JOURNAL OF VIROLOGY, Nov.1978,P.466-474 0022-538X/78/0028-0466$02.00/0
Copyright X)1978 AmericanSociety for Microbiology
Vol.28, No. 2 Printed in U.S.A.
Mutants of
Sindbis
Virus
III.
Host
Polypeptides
Present in
Purified
HR
and
tslO3
Virus
Particles
ELLEN G. STRAUSS
DivisionofBiology,CaliforniaInstitute of Technology, Pasadena, California 91125
Received for publication 5 June 1978
The amounts of host-encoded protein present in purified Sindbis virions of both the HR strain and of a mutant (ts103) whichmakes multicored particles wereexamined. Cells were labeled with [35S]methionine before infection and with [3H]methionine postinfection. Virions were purified by velocity sedimentation andisopycnic banding,and theirpolypeptideswere examinedbypolyacrylamide gels in a sodium dodecyl sulfate-containing discontinuous buffer system. Host prelabeled materialwasfoundprincipallyinasmallnumber of discrete polypep-tides in HRvirions,whichcontainedaslittleas0.2% host-encodedprotein. Virus-sized particles ofmutant tslO3 containedsignificantly more host material, and
multiploid particles from ts103 infection contained up to 12% host prelabeled
protein.
Sindbis virus and otheralphaviruses are the
simplest envelopedviruses(20).Theicosahedral nucleocapsid of the virus, which contains several hundred molecules of a single protein species surrounding the viralRNA,is assembled inthe cytoplasm of the infected cell. Upon diffusionto the cell surface, it interacts directly with the internalregions oftwovirus-encoded
glycopro-teins, El andPE2, present in theplasma mem-brane, leading to virus budding. During matu-rationaproteolytic cleavage of PE2to E2and E3occurs, and thematurevirionthus contains only three or four protein species, depending upon whetherE3 is retained in the virion. Other
envelopedviruses suchasrhabdoviruses,
ortho-myxoviruses, and paramyxoviruses mature by related mechanisms, but they contain a more complexnucleocapsidand alsocontain, in addi-tiontotheexternalglycoproteins,amatrix pro-tein. Thismatrixproteinisintimatelyassociated with the inner (cytoplasmic) surface of the
bi-layer and appears to interact with both the
capsid and the glycoproteins. In all cases the majorpolypeptide componentsof the virion are virusencoded (8, 14), whereas thelipid constit-uents arelargelyhostspecific, with little selec-tionby the virus (3, 9, 13, 15).
Early experiments on virus maturation sug-gested thathostcomponentswereexcludedfrom sitesof virusbudding (8, 14).Subsequent exper-imentsshowedthatthe virus-specific envelope polypeptidesarefree to diffuselaterally in the hostcell membranes and formpatches of modi-fied membranebyinteraction withintemal pro-teins ornucleocapsids (20). It wasthen tacitly assumed thatvirions contained only
virus-spe-cificpolypeptidesand that residual host material found in agiven virus preparation was the result ofinsufficientpurification(see review byLenard and Compans [11]). Recently, however, there hasbeen renewed interest in thepossibility that certaincellular polypeptides form integral mem-brane components of some enveloped viruses. Therefore, Sindbis virions were examined for the presence of host polypeptides which had been synthesizedbefore infection. The amount and type ofcellular material found in the HR strain andtslO3virionswerecompared. tslO3is amutantof Sindbis virus which forms aberrant
particles containing multiple nucleocapsids withinoneenvelope (19).
MATERIALS AND METHODS
Cell and virus. Preparation of monolayers of
chickenembryo fibroblasts has been described
previ-ously (16). Virus stocks usedwerethe HR strain of
Sindbis virus(2) and themutantts103 (19).
Labelingconditions.Rollerbottles (750 cm2) of
subconfluentchicken embryo fibroblasts were washed
for several hourswithEagle minimalessential medium
(6) containing Earlessalts, 2% dialyzed fetal calf
se-rum, and 1/3 to 1/5 the normal concentration of
essential amino acids. Then
[355]methionine
(460 to500 Ci/mmol; Amersham/Searle) wasadded to 4 to
4.2mCi/roller bottle (depending ontheexperiment)
in 150 ml ofEagle medium containing 1/3x of all
essential amino acids except methionine (which was
at0.017mM or1/6X)and 3%dialyzed fetal calfserum.
In thismedium, themonolayers reached confluency in
48h,asopposedto 24hin mediumcontaining normal
aminoacid concentrations.
At 3 hbeforeinfection,the35S-containingmedium
wasreplacedwithEagle medium containing unlabeled
methionineat 0.2mM (2x)and2%dialyzed fetal calf
466
on November 10, 2019 by guest
http://jvi.asm.org/
serum.Bottleswereinfected with10mlof
phosphate-buffered saline (5) containing 1% dialyzed fetal calf
serum, 1 ,igofactinomycin D per ml,and sufficient
HR or ts103 virus to give a multiplicity of5 to 10
PFU/cell. Mock-infected rollers received10ml of
dil-uentwithout virus.
At1hpostinfection the inoculumwasremoved and
replaced with 40 ml of low-salt medium (containing
NaCl at0.065M inplace of the normal concentration
of 0.115 M) per roller bottlewith 1/3X essential amino
acids exceptmethionine(whichwasat lxor2x)and
1 ,ug of actinomycin D per ml. After 1 h this was
replaced with mediumcontaining [3H]methionine (5
to10mCi/bottle),0.065MNaCl,1/3Xessential amino
acids exceptmethionine,3%dialyzedfetalcalf serum,
and atotalconcentrationof methionine of 1/2x to2x
depending upon theexperiment.
([Methyl-3H]methi-onine[5.5Ci/mmol]waspurchasedfromAmersham/
Searle, lyophilized to remove mercaptoethanol,
dis-solved in thelabeling medium, and sterilized by
filtra-tion.) Bottleswere incubated at30°C for 16 to 17 h
after HR virus infection and 20 to 21 h after tslO3
infection. The low-salt radioactive medium was
re-moved,and the virus was harvested by incubating the
monolayers with asmallvolume of medium containing
0.16M NaCl (16).
Purification of virus. The entire virus harvest
from oneroller bottle (-7ml)waslayered onto 32-ml
gradientsof15 to30%sucrosein 0.2MNaCl-0.05 M
Tris,pH 7.6-0.001 M EDTA-0.3% fetal calf serum and
sedimented inanSW27rotorfor90minat27,000 rpm
and4°C. Fractionswerecollectedby pumping,anda
smallportionof eachonewasassayed for 5Sand3H.
Fractions containing virion-sized tslO3, multiploid
tslO3 sedimenting 1.7x as rapidly, and 2.Ox
multi-ploidswerepooled separately.Comparable regions of thesucrosegradientscontaining HR and the harvest
fromamock-infection roller bottlewerealsopooled.
Pooledfractions were layered directly onto 11-ml
preparative isopycnic gradients containing 22 to44%
sucrose, 0.2M NaCl, 0.05 MTris (pH 7.6), 0.001 M
EDTA, and 0.3% fetal calfserumin 90%D20.
Centrif-ugation was for 12 h or more in an SW40 rotor at
32,000 rpm and4°C. Asmallamountof each of the
pools was also centrifuged in a parallel analytical isopycnic gradient with 32P-labeled HR virions as a
marker. In eithercasethegradientswerecollected by
pumping, and a small amount ofeach fraction was
counted inathree-channel Beckmanliquid
scintilla-tion spectrometer. Correcscintilla-tion of thecountsfor channel
overlapwasmadebycomputer.
Forquantitationof hostpolypeptides, pooled
frac-tions from the preparative isopycnic gradients were
diluted with5volumes of 0.2 M NaCl-0.05 MTris, pH
7.6-0.001M EDTA andfurtherpurified byasecond
cycle ofsucrose gradient velocity sedimentation
fol-lowedbyisopycnic separation.
Polyacrylamide gelelectrophoresis. Gels
con-taining 10%acrylamide, 0.26% bisacrylamide in 0.19M
Tris-chloride buffer(pH 8.6), and 0.1% sodium dodecyl
sulfate(SDS)werepreparedasdescribed byLaemmli
(10). Stacking gels contained 5%acrylamide, 0.067%
bisacrylamide,0.063MTris(pH6.8), and 0.1% SDS.
Sampleswerefirstdialyzed against buffer containing
0.05M Tris-chloride, pH 6.8, 1%SDS, 10% glycerol,
and 1to2%,8-mercaptoethanolandsubjectedto
elec-trophoresisuntil the bromophenol blue front was
ap-proximately0.5 cm from the bottom of the gel. Gels
werefrozen and sliced onaMickle gelslicer,and the
fractionswerecounted inascintillation cocktail
con-taining toluene,Liquifluor,andNCS.
Gelswerecalibrated for molecular weight by using
a series of standard proteins. Electrophoresis
condi-tionswerethesameasdescribed above, except that,
in the case of the unlabeledmarkers, the gels were
stained overnight with 0.05% Coomassie brilliant
blue-25% isopropyl alcohol-10% glacial acetic acid
and destained with several changes of
water-methanol-glacial acetic acid at 66:33:10. Gels were
scanned for absorbanceat580nm withagelscanning
attachment for an ISCO model UA-5 Absorbance
Monitor(Instrumentation SpecialtiesCo.).Standards
used in addition to the three virion proteins from
purified HR virus werephosphorylase A (molecular
weight, 94,000 [94K]), bovine serum albumin (66K),
pyruvatekinase(57K), heavy chain of gammaglobulin
(50K), actin (43K), D-amino acid oxidase (37K), and
soybeantrypsininhibitor(21K).
We have found that small changes inpH
signifi-cantly influence the rate ofmigration of the virion
polypeptidesrelativetothebromophenolblue marker
but havelittle effectonthemigrationof theSindbis
polypeptidesrelativeto oneanother.For thisreason allmeasurementsofmigrationrates weremade
rela-tivetothemigrationrateof Sindbiscapsidprotein.
Laemmli slab gels containing anexponential
gra-dient ofacrylamidefrom8 to20%wereusedtobetter
displaythelower-molecular-weightcomponents. Stan-dard proteinsused to calibrate this gelsystemwere
insulin (two chains of molecular weights 2.3K and
3.4K), a-bungarotoxin (8K), cytochromec(11.7K), ,8-lactoglobin (18.4K), and soybean trypsin inhibitor
(21K).
Immuneprecipitation.Asafinal method of
pu-rification of certain virusfractions, the materialwas
precipitated by atwo-stageimmuneprecipitation, in
which rabbit anti-Sindbis immunoglobulin G (IgG)
and goat anti-rabbitIgGwereused. The anti-Sindbis
IgGfractionwas agiftofC.Birdwellandwasabsorbed
three timesonmonolayersof uninfected chickencells
before use. Goat anti-rabbit IgG was obtained as a
lyophilized powderfrom Calbiochem. Fractions from
theisopycnicgradientswereprecipitatedat4°Cwith
anti-Sindbis serum for 4 h, followed by incubation
overnightat4°Cwith goatanti-rabbitserum.
Precip-itateswere collectedby centrifugation at27,000x g
for20minand dissolved in thesamebufferasused for
dialysisofgel samples.Bromophenolbluewasadded,
and thesampleswereboiled for 2 min.Electrophoresis,
slicing,andcountingwere asdescribed above.
RESULTS
Amountofprelabeled proteininHR and ts103. Several prelabeling experiments were
performed,withsomechangesinprocedural de-tail,
especially
inthespecificactivities of methi-onine used; with each experiment an attempt wasmade toimprove
theefficiency ofthe chase. Several preliminary experiments were done inon November 10, 2019 by guest
http://jvi.asm.org/
468 STRAUSS
which thepurification consisted of onlyasingle
cycle of velocityandisopycnic centrifugation.In theseexperiments,theamountof hostmaterial in lx tslO3 virions varied between 0.5 and 2.8%, dependingonthepreparation. Afinal
ex-perimentwitha two-cycle purificationreduced
the amount of host material to an apparent
minimum and yielded the quantitative data shown in Table 1.
Chicken embryo fibroblasts were labeled for
2dayswith [35S]methionineataspecific activity
of 1,700mCi/mmol,infectedwith eithertsl03or
HRormockinfected,andlabeledpostinfection
in low-salt medium containing [methyl-3H]me-thionine at 1,150 mCi/mmol; virus was
har-vested in normal salt. The entire virus harvest from each infection wassedimented in sucrose
velocitygradients,andmaterialssedimentingat
lx(the majorviruspeak-),1.7x(containing mul-tiploidparticles sedinenting1.7xasfastasHR
virions), and 2,OX werepooledfor further anal-ysis.
Eachof thepooledfractionswaslayeredonto both analytical and preparative isopycnic
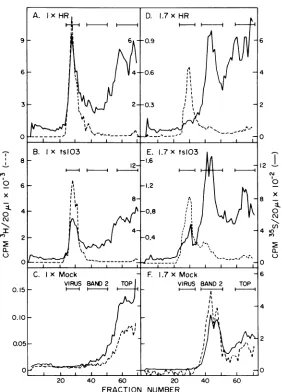
gra-dients. Figure 1 shows profiles obtained with preparative isopycnic gradients of lx (virion-sized) and1.7x material forHR,tslO3,andmock
infections. The gradients from virus-infected material contained well-definedpeaksof3H la-bel, which was coincident with the32p marker HR virus on the parallel analytical gradients.
Themock-infected monolayer showed no such
peak and illustrated thatmost of the
35S
label associatedwith the virus from the infectedcellswas specific to virus infection. The location of the virus region shown on this gradient was
determined byreference tothe32p markerHR
on theanalytical gradient. In all three samples
J. VIROL.
from the 1.7x region there was a secondpeak
(band 2; usually split into twopoorly resolved peaks) near the density of 1.19 g/ml (at 5°C),
midway between the sample overlay and the Sindbispeakat1.21g/ml. From the densitywe
postulate that this fraction contained membrane fragments which sedimented at greater than 400S.
The viruspeaks from such isopycnic gradients
were diluted and resedimented in velocity
su-crose gradients, and the appropriate fractions wereagain selected and rebanded on isopycnic
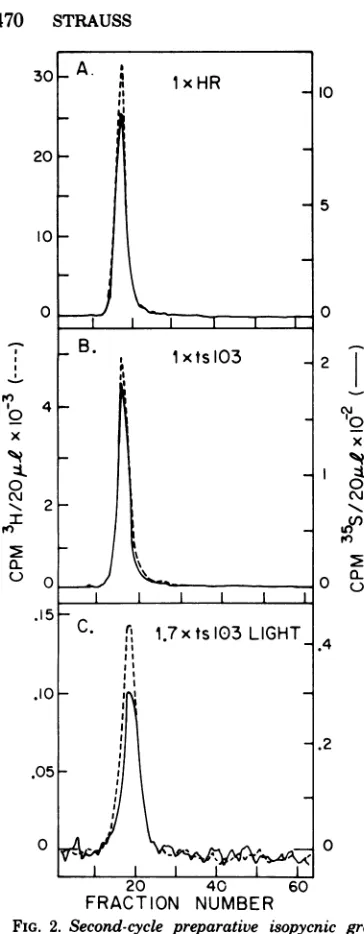
gradients. Selected isopycnic gradients from suchatwo-cycle purification areshownin Fig.
2. On these gradients, little or noradioactivity,
either 35S prelabel or 3H postlabel, was found
outside the virus peak. In the case ofthe 1x
ts103the twohalves of thefinal isopycnic
gra-dientwerepooled separately (lxtslO3"dense"
andlxtslO3 "light").In thecaseof 1.7x tslO3,
the virus band was split according to density
after thefirstisopycnic gradient (see below). Figure 3showstheresult ofthesecond-cycle velocitygradientofthe 1.7x HR. Themajority of the 3H label (>60%) sedimented at lx, with only25% ofthe materialresedimentingat1.7x. This is characteristic ofHRpreparations; mul-tiploid particles are produced in very small
amounts and are often obscured by reversible
aggregates of lx particles sedimenting in the
sameregion. (The1.7xparticles from the second
cycle all sedimentat1.7x,however.) It is readily apparent from thisfigure thattherewas much more35Sassociatedwith themultiploid particles
than with the lx virions (also seebelow). The
1.7x particles and the lx particles from this velocity gradient were pooled separately and
sedimented to equilibrium on isopycnic
gra-TABLE 1. Quantitation of host polypeptides in Sindbis virus
%Host Total[3H]methi-
%s
El+ Total[3S]methi- %asEl+Sample polypep- onine radioactiv- E2+C onineradioactiv-
Ey
+Ctidesb ity (cpm) o ity (cpm)o E2 + C
lxHR 0.24 385,600 94.3 9,690 68.0
lxHR from 1.7x HR pool 0.56 28,700 89.4 1,010 49.7
1.7xHR 4.01 5,300 77.0 770 24.1
lxts103dense 0.58 110,100 88.8 3,950 49.1
lxtslO3light 1.14 30,170 84.6 1,560 33.5
1.7x tslO3 intermediate den- 3.80 9,050 81.5 1,320 24.5
sity
1.7xts103light 12.31 2,650 75.5 1,070 18.2
aPreparationandpurification of these samplesisdescribed in thetext.
bHostpolypeptidesarecomputed as
follows:
% host polypeptides=(counts of 35S not inEl
+ E2 +C/countsof3HinEl+E2+C)x(specific activityof3Hin counts per minute per mole/specificactivity of 3S in counts
per minute permole). This formulaexpresses the 35S present corrected for the amountreutilizedin virion
proteins but uncorrected foranyother turnover andnormalizedtothe total amount of virion polypeptide on
thegel. Itgivesamassratioifone assumesthatthe average methionine content of the host polypeptides is
similartothatof the virion polypeptides.
c35Scountshave been corrected fordecay.
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.499.61.456.481.588.2]I0
x 0
N
I V
re
12 1
N
0 x 8
-0 N N CI) 4 )
CL u
20 40 60 20 40 60
FRACTION NUMBER
FIG. 1. Preparative isopycnic gradients of Ix and 1.7x regions from HR, ts103, and mock infections.
Samples (20 ,u) ofeach fraction were counted in Aquasol to determine the distribution of prelabel and
postlabel. (A)Ix HR; (B) Ix tsl03; (C) Ix mock;(D) 1.7xHR; (E) 1.7x tsl03; (F) 1.7xmock. ( )
[35S]-methionineprelabel; (---)[3H]methioninepostlabel. Sedimentationwasfromrighttoleft.
dients. They are referred to below as lx HR from 1.7x pool and1.7x HR,respectively.
The 1.7x tslO3 particles show considerable density heterogeneity on isopycnic gradients
(19).Therefore, after the firstisopycnic
centrif-ugation, three regions of the virus band were pooled:the denseside, the central(intermediate) region, and the light side. Each of these was diluted and resedimented separately on 15 to
30% sucrose gradients, where virtually all the
radioactivity resedimented at 1.7x. The 1.7x material from the three second-cycle velocity gradientswere pooledseparatelyandrebanded onthreeisopycnicgradients,where each formed asingleband. Thesepreparations are referred to
as 1.7x tslO3 dense, 1.7x ts103 intermediate,
and1.7x ts103light,respectively.
Selected fractions from these gradients were analyzed by acrylamide gel electrophoresis for radiolabelinvirion andhostpolypeptides. Rep-resentative gels are shown in Fig. 4, and the resultsof one suchexperiment are summarized in Table 1. Several conclusions can be drawn from these data. The first is that the amount of hostpolypeptide foundin HR virions was strik-ingly small, as little as 0.24%. The amount of prelabelin lx ts103 was severalfoldhigher, on theotherhand,anddependedonthedensityof the particle. Multiploid particles, whether HR or
tslO3,
contained more host material; foron November 10, 2019 by guest
http://jvi.asm.org/
[image:4.499.112.394.68.460.2]A.
density.
These resultssuggest
thatabnormalities30-xHR in
budding
lead to the inclusion of more hostmaterial in the virionsproduced.
Host polypeptides in purified virus par-ticles and in other fractions. Figure 4shows
20_ three of the profiles from which the data in
Table 1 were obtained andillustratesthat
puri-__5 fiedvirions from HR andts103 differ in
compo-sition of prelabeled material. In these
prepara-0_ tions, a
small
number of discrete species ofpre-labeled host polypeptides was identified. The same host polypeptides appeared reproducibly
O
0 / in Oallprelabeled preparations
examined and0 l have been numbered in order of decreasing
mo-B. lecular
weight.
lx HRvirions(Fig.
4A) containi-
*t
x11s
103 _2polpeptides
called 1 (molecular weight, 68K), 3 (41K), 7 (22K), 8 (21K), and F (polypeptides of4 - <20K which
migrated
with the solventfront).
o
'O
(Forreference,
theapparent
molecularweights
l x of El, E2, and C in this system are 54K, 48K,
IN _ I _ and
32K,
respectively.)
Prelabel35S
was alsoo 0 found in E1, E2, and C, presumably due to
(M_2 inefficiencies in the chase or from breakdown
roI andreutilization of the label afterinfection, but
E'
IK there was verylittle 35S
in otherregions
of theL\Egel. lx tslO3 (Fig. 4B) contained the host
poly-0 o peptides found in lx HR as well as peakscalled
C.L
1.7
xts
103
LIGHTj
4 1~~~~.7xHRl
12
.10~~~~~~~~~~~~~~~~~~~2
20-~~~~~~
021
,05
A: 15 8 Xx
-0
[image:5.499.64.246.49.516.2]FRACTION NUMBER5
FIG. 2. Second-cycle preparative isopycnic gra-L X /
dients of(A) Ix HR, (B) Ix ts103, and (C)1.7x ts103_
light. The origin of the samples isdescribed in the 20 40
text. ( ) [35S]methionineprelabel; (- --)[3H]me- FAT NNME
thionineFpostlabel.
NUFIG. 3. Second cycle ofpreparative velocity
sedi-ample,thelight 1.7x tslO3 containedup to 12% mentationof 1. 7x HR. 1. 7x HR material was purified host protein, 10 to 20 times as much as was byvelocity sedimentation followed by isopycnic
cen-found in lx tslO3. Despite these differences trifugation,and thevirusbandwasdiluted and
cen-earlierexperimentshave shown that thesemul- trifuged on a15 to30%sucrosegradient. Portions of
tiploids are fully infectious. The 1.7x HR are eachsamplewerecountedfor
'S
and3H. Samples14also
nlot beenasr weyocharacterized
naturaly
occurring multiploids,
but have through 23 werepooled,
banded on anisopycnic
mu 9 Ne
thnave
gradient, and designated 1.7x HR. Samples 30not been as well characterized (19). Note that through 36 were treated similarly and designatedlx
1.7x HRweremade inverysmallamounts and HRfrom 1.7x HR pool. (-)
[3SJmethionine
pre-contained levels ofprelabeled material compa- label; (---)[3H]methionine
postlabel. Sedimenta-rabletothatfound in1.7xts103 ofintermediate tion wasfrom right to left.470 STRAUSS J. VIROL.
--- ---I - - ---1
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.499.249.455.84.549.2] [image:5.499.264.455.355.549.2]polypeptides and in thesmall material migrating atthe front.
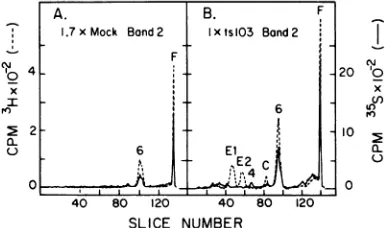
Selected fractions from first-cycle isopycnic gradients (such asthose shown in Fig. 1) were
also examined as acheck onvirus purification
and to determine the distribution of polypep-tidesthroughout thegradients. Figure 5shows gel profiles ofthe band 2 material from mock-infected and tslO3-mock-infected preparations. One predominant peak (6, 25Kto26K)wasfound in
addition toF. Polypeptide 6 (aswell asF) was
labeled with both 35S and 3H in both cases,
showing that it is a species which turns over
rapidly, is synthesized after actinomycin treat-ment of the cells, and is independent of virus infection. Band 2 fromtslO3 infection contained the Sindbispolypeptidesinadditionto6and F. The absence ofpolypeptide 6from the various purified virion preparations in Fig. 4
demon-To
stratesthatthe viruspreparationswerenotde-x tectably contaminated with band 2 material.
The
origin
of band 2 isunclear,
although
its isopycnicdensity of1.18to1.19indicates that ita.
o contains membrane
fragments.
Whatever itsor-igin, it is curious that it contains suchalimited
distributionofpolypeptides.
It shouldalso be pointed outthat host poly-peptides 1,5,and 7 couldnotbedetected inany
fraction examined from uninfected cells (whether the virus region, band 2,orthe soluble
material at the top of the isopycnic gradient). Thesepolypeptidesappeartobespecifically
as-sociated with virusparticlesinsomeway.
Thelow-molecular-weight material migrating with thesolventfrontonthesegels (F) has been
showntobecomposedofpolypeptides. Itis
non-dialyzable, precipitable in hot trichloroacetic acid, and insoluble in chloroforn-methanol. When Fwasdisplayedon8to20% Laemmli slab
SLICE NUMBER
FIG. 4. Acrylamide gel electrophoresis of virus
particles purified throughtwocycles ofsedimentation
followed by isopycnic density banding.
Electropho-resis wasfrom left toright; Fcomigrated with the
bromophenol blue marker. (A) Ix HR.Same
prepa-rationasinTable1,line 1.(B)Ix tsl03, dense side
ofviruspeak.SamepreparationasinTable1,line 4. (C) 1.7x tsl03, lightsideofviruspeak.Same
prepa-ration as in Table1, line 7. (-) [3S]methionine
prelabel;(---)[3H]methioninepostlabeL
4 (33K) and5 (29K).Allof the host peakswere more prominent in the tslO3 material, and in
additiontherewasmoreofabackgroundsmear
of35Sthroughout the gel. Thelight 1.7x ts103 particles (Fig. 4C) contained 35S primarily in peptides1andF, plusageneralsmear.InFig.4,
the 3Hpostlabel wasfoundonlyin theSindbis
C~4
I
24
10
0
20 '0
x
U)
10E
0
40 80 120 40 80 120
[image:6.499.58.225.51.490.2]SLICE NUMBER
FIG. 5. Polyacrylamidegel electrophoresis ofband
2fractions from isopycnic gradients. Fcomigrated
with the bromophenol blue marker. (A) 1.7x mock, band 2(pool of fractions41,42, 43fromgradientin
Fig. IF). (B)1.7x tsW03,band 2(pool offractions 37,
38, 39fromgradientinFig.IE). (-)
[3S]methio-nineprelabel; (---)[3H]methionine postlabel.
I0
x
x
I
A. B. F
1.7xMock Bond2 IxtsIO3 Bond2 F
4 ~~~6
6 E
'E2
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.499.254.448.482.596.2]472 STRAUSS
gels with appropriate low-molecular-weight markers, both theprelabelandpostlabelshowed a small number of discretebands, betweentwo andfourdependingonthesample, with molec-ular weights between10Kand 20K.F was pres-ent in the most highly purified virus prepara-tions examined (Fig. 4A and B) and was not removed by immunoprecipitation with anti-Sindbis antibody, implying that some orall of these smallpolypeptides may be integral
com-ponents of the virion. On the other hand, it is possible that these low-molecular-weight
com-ponents are in part generated by proteolysis
occurringduring purification. DISCUSSION
Hostpolypeptides in purified Sindbis virions have been examinedtodetermine the absolute level of host proteinpresent invirusparticles,to ascertain thenatureanddegree of heterogeneity of hostproteinfound invirions,andtodetermine whether the amount or type of host material dependsupontheefficiencyofbudding.First of all, the host materialfound in HR virionswas a verysmall percentage of theprotein of the Sind-bisvirion,althoughit isalwaysdifficulttoknow whether residual host label invirions is dueto truevirioncomponentsor toresidual nonvirion contaminants. Pfefferkorn and Clifford (14) placed an upper limit of 5% host material in Sindbisvirus,butimproved techniquesandthe
availabilityofhigherspecificactivitylabels have reduced this limitconsiderably. Thepresent re-sults indicate that during a standard infection withthevirus, theamountof host protein found inpurified virionswas aslow as0.2 to 0.5% of the totalprotein,whichtranslatestoonlytwo to four host polypeptides per particle. What pro-portion of theprotein ofthe infected cell
plas-malemma isrepresented byEl and PE2 is un-known, but it isprobablyless than50%,
consid-eringthelengthof theinfection and the relative ratesofproteinsynthesis in infected and unin-fectedcells. Thus, anarea of membrane which contains morethan 400 host proteins and less than 400virus-specificmoleculesatthe onset of budding produces a mature virion containing 800viruspolypeptidesand not more than2to 4 host polypeptides in its envelope. This rather remarkableselectivityin anessentiallyfluid sys-temisprobably provided bytheinteraction be-tweenthenucleocapsidand the transmembran-ousportionsoftheglycoproteins (20).
In comparing HR virus with tslO3, it was found that lx ts103 contained more prelabel materialthan didthecomparable HR fraction. In addition to the discrete species of polypep-tides seen in HR particles, tslO3 particles also
containedmore of a smearofmanypolypeptides, suggestinganonspecific inclusionofhost mate-rial.Larger tslO3 virionparticles, the multiploids which contain four to eightnucleocapsids in a single envelope, contained significantly more host polypeptides than did the lx virionswith asinglecore,and thesmearofbackground poly-peptides was quite pronounced. Furthermore, thetslO3multiploids of lowestdensity contained up to 12% 35S prelabel; these particles have a higherproportion ofenvelope tocore, in other words, loosely fitting membranes. It is also of notethatmultiploids of HR containmore host protein than does the standard HRvirion; this isreadilyapparentinFig. 3and Table1.
There are three possible origins for the host material whichwasfound associated with virus. The first is that the 35S material consists of residualcontaminants whichareadsorbedonto the surface of the virions. Such a mechanism wouldnotexplain the differencesseenbetween lxHR and1.7xHRorbetween lxHR andlx tslO3, which should have very similar surface properties foradsorption. The second possibility is that the35S-labeledprotein is duetointrinsic contamination, i.e.tomaterial whichcopurifies withparticular virus classes. Thisseemshighly unlikely because of the absence ofcomparable prelabeled material in mock-infected prepara-tions and because of thedisparate levels of host polypeptides present in the various virus sub-classes. Ipersonallyfavor thethirdalternative, i.e., thatsomeof theprelabeled material (above the0.2 to0.5%level whichmaybeameasureof thepurity of the preparations) represents inte-gral host membrane componentswhich are in-serted intheenvelope of the virion.
All of the results support the general model forSindbis maturationpreviouslypresented (20) as well as our explanation of the defect inthe mutanttslO3 (19). The tslO3 defect isprobably inthenucleocapsidprotein;wehave shown that themajorityofnucleocapsidssynthesized early ininfectionare aberrant in their sedimentation behavior, but thatmostofthese defectivecores do not bud and are not recovered in released virions.Wehave postulatedthatallof the man-ifestations of the tslO3 mutation can be ex-plained if the ts103 nucleocapsidcanonly inter-act weakly with the glycoproteins in the cell surface. Whereas tslO3 veryseldombudssingly and usually buds as multiploid particles, it is easytoseehowhost materialcouldbeincluded adventitiously.
Themosthighly purifiedpreparationsofHR virionscontain theequivalentofonly two to five molecules of hostpolypeptides pervirion, and thesearepresentasseveralpolypeptide species.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
Incontrast tothis, actin has been foundtobea
regular component of both paramyxoviruses, suchasSendaivirus(17),andtheavian, monkey,
and murine RNAtumorviruses(21).Comprising
upto8% of the proteinmass ofthe
Rous-asso-ciated virus-2 virion, this large amount of a
cellularprotein incorporatedisthoughttoresult fromabudding mechanism which requires the
juxtapositionofmicrofilamentstotheemerging virion. Actin,whichshould migrateatthe posi-tionof hostpolypeptide 3, could be present in only insignificant quantities in Sindbis virions. Inaddition, there have been numerousreports whichsuggestan associationof polypeptides of
the major histocompatibility complexwith
en-veloped virus particles. On the one hand, H-2
antigenshavebeenreportedtobeintegral
com-ponentsofvesicular stomatitis virionsgrownin
mouse L-cells (7). On the other hand, studies
with several viruses, including lymphocytic cho-riomeningitis virus and Rauscherleukemia
vi-rus,haveshown thatsensitizedcytotoxic
T-lym-phocytes are most efficient in killing
virus-in-fectedcellswhen the target and thekillercells
sharethe samehaplotypeofthe H-2
histocom-patibility locus, suggesting that the cell surface antigen recognized might be a hybrid of viral
andcellularcomponents (4, 18). Sindbis virions havenotbeen testeddirectly for suchantigens, but they could be present only in very small
quantitiesinthesepreparations.
There have also been numerous reports of
host cell enzymatic activities associated with purified viruses, but whether these represent polypeptides necessaryfor production of
infec-tiousvirions, adventitiouscomponentswhichare
engulfedorincluded in theenvelopeatthe time of budding, or stubborn contaminants which
copurifywith virions is unclear(1). It has been suggested that the host-encoded ribosomes which are found in the arenaviruses belong to
the second category and are not required for virusreplication (12).
Finally, there is supporting qualitative evi-dence indicating that alphaviruses are much more selective in their envelope components
thanother virusgroups.Arecentseries of phe-notypic mixingexperimentshas shown that it is possible to produce pseudotypes containing either vesicular stomatitis virusoraviantumor
virusgenomeswithSindbis envelope
glycopro-teins. However, noparticles could be detected
containing Sindbisnucleocapsids and
heterolo-gous viral antigens (22). This implies that the interactionof thealphavirus nucleocapsidwith its envelope is highly specific, much more so
thanthe interaction between glycoproteinsand the matrixproteins ofrhabdoviruses or
oncor-naviruses. Thismay
explain
the almostcomplete
exclusion ofhost polypeptides fromalphavirus
virions which has beenobserved, in contrastto the situationwith other enveloped viruses dis-cussed above.ACKNOWLEDGMENTS
IthankEdithLenchesandMaryStammreich-Martinfor excellenttechnical assistance, Charles Rice for his participa-tion inthepreliminaryexperiments, and James H.Straussfor stimulatingdiscussions and invaluable assistance in the prep-aration of thismanuscript.
This research was supported by Public Health Service grant AI-10793 from the National Institute of Allergy and Infectious Diseases.
LITERATURE CITED
1. Bishop, D.H. L., and M. S. Smith. 1977. Rhabdoviruses, p.167-280. In D. B. Nayak (ed.), The molecular biology ofanimal viruses. Marcel Dekker, Inc., New York. 2. Burge, B.W.,andE.R.Pfefferkorn. 1966. Isolation
and characterization of conditional lethal mutants of Sindbis virus. Virology 30:204-213.
3. David, A. E. 1971. The lipid composition of Sindbis virus. Virology 46:711-720.
4. Doherty, P. C., and R. M. Zinkernagel. 1975. H-2
compatibility isrequired for T-cell-mediated lysis of
targetcellsinfected withlymphocytic choriomeningitis virus. J.Exp. Med. 141:502-507.
5. Dulbecco, R., andM.Vogt.1954.One-step growth curve ofwesternencephalomyelitisvirusgrown in vitro and analysisof the virus yieldsfromsingle cells. J. Exp. Med.99:183-199.
6. Eagle, H. 1959. Amino acidmetabolism inmammalian cellcultures. Science 130:432-437.
7. Hecht,T.T.,and D. F. Summers. 1976. Interactions of vesicular stomatitis virus with murinecellsurface an-tigens. J. Virol. 19:833-845.
8. Holland,J.J., and E. D.Kiehn.1970.Influenza virus effects on cell membraneproteins. Science 167:202-205. 9. Klenk, H.-D.,and P. W.Choppin.1969.Lipids of plasma membranes of monkey and hamster kidneycellsand of parainfluenza virions grown in these cells. Virology 38:255-268.
10. Laemmli, U. K. 1970. Cleavage of structural proteins during theassemblyof thehead ofbacteriophage T4. Nature(London)227:680-685.
11. Lenard, J.,and R. W.Compans.1974.The membrane structureoflipid-containingviruses. Biochim.Biophys. Acta 344:51-94.
12. Leung,W.-C.,and W. E. Rawls. 1977. Virion-associated ribosomes are notrequired for thereplication of Pi-chindevirus.Virology81:174-176.
13. McSharry,J.J.,and R. R.Wagner. 1971.Lipid com-positionofpurifiedvesicular stomatitis virus. J. Virol. 7:59-70.
14. Pfefferkorn,E.R., and R. L.Clifford.1964.The origin oftheproteinofSindbis virus.Virology23:217-223. 15.Pfefferkorn,E.R.,and R.L. Clifford.1964.The origin
oftheproteinof Sindbis virus.Virology 20:446-456. 16.Pierce, J.S.,E.G.Strauss, and J.H.Strauss. 1974.
Effectof ionicstrengthonthebinding of Sindbis virus tochickcells.J. Virol. 13:1030-1036.
17.Scheid,A., and P. W.Choppin.1974.Identification of biologicalactitities ofparamyxovirus glycoproteins. Ac-tivationofcellfusion,hemolysisandinfectivity of pro-teolytic cleavage of an inactive precursor protein of Sendai virus.Virology 57:475-490.
18. Schrader, J.C.,B.A.Cunningham,and G. M. Edel-man. 1975.Functional interactions of viral and
on November 10, 2019 by guest
http://jvi.asm.org/
474 STRAUSS
compatibility antigens at tumor cell surfaces. Proc. Natl. Acad. Sci. U.S.A. 72:5066-5070.
19. Strauss, E. G., C. R. Birdwell, E. M.Lenches, S. E. Staples, and J. H.Strauss.1977. Mutantsof Sindbis virus. II. Characterization ofa maturation defective
mutant,tslO3.Virology 82:122-149.
20. Strauss,J. H., and E. G. Strauss.1977.Togaviruses,p.
111-166. InD.Nayak (ed.),The molecularbiologyof animalviruses, vol,1.MarcelDekker,Inc., New York. 21. Wang, E., B. A. Wolf, R. A. Lamb, P. W. Choppin,
and A. R. Goldberg. 1976.Thepresenceof actinin enveloped viruses,p.589-600.InR. Goldman, T.
Pol-lard, and J. Rosenbaum (ed.), Cold Spring Harbor Con-ferencesonCell Proliferation, vol.3.Cold Spring Har-bor, N.Y.
22. Zavadova, Z., J. Zavada, and R.Weiss. 1977. Unilat-eral phenotypic mixing of envelope antigens between togaviruses and vesicular stomatitis virusoravian RNA
tumorviruses. J. Gen. Virol. 37:557-567.
J. VIROL.
on November 10, 2019 by guest
http://jvi.asm.org/
![TABLE 1. Quantitation of host polypeptides in Sindbis virus [3H]methi-%s El](https://thumb-us.123doks.com/thumbv2/123dok_us/1523069.104893/3.499.61.456.481.588/table-quantitation-host-polypeptides-sindbis-virus-methi-el.webp)