Copyright i 1974 American Society for Microbiology Printed in U.S.A.
Role
of
Genetic Recombination
in DNA
Replication
of
Bacteriophage Lambda
I.
Genetic Characterization
of
the
Delta
Gene
K. BARTA, P. TAVERNIER, AND J. ZISSLER
Department ofMicrobiology, University of Minnesota Medical School,Minneapolis, Minnesota55455
Received forpublication 12February 1974
We describe the isolation and genetic characterization of point mutations in
genedelta, includingatemperature-sensitive mutation
(del206,).
Genetic methodsenable the extraction ofadelta mutation from the triplemutant (del,red,gam)
and the construction of new genotypes, including del,red and del,gam double
mutants. Tests of plating efficiency indicate gene delta is essential for normal
phase growth on the polA host. The possible association of delta in a system
involving alpha, beta, andgamma is considered.
Bacteriophage lambda encodes proteins
which act in genetic recombination (the Red
system).Theredalpha gene codes for X
exonu-clease (2, 7-9, 11, 12), and the red beta gene
codes for beta protein (2, 8, 11, 12). The
phage gamma gene codes for gamma protein (15), which interacts with the host
recombina-tion protein, recBC nuclease (10); gamma also
mayparticipate in the A recombination process.
Although theseproteins function in
recombi-nation, it is clear that they also affect DNA
replication (3). Phagesmutant inAexonuclease
orbeta protein make somewhat lower levels of
DNAthanX+ (approximately50% ofXA in rec+
hosts), and concatemers are somewhat shorter
than thoseobserved forX+.
Phages mutant in gamma also make lower levels of DNAthan X+ (30%). Gamma mutants
makesignificantly fewer concatemers than
X+,
andtheycontinue to make throughout infection
early replication intermediates consisting of
nicked circles and supercoils. These results
suggest recombination functions might act
di-rectly in DNAreplication, possibly topromote formation of concatemers (3).
In aseries of experiments (1), we are
investi-gating the role of the delta gene in DNA replication. In this paper are reported the isola-tion and characterization ofpoint mutations in gene delta, and the observation that delta
mutants plate with somewhat reduced
effi-ciencyonhostsdeficientinDNApolymerase I. Together with biochemical studies on DNA
replication reportedelsewhere (1), theseresults
suggest that gene delta may be an additional
component in a system
involving
genes for Xexonuclease, beta protein, andgamma
protein.
MATERIALS AND METHODS
Media, procedures, and phages have been
de-scribed previously (12). The bacterial strains used
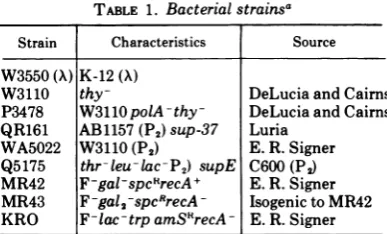
here aredescribedinTable 1.
RESULTS
Isolation of delta mutants. Phage lambda
failstoplateon
lysogens
ofthe unrelatedphage
P2; thisphenotypeiscalled
Spi+ (sensitive
toP2 interference) (5). Lambda phages able to plateon P2
lysogens
(Spi-)
wereoriginally
detected by Lindahl et al.(5)
asbiotin-transducing
phages deleted for lambda genes between the
attsiteand the CIII gene (Fig. 1). Zissleret al.
(15)
subsequently
demonstrated that mutationsin three genes are requiredforthe
Spi-
pheno-type. Theseare mutations ina redgene
(alpha
or beta), in the gamma gene, and in the delta
gene.
In previous
studies,
the delta gene was de-finedby
studies using the bio deletionphage,
bio7-20,
which removes genes between the attsite and the gene for A exonuclease
(Fig. 1).
Thus,aseries of
Spi- phages
wasconstructedofthe general family: bio7-20,
red,gam.
Thesephages are notsuitableforgeneticor biochemi-cal studies to elucidate the function of gene
delta because the bio7-20
phage
is deleted forseveral genes, and because Xbio
phage
behaveanomalously in other respects,
perhaps
due tothe bio insertion (see [6]).
Therefore,
we setout(i) to isolate a series ofpointmutations in
delta,
1445
on November 10, 2019 by guest
http://jvi.asm.org/
TABLE 1. Bacterial strainsa
Strain Characteristics Source
W3550 (X) K-12(A)
W3110 thy- DeLuciaandCairns
P3478 W3110polA-thy- DeLucia andCairns
QR161 AB1157(P2)sup-37 Luria
WA5022 W3110 (P2) E. R.Signer
Q5175 thr-leuilac-P2) supE C600 (P2)
MR42 F-gal-spcRrecA+ E. R. Signer
MR43 F-gal2-spcRrecA- Isogenic to MR42
KRO F-lac-trpamSRrecA E. R.Signer
aGenetic symbols used: K-12, E. coli K-12;
(X),
lysogenic for phage A; thy-, nutritional requirement
for thymine; polI, deficiency in DNA polymerase
I; (P), lysogenic for phage P2; sup, carries amber
suppressor; thr-, nutritional requirement for
threo-nine; leu-, nutritional requirement for leucine; lac-,
unable to ferment lactose; gal-, unable to ferment
galactose; F-, female strainmissing Fsexfactor;spcR,
spectinomycin resistance; recA-, deficient in
recom-bination; trpam, suppressible nutritional
require-ment for tryptophan; and SI', streptomycin resist-ance.
and (ii) to construct a new series of phages
carrying delta mutations in combination with other mutations.
Ethyl methane sulfonate-mutagenized stocks
ofgam phage were plated on P2 lysogens (see
Table 1 for strains). Small plaques arise from
mutagenizedgam mutants at afrequencyof2x
10-'. Thisisapproximately a 100-fold
stimula-tion in mutastimula-tion ascompared with
unmutagen-ized stocks. Analysis of these Spi- mutants
from Agamphage shows that these phages carry
three mutations: the original gam mutation, a
mutation indel, and athirdmutation ineither
alphaorbeta.
Spi- phage derived from Fec-
(red,gam)
phage are obtainedwith difficulty because the leaky growth ofthered,gam phage reduces the
background
against whichaSpi- plaquecanbe visualized. However, when ideal platingcondi-tions are achieved small Spi- plaques are
de-tected.
Tem perature-sensitive del mutations were
obtainedby a different technique. We
mutagen-ized with ethyl methane sulfonate
beta270gaM210, which has an amber mutation in
bothbeta and gamma. In Su- P2 lysogens, this
phage is already partially Spi-. In Escherichia
coli strain QR161, which is a P2 lysogen of
AB1157 carrying the weak amber suppressor
sup-37,
beta270gaMi210
is unable to form aplaque. Since sup-37 incompletely suppresses
the beta and gam mutations, the phage is still
partially beta,gam. Nevertheless, partial
sup-pression of beta,gam eliminates the leaky
growth of this phage on Su- P2 lysogens. This
enables us to detect more easily Spi- phage
which contain new mutations in delta. Among
these phage tested, approximately 1% are spi
at40C and spi+ at32C.
All the delta mutationsdescribed hereappear
tobe of thesamegeneralclass.They constitute
a Spi- phenotypein combinationwith redand
gam mutations, and they allextractand
back-cross as described below. At a much lower
frequency (106 to 10-8), Spi- phages arise
which are also Fec+. These may be A reverse,
which are also fec+ because they contain an
insertion of new recombination genes ([16]);
Zissler, unpublished data). Another subclass
ap-pears to have a chi mutation such as that described originally by D. Henderson (personal communication; [4] and [8]). By genetic and
physiological tests, chi mutations differ from
the del mutations described here (Barta and
Zissler, unpublished data).
Extraction of the del mutation from the
del,beta, gam triplemutant. To have the del
mutation by itself, we extracted it from the
triple mutant del,beta,gam which have been
isolated previouslyas aSpi-phage (see above).
The extraction involves a cross between the
del,beta,gam phage and Xbio72 (Fig. 2). Each
parentfails toplateonthe polA host (deficient
in DNA polymerase I). The del,beta+,gam+
recombinant canbe isolatedas aplaque onthe
polA host (even though the efficiency of plating
(EDP) of the isolated delta mutant onpolA is
0.03 [see below]). The deltamutantextracted in
thiswaywasthen confirmedbyabackcross; the
cross between del and beta,gam produces Spi-phage. The delta mutations which have been
extracted aregiven in Table 2.
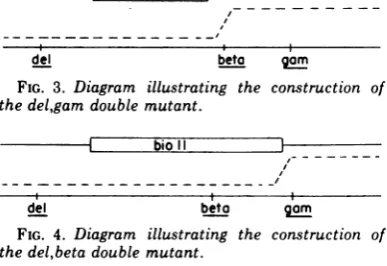
Construction of the del,gam double
mutant. We constructed new genotypes with
the delta mutation in combination with other
mutations because we hoped that additional
del b2 Ott imt alpha betagum Cm NCI
bio
17.2
biobio 11
b2 I
I b13191
FIG. 1. Map oftheAchromosomeshowingtheorder
ofrelevant genes.
-4
del beta gam
FIG. 2. Diagram illustrating the extraction of del.
1446
on November 10, 2019 by guest
http://jvi.asm.org/
[image:2.499.265.459.499.663.2]TABLE 2. Genotype and growth properties of 6 mutantsa
Phenotype Growth
Phage
6(delta) Exo(exo) | (beta) -y(gamma) polA recA (P,)lysogen Single
62 NC + + + + ++
_-61601 NC + + + + ++
_-6206 ts + + + + ++
_-Double
62Y5
NC + + NC - ++-627210 NC + + AM11 - + +
-62.113 NC + NC + - ++
-640175 NC + + NC - ++
-64017201 NC + + AM11 - + +
-620.75 ts + + NC _ ++
-620,7210 ts + + AM11 - ++
-Triple
6201137210 NC + NC AMW - __ +
6206 2707210 NC NC + AM11 - _- +
61575 NC ts + NC -
_-
+62red329Y210
NC NC +AM,1
- -- +6401#11375 NC + NC NC - __ +
6401161137210
NC + NCAM,1
- _ +6401redlY210
NC NC +AM,1
-_-
+6206011837
ts + NC NC - __ +620&82?0Y210
ts + AM,, AM,, -I
+aGenetic symbols used: NC, non-conditional mutation; AM1, suppressible by supE (carries amber
suppressor); ts, temperature-sensitive mutation; +, forms small- to medium-size plaque; ++, formslarge
plaque;and -, forms no plaque.
cluestothefunction of delta could beobtained fromdouble mutants.
Toconstructthe
del,gam
doublemutants, wecrossed the triple mutant
del,beta,gam
andXbio72,
gam. Each of the parents fails toplate
on the recA
host;
thedel,gam
recombinant,
however,
isFec+ andplates
onthis host(Fig. 3). The genotype of thedel,gam
mutant, onceisolated, was confirmed
by
additional tests;crosses between the
del,gam
mutantand a redsingle
mutantproduce
thetriple
mutantdel,red,gam
at the expected frequency. Thedel,gam
double mutant checked as having thegam mutation because it plated on the
polA
hostwith an
extremely
lowefficiencyofplating
EOP
(10-8).
Construction of the
del,beta
doublemutant. We constructed
del,beta
doublemu-tants
by
crossing the triple mutantdel,beta,-gam with Xbioll (Fig. 4). Each parent fails to
plate on the recA host, i.e., is Fec-. Since the gaM210 mutation maps to the right ofthe bio end point, the cross produces Fec+
recombi-nants which are
del,beta.
The genotype oftheFec+ mutants selectedinthis way isconfirmed
because crossesbetween
del,beta
and gam pro-duceSpi-
phage. The presence of the betabio72 (aam
[image:3.499.252.446.404.537.2]del beta 22m!
FIG. 3. Diagram illustrating the construction of
the
del,gam
doublemutant.bio~~~~~~
del beta gaOi
FIG. 4. Diagram illustrating the construction of
thedel,betadouble mutant.
mutationin the double mutant is indicated
by
thelow EOPofthedouble mutantonthepolA
host (Table3).
Summary
of delta mutants constructed.The mutations in deltanowavailablearelisted
inTable 2. Deltamutations del2 and
del40,
arenonconditional, whereas del20, is conditional.
(Mutation
del20,,
istemperature-sensitive
indelta.)
Genotypes
involving
delta pointmuta-tions in combination with other mutations are
also listed. Fordouble mutants, different delta
1447
I
on November 10, 2019 by guest
http://jvi.asm.org/
BARTA,TAVERNIER, AND ZISSLER
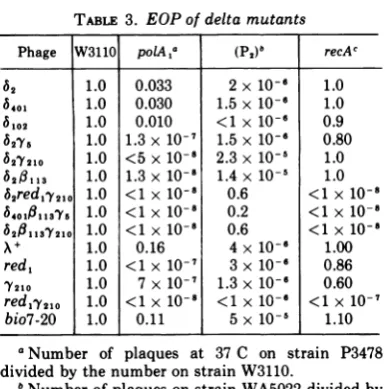
TABLE 3. EOPof delta mutants
Phage W3110 polAa (P2)8 recAc
62 1.0 0.033 2x 10-6 1.0
6401 1.0 0.030 1.5x 10-6 1.0
6102 1.0 0.010 <1x 10-6 0.9
6275 1.0 1.3x 10-7 1.5 x10-6 0.80
62Y210 1.0 <5x 10-8 2.3x10-5 1.0
62 113 1.0 1.3 x 10-8 1.4 x 10-5 1.0
62redl'Y210 1.0 <1 x 10-8 0.6 <1 x
10-6401011375 1.0 <1 X 10-8 0.2 <1x 10-1
6201137210 1.0 <1X 10-8 0.6 <1 x 10-8
A+ 1.0 0.16 4x 10-6 1.00
red,
1.0 <1X 10-7 3x 10-6 0.867210 1.0 7x 10-f 1.3x 10-1 0.60
redl.Y210
1.0 <1X 10-8 <1 X 10-6 <1 X 10-7bio7-20 1.0 0.11 5 x 10-5 1.10
aNumber of plaques at 37C on strain P3478
dividedby the numberonstrain W3110.
bNumber of plaques on strain WA5022 dividedby
the number on strain W3110.
cNumber ofplaques on strain MR43dividedbythe
number on strain MR42.
point mutations exist in combination with a
beta mutation (i.e.,
del,beta)
orwitha gammamutation (i.e.,
del,gam),
wheregamma iseither nonconditional (gaM5) or conditional (gaM210,an amber mutation suppressed by SuII). Triple
mutants are listed, which mutants have
differ-ent delta mutations reconstructed with various
other
red,gam
combinations. The type of the point mutation in any gene (conditional, ornonconditional) is given for each mutant.
Role ofgene delta in
phage growth.
The delta mutants constructed in crosses (see above) are listed in Table 2,together
with the phenotype for growth on various hosts. TheEOP valuesare presented inTable 3.
Whereas the red or gam mutants have an
extremely low EOP on the
polA
host (10-7), delta mutants have a higher EOP (3 x 10-2, Table 3). The delta mutants, however, have alowerEOPonthe
polA
host than A+. The EOPofdelta mutants on
polA
is 0.03, whereas theEOP of X+ under these conditions is 0.16. In
addition, delta mutantshave alower burstsize
(8) than A+ (62) in the
polA
host.Table 4 gives data for the burst size ofthe del,gam double mutant on the recA host. These data show that the del mutation in
combinationwith agam mutation (either gaM5
orgaM210) significantly increases the burst size
inthe recA hostfrom the low level observed for
the gam control. This indicates that the
in-creased growth observed for del,gam does not
depend on an intact host rec system. This
contrasts with the gam,chi mutant, which
others (D. Henderson and J. Weil, unpublished
data)have shown doesrequireanintact hostrec
system for increased growth.
Mapposition of delta. The delta
point
muta-tionsdescribed hereappear to maptothe left of
alpha. We conclude this form crosses between
del,red,gam and Xbio72 (Fig. 2).Ten
independ-entrecombinantsselected from these crosseson
a polA indicator were del, as tested in
subse-quentbackcrosses withred,gam phageto create
Spi-
phage. This indicates the del mutationsare to the left of the beta113 mutation and are
possibly to the left of the deletion endpointin
Xbio72.
We obtained furthermappinginformation in
the crossbetween del mutants(del2 and
del206)
and the deletion phage, A b1319,CI857 (Fig. 5). Mutant A b1319,CI857, which produces a min-uteplaqueon aP2lysogen,isdeletedfor A genes
from the att site to gene CIII. Del mutations
successfully backcross with A b1319,CI857 to
producealarge Spi- plaqueon aP2 lysogenat a
frequency of 1%. This maps del mutations outside the regionfrom attto CIII.
The del2 and
del201
mutations can bereex-tracted from the corresponding
del,b1319
re-combinant with a crossbetween
del,b1319
andXsusJ,6.
Neitherparent grows ontherecA strainKRO. Recombinants (dashed line in Fig. 6) arise on this host at a frequency of 1 to 2%. Backcrossing ofthese recombinants with either b1319 or
red,gam
againyields Spi-
phage,
indicating that del is to the leftand outsideof
the b1319 deletion.In thecase of
del206
(atem-perature-sensitive del mutation), only crosses
plated at 41 C yield Spi- recombinants. Thus, thetemperature-sensitive marker ofthe
original
isolate maps to the left and outside the b1319 deletion. Crosses between
red,gam
and A b2, aTABLE 4. Burst-sizeafter infection ofarecAhosta
Phage (5.0 input phage Infective Yield Burst
per cell) centers" size'
del2gam5 0.75 8.8 117
gami 0.91 5.3 58
del2gam210CI857
1.30 6.3 49gam210CIs57
1.50 3.0 20aStrain KRO(recA
Su-)
wasgrown to108/mlandinfected. After 15 min of adsorption at 37 C, the
mixture was washed in warm broth, diluted
1,000-fold, grown for 90 min at 37 C, and treated with chloroform.
'Number of cells releasing at least one phage,
measured at 15 min after infection. Number equals
valuetimes 10-7.
'Values to bemultiplied by
10-6.
dCalculatedastheratioofyield to infective centers
x 1,000(dilution factor).
1448
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.266.459.480.563.2][ b 1319 Ott
FIG. 5. Diagram illustratingthe back
mutation tothe deletionmutant b1319.
d.cJ
Ott
b 1319
SUsJ
FIG. 6. Diagram illustrating the extrc
from
thedel,b1319
recombinant.phage deleted for 13% of thegenome
ofatt, fail toyield Spi- phage,sugg
del mutations maptothe left of this
DISCUSSION In this paper, we describe the i
point mutations ingene delta. Ate(
extracting the delta mutation from
mutant (del,red,gam) permits isola
del mutation itself. Other techni(
construction of thedelmutation inc
with red mutations (del,exo; del,i
gamma mutations (del,gam) and wi gamma mutations (del,red,gam). Si genes may act with delta insome pi
helpful to study the delta gene in
where theproducts of othergenes (a
and gamma) also can be controlled
tion.The series ofpointmutantsdes
make this possible.
The Spi- phenotype of the
trip
(del,red,gam) implicates delta in so
involving the function of the red s
gamma. First, however, consider t gamma. Gamma protein has beer
inhibit directly the host recBC nu
Sironi et al. (13) discovered that r(
tants are killed by prophage P2 ift]
old+ but not if the phage is old
thesefactssuggest that infection of
by Xgam+ converts the host to
phenocopy (13) leadingtothe death
and perhaps also the infecting A.
Since a gamma mutation is requi
Spi- phenotype, DNA replication b
P2 lysogen should be restricted to
modeproposed byEnquist and Skall
model could provide a clue to the
delta inthe Spi phenomenon. In the
gamma, delta inhibits phage growth. Thus,
when concatemer synthesis is prevented by a
gam mutation, delta might in some way block
theDNA replication occurring in theabnormal
cm
mode.cm'
The low burst size ofdel mutants on polAcross
of
adel strains issignificantinterms of the possible roleof delta in DNA replication. Other mutants
such as red orgam fail to grow on polA hosts
c}m
(15) and are defective in DNA replication inthese hosts (L. W. Enquist and A. Skalka, personal communication).
Inthe recA host, delta also has a phenotype
and inhibits phage growth when gamma is
action of del
missing.
Perhaps
deltaaffectsDNAreplication
inthis case, since X replication isabnormal in
the recA host whengamma is missing (3).
DNAcanbecut intomonomersduring
matu-tothe left ration
only
if it contains at least two cosresting
that sequences, andthustheminimumpackageable
deletion. DNA size must be longer than a monomer; when concatemer synthesis in a recA host is
blocked by a gamma mutation, circular DNA
Lsolation
ofmight
be recombined to a DNAform thatcanchnique
for bepackaged,
and this could occurby phage
then
triple
recombination functions alpha andbeta (3). Ifti
ofthe
delmutations increase gam phage production inques
allow a recA hostthrough
an effect onreplication
)mbination
(possibly through
an effect on circleproduc-beta)
with tion),we therefore mightexpectredgenes tobeith
redan
essential, perhaps for making packageabletnce
severa
DNA. Infact, weobservethatredgenes arestillrocess, it is essential for
phage growth,
because thedel,red,-situations gam
triple
mutant fails to grow on the recAtlph,
bta,host.
i by muta- A paradox arises in the Spi phenomenon,
cribed
here because the phage recombination proteinsalpha
and beta apparently do not promotele mutant phage growth. If
circles
predominate in theime process replication of
Xspi,
it is not clear whythe Redmesprocess
system (which "rescues" the DNA in a recAthe
role ofhost)
islethal in theP2
lysogen.
a found to ACKNOWLEDGMENTS
clease (10). We thank PhillyPengforexcellent technical assistance.
ecBC- mu- Thisinvestigationconstitutespartof the thesis submittedby
he
phage is K. Barta in partial fulfillment ofthe requirementsfortheTogether Ph.D.degreefromtheUniversityofMinnesota,Minneapolis, Minn.
P2lysogens This investigation was supported by research grant a recBC- GB20677A1 from the National Science Foundation and a
ofthe host grantfrom the Graduate SchooloftheUniversityof
Min-nesota.K. Barta and P. Tavernier were supportedbyU.S. Public HealthService traininggrant No.AI-00090fromthe ired for the National Instituteof Allergyand Infectious Diseases.
oy Xspiin a
the circle LITERATURE CITED
ka(3). This 1. Barta, K., and J.Zissler. 1974. Role ofgenetic
recombina-tion in DNA replicarecombina-tion of bacteriophagelambda.II.
function of Effect in DNA replication by gene delta. J. Virol.
> absenceof 14:1451-1457.
1449
-.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.47.240.56.233.2]BARTA, TAVERNIER, AND
2. Echols, H., and R. Gingery. 1968.Mutants of bacterio-phage defective in vegetative geneticrecombination.J. Mol. Biol.34:251.
3. Enquist, L. W., and A. Skalka. 1973. Replication of
bacteriophagelambdaDNAdependentonthefunction ofhost andviralgenes. I.Interaction ofred,gam and
rec.J.Mol. Biol.75:185.
4. Lam,S., M. M. Stahl,K. D.McMilin,and F. W.Stahl. Rec mediated recombinational hot spot activity in bacteriophagelambda.II. A mutation whichcauseshot
spotactivity. Genetics77:425.
5. Lindahl, G., G. Sironi, H. Bialy,and R.Calender. 1970. Bacteriophage lambda: abortive infection ofbacteria lysogenic for phage P2. PNAS 66:587.
6. McMilin, K. D., M. M. Stahl, and F. W. Stahl. Red mediated recombinational hot spotactivity in bacte-riophagelambda.I. Hotspotactivity associated with
Spi- deletions and bio-substitutions. Genetics 77:409. 7. Manly, K. F., E. R. Signer, and C. M. Radding. 1969. Nonessential functions of bacteriophage A. Virology
37:177.
8. Radding, C. M., and D. C. Schreffler.1966.Regulationof
Aexonuclease. II. Increased synthesisoftwoproteinsby A+. J. Mol. Biol. 18:251.
9. Radding, C. M., J. Szpirer, and R. Thomas. 1967. The structural gene for A exonuclease. Proc. Nat. Acad.
Sci. U.S.A. 57:277.
10. Sakaki, Y., A. E. Karu, S. Linn, andH. Echols. 1973.
Purification andpropertiesof the-yprotein specifiedby bacteriophageA:aninhibitorofthe hostrecBC recom-binationenzyme.Proc. Nat. Acad.Sci.U.S.A. 70:2215.
11. Signer, E. R., and J. Weil. 1968. Recombination in
bacteriophageA.I.Mutants deficient ingeneral
recom-bination. J. Mol. Biol. 34:261.
12. Signer, E. R., H. Echols, J. Weil, C. Radding, M.
Shulman,L.Moore,and K.Manly.1968.Thegeneral
recombination system ofbacteriophageA.ColdSpring
HarborSymp.Quant.Biol. 33:711.
13. Sironi, G., H.Bialy,H.Lozeron,andR.Calendar.1971.
BacteriophageP2: interaction withbacteriophageand
withrecombination-deficientbacteria.Virology46:387.
14. Stahl, F. W., K. D. McMilin, M. M. Stahl, J. M.
Craseman,andS. Lam. Thedistribution ofcrossovers
along unreplicated lambda bacteriophage
chromo-somes.Genetics77:395.
15. Zissler, J.,E.Signer,and F. Schaefer.1971a.The roleof recombination ingrowthofbacteriophage lambda.I.
The gamma gene,p.455-469. In A. D. Hershey (ed.),
Thebacteriophagelambda. ColdSpring Harbor,New
York.
16. Zissler, J.,E.Signer,and F. Schaefer. 1971b. The roleof recombination ingrowthofbacteriophagelambda.II. Inhibition ofgrowthby prophage P2,p.469-477. In A. D. Hershey (ed.), The bacteriophage lambda. Cold
SpringHarbor,New York.
1450 J. VIROL.