1982, p. 636-648 Vol.
0022-538X/82/050636-13$02.000/
DNA
Synthesis
and Gene
Expression
in Bacillus
subtilis
Infected with
Wild-Type and Hypermodification-Defective
Bacteriophage
SP10
HEMANWITMER*ANDMICHAELFRANKS
Departmentof Biological Sciences, University ofIllinois atChicago Circle, Chicago,Illinois60680 Received25September 1981/Accepted24 December1981
A hypermodified base (Y-Thy) replaces 20%o of the thymine (Thy) in mature
DNAof Bacillus subtilis phage SP10. Two noncomplementing hypermodification-defective(hmd)mutants aredescribed. At 30°C, hmd phage carriedout a normal program,butattemperaturesof.37TC,the infectionprocess wasnonproductive. When cellswere infectedat37°C with hmd phage, DNA synthesis started atits usual time(12 mmin), proceededat about half the normalratefor 6to8min, and
then stopped ordeclined manyfold. All, ornearly all, of the DNA made under
hmd conditions consisted offully hypermodifiedparental DNA strands H-bonded
to unhypermodifled nascent strands. The reduced levels of DNA synthesis
observed under hmd conditions were accompanied by weak expression oflate genes. A sucrosegradientanalysis ofSP10hmd+ replicatingDNA intermediates wasmade. Twointermediates, calledVFandF,wereidentified. VF consisted of condensed DNAcomplexedtoprotein;VF also containednegatively supercoiled
domains covalentlyjoinedtorelaxedregions.Fwascomposedof linear
concaten-ates from which mature DNA was cleaved. None of those intermediates was evident in cells infected at 37°C with hmd phage. Shiftup experiments were performed wherein cells infected with hmdphageat30°Cwereshiftedto37°Cata time when replication was well underway. DNA synthesis stoppedor declined manyfold10 minaftershiftup.ThehmdDNAmade aftershiftupwasconservedas
aformsedimentationally equivalenttotheFintermediate,but littlematureDNA
was evident. It is proposed that Y-Thy is required for replication and DNA maturation because certain key proteins involved with theseprocesses interact preferentially withhypermodifiedDNA.
Bacillus subtilis phage SP10 belongs to a
group of viruses in which thymine (Thy) in mature DNA is partially replaced by a hypermodified base (10,37,39). The hypermodi-fied base in SP10DNA(designated Y-Thy) has not beencompletely characterized with respect to chemical structure (37). Other members of this group include B. subtilis phage SP15 and Pseudomonas acidovorans phage 0W-14, in which Thy is replaced by 5-(4',5'-dihydroxypen-tyl)uracil (DHP-uracil) and a-putrescinylthy-mine, respectively (4, 21). In the case of SP15,
DHP-uracil and Thy are assembled as free
de-oxynucleoside triphosphates that are subse-quently inserted intothe DNA polymer (36). In thecaseof SP10and 0W-14, however, neither Thy nor the respective hypermodified base is assembledas afreemononucleotide but,rather, each originates by post-replicational modifica-tion ofacommonprecursor base that is
proba-bly 5-hydroxymethyl(HM)uracil(10, 27, 31,37, 39). SP10 and0W-14aretheonly known
biolog-icalsystemsin whichThyin DNA does not arise via the deoxythymidylate synthetase reaction
(37).
Little is knownabout thebiologicalrelevance ofhypermodified basesin DNA. Recent studies suggestedthatDHP-uracilplaysnomajor role in
differentialtranscription of SP15 DNA (16) but
thatnormallevelsofthehypermodifiedbase are required for proper DNA maturation, phage assembly, orboth (6, 36). These earlier studies were confounded by the fact that no situation couldbe definedinwhichSP15DNAdevoid of
DHP-uracilwas made (6, 36).
This paperconcernstheisolation and charac-terization of hypermodification-defective (hmd)
mutants of phage SP10. The results of these experimentssuggest that Y-Thyis indispensable to SP10 development. Specifically, Y-Thy
636
on November 10, 2019 by guest
http://jvi.asm.org/
ROLE OF Y-Thy IN SP10 DEVELOPMENT 637 seems to be required for proper replication of
thephagegenome aswellascleavageof concat-enatestounit-size DNA.
MATERIALSANDMETHODS
Phageand bacteria.SP10hmdl andSP10hmd2are
heat-sensitivemutantsdefective in hypermodification (see below and reference39). SP10 hmd+is the same
clear-plaque variant used in allprevious studies (29, 30).B.subtilis W23wasthe hostin all experiments.
Mutagenesis.Cells infectedwithSP10 hmd+at30°C
weremutagenizedwith
N-methyl-N'-nitro-N-nitroso-guanidineasdescribedby Kahan (20).
Screening forhmdmutants.Survivorsof
mutagene-sis were plated at 30°C. Plaques were picked with
steriletoothpicks and scoredfortheirabilitytoplateat
42°C.Lysatesofheat-sensitivemutantswereprepared at30°C and usedtoinfect cellsat42°Cin thepresence
of6-(p-hydroxyphenylazo)uracil,aninhibitor of
bacte-rialDNA polymeraseIII (5, 32) that hasnoremarkable
effect on SP10development(29, 30). Thecells were
labeledwith[6-3H]uracil(10 uCi/ml)for 15min. Cellu-lar DNA was isolated(6) and acid hydrolyzed to a
mixtureofpurine basesandpyrimidine deoxynucleo-sides(36). The digestswerechromatographedinone
direction on unmodified cellulose thin layers (10),
usingdeoxythymidine (dThd) andY-dThdas optical
markers. Spotscorrespondingto the optical markers
wereeluted (10, 29),and theamountof label present
was determined. In those caseswhere no labelwas
recoveredin theY-dThdspot,asampleof [3H]DNA
wasenzymatically degradedtomononucleotides (36, 39) thatwerefractionated by two-dimensional chroma-tography onunmodified cellulose thin layers (7). Of
the 184 single-plaque isolates tested to date, only 2 generatedDNAat42°C whichwastotally devoid of
Y-dThd. The basesreplacingY-Thy inSP10 hmdl and SPO hmnd2 DNAs, undernonpermissive conditions,
are 5-(hydroxymethyl-O-pyrophosphoryl)uracil and
HM-uracil,respectively (39).
Sucrosegradient centrifugational analysisof
replicat-ingDNA.Cultures were infectedasdescribed previ-ously (29, 30).Thelabeledprecursor,typically32p, (10 ,Ci/ml),wasintroduced12minafterinfection. Atthe desired times,5-ml sampleswereremovedand mixed with 20 ml of ice-cold medium that contained 10mM KCN. Thecellswereharvestedbycentrifugationand suspendedinto 2 mlof 10mMTris-hydrochloride (pH 7.9),10mM disodium EDTA(pH 7.9),1 mMKCN, and 1mgofeggwhitelysozymeperml. Thecellswere
incubatedat37°Cfor 20min,and 0.1 ml of11%(wt/
vol)sodiumlauroylsarcosinewasadded tocomplete lysis.
A100-1l portionofthe extractwasmixed with100 p.1 ofasolution containing marker [14C]DNA. Each
markerwaspresentat0.5 pLg.The entire mixturewas
layeredontoa5.8-mllinearsucrosegradient (5to25% sucrose in10 mMTris-hydrochloride, pH7.9-1 mM
disodiumEDTA,pH7.9-1 MNaCI-0.01% sarcosine). Allgradientswere spuninan SW50.1 rotor at150C.
Theexactconditionsofcentrifugationaregivenin the
appropriate figurecaptions. Fractions (100 pLI) were
collectedfrom thetopwithanISCOdensitygradient fractionator,usingFluorinert FC-48asthechase
solu-tion.The fractionswereindividuallyprecipitated (6), andthe amount of labelinDNAwasdetermined.
Isolation of RNA.Unlabeled RNA was isolated from
2-litercultures by the method of Bolle et al. (2) as modified by Legault-Demare and Chambliss (22).
[3H]RNA was isolated from 50-ml cultures labeled with[5-3H]uracil(5 uCi/ml).
Isolation of SP10 DNA strands. Strands of SP10 DNA were separated in CsCl on the basis of their differential binding of polyuridylate-polyguanidylate
(16). TheCsCl-purified strands were self-annealed and passed through hydroxyapatite columns to separate single strands from residual duplexes (19).
Other methods. DNA-RNA hybridization and hy-bridization-competition were carried out as described byGillespieandSpiegelman (14). Enzyme assays and definition of activity units are given elsewhere (29, 39). Serum blocking power was determined by the method of DeMars (8). Protein concentration was measured by the method of Lowry et al. (26). The incorporation of radioactive precursors into DNA was measured as described by Lembach and Buchanan (23).
Materials. Radioactive materials were purchased from New England Nuclear Corp. (Boston, Mass.), except [6-3H]HM-deoxyuridine, which was synthe-sized by the method of Alegria (1).Nitrosoguanidine was purchased from Aldrich Chemical Co. (Milwau-kee,Wis.). Enzymes were obtained from Boehringer-Mannheim (Indianapolis, Ind.). Sucrose and Fluorin-ert FC-48 were purchased from Schwarz/Mann (Orangeburg, N.Y.) and Instrumentation Specialties Co. (Lincoln, Neb.), respectively. All nonradioactive materials werepurchased from Sigma Chemical Co. (St. Louis, Mo.).
RESULTS
Thermolability of hmd phage. Wild-type (hmd+) phage produced high bursts at all tem-peratures but, by this criterion, the infection processwasmostefficient between 30 and37°C
(Table 1). At temperatures of < 30°C, mutant (hmd) phagegave bursts that werewithin60% of normal, butathigher temperaturesthe burstsize declinedsharply(Table1).Forthispaper,hmd+
and hmd phagewerecomparedat37°Csincethis is the temperature at which most biochemical studies with SP10 were performed (10, 29, 30,
39,40).
TABLE 1. Burstsize ofSPIOhmd+andSP10hmd phagesatvarioustemperaturesa
Burstsize
Phage
200C 300C
330C
37°C420C
SPIOhmd+ 31 87 89 83 63
SPno
hmdl 20 54 12sl
x 10-4sl
x 10-4SP10hmd2 21 58 12 sl x i-0sl x
i0-4
a
Cells
weregrown at37°Cto adensityofca. 2 x108/ml.Thecellswereharvestedbycentrifugationand
resuspendedinto0.95 volume of fresh medium
equili-bratedtotheindicated temperature. Ten minuteslater, the cells were infected by adding 0.05 volume of medium thatcontainedthephage(inputmultiplicityof infection=0.5PFU/cell).Theremainder of the
proce-durewasidenticaltotheoneusedbyBottand Strauss (3). Allplatingsweredoneat300C.
VOL.42, 1982
on November 10, 2019 by guest
http://jvi.asm.org/
638 WITMER AND FRANKS
E L 0 _ 2 * 04zz 0 ; X X M p
w 200 hmd50o 20
Iytice ( S ( 0 (S0m 0
0 3 6 912 15 1821 012 1824 3036 42 4854 012182430 36 4248 54 Minutespost infection
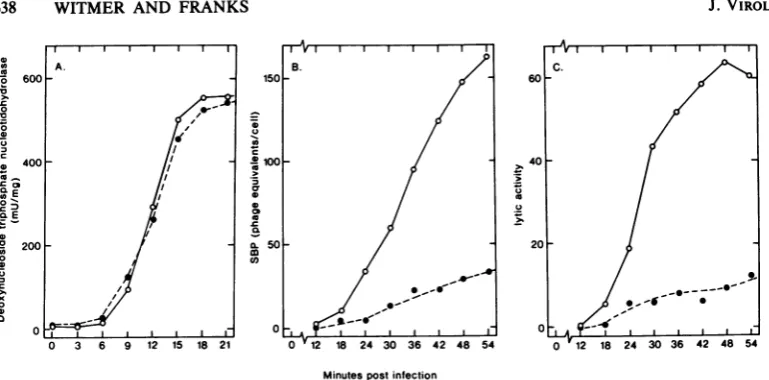
FIG. 1. SynthesisofSP10-codedproteinsunderhmd' andhind conditions. Cultures(300 ml)wereinfected
with SP10 hmd' or SP10hmdl at 370C. Atthe times specified inthe graph, 25-mi samples were removed.
Extractswere preparedand assayedfordeoxynucleoside triphosphatase (A), serumblocking power(B), and
lyticenzymes(C). Symbols: (0)SPIOhmd';(0)SP10hmdl.
Complementation of hmd mutations. When
cells weredoublyinfected withhmdlandhmd2, the burst sizewas nogreater than that obtained
when cells were infected with only one of the
mutants (data not shown). These data suggest thathmdl and hmd2do notcomplement, which, inturn, suggests that both mutations arein the
samegene.
Synthesis of phage-coded proteins. The
condi-tionally lethalnature of hmd mutations
suggest-ed thatsomeimportantaspectsofSP10
develop-ment were aberrant in the absence of
hypermodification. Toseeif thedefect involved early or late functions, synthesis of SP10
pro-teinswascompared under hmd+ and hmd
condi-tions. In this and all other experiments, only dataobtainedwithhmdl phage will be presented unless statedotherwise.
A typical early enzyme, deoxythymidine
tri-phosphate nucleotidohydrolase (29), was
syn-thesized normally under hmd conditions (Fig. 1A), whereastwolatefunctions, structural
pro-teinsresponsible forserumblockingpower(Fig.
1B) and lytic enzymes (Fig. 1C), were
synthe-sized poorly. Therefore, under the conditions
defined, theabsence of hypermodification limit-edlate geneexpression.
Synthesis of phage-coded RNA. Since
hyper-modification is an event at the DNA polymer
level (39), it seemed reasonable to assumethat
thelow levels of lateprotein synthesis observed
under hmd conditions were due to poor
tran-scription oflate genes. To confirm this,
DNA-RNA hybridization competition experiments
weredone (29). For these experiments, parallel
cultures of B. subtilis W23were infected under
hmd+ conditions. One culture waslabeled with
[5-3H]uracil from 4to12min postinfection(early
[3H]RNA), and the other was labeled from 12 to 20min postinfection (late [3H]RNA) (29). Unla-beledcompetitor RNAs were isolated 12 and 20 min postinfection by hmd+ and hmdphage.
Unlabeled 12-min RNAs isolated from cells infected underhmd+ and hmd conditions were equivalent with respect to theirability to com-pete against early [3H]RNA (Fig. 2A). By the same token, hmd+ and hmd 12-min [3H]RNAs each competed against sequences that account-ed for about 60% of the label in late [3H]RNA (Fig. 2A). Thesedata indicate that 12-min RNAs present in hmd+- and hmd-infected cells were equivalent. This conclusion is supported bythe observation that unlabeled hmd+ and hmd 12-minRNAscompeted equallywellagainsthmd 4-to12-min [3H]RNAs (datanotshown).
On the other hand, hmd+ and hmd 20-min RNAs displayed marked differences (Fig. 2B). Specifically,sequencesamountingto30% ofthe label in late [3H]RNA were either absent from 20-min hmd RNA or presentin low concentra-tions. This difference is better illustrated by a "mixed competitor" experiment (2) in which late [3H]RNA was incubated simultaneously with asaturating concentration of hmd+ 12-min RNA and various concentrations of hmd+ or hmd 20-min RNA(Fig.2C). Overall,these data areconsistentwiththeinterpretationthat genes transcribeduniquelyattimeslaterthan 12minof anormal SP10program were weakly expressed underhmd conditions.
Kinetics of DNA synthesis. In many phage systems, expression of late genes is, in some measure,dependenton DNAsynthesis (15, 18). Although the screening procedure showed that some DNA synthesis took place under hmd conditions, the possibility still existed that the
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.505.57.442.56.246.2]ROLE OF Y-Thy IN SP10 DEVELOPMENT 639
100
z
cc
I
CcO
E D
D >E
E
--E
0
50I
0
.I
B._-A.
-l
I
-
I-I
a
0\
_ 0
0 0.5 1.0 0 0.5 1.0 0 0.10.20.3 0.
Competitor RNA (mg/ml)
FIG. 2. DNA-RNAhybridizationcompetition between early and lateSP10RNAs made under hmd' andhmd conditions. Early[3H]RNAwaslabeled from 4 until 12min postinfectionbyhmd+ phage; late [3H]RNAwas
labeled12 to 20minpostinfection by hmd+ phage (29). Unlabeled RNAs were isolated 12 or 20minpostinfection
byhmd+ or hmdl phage.Hybridizationmixtures contained 1 ,ug of[3H]RNAandfilters that contained 20p.gof
heat-denaturedphage DNA. (A)Hybridization competition of early[3H]RNA.Thespecific activity of[3H]RNA
wasca. 32,000cpm/4gand, intheabsence of unlabeledcompetitionRNA,ca.11% ofthe input was retained. Unlabeled RNA was extracted 12 min postinfection by hmd+ (0) or hmdl (0) phage. (B) Hybridization
competitionof late [3H]RNA. The specific activity of
[3H]RNA
was 23,500 cpm/p.gand, in the absence ofcompetitor, 33%of the input was retained by theifiter.Theunlabeledcompetitorswere(0) hmd+12-minRNA, (0)hmd+20-min RNA, and (0) hmdl 20-min RNA. (C) Mixed competition of late [3H]RNA. Late [3H]RNA was simultaneouslycompeted by 1 mg of hmd+12-minRNA and theindicatedamountsofeither(0) unlabeledhmd+
20-minRNA or(0)unlabeled hmdl 20-minRNA.
I D
apparently weak expression ofSP10late genes observed under hmd conditions could reflect someupsetin themechanism ofDNAsynthesis. Kineticsof DNA synthesiswasmonitored by the incorporation of [6-3H]uracil into
acid-pre-cipitable, alkali-labile material (23). By this
cri-terion, replication of the viral genome, under
hmd+ conditions, was firstevident 9 to12 min
postinfection and continued for the duration of
the latent period (Fig. 3). During an hmd pro-gram,DNAsynthesisstartedatthe normaltime,
proceededatabout half the normalrateuntil18
to21 min, and thenstopped ordeclined
many-fold (Fig. 3). The limited capacity for DNA
synthesis under hmd conditions could not be
attributed toadearth of substratesinasmuchas dATP,dGTP, dCTP,and HM-dUTPpools
actu-allyexpanded duringanhmdprogram(datanot
shown). It is worth noting that depletion of dTTPpools occurred normally underhmd con-ditions(datanotshown),which agreeswithour
observations that many early functions were
expressed normally underthese mutant
condi-tions(Fig. 1A and2A).
Intermediates inSP10replication. Inthe case
ofSPO0,severalkeyintermediates in the
replica-tioncyclehavebeenidentified
by
sucrosegradi-entanalysis (24, 25). Initially, parental DNAis
converted to a form that sediments
through
neutral sucroseatca. 300S;this is the so-called
VF ("very fast") form. Approximately 10 min
later, theVFform disappearsand isreplacedby
100,000' '
50.000
0-,o'
/C
0
10,000_
5,000
L
0/
1,000_
500 0'
0@
100 *
[image:4.505.125.411.71.247.2]0 6 12 18 24 30 36 42 48 54 Minutes postinfection
FIG. 3. Incorporation[6-3H]uracilintoSP10hmd+ andSPIOhmdl DNA.Cultures(12ml)wereinfected at37°C (29).Various timespostinfection,500-1l
sam-plesweretransferredtoafresh tube thatcontained20
p.
of[63H]uracil
(200 ,uCi/ml). The samples werelabeled for 3 min and frozen inanacetone-solidCO2
bath. At the endof theexperiment,the
samples
werethawed, and theamountof label in DNA was deter-mined(25).Symbols:
(0)
SP10hmd ;(0)
SPIOhmdl.VOL.42,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.505.297.449.441.590.2]640 WITMER AND FRANKS
.2,000
4,000 T SP15
C. S1
2,000-splo
0 1 2 3 4 5
Distance from meniscus(ml)
FIG. 4. Sampleneutralsucrosegradientprofilesof intermediates in SP10 hmd+ DNAreplication. A cul-ture was infected with SP10 hmd+ at 370C. 32p; (10
tLCi/ml)
was added at 12 min. At the times givenbelow, [32P]DNA was extracted as described in the text.(A)[32P]DNAextracted 18 minpostinfectionthat wassedimented at 11,000 rpm for 4.5 h. (B)[32P]DNA
extracted 24 min afterinfection thatwassedimentedat 11,000 rpmfor 4.5 h. (C) [32P]DNAextracted 33 min after infection that was sedimented at 11,000 rpm for 15h.Arrowsmark the position of SP10 or SP15 virion
[14C]DNA(ca. 2,000 cpm) used as marker.
the more slowly sedimenting F form composed of linearconcatenates that initially contain up to 20 phage equivalents of DNA. Mature DNA (54S) appears roughly 5 min after the F form, and astheamount of mature DNAincreases,the average size of the F formdiminishes.
Asimilarsequence of events seems adequate to explain the replicative cycle ofSP10 hmd+. Figure4depicts sample gradient profiles. Figure 5presents a detailed summary of the amount of label inVF, F, and M (mature) forms ofSP10 hmd+ DNA. For these particular experiments, cells were labeled with3
Pi
beginning at the 12thmin postinfection. At various times, extracts
were prepared andsedimented through sucrose
at areducedspeedtodemonstratethe VF and F formsor at ahigher speedtoresolve the F and M forms. The internal markers were SP10 hmd+
virion [14C]DNA (80 megadaltons; 50S) and SP15 virion [14C]DNA (250 megadaltons; about lOOS).
Until 18 min, all [3H]DNA sedimented through sucrose at a rate 2.5 to 3 times faster than did the SP15 [14C]DNA marker; this evi-dently represented the VF form of SP10 hmd+ replicating DNA (see below). Unlike the situa-tion withSPO (24),the VF intermediate ofSP10 replication usually remained evidentuntil latein the program. Between 18 and 21 min, a form sedimenting in the vicinity of SP15 [14C]DNA appeared in SP10 hmd+-infected cells which presumably correspondedtothe F intermediate. TheFintermediatereached its maximal levelat
24 min and was maintained at approximately
that level for theduration ofthe infection pro-cess. At 24 min, the F intermediate had a
nominalsedimentation coefficientof 105to120S whichcorresponded to linear duplexesofmass 350 to 500 megadaltons (9). Experiments with alkaline sucrose gradients (not shown) suggest-ed, by comparisonwith othersystems (11, 25),
60,000
45,000~
30,000[
15,000[
0 6 12 18 24 30 36 42 48
Minutespostinfection
FIG. 5. Abundance of replicative intermediates various times postinfection by SP10hmd+. Sucrose gradient analyseswerecarriedoutasdescribedin the text.Duplicate100,-ulsamples of extracted [32P]DNA
werecentrifugedateither 11,000rpmfor4.5h(VFand
Fforms)or11,000rpmfor15 h (Fand M forms). The
totalamountof radioactivityrecovered inaparticular
formisrecorded. Since theFform is definedin both regimens used, the reported amount of label in this intermediate is theaverage oftwovalues. Symbols: (0) VF; (0)F;(O) M.
I
0
II
*-
I1,
I X
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.505.71.217.57.403.2] [image:5.505.274.423.368.573.2]ROLE OF Y-Thy IN SP10 DEVELOPMENT 641 that the F intermediate was a linear
multige-nomic structure containing, at 24 min, between four andsix SP10 phage DNA equivalents. The MformofSP10hmd+ DNA appeared between 24 and 27 min postinfection and, thereafter, increased in level until it accounted for 60 to 70% of the [3H]DNA present at 51 min (i.e., 4 minbefore lysis).
Apulse-chase experiment was performed to determine the precursor relationship between VF,F,andMformsofSP10DNA.Label, as
[6-3H]HM-deoxyuridine,wasintroducedat 12 nun,
andincorporation was terminated at 17.5 min by the addition of dThd (25). Initially, all
radio-activity was recovered in the VF intermediate
(datanotshown). Between 18 and 24 min, label appeared in the Fintermediateatapproximately thesame rate atwhich itwaslostfrom the VF
intermediate.Labeldidnot appearin theMform
until 24to 27minpostinfection. These dataare consistent with the
progression
VF-3 F-- M.Under hmdconditions,replicationof the SP10 genome waslimitedto anintervalof time when VF was the only observable intermediate in normally infected cells (Fig. 3 and 4). This
impliesthat, under hmd conditions,formationof
the VF intermediate should nottake place.
In-deed,all[3H]DNAmadebetween12and 18 min
ofanhmdprogram sedimented throughsucrose
at60S, i.e., 20%6faster than matureDNA (data
notshown). The fate of this intermediate hasnot been studied.
Characterization of the VF and F intermedi-ates. The VF form was stable in sarcosine,
which indicates that binding to the cell mem-braneplays little, ifany,roleinmaintainingthe structure of this intermediate (25). Similarly,
treatmentof VFwithRNaseAhad noobvious effect on the sedimentation properties of this intermediate(Fig. 6A).On the otherhand,
treat-mentwithpronaseconvertedVF toaformthat sedimented at a rate characteristic of the F intermediate (Fig. 6B); material produced by pronase treatment of VF will be calledF*. These dataindicate thatVF represented acondensed intermediate caused by
complexing
of DNAtoprotein.
Novobiocin,aninhibitorofDNAgyrase(13),
immediatelyarrestedSP10DNA
synthesis
atalltimespostinfection(datanotshown).
Evidently,
negatively supercoiled(underwound) regions
must be maintained for continuous replication
(13). Sedimentation coefficient of
negatively
su-percoiled DNA varies with ethidium bromideconcentration in a characteristic fashion (35).
Lowdye concentrationsunwind the
supercoils,
which results in a reduction in sedimentation
coefficient;higher
dye
concentrations introducepositive supercoils with a
corresponding
in-crease in sedimentation coefficient. Forms of
DNA thatcannotbenegatively supercoiled (i.e., linear duplexes and open circles) also show characteristic changes in sedimentation coeffi-cient;inthiscase,the .sedimentationcoefficient decreases approximately linearly with increas-ing dye concentration.
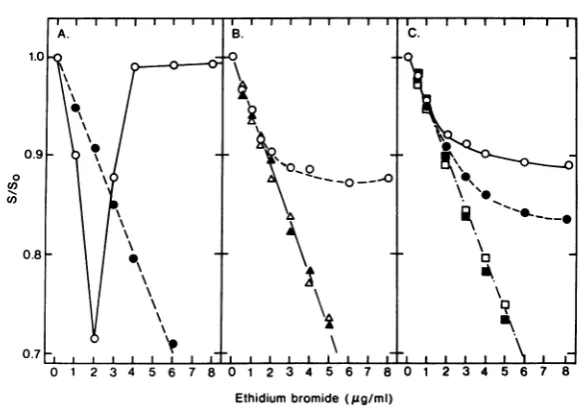
Toshow thatour system would give reliable
results, we compared the effect of ethidium bromide concentration on the Escherichia coli nucleoid (41) in its native, negatively super-coiled form and its DNase I-relaxed form (Fig. 7A). For the native form, the characteristic biphasic curve, indicative of negatively super-coiledDNA, was readily obtained, whereas the sedimentation coefficient of the relaxed form decreased linearly across therange ofdye con-centrationstested.
The F and F* forms of SP10 hmd+ DNA responded to increasing dye concentrations as
relaxedforms(Fig. 7B). VF, onthe other hand,
responded in a peculiar fashion (Fig. 7B). For dyeconcentrations ofc2,ug/ml,the sedimenta-tion coefficient of VF decreased linearly in a manner similar to that obtained for F and F*. Higher dye concentrations, however,produced
vI ,
4,000 A. SP15
2.000-0 vi
4,000*
B. 'P
2,000
-ft
SP15
0
0 1 2 3 4 5
Distance from meniscus (ml)
FIG. 6. Effect of pronase and RNase A on sedi-mentation coefficient of the VF intermediate. Cells infectedat370C with SPIOhmd+received32P,(10 F.Ci/ ml)at12min.DNAwasextractedat18 min.A100- l
portionoftheextractwastreatedwith either RNaseA
(10pg/ml)orpronase (10,ug/ml)at370C for1 hand
thencentrifugedat11,000rpmfor 4.5 h.(A)RNaseA; (B)pronase.Symbols:(0)incubatedwithoutenzyme; (0)incubated with enzyme. Arrowslocatethe posi-tion of marker[14C]DNA.
VOL. 42,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.505.299.443.365.585.2]642 WITMER AND FRANKS
'~~~~~~~~~~Al
0.8o
0_7 ,_,1_J.
01 23 4 5 6 7 8 0 1 2 3 4 5 6 7 8 0 1 2 3 4 5 6 7 8
[image:7.505.100.392.70.274.2]Ethidium bromide(pg/ml)
FIG. 7. Effectof ethidium bromideonsedimentation behavior ofreplicativeintermediates. Extracts of
32P-labeled cells received the indicated concentration ofdyeandweresedimented ingradientscontaininganidentical concentration ofdye. (A)E. colinucleoid(41) inits native,negatively supercoiledform(0)orits DNase I-relaxed form(0).(B) The F(A),F*(A),and VF(0)forms ofSP10hmd+replicatingDNA. F* isgeneratedby
pronase treatment of VF (see text). (C) DNase I-relaxed VF. Extracts were first treated with increasing
concentrations of DNase I inanice-water bath for 10 min under conditions that limit nucleolytic damageto
single-strandbreaks(41).Symbols:(0)noDNaseI;(0)0.5 ,ugof DNaseI/ml;(m)1 1Lgof DNaseI/ml; (O)2 ,g of DNaseI/ml.
nofurtherreduction inthesedimentation coeffi-cientof VF. That istosay,atcohcentrationsof dye where positive supercoiling of restrained DNA would be expected, VF underwent no furtherreduction in sedimentation coefficient.
Such data could be interpreted to mean that VF DNA contains underwound and restrained regions that are covalently joined to relaxed regions. High concentrations ofdye introduced
positivesupercoiling intothe restrainedregions
whichcompensatedforthecontinuedunwinding of the relaxed regions. If so, introduction of sufficient single-strand breaks into VF DNA should relax therestrained regions, thereby re-sulting ina structure whosesedimentation coef-ficientdeclines linearlyatdyeconcentrationsof >2 ,g/ml. Controlled incubation of VF with DNase I did, indeed, generate a form that re-spondedtoethidiumbromide concentrationas a fullyrelaxed structure (Fig. 7C).Theamountof DNase I treatment required to give a fully re-laxed formofVF DNA did not affect
the
initial sedimentation coefficient (data not presented). Therefore, ifwe are correct inassumingthat VF DNAis acondensed, protein-complexed inter-mediate, thensupercoiling makes only anminor
contributiontothe rapidly sedimentingnature of this DNA. Similar conclusions have been reached for thecondensedform of T4 DNA (17). CharacterizationofDNAmade under hmd
con-ditions. The smallamount of DNA made under hmdconditions seems to beinaform thatdoes not occurin hmd+ infections (unpublished data described above). Inasmuchashmdl andhmd2 DNAsreplaceY-Thy with different bases(39),it followed that DNAmade under hmd conditions would have abuoyantdensity in CsCl different from that of hmd+ DNA and that this density
differentialcould beused asaprobe toexplore the relationship between parental and newly replicated strands generated under hmd condi-tions.
For these experiments, cellswere infectedat 37°C with hmd phage thatcontained [32P]DNA. At 12 min, [6-3H]uracil was added. DNA was extracted 20 min postinfection and banded in CsClgradients.In the caseofhmdl,about75% of parental [32P]DNA and all of the nascent [3H]DNA were denser than the SP10 hmd+ marker [14C]DNA (Fig. 8A). In the case of hmd2, 65% of the parental [32P]DNAand all of thenascent[3H]DNAwererecoveredin a band that was less dense than hmd+ marker DNA (Fig. 8B). These data are consistent with the observation thattheuniquebase inhmdl DNA is more acidicthan Y-Thy, whereas the one in hmd2 DNAisneutral (39).
When newly replicated hmdDNA was dena-tured andrebandedin neutralCsCl,theparental [32P]DNAwasrecoveredatthe samedensityas
on November 10, 2019 by guest
http://jvi.asm.org/
ROLE OF Y-Thy IN SP10 DEVELOPMENT 643
3.000
2.000[
1,0001
3,000
2.000
IL0
1.000
01 2 3 4 5 6
W23 SP1O SPOl B. '
A_**w0~~~~~~w*-J
j|*
*~~~~~
1,500 Shiftupexperiments. Strictly speaking, there-sults shown above indicate that hypermodifica-tion is required for the first round of DNA synthesis. However, is hypermodification
con-tinuously required for DNA synthesis? To
ob-,i
tainsomeideaonthis point, shiftupexperiments<, were
performed.
Cells wereinfected with hmdphageat
30°C.
Various timeslater,asamplewas S00-2,000 removed andheat shockedat50°Cfor 2minto,
rapidly
destroy
thethermolabile function. After_1.000 < the heat shock, the samplewas placed at 37°C
and
32P,
was added. Control experimentsdem-O °0 onstrated that heatshock hadnoeffecton
hmd+
infections (datanotshown).
Only the results ofa typical experiment are
shown here (Fig. 10). In thiscase, shiftupcame
1,500 50
min
postinfection at30°C,
which is roughlyequivalentto36minpostinfectionat37°C.When
an hmd+ culture was shifted up in the manner
1,000o described, incorporation of 32p; continued for
about 21 min,atwhich timethe culture lysed. In
the hmd culture, however,the apparent rate of
500 2,000 9
0
1,000 0
O1 2 3 4 5 6 Distancefrommeniscus (ml)
FIG. 8. Isopycniccentrifugation of DNA extracted fromcells infected with SP10 hmdl or SP10 hmd2.
Bacteria wereinfected at370C withhmdphage that contained[32P]DNA. At 12 min, [6-3H]uracil (10 ,uCi/ ml)wasadded. DNAwasextracted 18min
postinfec-tion and banded in neutral CsCl density gradients (6). (A) SPIO hmdl. (B)SP10 hmd2. Symbols: (0) parental
[32P]DNA;(0) newly replicated[3H]DNA; (0)
mark-er[(4C]DNAs runinaparalleltube.
denatured hmd+ DNA, whereas the nascent
[3H]DNAwasrecovered inaseparateband(Fig.
9). [3H]DNA annealed tobothH and L strands
ofSP10 hmd+ DNA (Table 2), suggesting that
newly replicated hmdDNA contained both
com-plementary strands. Thus, itappears that repli-cation under hmd conditions generated hybrid DNAmoleculescomposedofafully
hypermodi-fiedparental strand H-bonded to anascent
un-hypermodified strand. Inotherwords, fully
un-hypermodified duplexes couldnotbegenerated
in vivo.
In ethidium bromide, material made under
hmd conditions behaved as if it were fully
re-laxed,and treatmentwith pronase hadnoeffect
on its sedimentation coefficient (data not
shown).Therefore,itisnotimmediatelyobvious
whymaterial madeunderhmdconsistently sedi-mented more rapidly than matureDNA. Many possibilitiesexist, butnonehasbeentested.
2.000
1.000
1.500
1.000 L, x
0
500-2:000
2
10 JO
FIG. 9. Isopycnic centrifugation of denatured DNAextracted from cells infected withSP10 hmdl andSPIO hmd2. This is the companion experiment to the one reported in Fig. 8. Material with adensity
different from hmd+ native DNAwas isolatedfrom pooled gradient fractions and denatured by dialysis againstdistilledwaterfollowedby heatingfor 2minin
a boiling water bath (39). The denatured DNA was rebanded inneutral CsClgradients. (A) SP10 hmdl; (B) SP10hmd2.Symbols: (0) parental [32P]DNA; (0) newly replicated [3H]DNA; (0) denatured marker [14C]DNAsruninaparalleltube.
.A. W23 S;1o 2 ISPOll
LA
I
I.q$o %1.0~.I
I I
I 3
B ~ IQ
ILA
VOL. 42,1982
NL
9 0
.00 r.
3.000
2
a.
u
(L
11
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.505.71.257.61.369.2]TABLE 2. Annealingof DNA made in vivotothe L andH strands ofSP10hmd+ DNA'
Si nuclease-resistantcpm Labeled DNALabeledDNA UnlabeledDNA
Input
cpmb Mock bHbiiehybridized" yrdzd
32P-labeledparental None 2,463 13 82
L strand 2,876 71 686
H strand 2,626 66 649
3H-labelednascent None 1,251 18 47
L strand 1,338 41 305
H strand 1,119 27 381
32P-labeledLstrandc None 2,476 41 93
L strand 2,509 73 176
H strand 2,818 81 1,572
32P-labeled H strandc None 2,011 45 87
Lstrand 2,256 53 1,703
H strand 2,256 58 201
aCellswereinfected,at
37°C,
withSP10hmdl thatcontained[32P]DNA
and labeled with[6-3H]uracil
(10,uCi/
ml)from 12 until 18 minpostinfection.DNAwasextracted and banded in neutralCsCl density gradients(Fig. 8A). All material thatbanded more heavily thanmarker SP10hmd+ DNAwascollected and denatured by dialysisagainstdistilledwaterfollowedbyheatinginaboilingwaterbath for 2 min(36, 39).The denatured DNAs were rebanded in neutral CsCl; material corresponding to parental [32P]DNA and nascent [3H]DNA was reisolated(Fig. 9). Hybridization was carriedoutin 200 mM NaCl-10mMTris-hydrochloride (pH7.9).The reaction mixtures contained'0.1,ugof labeled DNA and 25jigof unlabeled DNA. The final volumewas200 ,u. The reaction mixtureswereincubatedat67°Cfor 4 h. This isnotenoughtimetogivemaximalhybridizationto unlabeledDNA,butself-annealingof the labeled DNA isnegligible duringthis interval. The reaction mixtures weredilutedtwofold with 60 ,uMZnSO4and 250 ,ug of denatured calfthymusDNA perml. The Si nuclease of
Aspergillusoryzae(250U)wasadded. The mixturewasincubatedat50°Cfor4h,and theamountof precipitable labelwasdetermined.
b Threehybridizationmixtureswereconstructed. Onewasprecipitatedimmediatelyand usedtodetermine the
input label. The secondwas mock hybridized byincubation at 2°Cfor4h andwasthen incubated with S1
nuclease at 50°C. The third reaction mixturewashybridizedat67°Cfor 4 h anddigestedwith S1 nucleaseat50°C.
cPrepared from SP10hmd+ [32P]DNA.
DNA synthesis decreased rapidly and stopped within 10 to 12 min of the shiftup. The hmd culturelysed 30min after the shiftup.
In a subsequent experiment, the shiftup was
performedasdescribed above.After shiftup, the
cellswerelabeled with32p,for 10min.TheDNA was extracted and centrifuged through neutral sucrose gradients. Approximately 50% of the
hmd+ [32P]DNAwas recoveredas theMform,
whereas the remainder sedimentedasthe Fform
(Fig. 11A). In the case of hmd [32P]DNA, no
morethan10% of the material sedimentedasthe
Mform(Fig. llB). When denaturedand banded
in neutral CsCl gradients,thebuoyant densityof
hmd[32P]DNA made after the shiftup was
be-tween that of fully hypermodified strands and
unhypermodified strands (Fig. 12), implying
that, under these conditions, the
unhypermodi-fied DNA was covalently linked to
hypermodi-fied DNA made before shiftup. All in all, these
data are consistent with the interpretation that concatenates containingunhypermodified DNA
areinefficientlycleavedtothe Mform.
Shiftdown experiments. Shiftdown experi-mentswereperformedtoseeifrestorationof the
thermolabile hmd functions permitted
comple-tion ofthe infection process. For the sake of
comparison, dataarealso shown forSP10repd, a heat-sensitive mutant defective in an as yet undefined aspectof replication.
Inthe caseofrepd, shiftdown resulted inthe reestablishment of DNA synthesis (Fig. 13) and production of viable phage (not shown); this mutantcould be incubatedat 37°C forup to42 minwithoutaffecting the subsequent production of DNA andphage at30°Cinanymajorway. In the case of hmd mutants, normal amounts of DNA andprogeny were generated at 30°C pro-vided the shiftdown took place <12 min post-infection at the nonpermissive temperature; thereafter, the amount of DNA and progeny generatedat30°C decreasedsharply (Fig. 13 and unpublished data). Under hmd conditions, a shiftdown laterthan18min postinfectionyielded virtuallynoDNA synthesis or progeny produc-tionat 30°C (Fig. 13 andunpublished data).
DISCUSSION
Eventhough itis a minorcomponentofSP10
DNA (37, 39), Y-Thy is clearly essential for intracellular development. At nonpermissive temperatures, SPIO hmdl andSP10hmd2 gener-ateDNAs in whichdifferentintermediates
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.505.48.447.87.238.2]ROLE OF Y-Thy IN SP10 DEVELOPMENT 645
80,000
60,000
a. 40,000[
20,000
0
0 10 20
Minutesafter shift-up"
FIG. 10. Effectofa shiftuponDNA synthesis in cellsinfected with SP10hmd+andSP10hmdl.
Bacte-ria were infected at 30°C, which is a permissive temperaturefor hmdl. At 50 min, the culture (5 ml)
was transferred to a waterbath (50°C) and swirled gently for 2min. The culturewasthenplacedat37°C. Periodically, 500-p,lportions werepulse-labeled with
[6-3H]uracilasdescribed in Fig.3.Symbols:(0)SP10 hmd';(0)SPlO hmdl; (A)SPnorepd,aheat-sensitive mutantthatisdefective in DNA synthesisat37°C(see text).
mulateinplace of Y-Thy (39),yetboth mutants showbasicallythesameproperties withrespect tointracellular development (thispaperand
un-published data). Therefore, intermediatesin hy-permodification cannotsubstituteforY-Thy.
Under hmd conditions, replication of SP10 DNAstopped ordeclined manyfoldafter afew
minutes (Fig. 3). The principal, if not only, product of SP10 DNA replication under hmd conditions consisted ofduplexes composed of parental (hypermodified) strands H-bonded to nascent(hmd)strands(Fig.8 and9).DNA-DNA annealingexperiments (not shown)indicatethat hmd duplexes can be formed quite readily in
vitro, sothereis no reasontobelieve thatany
intrinsic properties ofhmd strands per se pre-ventformationof suchduplexesinvivo. Conse-quently, it seems reasonable to propose, as a
working hypothesis, that the replicational
ma-chinery of SP10 cannot use an hmd strand as
template,soreplicationstopsat thepointwhere
these strands wouldhave toserve as templates
tosustain DNAsynthesis.
Shiftdown experiments indicated that the
thermolabile hmdfunction could notbe rescued
later than 18 min postinfection (Fig. 13).
Al-though it could be thatinactivation of the
ther-molabile hmd function becomes irreversible at
18min, another explanation seems equally val-id. The time at which hmd functions become unrescuable by shiftdown corresponds to the time at which DNA synthesis stops at the non-permissive temperature (Fig. 3), i.e., the time at which hmd+-hmdhybrid duplexes are complet-ed (Fig. 8 and 9). Such hybrid duplexes are evidently inert as templates for DNA synthesis
(Fig. 3 and 10). At 37°C, hypermodification of
nascentSP10 hmd+ DNA occurs no more than 8 to 10s afterpolymerization (39). It couldwell be that hypermodification can occur only within a short time interval of DNA synthesis and that,
beyond this interval, hypermodification cannot
occureven if thehmdfunctions are restored. In thiscontext, it is worth noting that hmd+-hmd
hybrid duplexes cannot be rescued by fection with hmd+ phage even though
superin-fectingphagedevelops normally (H. Witmer and
M. Franks, unpublished data).
Insofarasreplication under hmd+ conditions
was concerned (Fig. 4 and 5), SP10 showed
basically the same intermediates as SPOl (25).
10,000 _ SP1O SP15
A. * 4
5,000
10,000 SPl
B.
5,000-0 1 2 3 4 5
Distance from meniscus(ml)
FIG. 11. Neutralsucrosegradientcentrifugationof DNA made after a shiftup. Cultures (5 ml) were
infectedat30°CwitheitherSP10hmd+orSP10hmdl. At 50 min, the cultures were shifted up to 37°C as
describedinFig.12.32P,(10FCi/ml)wasadded 1 min
aftertheculturewasplacedat37°C.DNAwas extract-ed 10 minlater and sedimented through neutral
su-crosegradientsat11,000rpmfor 15 h.Onlythe F and M forms of DNA (see text) are resolved by this
procedure. (A) SPIO
hmd';
(B) SPIO hmdl. Arrows locate thepositionof marker[14C]DNAs.
0
t
\
IE
O.I .h *...x.s
VOL. 42,1982
on November 10, 2019 by guest
http://jvi.asm.org/
[image:10.505.91.237.70.290.2] [image:10.505.299.438.356.578.2]646 WITMER AND FRANKS
a-0
ctJ
splo
spol
3 4
2,000
Ci:
1,000 c0
[image:11.505.97.394.64.221.2]
-Distance from meniscus (ml)
FIG. 12. Isopycnic centrifugationofdenatured DNA made afterashiftup.Material that sedimentedthrough
sucroselikethe Fintermediate(Fig. 11)wasreisolated,denatured,and banded in neutralCsCldensity gradients
(6, 39).Symbols: (0) [32P]DNAextracted fromSP10hmdl-infectedcells;(0)denatured marker
["4C]DNAs.
Therefore, atthepresenttime, it doesnot seem
necessary to assumethat replication of hyper-modified DNAs requires novel intermediates. However,onepointof interest should be noted.
Replication of the SP1O genome is novobiocin
sensitive, yet underwinding of SP1O DNA is
seemingly limitedtorelatively small domains in
the VF intermediate (Fig. 7B). Maintenance of theseunderwoundregions, whichweassume to be essential forreplication,requires continuous presenceofproteinsbecause F* DNA(Fig. 6B)
behaves as if it were fully relaxed (Fig. 7B).
Whencellswereinfected underhmd
conditions,
SP1ODNA assumednothing comparable to the
VFform (Witmer and Franks, ourunpublished
data), so it is possible that some proteins
re-quiredtogenerateand maintain theunderwound
regions requireafully hypermodifiedDNA. We arecurrently attempting toidentify SPlO-coded DNA-binding proteinsto seeifanybind
prefer-entiallytofullyhypermodifiedDNAs.
Consider-ing the complex and reactive nature of the substituents in hypermodified bases (37), they would be ideally suited as target molecules for DNA-protein interactions.
DHP-uracil, the hypermodified base in SP15 DNA(4), isrequired for proper DNA maturation or phage assembly (6, 36). Evidence for this conclusion stemsfrom the fact that, under con-ditions where only5% of the replicating pool has anormal DHP-uracil content, over50% of the DNA packaged into phage is normal with re-spect to base composition (6, 36). A similar conclusion seems to hold for Y-Thy in SP1O
DNAinasmuchas little, if any,hmdDNA made aftera shiftup matured (Fig. 11). Late protein synthesis isessential for DNA maturation, since
chloramphenicol, added 12minpostinfection by
hmd+ phage, allowsthe VF and Fintermediates toaccumulate but no mature DNA is generated (datanotgiven). Atpresent, it is not known if
SP1Omature DNA is cut fromtheconcatenates beforepackaging, as is the casewith SPO1 (24), or, asin thecase ofcoliphages 0X-174, T4, and
X (12,33, 38), cleavagefromtheconcatenate is the final step ofpackaging. Eitherway, one or
more late proteins must interact with the F
intermediate to generate mature DNA and,
when the F intermediate contained
unhyper-modified regions (Fig. 12), this interaction was
inefficient(Fig. 11).
60,000[
Shift-down(minutes post infection)
FIG. 13. Restoration of thermolabile functions: Shiftdown experiment. Parallel 12-ml cultures were
infectedat37°Cwith eitherSP10hmdlorSP10repd
(see text). Varioustimes postinfection, 1-ml samples were transferredto afresh tube and incubated, with
gentleagitation,at20°C for2min. The sampleswere
placedat30°C, and32Piwasaddedtoafinal
concentra-tionof10,Ci/ml.The sampleswereincubatedat30°C until all the SP10 repd-infected cultures lysed. The
amount of label in DNAwas determined (23). SP10 hmdl-infected culturesfailedtolyse orlysedpoorly if
shiftdown came later than 18 min after infection at
37°C. Symbols: (0) SP10repd; (0)SP10hmdl.
-jr--a-I, E
1;--111
\\
IIz
I *
2
(L
L)
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.505.270.420.395.556.2]ROLE OF Y-Thy IN SP10 DEVELOPMENT 647
Thus far, it is clear that Y-Thy is required for replication as
well
as cleavage of mature DNAfrom concatenates. In turn, it is proposed that both defects arise through faulty interaction of certain key proteins with unhypermodified DNA. It is difficult to judge from the available data if Y-Thy plays a role in differential tran-scription of theSP10genome. Although it is true that SPIO late genes were weakly expressed under hmd conditions (Fig. 2), this could just as easily be attributed to the low levels of DNA synthesis that occur under those conditions (Fig. 3), since late transcription of numerous phage genomes is, to some extent, coupled to
replica-tion (15, 20). The importance of unusual bases in differential gene expression varies somewhat from system to system. In the case of coliphage T4, late genes can be transcribed only from an HM-cytosine-containing template (42). Indeed, the alc gene product of phage T4 seems to modify
cellular
RNA polymerase such that HM-cytosine-containing DNA is a preferredtemplate (34). On the other hand, Thy can substitute for, at least, 20% of the HM-uracil in 4e DNA without affecting phage functions (28).Similarly, Thy can replace up to 70% of theDHP-uracilinSP15DNA without affecting gene expression (6, 36). Thus, there is no a priori reason to believe that Y-Thy would play a direct role in transcrip-tion of the SP10 genome.
ACKNOWLEDGMENT
This research was funded by grant PCM 79018-03 from the National Science Foundation.
LITERATURECITED
1. Alegria, A. H. 1967. Hydroxymethylation of pyrimidine nucleotides with formaldehyde. Biochim. Biophys. Acta 149:317-324.
2. Bolle, A., R. H. Epstein, W. Salser, and E. P.Gelduschek. 1968. Transcription during early bacteriophage T4 devel-opment: synthesisand relative stability of early and late RNA. J. Mol. Biol. 31:325-348.
3. Bott, K., and B.Strauss.1965.The carrier state ofBacillus subtilisinfected with the transducing bacteriophageSP1O. Virology 25:212-225.
4. Brandon,C., P. M.Gaflop,J.Marmur,H. Hayashi,and K. Nakanishi. 1972.Structure of a new pyrimidinefrom Bacillussubtilisphage SP-15. Nature (London) New Biol. 239:70-71.
5. Brown, N. C. 1970. 6-(p-Hydroxyphenylazo)uracil: a
se-lective inhibitor of host DNAreplicationinphageinfected Bacillussubtilis. Proc. Natl. Acad. Sci. U.S.A. 67:1454-1461.
6. Csella,E.,0.Markewych,M.Dosnr, and H.Witiner. 1978. Production and expression of dTMP-enriched DNA of bacteriophage SP15.J. Virol. 28:753-766.
7. Dawid, I. B., D. D. Brown, and R. H. Reeder. 1970. Composition and structureofchromosomal andamplified ribosomal DNA's of Xenopus laevus. J. Mol. Biol. 51:241-260.
8. DeMars, R.I. 1955. The production of phage related material when bacteriophage developmentisinterrupted by proflavine. Virology 1:83-99.
9. DoneUl, G., E. Dore, C.Frontali, andJ.Grandolfo. 1975.
Structure and physico-chemical properties of bacterio-phageGIll. A homogeneous DNA of molecular weight 5
x 16§.J. Mol. Biol.94:555-565.
10. Dosmar, M., and H. Wltmer. 1979. An atypical deoxynu-cleoside triphosphate in Bacillus subtilis infected with bacteriophage SPlOc. Curr.Microbiol. 2:361-364. 11. Frankel,F. R.1968. Evidence for long DNA strands in the
replicatingpool after T4 infection. Proc. Natl.Acad. Sci. U.S.A.59:131-138.
12. Frankel, F., M. Batcheler, and C. Clark. 1971. The role of gene 49in DNA replication and head morphogenesis in bacteriophageT4. J. Mol. Biol. 62:439-463.
13. Gdlert,M., M. O'Dea, T.Itoh, and J.Tomizawa. 1976. Novobiocin and coumermycin inhibit DNA supercoiling by DNA gyrase. Proc. Natl. Acad. Sci. U.S.A. 73:4474-4478.
14. GIllespie, D., and S. Splgeman. 1965. A quantitative assayfor DNA-RNA hybrids with DNAimmobilizedon a membrane. J. Mol. Biol. 12:829-842.
15. Glamberg, J., R. A.Somiany, and C. R. Stewart. 1977. Selective screening procedure for the isolation of heat-andcold-sensitive, DNA replication-deficient mutantsof bacteriophage SPOl and preliminary characterization of the mutants isolated. J. Virol. 21:54-60.
16. Guha, A., and W. Szybalski. 1968. Fractionation of the complementary strands of coliphage T4DNA based on the asymmetric distribution of the PolyU andPoly U, G bindingsites. Virology 34:608-616.
17. Hamilton, S., and D. E. Pettljohn. 1976. Properties of condensed bacteriophage T4 DNAisolated from Esche-richia coli infected with bacteriophage T4. J. Virol. 19:1012-1027.
18. Hemphill, H. E., and H. R. Whiteley. 1975. Bacterio-phages of Bacillus subtilis.Bacteriol. Rev.39:257-315. 19. Jayaraman,R., and E. B. Goldberg.1969. Agenetic assay
for mRNA's of phage T4. Proc. Natl. Acad. Sci. U.S.A. 64:198-204.
20. Kahan, E. 1966. A genetic study oftemperature-sensitive mutants of thesubtilis phageSP82.Virology 30:650660. 21. Kropinskl, A. M. B., R. J. Bose, and R. A. J. Warren. 1973. 544-Aminobutylaminoethyl)uracil, an unusual py-rimidine fromthedeoxyribonucleicacid ofbacteriophage 0W-14. Biochemistry12:151-157.
22. Legault-Demare, L., and G. H.Chambliss. 1974. Natural messenger ribonucleicacid-directed,ceil-free protein syn-thesizing system of Bacillus subtilis. J. Bacteriol. 120:1300-1307.
23. Lembach, K., and J. M.Buchanan.1970.Therelationship of protein synthesis to early transcriptive events in bacte-riophageT4-infected Escherichia coli B. J. Biol. Chem. 245:1575-1587.
24. Levner, M. H. 1972. Replication of viral DNA in SP01-infected Bacillus subtilis. II. DNA maturation during abortiveinfection. Virology 48:417-429.
25. Levner, M. H., and N. R. Cozzarefl. 1972.Replicationof viral DNAinSPOl-infectedBacillus subtilis.I. Replica-tiveintermediates.Virology 48:402-416.
26. Lowry, 0. H., N. J.Rosebrough, A. L. Farr,and R. J. Randall. 1951. Protein measurement with Folin phenol reagent.J. Biol. Chem. 193:265-275.
27. Maltman, K. L., J. Neuhard, H. A.Lewis, and R. A. J. Warren. 1980. Synthesis of thymine and a-putrescinylthy-mine in bacteriophage 0W-14-infected Pseudomonas aci-dovorans. J. Virol. 34:354-359.
28. Marcus, M., and M. C. Newlon. 1971. Control of DNA synthesis inBacillus subtilisbyphageoe.Virology 44:83-93.
29. Markewych, O., A.Boghosan, M.Dosmar, D.Ende, and H. Witmer. 1977. SP-10 bacteriophage-specific nucleic acid and enzyme synthesis in Bacillus subtilis W23. J. Virol. 21:84-95.
30. Markewych,O.,E. Casella,M.Dosmar,and H. Wltmer. 1979. Deoxythymidine nucleotide metabolism inBacillus subtilis W23 infected withbacteriophage SPlOc: prelimi-naryevidence that dTMP inSPlOc DNA issynthesizedby VOL.42,1982
on November 10, 2019 by guest
http://jvi.asm.org/
648 WITMER AND FRANKS
anovel,phage-specific mechanism. J. Virol. 29:61-68. 31. Neuhard, J., K. L.Maltman,and R. A.J. Warren.1980.
Bacteriophage 0W-14-infected Pseudomonas
acidovor-ans synthesizes hydroxymethyldeoxyuridine triphos-phate. J. Virol. 34:347-353.
32. Neville,M.M., andN. C. Brown.1972. Inhibition ofa
discrete bacterial DNA polymerase by 6-(p-hydroxyphen-ylaso)-uracil and 6-(p-hydroxyphenylazo)-isocytosine. Nature(London) NewBiol. 240:80-82.
33. Sinsheimer, R. L.,R. Knippers, and T. Komano. 1968. Stages in thereplication of bacteriophage 0W174 DNAin
vivo.Cold Spring Harbor Symp. Quant. Biol. 33:443-448. 34. Snyder, L., L. Gold, and E. Kutter. 1976. A gene of
bacteriophage T4 whose productprevents truelate
tran-scription on cytosine-containing T4 DNA. Proc. Natl. Acad.Sci. U.S.A. 73:2098-2102.
35. Vinograd,J., J.Lebowitz,R.Radloff,R. Watson, and P.
Laipis.1965. The twisted circular form of polyoma viral DNA. Proc.Natl. Acad. Sci. U.S.A. 53:1104-1111. 36. Walker, M. S.,andM.Mandel.1978. Biosynthesis of
5-(4',5'-dihydroxypentyl)uracilas anucleosidetriphosphate inbacteriophageSP15-infected Bacillus subtilis. J. Virol.
25:500-509.
37. Warren, R.A.J. 1980.Modified bases in bacteriophage DNA's. Annu.Rev.Microbiol. 34:137-158.
38. Weissbach, A., P. Barti, and L. Salzman. 1968. The structureofreplicative lambda DNA-electron microscope studies.Cold Spring Harbor Symp. Quant. Biol. 33:525-532.
39. Witmer, H. 1981. Deoxythymidylate and the unusual nucleotide inmatureSP10 phage DNAaresynthesized by postreplicational modification of 5-hydroxymethyldeoxy-uridulate. J. Virol. 39:536-547.
40. Witmner, H., and M. Dosmar. 1978. Synthesis of 5-hydroxymethyldeoxyuridine triphosphate in extracts of SPlOc-infected Bacillus subtilis W23. Curr. Microbiol. 1:289-292.
41. Worcel,A., andE. BurgL.1972. On thestructureof the folded chromosome of Escherichia coli. J. Mol. Biol. 71:127-147.
42. Wu,R., and E. P.Geiduschek. 1975.Therole of replica-tionproteins in theregulation of bacteriophage T4
tran-scription. I. Gene 45 and hydroxymethyl-C-containing
DNA.J. Mol. Biol. 96:513-538.
on November 10, 2019 by guest
http://jvi.asm.org/
![FIG. 2.conditions.bylabeledheat-denaturedwascompetitionUnlabeledcompetitor,(0)20-minsimultaneously DNA-RNA hybridization competition between early and late SP10 RNAs made under hmd' and hmd Early [3H]RNA was labeled from 4 until 12 min postinfection by hmd](https://thumb-us.123doks.com/thumbv2/123dok_us/1461989.98874/4.505.297.449.441.590/conditions-bylabeledheat-denaturedwascompetitionunlabeledcompetitor-minsimultaneously-hybridization-competition-labeled-postinfection.webp)
![FIG. 4.intermediatesbelow,text.tureextractedwasaftertLCi/ml)[14C]DNA11,00015 Sample neutral sucrose gradient profiles of in SP10 hmd+ DNA replication](https://thumb-us.123doks.com/thumbv2/123dok_us/1461989.98874/5.505.274.423.368.573/intermediatesbelow-tureextractedwasaftertlci-sample-neutral-sucrose-gradient-profiles-replication.webp)

![FIG. 9.DNAanddifferentpooledagainsttherebandeda(B)newly[14C]DNAs boiling Isopycniccentrifugationof denatured extracted from cells infected with SP10 hmdl SPIO hmd2](https://thumb-us.123doks.com/thumbv2/123dok_us/1461989.98874/8.505.71.257.61.369/dnaanddifferentpooledagainsttherebandeda-newly-boiling-isopycniccentrifugationof-denatured-extracted-infected-spio.webp)


