ( Bain , 1 967b ) , thereby pre c eding the product ion of PMSG by the endometrial cup s .. Se cret ion. of PMSG is halt ed as the mare ' s defenc e me chanisms rec ogniz e the cup s as f ore ign t i s sue and mount an immuno logical re j e c t ion .. The fo etal genotyp e has a profound inf luenc e on PMSG. product ion and the lif e span of the endometrial cup s ( A l len ,.
Figure




+7




Related documents
The use of sodium polyacrylate in concrete as a super absorbent polymer has promising potential to increase numerous concrete properties, including concrete
Junior Positive mental health inventory prepared by Agashe and Helode (2009) was used as psychological instrument in the present study.Results reveal significant impact of yoga
Radio and TV broadcast music related programmes which provide opportunities not just. to listeners but also to the
21 Department of Neurosurgery, Tangdu Hospital, The Second Affiliated hospital of the Fourth Military Medical University, 1 Xinsi Road, Xian, Shanxi Province 710038, People ’ s
In this paper we trace the early history of relativistic su- pertasks, as well as the subsequent discussions of Malament-Hogarth spacetimes as physically-reasonable models
Nitri fi cation and sedimentary denitri fi cation occurred near the river mouth, nitri fi cation prevailed further offshore under the plume, and fi nally, phytoplankton
2 Equity Market Contagion during the Global Financial Crisis: Evidence from the World’s Eight Largest Economies 10..
In 2008, the first pilot experiments were conducted in seven organic winter oilseed rape fields (total area: 4.2 ha) in North-western Switzerland.. One half of each field was
