Copyright01975 AmericanSocietyforMicrobiology Printed in U.S.A.
Nucleotide
Sequence Complexities,
Molecular
Weights, and
Poly(A) Content of the Vesicular Stomatitis Virus
mRNA Species
JOHN K. ROSE* AND DAVID KNIPE
Department ofBiology,Massachusetts Institute ofTechnology, Cambridge,Massachusetts02139
Received forpublication23October 1974
Poly(A)-containing vesicular stomatitis virus mRNA species synthesized in vesicular stomatitis virus-infected cells have been separated into four bandsby electrophoresisonformamide-polyacrylamide gels.Two-dimensionalfingerprints
of ribonucleaseTi and ribonuclease Adigests of the RNA from each bandshow
that they contain unique oligonucleotide sequences as well as 60 to 125 nucleotides of poly(A). The fingerprints were usedto determine thenucleotide
sequencecomplexitiesof RNA from threeof the bands.Two containnucleotide
sequences whichaccount completelyfor their molecularweights (0.70 x 106 and
0.55 x 106) determined by gel electrophoresis and sedimentation rate, and,
therefore, these are radiochemically pure RNA species. The most rapidly migrating band mustcontain two orthree different RNA species since it hasa
molecular weight of 0.28 x 106, determined by physical methods, and a
nucleotide sequencecomplexitytwotothree times thatexpected forapureRNA
species of thissize. These dataareincompleteaccord with translationalstudies
(accompanying paper) which show that each of thetwo pureRNAspecies codes fora distinct viral protein, whereas the third codes fortwoviralproteins. From the molecularweight andsequence complexity determinations on mRNA from
the bands, we conclude that most of the vesicular stomatitis virus genome is transcribed into discrete mRNAspecies.
Two vesicularstomatitis virus (VSV) mRNA size classes (28S and 13 to
15S)
have been resolved previously by centrifugation on sucrosegradients (5). The 13 to 15S species have been resolved further into three species by
electro-phoresis on polyacrylamide gels, whereas the
28S RNA is notresolvedfurther(M. Stampfer, Ph.D. Thesis, Massachusetts Instituteof Tech-nology, Cambridge, 1972; D. Baltimore, T.
Morrison,
M.Stampfer,
and H.Lodish,
Nega-tive Strand Virus Meet.
Abstr.,
1973, inpress). These mRNA speciesarecomplementarytothe single strand of RNA contained within thevirion (5) and contain poly(A) (4, 13).
We describe here a chemical analysis ofthe
number of VSV mRNA species contained within three of thebandsseparated by Forma-mide-polyacrylamide gel electrophoresis. The determination is based on analysis of several large, unique oligonucleotides obtained from fingerprints of the RNA from each band. The radioactivity per nucleotide length is deter-mined for each oligonucleotide, and this
num-berisdivided into the total radioactivity in the fingerprint to obtain an apparent chain length
innucleotides(nucleotidesequencecomplexity)
for the mRNA species in each band. The average chain length of the poly(A) in each mRNA species is calculated in a similar man-ner.Thereliabilityofthis technique depends on the relative abundance of mRNAs within a
band, since oligonucleotides from a minor spe-ciesmightnotbe detected andwouldcontribute
only in proportion to their abundance to the total sequence complexity. Arelated approach
has been used previously to determine the
molecular weightofthe VSVvirion RNA (11).
MATERIALS AND METHODS
Virus and cells. Standard B particles of VSV
(Indiana serotype) were grown in Chinese hamster
ovary (CHO) cells and purified as described
previ-ously(6, 14, 15, 17).
32P labeling and purification of VSV mRNA.
CHO cells(2 x 108to 4 x 108)growing at 37 C were
infected at a multiplicity of 3 with VSV as described
by Huangetal. (5), except that the medium lacked
phosphate and contained fetal calf serum which had
been dialyzed for 6 h against three changes (50
volumes)of0.9% saline. The actinomycin D
concen-tration was 5 usg/ml, and 20 mM N-2-hydroxyethyl-994
on November 10, 2019 by guest
http://jvi.asm.org/
piperazine-N'-2-ethanesulfonic acid (pH 7.2) was
added. Thirty minutes after infection, the cells were
centrifuged and resuspended in phosphate-free
me-dium. At 1 h following infection, 10 to 20 mCi of
carrier-free 32P (New England Nuclear Corp.) was
added. Cells were harvested by centrifugation at 4 h postinfection, resuspended in 6 ml of RSB (0.01 M
Tris [pH7.3], 0.01 M NaCl, and 1.5 mM MgCl2), and
allowed to swell for 10 min at 0 C. Cells were
disrupted with a Dounce homogenizer, and the nuclei
were removed by centrifugation (5,000 x g, 5 min).
Nuclei were washed with 1 ml of RSB containing 1% Nonidet P-40 (Shell Oil Co.) and 0.5% desoxycholate
and recentrifuged (9). The combined supernatants
were adjusted to final concentrations of 0.01 M
EDTA, 0.4 M sodium acetate (pH 5.2), and 1%
sodium dodecyl sulfate (SDS). Five milliliters of a
solution containing 50% redistilled phenol, 49%
chlo-roform, and 1% isoamyl alcohol was added, followed
by brief mixing. The mixture was centrifuged for 10
min at 12,000 x g (4 C), and the aqueous layer was
precipitated with2volumes of ethanol. After
centrifu-gation, the RNA was dissolved in 0.4 M sodium
acetate (pH 5.2) andreprecipitated with two volumes
ofethanol. Usually 3 x 107 to 108 acid-precipitable
counts per minute were recovered in 2 mg of RNA
when2 x 108 cells were labeled with 20 mCi of 32P.
To obtain a quantity of VSV mRNA suitable for
preparative gel electrophoresis, the RNA was
dis-solvedin0.5 ml ofbindingbuffer (0.4 M NaCl, 0.01 M
Tris, pH 7.4, and 0.02% SDS), applied to 0.3 g of oligo(dT)-cellulose in a disposable pipette column,
and washed with 5 ml of binding buffer. The bound
RNA was eluted with 1 to 2 mlof0.1 mMEDTA(pH
7.4), precipitated with 2 volumes of ethanol, and
recovered bycentrifugation at 100,000 x g for1h. For
preparative formamide-polyacrylamide gel
electro-phoresis, the pellet was dried, resuspended in 20 to 50
glofsterile water, and lyophilized. The RNA was then
dissolvedinsample bufferforgelelectrophoresis.
Electrophoresis in formamide-polyacrylamide
gels. The method described by Duesberg and Vogt (3)
was used with the modification that an E.C. vertical
slab gel apparatus (E. C. Apparatus Co.) was used
insteadofcylindricalgels. Eitherthe four oreightslot
well former was used, and wells were filled with
formamide followed by underlayering of the RNA
sample dissolved in formamide-glycerol as described
(3).Electrophoresisat roomtemperaturewasfor 16to
20h at 100 V in 3.75% (wt/vol)acrylamide gels. Wet
gels were covered with Saran Wrap and
autoradio-graphedfor 20 s to 1h,depending upon theamountof
radioactive RNA used.
Purification of RNA from
formamide-polyacryl-amidegels. Using the autoradiogram as atemplate,
the regions of the gel containing the labeled RNA
bands were excised andforced througha disposable
syringe (20-gauge needle) into a centrifuge tube. A
volume ofelution buffer(0.01 MTris-hydrochloride,
pH 7.4, 0.4 Msodium acetate,and 0.02%SDS) twice
thegelvolume was added (usually about 1to 2 ml),
and thesamplewasmixedvigorouslyfor10 s at room
temperature. Large gel particles were removed by
centrifugationfor20min at30,000 x g(0 C).
Extrac-tion ofthegel was repeated twice, and the
superna-tants were pooled. Greater than 90% of the RNA is
eluted by this procedure. Two volumes of ethanol
were added tothe pooledsupernatants, and the white
precipitate was recoveredby centrifugation at10,000
x gfor 10min(0C). The precipitatewasresuspended
in binding buffer (see above) and bound to a0.2-g
oligo(dT)-cellulose column followedbya 10-ml wash
with binding buffer. The RNA was elutedfrom the
column with 1 to 2 ml of 0.1 mM EDTA (pH 7.4).
Sodiumacetate(pH 7.0)wasaddedto afinal
concen-trationof0.4M, andthe RNAwasprecipitatedwith2
volumes ofethanol at 100,000 x gfor 1 h (0C). The
bindingtoand elutionfromoligo(dT)-cellulose
effec-tively purifies the RNA away from a stickygel-like
material and gives excellent recovery (>90% forthe
three smaller VSV RNA species) ifperformedon an
eluted sample within a fewhours after
electrophore-S1S.
RNA fingerprinting and oligonucleotide
analysis. The homochromatography method of
fin-gerprinting employed is essentially that describedby
Barrell (1). Unlabeled carrier yeast RNA (40 Mg)was
addedtoeach3"P-labeledRNAsample prior to RNase
digestion. The samplewasevaporatedtodrynessin a
vacuum desiccator and resuspended in a solution
containing
200k4g
ofRNase TI(Calbiochem) per mlor200 Mg of RNase A (Worthington) per ml, 10 mM
Tris-hydrochloride, and1mMEDTA(pH 7.4).
Diges-tion was for 30 min at 37C. Separation in the first
dimensionwasby electrophoresis atpH3.5 on
cellu-lose acetate strips (3 by 55 cm; Schleicher and
Schuell) at 7000 V for 25 min. Separation in the
second dimension was by homochromatography on
plastic-backed thin layers (20 by 20 cm) of either
DEAE-cellulose or polyethyleneimine-cellulose
(Brinkman Instruments). Homomixture C (1),
di-gested for 10 min with 1 N KOH, was used for the
second dimension with both DEAE and
polyeth-yleneimine thin layers. Autoradiography of the
fingerprintswas from 4hto5days,dependingonthe
amount ofradioactivity used.
The polyethyleneimine plates were used only for
theRNaseTi fingerprints and gave betterresolution
of oligonucleotides than DEAE-cellulose.
Oligonu-cleotides wereeluted anddigested with0.5 NKOH,
andmononucleotideswereseparatedby
electrophore-sis at pH3.5 onWhatman 3MM paperas described by Barrell (1).
Preparationof32P-labeledrRNAmarkers.
Esch-erichia coli(strain W3110) 16Sand 23SrRNAs were
labeledwith 32P by theprocedureofCohenetal. (2),
except thatlabelingwas for 2hwith1mCi of 32P, and
tryptophan starvation was not imposed. Cells were
lysed by freeze-thawing three times in a medium
containing0.01 M Tris(pH 7.4), 0.01 M MgCl2, 0.03
mgofDNaseper ml, and 0.6mgoflysozyme perml.
Phenol extraction and ethanol precipitation were
performed as described above. 18S and 28S rRNA
wereobtained from 2x 10' CHOcellswhich had been
labeledfor 10 h with 2 mCi of "P in phosphate-free
medium.Preparationofthe RNA was asdescribedfor
VSV mRNA and was carried throughtothe stepprior
tooligo(dT)-cellulose binding.
on November 10, 2019 by guest
http://jvi.asm.org/
RESULTS
Separation and molecular weight determi-nation ofthe VSV mRNA species. To obtain
uniformly labeled VSV mRNA species, VSV-infectedCHO cellswerelabeled with32P from 1 to 4 h after infection. Actinomycin D was present to preventhostRNAsynthesis. Seventy
to ninety percent of the labeled RNA obtained from these cells bound to oligo(dT) cellulose,
indicating that it contained regions ofpoly(A).
The oligo(dT)-bound RNA was then fraction-ated on a formamide-polyacrylamide gel into the fourmajor bands seenin thegel autoradio-graph (Fig. 1). The bands, numbered 1 to 4, were reproducible, although the relative
amounts of 32P in each varied in different
ex-periments. Bands 1 and 2 generally each
con-tained from 5 to 10% of'the total radioactivity in all four bands, whereas bands 3 and 4
con-tained 20 to 40 and 30 to 50%, respectively.
RNA from the four bands were extracted from the gel andanalyzed in parallelslots of another gel (Fig. 1), showing that each is largely intact and free of contaminationbyother species.
The molecular weight ofthe mRNA in each band was estimated relative to rRNA markers
analyzed in adjacent slots on slab gels. The distances migrated bythese marker RNAs and the VSV RNAs are plotted versus molecular
weight in Fig. 2. For 23S, 18S, and 16S ribo-somal RNA, an approximately linear
relation-ship between the logarithm of' the molecular weight and the distance migrated is observed.
However, inthe higher-molecular-weight region the linear relationship does not hold, and thus molecularweight assignmentsinthis region are likely to be inaccurate. In fact, the VSV virion
RNA (about 3.5 x 106daltons, see ref'erence 11 for a discussion of various determinations) is not well resolved from 28S rRNA (1.65 x 106 daltons) on these gels.
Because of' the uncertainty of' molecular weight assignment from gel mobility, the RNA bands which had been eluted fromthe gel were
sedimented on sucrose gradients to assess mo-lecular weight by diff'erent physical criteria. The sucrose gradient profile (Fig. 3) shows that bands 2, 3, and 4 have sedimentation coeffi-cients of' approximately 18S, 15S, and 12S, respectively, values which are consistent with themolecular weight calculated from gel mobil-ity (see Table 3). However, band 1, which appearsto be a single specieswhen rerun on the gel, shows a broad distribution around 28S on
the sucrosegradients, suggesting that it may be
28S mRNA contaminated with other RNA
spe-cies.
Thus, bands 2, 3, and 4 are the RNA species which have been designated 13 to 15S on
sucrose gradients of VSV mRNA (5), whereas band 1 presumably contains mainly the 28S species which codes for the large viral protein
(8).
Fingerprint analysis of the isolated RNA species. To examine the nucleotide sequence relationships among the purified RNA species, and to examine their nucleotide sequence
com-plexities, RNase A digests of each were sepa-rated byelectrophoresis and homochromatogra-phy. The autoradiographs of these fingerprints
are shown in Fig. 4. Separation in the first dimension (cellulose acetate, pH 3.5) is mainly by charge, whereas the second dimension (ho-mochromatography) separates mainly by size with the smaller oligonucleotides moving fur-thest toward the top of each plate (1). The sequencesofthedinucleotides atthe top of the figures are indicated. Extending from these dinucleotides are lines of oligonucleotides of increasing size containing additional adenylic acid residues.
The compositions ofseveral ofthese oligonu-cleotides were determined and are indicated in the figures. Note that band 4 contains the sequences AC and AAC, but lacks larger se-quences inthe
AnC
line, whereas in band3this line is complete to A6C. Band 2 contains the sequenceAC (displaceddue to uneven chroma-tography), AAC, andA,C,
but lacks A4C. Inband 1the line is complete only toA4C. Band2
also contains a characteristic oligonucleotide doublet (numbered4and 5)which isnever seen in band 1 fingerprints. Thus, band 1 could not be a precursor of bands 2 or 3 since it lacks sequenceswhich they contain. Sequence
differ-ences are also clear in the adjacent line
AnGC,
where the band 1 and 4 fingerprints contain an oligonucleotide with the composition A4GC, whereas bands 2 and 3 lack this oligonucleotide (position indicated by arrows in fingerprints of' bands2 and 3). The large oligonucleotides near the bottom of' the plates show patterns which
are characteristic of' each RNA species, again indicating that the RNA bands are not
precur-sors orproducts of' each other.
RNaseTI f'ingerprints of bands 3 and 4 were also obtained for nucleotide sequence complex-ity analysis. These f'ingerprints are shown in Fig. 5 with sequences of the mono- and dinu-cleotides indicated. As in the RNase A f'inger-prints, the larger oligonucleotides from bands 3 and 4 differ signif'icantly. Analysis of oligonu-cleotides from both RNase A and RNase TI
f'ingerprints indicated that all four nucleotides 996
on November 10, 2019 by guest
http://jvi.asm.org/
origin
i
23
1
.III~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~
a~
FIG. 1. Autoradiograph of the VSV mRNA species separated on a formamide-polyacrylamide gel and
electrophoresis ofthe isolatedRNA bands.Electrophoresiswason3.7%oformamide-polyacrylamideslabgelsas
described in Materials andMethods.
997
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.507.57.452.80.602.2]werelabeled tothesame specific radioactivity,
indicating uniform labeling of the RNA species. Analysis of the nucleotide sequence
complex-ity of bands 2, 3, and 4 was carried out from RNase A fingerprints of band 2 and from both RNase A and Ti fingerprints of bands 3 and4.
Band 1 contained insufficient radioactivity to
permit analysis. To determine complexity,
sev-eral large isolatedoligonucleotides wereexcised
from the fingerprints, and the radioactivity in each was determined. The size of the
oligonu-cleotide was determined from analysis of its
20
I
3.
5
205 (D
10 2 0 30 40 50 60 DISTANCE MIGRATED(mm)
FIG. 2. Plot oflogarithm of molecular weight
ver-sus distance migrated for marker rRNAs and VSV mRNAs. The marker RNAs (positions indicated by points) were E. coli 23S and 16S rRNA, molecular weights of 1.1 x 106and 0.55x 106,respectively (16), and CHOcell 28S and 18S rRNAs,molecularweights of 1.65 x 106 and0.65 x 106, respectively (10).
base composition, assuming thatasingle
uridy-lateorcytidylatenucleotidewaspresentineach RNase Aoligonucleotideorthatasingle
guany-late nucleotide was present in each RNase TI oligonucleotide. The oligonucleotides selected
arenumbered on the fingerprints, and analysis
isshown inTable 1.Ineachcase, approximately
thesame radioactivity pernucleotide length is seen foroligonucleotides from the same
finger-print. The most reasonable interpretation of
thisresult is that eacholigonucleotide contains
asequence whichoccurs oncewithineach RNA
species, and that each band contains one or
more RNA moleculeswith thesame radioactiv-ity per nucleotide length. The analysis was
completed by determining the total radioactiv-ity ineach fingerprintand dividing this number by the radioactivity per nucleotide (Table 2).
Thisnumber should equal the lengthin nucleo-tides ofthe RNA molecule ifasingle molecular
species is present. For bands 2 and 3, these numbers (2,156 and 1,545 nucleotides, respec-tively) are very close to the nucleotide lengths expected for single species ofmolecular weights 0.78 x 106and0.56 x 106(Table3). This result
indicates that bands 2 and 3 are
radiochemi-cally pure RNA species. For band 4, however,
therange ofthepossiblesequencecomplexities
istwo tothree times that expected for a single
molecular species. The most likely interpreta-tionof thisresult is that band4containstwoto
'CM
0
4 8 2 16 20 24 28 32
FRACTION NUMBER
FIG. 3. Sedimentation of isolated VSV mRNA bandson SDS-sucrose gradients. 32P-labeled VSV mRNA
species isolated fromaformamide-polyacrylamidegelasdescribedinMaterialsand Methodsweresedimented
onseparate15-30%linearsucrose(wt/wt) gradients(0.5%SDS,0.1M NaCI,0.01M Tris, pH7.5,1mM EDTA) inanSW41rotorfor14hat25,000rpm.Positions oftheCHO 28S,18S,and4to5Smarker RNAsareindicated.
Symbols: 0, band1 RNA; *, band2RNA; 0, band3RNA; x, band4RNA.
i~~~~~~~~~~~~~~
I-I
998
on November 10, 2019 by guest
http://jvi.asm.org/
[image:5.507.63.259.208.324.2] [image:5.507.120.408.399.602.2]U
).
,w-2
E ' : $o
3
Nz~4G
o
E
0 G
u, BANDI1
Lceltloseacetate
3.5
I
C .4
NANDO
NANO3
ACM4
AAI
a
[image:6.507.57.452.72.464.2]BAND4
FIG. 4. Autoradiographs oftwo-dimensional separations by electrophoresis and thin layer
homochromatog-raphy(DEAEplates) ofoligonucleotides produced by completeRNaseAdigestion ofVSV mRNA bands1to 4.
The spotsjust above and to the right of the dinucleotides AU and GUare their cyclic 2',3'-phosphates.
Extensive RNaseA digestion conditions which minimize these also result in digestionafterAresidues.
three RNA species (seeDiscussion).
Size analysis of poly(A) from the fingerprints. The large oligonucleotide(s)
visi-bleattheoriginofthesecond dimensionineach
ofthefingerprints(Fig.4and 5)waselutedfrom
the thin layer and
digested
with 0.5 NKOH,
and theproductswereanalyzedby pH3.5paper
electrophoresis.Theanalysis showedthat itwas
greater than 90% adenylate and therefore
pre-sumablyrepresentsthe
poly(A)
fromeachmes-senger which has been identified in the total
VSV messenger population by other methods (4, 13).Sincethecountsperminuteper
nucleo-tideareknownforbands 2, 3, and 4, thecounts
per minute in thepoly(A) region of the
finger-prints can be used to calculate the average
length of poly(A) on each messenger
species.
The calculation
(Table
3) shows that theaver-age
poly(A)
sizeonthesespeciesrangesfrom64 to 124nucleotides.Thecalculationforband4is made assuming that twodifferent RNAspecies
are present within the band, and that the
poly(A) is equally distributed between them.
Since the counts per minute per nucleotide
length could not be determined
accurately
for the band1fingerprint, thelength
ofthepoly(A)
wascalculated byassuming that the fractionof
thetotal fingerprint radioactivity in
poly(A)
is999
---Ogg" owmikw
on November 10, 2019 by guest
http://jvi.asm.org/
ROSEANDKNIPE
G
CACw
UGS
C
~
C
0
E
0
E
_,0
O)
S
s~~~~~~*
*
"
O
vBAND 3
polylAl
I
cellulose
acetate
pH 3.5
c
Af
S_
_
+~~~~
15
bands 3and 4.
J VIROL.
1000
on November 10, 2019 by guest
http://jvi.asm.org/
SEQUENCE
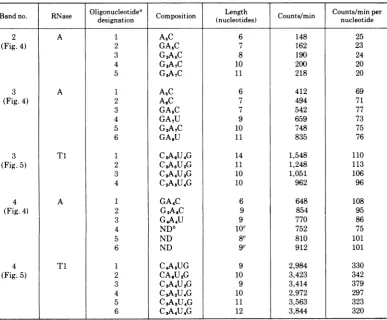
TABLE 1. Analysis of oligonucleotides in RNase A and RNase Tl fingerprints of bands 2, 3, and 4
Oligonucleotidea C Length Counts/minper
designation (nucleotides) nucleotide
2 A 1 A5C 6 148 25
(Fig. 4) 2 GA,C 7 162 23
3 G2A5C 8 190 24
4 G2A7C 10 200 20
5 G3A7C 11 218 20
3 A 1 A5C 6 412 69
(Fig. 4) 2
AsC
7 494 713 GA5C 7 542 77
4 GA7U 9 659 73
5 G2A7C 10 748 75
6 GAOU 11 835 76
3 Ti 1
C3ASU4G
14 1,548 110(Fig. 5) 2 CSA5U2G 11 1,248 113
3 CSA4U2G 10 1,051 106
4 C2ASU4G 10 962 96
4 A 1 GA4C 6 648 108
(Fig. 4) 2 G2A6C 9 854 95
3 G4A4U 9 770 86
4 NDb 1C 752 75
5 ND 8c 810 101
6 ND 9c 912 101
4 T1 1
C4A3UG
9 2,984 330(Fig. 5) 2 CA6U2G 10 3,423 342
3 C2A4U2G 9 3,414 379
4 CSA2UAG 10 2,972 297
5 CsAsU4G 11 3,563 323
6 C3A4U4G 12 3,844 320
aTheoligonucleotides includedinthetablearethose which gave minimal valuesof countsperminuteper
nucleotide length. Other oligonucleotides which gave integral multiples ofthese values were analyzed. For
example, A4Cintheband3RNase A fingerprint contained145counts/minpernucleotide andwaspresumedto
be presentintwocopies per molecule. Suchmultiple-copyoligonucleotideswereomitted from theanalysis.
bND, Not determined.
cEstimatedfrom positiononfingerprint.
TABLE 2. Calculation of nucleotide sequence complexities and poly(A) length for bands 2, 3, and 4
Counts/minper Totalcountsper Countspermin
nucleotide Total Counts/min in min/countsper inpoly(A)/ Bandno. RNase (average or counts/minin poly(A) min per countsper
range) fingerprinta nucleotide mnu per
nucleotide
2 A 23 49,600 2,871 2156 124
3 A 73 110,539 5,100 1514 70
3 Ti 106 167,130 6,105 1576 58
4 A 75-101 210,424 15,909 2,083-2,805 170
4 Ti 297-379 799,812 61,810 2,110-2,692 180
aDeterminedby excising allradioactiveregions
fromi
the thinlayer and counting theminascintillationspec-trometer. InRNaseAfingerprints, thelossofCMP (not transferred to2nddimension) wascorrectedfor.
the fractional length of the molecule which is poly(A). The major species present is assumed to be 28S with a molecular weightnear 1.65 x 106(Fig. 2and 3),corresponding toabout4,500
nucleotides, and the fraction ofthetotal 32pin poly(A) iscalculatedtobealengthofpoly(A)of
about 110nucleotides.
Determination of average poly(A) length for
1001
on November 10, 2019 by guest
http://jvi.asm.org/
[image:8.507.63.451.86.406.2]TABLE 3. Molecularweights, nucleotide sequencecomplexities, poly(A)content, andcoding capacities ofthe VSVmRNAspecies
Mol wt from Sedimentation Nucleotide Avg poly(A) Amino acidcoding
Band no. rr- sequence
.e.hcapacities
gelmobility coefficient complexity length (daltons)
1 >1.65 x 106 Heterogeneous NDa 110 170,0OOb
(4,600nucleotides) (28S)
2 0.70 x 106 18S 2,156 124 74,500c
(1,940nucleotides)
3 0.55 x 106 15S 1,545d 64 54,200c
(1,530nucleotides)
4 0.28 x 106 12S 2,100-2,800 90 74,000-100,000c
(780nucleotides)
aND, Not determined.
'Calculated frommolecularweight determined bysucrosegradient sedimentation.
cCalculatedfromnucleotidesequence complexity assuming poly(A)isnoncoding andanaverage molecular
%wightof110 foramino acids.
dAverageofRNaseAand RNase
Ti
determinations shown inTables 1 and 2.the VSV mRNA species showed considerable variation in different experiments, although it wasalways greater than50nucleotidesforeach species.This variation may reflect uncontrolled variation in the rate of poly(A) synthesis or degradation indifferent preparations.
Since no distinct RNA species were found in
the 10 to30% ofthe RNA which didnotbindto
oligo(dT)-cellulose, we conclude that all VSV mRNA species analyzed contain poly(A)
se-quences. The averagelengthsofthese sequences
are consistent with the wide range of poly(A)
sizes reported previously (4) for a mixture of
VSV mRNAs.
DISCUSSION
These results show that two-dimensional RNA fingerprinting techniques can be used to
assesssequencerelatedness oflargeviral mRNA
moleculesaswellastoexaminetheir nucleotide sequence complexity, poly(A) content, and pu-rity.
A major difficulty was encountered initially inattempting todetermine the molecular
com-plexities ofthe VSV mRNAs from fingerprints
ofRNase A digestions. Ifsufficient carrierRNA was not added during the digestion, significant digestionofthepoly(A) wasobserved aswellas
loss ofotherlargeoligonucleotides, presumably
due to a low level ofcleavage after adenylate. Thisproblem wasnot encountered with finger-prints of RNase Ti digests. Also, streaking of the RNAs during gel electrophoresis due togel overloading must be avoided, since it resultsin
contamination ofeach mRNA band with
oligo-nucleotides from the other bands and
overesti-matesofsequence complexities.
The validity of the sequence complexity de-terminations are confirmed by the agreement between complexities determined from RNase A and RNase Ti fingerprints of bands 3 and 4
and the close agreement between sequence complexity and molecular weight for bands 2 and 3. Furthermore, the RNAs extracted from bands2 and 3 code for the viral proteins G and N, respectively (accompanying paper), and their nucleotide sequence complexities are just
sufficient to encode these proteins.
The sequence complexity of band 4 RNA relative to itsmolecular weight suggested that it contained two tothree RNA species. Analysisof band 4 RNA is complicated, however, because the largeoligonucleotides analyzed are presum-ably derived from RNA species which may differ in length and counts per minute per nucleotide length. In fact, more variation in counts per minute pernucleotide was observed in band 4 oligonucleotides than in oligonucleo-tides from the other species. For this reason, only a possible range of sequence complexities is given in Table 3. That band 4 does contain at least two mRNA species is shown by its ability todirect synthesis of two VSV proteins, M and NS, which require at least twice the coding capacityof asingle mRNAofthe size of band 4. However, until the band4RNAs can be separated, their total nucleotide sequence com-plexity must be considered tentative.
Theformamide-polyacrylamide gelsseparate bands 2, 3, and 4 into species which are appar-ently of homogeneous size when analyzed by
sucrosegradient sedimentation, and their
sedi-mentation rates are consistent with molecular weightscalculated from gel mobility relative to
on November 10, 2019 by guest
http://jvi.asm.org/
markers. However, band 1 RNAshows a broad sucrose gradient profile, suggestingthat it may be a mixture of RNAs of different sizes which are not resolved on the gel. The band 1 finger-print shows that it lacks nucleotide sequences found in the smaller RNAs, and thus the
heterogeneity isnot due to contamination with smallerRNAs or large amounts of a precursor to thesmaller RNAs. We feel that the most likely
explanation for the heterogeneity of band 1 is
thatit contains mainly the28SmRNA(5, 8) as well as some large degradation products of it,
and perhaps a small amount of the 40S (+) strand copied from the entire (-) strand VSV genome (D. Baltimore, T. Morrison, M.
Stampfer, and H. Lodish, Negative Strand Virus Meet. Abstr., 1973, in press).
The total nucleotide sequence complexityof
bands 2, 3, and 4 is about 4,700 [without
poly(A)], and the 28SL protein messenger (8), which is presumably contained within band 1, must contain about 4,400 nucleotides (see
above). Thus, the total nucleotide sequence
complexity of the five monocistronic mRNAs codingforthe five viral proteins mustbe about
9,110,correspondingto atotal molecular weight of 3.2 x 106. Thus, the nucleotide sequence
complexities of the VSV mRNAs account
al-most entirely for the 3.5 x 106molecular weight ofthe (-) strand virion RNA.
ACKNOWLEDGMENTS
WethankCraig Squiresforpatient instructionin nucleo-tidesequencingtechniques and Harvey F. LodishandDavid Baltimore forhelpful discussions ofthe work. Weare espe-cially gratefultoAlanJacobsonfor instruction intheuseof formamide-polyacrylamide gels.
This work was supported by American Cancer Society grant no.VC-4D and Public Health Servicegrant AI-08814 from the National Institute of Allergy and Infectious Dis-eases.J.K.R. is apostdoctoralfellow ofthe NationalCystic Fibrosis Research Foundation, and D.K. is a predoctoral fellow ofthe National Science Foundation.
LITERATURE CITED
1. Barrell, B. G. 1971.Fractionation and sequence analysis ofradioactive nucleotides, p. 751-828. G. L. Cantoni and D. R. Davies (ed.), Procedures in nucleic acid research,vol.II. Harper and Row, New York.
2. Cohen,P.T., M. Yaniv,and C.Yanofsky. 1973. Nucleo-tidesequencesfrom messenger RNAtranscribedfrom theoperator-proximalportionofthetryptophanoperon ofEscherichiacoli.J. Mol. Biol.74:163-177. 3. Duesberg,P.H., andP. K.Vogt. 1973.Gel
electrophore-sisofavianleukosisand sarcomaviralRNA in formam-ide: comparisonwithotherviralandcellular species. J. Virol. 12:594-599.
4. Ehrenfeld, E., and D. Summers. 1972. Adenylate-rich sequencesinvesicularstomatitisvirusmessenger ribo-nucleic acid. J. Virol.10:683-688.
5. Huang,A., D. Baltimore,and M. Stampfer.1970. Ribo-nucleicacidsynthesisof vesicularstomatitis virus.III. Multiple complementary messenger RNA molecules. Virology 42:946-957.
6. Huang, A.,J.Greenwalt, and R. Wagner.1966.Defective Tparticlesofvesicular stomatitis virus.I.Preparation, morphology and some biological properties. Virology 30:161-172.
7. Huang, A., andR. Wagner.1966.Comparative sedimen-tation coefficients of RNA extracted from plaque-form-ing and defective particles of vesicular stomatitis virus. J.Mol.Biol. 22:381-384.
8. Morrison, T., M. Stampfer, D. Baltimore, and H. F. Lodish. 1974.Translationof vesicularstomatitis mes-senger RNA byextracts from mammalian and plant cells. J. Virol. 13:62-72.
9. Penman, S. 1966. RNA metabolism in the HeLa cell nucleus. J. Mol. Biol. 17:117-130.
10. Petermann, M. L., andA.Pavlovec. 1966.Thesubunits and structural ribonucleic acids of Jensen sarcoma ribosomes.Biochim. Biophys. Acta114:264-276. 11. Repik, P., andD. H. L.Bishop. 1973. Determinationof
molecular weights ofanimal RNA viral genomes by nucleasedigestion.I.Vesicularstomatitis virus and its defective Tparticle.J. Virol. 12:969-983.
12. Schincariol, A., and A. Howatson. 1972. Replicationof vesicularstomatitis virus. II. Separation and charac-terization of virus specific RNA species. Virology 49:766-783.
13. Soria, M., and A.Huang. 1973. Association ofpoly(A) with mRNA of VSV. J. Mol. Biol.77:449-455. 14. Stampfer, M., D. Baltimore, and A. Huang. 1971.
Ab-sence ofinterferenceduring highmultiplicityinfection by clonally purifiedvesicular stomatitis virus. J. Virol. 7:407-4 11.
15. Stampfer, M., A. Huang,and D. Baltimore. 1969. Ribo-nucleic acid synthesisofvesicular stomatitis virus. I. Species ofribonucleic acid found in Chinese hamster ovarycells infected withplaque-formingand defective particles.J. Virol. 4:154-161.
16. Stanley,W.M., Jr.,and R. M. Bock. 1965. Isolation and physical propertiesoftheribosomal ribonucleic acid of Escherichia coli.Biochemistry4:1302-1311.
17. Wagner, R., A. Levy, R. Snyder, G. Ratcliff, and D. Hyatt. 1963. Biological propertiesoftwoplaque vari-ants of vesicular stomatitis virus. J. Immunol. 91:112-122.
1003