0022-538X/05/$08.00⫹0 doi:10.1128/JVI.79.16.10547–10560.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Primary Sooty Mangabey Simian Immunodeficiency Virus and Human
Immunodeficiency Virus Type 2
nef
Alleles Modulate Cell Surface
Expression of Various Human Receptors and Enhance Viral
Infectivity and Replication
Jan Mu
¨nch,
1† Michael Schindler,
1† Steffen Wildum,
1† Elke Ru
¨cker,
1Nicola Bailer,
1Volker Knoop,
2Francis J. Novembre,
3and Frank Kirchhoff
1*
Abteilung Virologie, Universita¨tsklinikum, 89081 Ulm, Germany1; Institut fu¨r Zellula¨re und Molekulare Botanik,
Universita¨t Bonn, 53115 Bonn, Germany2; and Yerkes National Primate Research Center,
Emory University, 954 N. Gatewood Rd., Atlanta, Georgia 303223
Received 26 October 2004/Accepted 24 May 2005
Thenef gene of the pathogenic simian immunodeficiency virus (SIV) mac239 clone has been well charac-terized. Little is known, however, about the function ofnefalleles derived from naturally SIVsm-infected sooty mangabeys (Cercocebus atys) and from human immunodeficiency virus type 2 (HIV-2)-infected individuals. Addressing this, we demonstrate that, similarly to the SIVmac239nef, primary SIVsm and HIV-2nefalleles down-modulate cell surface expression of human CD4, CD28, CD3, and class I or II major histocompatibility complex (MHC-I or MHC-II, respectively) molecules, up-regulate surface expression of the invariant chain (Ii) associated with immature MHC-II, inhibit early T-cell activation events, and enhance virion infectivity. Both also stimulate viral replication, although HIV-2nefalleles were less active in this assay than SIVsmnefalleles. Mutational analysis showed that a dileucine-based sorting motif in the C-proximal loop of SIV or HIV-2 Nef is critical for its effects on CD4, CD28, and Ii but dispensable for down-regulation of CD3, MHC-I, and MHC-II. The C terminus of SIV and HIV-2 Nef was exclusively required for down-modulation of MHC-I, further demonstrating that analogous functions are mediated by different domains in Nef proteins derived from different groups of primate lentiviruses. Our results demonstrate that none of the eight Nef functions investigated had been newly acquired after cross-species transmission of SIVsm from naturally infected mangabeys to humans or macaques. Notably, HIV-2 and SIVsmnefalleles efficiently down-modulate CD3 and C28 surface expression and inhibit T-cell activation more efficiently than HIV-1nefalleles. These differences in Nef function might contribute to the relatively low levels of immune activation observed in HIV-2-infected human individuals.
Both simian immunodeficiency virus (SIVmac) infection of captive rhesus macaques and human immunodeficiency virus type 2 (HIV-2) infection of humans originate from zoonotic transmission of SIVsm in naturally infected sooty mangabeys (16, 22). SIVsm usually causes a persistent but asymptomatic infection in its natural simian host (48, 54), although the de-velopment of AIDS in a sooty mangabey after an 18-year natural SIVsm infection has recently been reported (35). In comparison, HIV-2 is frequently pathogenic in humans, but infected individuals progress more slowly to AIDS than HIV-1-infected individuals (37, 44). Finally, infection of macaques with SIVmac typically results in rapid immunosuppression and represents one of the best animal models for AIDS in humans (reviewed in references 13 and 15). The well-characterized molecular SIVmac239 clone (44) has been proven particularly useful in studying the determinants of viral pathogenicity
be-cause it usually be-causes simian AIDS within 1 year after infec-tion (28).
More than a decade ago, it was demonstrated that a large deletion in the wild-type SIVmac239 (239wt)nefgene results in low viral loads and a strongly attenuated clinical course of infection (29). Subsequent in vitro studies identified a variety of 239wt Nef activities, including down-regulation of cell sur-face expression of CD4 (4, 24), CD3 (3, 23), CD28 (60), major histocompatibility complex class I (MHC-I) (58), and MHC-II (51, 57), up-regulation of the invariant chain (Ii) associated with immature MHC-II complexes (51, 57), enhancement of virion infectivity (34), and stimulation of viral replication (1, 20, 36). Accumulating evidence indicates that many of these Nef functions are genetically separable and contribute to the maintenance of high viral loads and to disease progression in SIVmac-infected rhesus macaques (14, 26, 39, 40, 52).
The 239wtnefgene has been thoroughly characterized (29, 47, 52). Much less is known, however, about the function of primary SIVsm and HIV-2 nefalleles. As mentioned above, Nef is multifunctional and a major pathogenesis factor in SIV-mac239-infected rhesus macaques (29). SIVsm strains also contain intactnefgenes, but infected mangabeys usually do not develop AIDS despite high levels of viral replication (35, 48, 54). Immunodeficiency viruses are highly variable, and Nef
* Corresponding author. Mailing address: Abteilung Virologie, Uni-versita¨tsklinikum, Albert-Einsteinallee 11, D-89081 Ulm, Germany. Phone: 49-731-50023344. Fax: 49-731-50023337. E-mail: frank.kirchhoff @medizin.uni-ulm.de.
† The first three authors are in alphabetical order and contributed equally to this work.
10547
on November 8, 2019 by guest
http://jvi.asm.org/
function might have changed after cross-species transmission of SIVsm from mangabeys to macaques or humans. Therefore, we investigated the functional activity of primary SIVsm and HIV-2nefalleles in eight previously established in vitro assays for Nef function. We found that SIVsmnefalleles down-mod-ulate cell surface expression of human CD4, CD28, CD3, and class I or II MHC molecules, up-regulate surface expression of the Ii typically associated with immature class II MHC, and enhance viral replication. They were more active than HIV-1
nef alleles in down-regulating CD3 and CD28 from the cell surface and in inhibiting T-cell receptor (TCR) signaling. Compared to SIVsm, the activity of HIV-2nefalleles was more variable and the ability to stimulate viral replication was sig-nificantly reduced. However, as a group the HIV-2 Nefs were capable of performing all eight functions investigated. Our findings indicate that no additional Nef functions were ac-quired after zoonotic transmission of SIVsm from sooty mangabeys to humans or macaques. Concordant with a previ-ous report (61), we found that some HIV-2 nef genes are truncated or nonfunctional, suggesting that a lack of Nef func-tion might contribute to the limited pathogenicity of HIV-2 in some infected individuals.
MATERIALS AND METHODS
Nef expression plasmids.The generation of bicistronic cytomegalovirus-based pCG vectors coexpressing green fluorescent protein (GFP) and the SIVmac239 or HIV-2 ROD or BENnefalleles has been described previously (51). To generate thenef-defective control vector, 239wtnefwas amplified with p239 nef-Xba (5⬘-TTTTTCTAGAgTGGGTtGAGCTATTTCCtaGAGGCGGT) and
[image:2.585.44.541.81.296.2]p239MluI (5⬘-GTCCCTACGCGTCAGCGAGTTTCCTTCTTG-3⬘) (mutated positions are lowercase; underlining represents the XbaI and MluI restriction sites), resulting in an M1V change and introducing two premature stop codons at positions 3 and 7 of thenefreading frame. The HIV-2 CBL23nef(53) was PCR amplified using primers HIV-2Xba1 (5⬘-CGGTCTAGATGCAATATGG GTGCGAG-3⬘) and pHIV-2MluI (5⬘-CCACGCGTTAACTAAATGGTATTC CT-3⬘) (underlining represents XbaI and MluI restriction sites). The remaining HIV-2nefgenes were amplified from the CDC 310248, 310072, 310319, and 60415K strains (17, 42) obtained through the AIDS Research and Reference Reagent Program, Division of AIDS, National Institute of Allergy and Infectious Diseases, National Institutes of Health, from Mark Rayfield, Stephen Wiktor, Feng Gao, and Beatrice Hahn. Primers were pHIV-2Xba2 (5⬘-CGGTCTAGA TGGGATCAGCTGGTTC-3⬘) or pHIV-2Xba3 (5⬘-CGGTCTAGATGGGTGC GAGTGGAT-3⬘) (for 60415K) paired with either p310248Mlu (5⬘-GTACGCG TCTAATCTGTAGGTATTCCTCTT-3⬘), p310319Mlu (5⬘-GTACGCGTCTAA TCTGTTGGTATTCCCCTT-3⬘), p7924AMlu (5⬘-GTACGCGTTCAGTTAAA TGGAATTCCCCTT-3⬘), or p60415KMlu (5⬘-GTACGCGTTTAATTAAATG GTATTCCT-3⬘). SIVsmnefalleles were amplified by nested PCR from the blood of three naturally infected sooty mangabeys, designated FWr, FYr, and FFm (Table 1). The inner primers pSIVsm(Xba) (5⬘-ACCTATCTAGAGCCC TTATGGGTGGCGTTACC-3⬘) and SIVsm(MluI) (5⬘-GTCCCTACGCGTCA GCTTGTTTCCTTCTTG-3⬘) introduced XbaI and MluI restriction sites (und erlined) flanking thenefgene. These sites were used for cloning into the bicist ronic pCG vector coexpressing Nef and GFP via an internal ribosome entry site element (21). Five to six individualnefclones per SIVsm or HIV-2 sample were sequenced, and one or two representative alleles were selected for functional analysis (Table 1). Notably, the SIVsm strains were never passaged on human cells. For Western blot analysis, we also amplified allnefopen reading frames (ORFs) with primers resulting in fusion of the AU1 peptide tag to the C terminus of Nef. Site-directed mutagenesis ofnefwas performed by splice overlap ext ension PCR essentially as described previously (26, 39, 40). All PCR-derived inserts were sequenced to confirm that no undesired nucleotide changes were present.
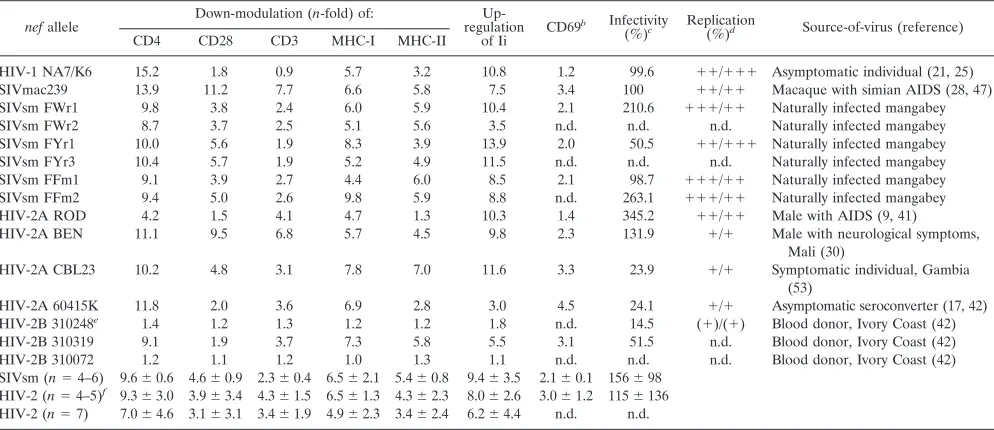
TABLE 1. Functional activity of SIVsm and HIV-2nefallelesa
nefallele
Down-modulation (n-fold) of: Up-regulation
of Ii CD69
b Infectivity
(%)c Replication
(%)d Source-of-virus (reference)
CD4 CD28 CD3 MHC-I MHC-II
HIV-1 NA7/K6 15.2 1.8 0.9 5.7 3.2 10.8 1.2 99.6 ⫹⫹/⫹⫹⫹ Asymptomatic individual (21, 25) SIVmac239 13.9 11.2 7.7 6.6 5.8 7.5 3.4 100 ⫹⫹/⫹⫹ Macaque with simian AIDS (28, 47) SIVsm FWr1 9.8 3.8 2.4 6.0 5.9 10.4 2.1 210.6 ⫹⫹⫹/⫹⫹ Naturally infected mangabey
SIVsm FWr2 8.7 3.7 2.5 5.1 5.6 3.5 n.d. n.d. n.d. Naturally infected mangabey
SIVsm FYr1 10.0 5.6 1.9 8.3 3.9 13.9 2.0 50.5 ⫹⫹/⫹⫹⫹ Naturally infected mangabey SIVsm FYr3 10.4 5.7 1.9 5.2 4.9 11.5 n.d. n.d. n.d. Naturally infected mangabey SIVsm FFm1 9.1 3.9 2.7 4.4 6.0 8.5 2.1 98.7 ⫹⫹⫹/⫹⫹ Naturally infected mangabey SIVsm FFm2 9.4 5.0 2.6 9.8 5.9 8.8 n.d. 263.1 ⫹⫹⫹/⫹⫹ Naturally infected mangabey
HIV-2A ROD 4.2 1.5 4.1 4.7 1.3 10.3 1.4 345.2 ⫹⫹/⫹⫹ Male with AIDS (9, 41)
HIV-2A BEN 11.1 9.5 6.8 5.7 4.5 9.8 2.3 131.9 ⫹/⫹ Male with neurological symptoms, Mali (30)
HIV-2A CBL23 10.2 4.8 3.1 7.8 7.0 11.6 3.3 23.9 ⫹/⫹ Symptomatic individual, Gambia (53)
HIV-2A 60415K 11.8 2.0 3.6 6.9 2.8 3.0 4.5 24.1 ⫹/⫹ Asymptomatic seroconverter (17, 42) HIV-2B 310248e 1.4 1.2 1.3 1.2 1.2 1.8 n.d. 14.5 (⫹)/(⫹) Blood donor, Ivory Coast (42)
HIV-2B 310319 9.1 1.9 3.7 7.3 5.8 5.5 3.1 51.5 n.d. Blood donor, Ivory Coast (42) HIV-2B 310072 1.2 1.1 1.2 1.0 1.3 1.1 n.d. n.d. n.d. Blood donor, Ivory Coast (42) SIVsm (n⫽4–6) 9.6⫾0.6 4.6⫾0.9 2.3⫾0.4 6.5⫾2.1 5.4⫾0.8 9.4⫾3.5 2.1⫾0.1 156⫾98
HIV-2 (n⫽4–5)f 9.3⫾3.0 3.9⫾3.4 4.3⫾1.5 6.5⫾1.3 4.3⫾2.3 8.0⫾2.6 3.0⫾1.2 115⫾136
HIV-2 (n⫽7) 7.0⫾4.6 3.1⫾3.1 3.4⫾1.9 4.9⫾2.3 3.4⫾2.4 6.2⫾4.4 n.d. n.d.
aDown-modulation (n-fold) of CD4, CD28, CD3, MHC-I, and MHC-II and up-regulation (n-fold) of Ii surface expression levels were determined for cells expressing
high levels of GFP and calculated as previously described (7, 49, 50). In all analyses, at least 500 cells expressing high levels of GFP were analyzed, and the results were confirmed in three to five independent experiments (standard deviations were generally⬍10%).
bValues given-fold reduction of CD3-induced CD69 expression and were determined for cells expressing medium levels of GFP. cViral infectivity was determined as described in the legend to Fig. 8. n.d., not done.
dThe efficiency of virus replication was measured in 221-89 cells in the presence/absence of IL-2. The results represent reverse transcriptase production measured
in the 221-89 culture supernatants at seven time points between 3 and 17 days postinfection with virus stocks containing 1 ng p27 antigen. Values were obtained from duplicate (with IL-2) or quadruplicate (without IL-2) infections of 221-89 cells with two different virus stocks. The following symbols indicate reverse transcriptase production:⫹⫹,⬍120%;⫹⫹, 80 to 120%;⫹, 50 to 80%, and (⫹), 30 to 50% compared to 239wt-infected cells. The SIVmac239 nef* mutant containing a disrupted nefORF replicated with about 30% of 239wt efficiency.
eSubtype B is based on Nef analysis;envclusters with HIV-2B strains (42).
fThe inactive 310248 and the truncated 310072nefalleles were excluded from the analysis.
on November 8, 2019 by guest
http://jvi.asm.org/
Proviral constructs.A modified form of the Nef-SHIV-K6 clone (31) was used for the insertion of HIV-2 and SIVsmnefgenes into the SIVmac239 genome. This chimeric SIVmac239 construct contains the HIV-1 K6nefallele down-stream ofenvand upstream of the polypurine tract and the core enhancer region of the SIVmac239 long terminal repeat. The original SIVmac239nefinitiation codon and a second ATG at codon 7 of thenefORF are mutated without changing the coding sequence of the overlappingenvgene (31). In addition, we eliminated the SacI restriction site at position 6015 of the proviral SIVmac239 sequence (M33262) without altering the predicted Vif sequence. All HIV-2 and SIVsm nefalleles were PCR amplified with 5⬘ primers containing the SacI restriction site (underlined) located upstream of the SIVmac239envtermination codon (bold) and the first 18 nucleotides of the respectivenef genes (e.g., pHIV-2Sac1, 5⬘-GGGCTTGAGCTCACTCTCTTGTAAGATGGGTGCGAGT GGATCC-3⬘) together with a 3⬘primer introducing an MluI restriction site just downstream ofnef. All PCR fragments were cloned into the proviral SIVmac239 vector using the SacI and MluI sites. Thus, the resulting proviral SIVmac239 clones do not contain overlappingenv-nefsequences and differ exclusively by their Nef coding sequences.
Cell culture and transfection.HeLa CIITA and Jurkat T cells were cultured as described previously (7, 50, 51, 57). Transfection of Jurkat T cells was performed using the DMRIE-C reagent (Gibco BRL) following manufacturer’s instructions. HeLa CIITA cells were transfected with Metafectene (Biontex). Briefly, 2.5g DNA in 100l optimized minimum essential medium (OMEM; Invitrogen) was mixed with 10l Metafectene in 90l OMEM and incubated for 30 min at room temperature. Subsequently, the mixture was added to 2⫻105cells and incubated for 6 h at 37°C. Thereafter, the medium was changed, and cells were analyzed by fluorescence-activated cell sorter on the following day. Transfection efficiencies varied between 20% and 35%.
Western blotting. 293T cells were transfected with 5g pCG expression vectors coexpressing GFP and Nef using the calcium phosphate method as described previously (7). Two days posttransfection, cells were pelleted and lysates were generated and separated through 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Expression of Nef proteins in whole cellular lysates was analyzed by immunoblotting. Nef was detected with 1:500 diluted mouse anti-AU1 antibody (Ab) (Covance).-Actin and GFP were detected with 1:2,000 dilutions of rabbit anti--actin and rabbit anti-GFP Abs (Abcam). West-ern blotting was performed using KPL’s protein detector kit as recommended by the manufacturer with 1:500 dilutions of alkaline phosphatase-coupled goat anti-mouse and goat anti-rabbit Abs (Jackson Immuno Research).
Flow cytometry.CD4, MHC-I, CD28, CD3, and GFP reporter molecules in Jurkat T cells transfected with a bicistronic vector coexpressing Nef and GFP (21) were measured as described previously (7, 52). Down-modulation of MHC-II and up-regulation of Ii were measured on transfected HeLa CIITA cells (50, 57). The following phycoerythrin-conjugated antibodies were used: human CD4, human CD3, and LeuTM-28 (BD Biosciences); anti-CD74/R-PE M-B741 (Ancell); anti-HLA-ABC antigen/RPE (DAKO); mouse anti-human HLA-DR TUE36 (Caltag laboratories); and L243 (BD Biosciences). Nef-mediated down- or up-regulation of cellular receptors was quantitated as described previously (7, 52). To test the inhibitory effect on T-cell activation, Jurkat cells were transfected as described previously (7, 52), except that no phytohemagglutinin (PHA) was added to the culture medium. Approximately 24 h posttransfection, cells were stimulated using 0.3g/ml HIT3a monoclonal antibody (MAb) (BD Biosciences), specific for theεsubunit of CD3, or by the addition of 1g/ml PHA and cultured for an additional 15 h prior to flow cytometric analysis of CD69 expression (clone FN50; BD Biosciences).
Intracellular Nef staining.Jurkat T cells were permeabilized as described previously (51). Intracellular Nef expression was detected with an anti-AU1 Ab, and two-color flow cytometric analysis for Nef and GFP expression was per-formed as mentioned above.
Virus stocks, infectivity, and replication.For virus production, 293T cells were transfected by the calcium phosphate method with 10g of the proviral con-structs as described previously (7, 40). The medium was changed after overnight incubation, and virus was harvested 24 h later. The p27 antigen concentrations were quantified by using an SIV enzyme-linked immunosorbent assay provided by the National Institutes of Health AIDS Reagent Program. Cells were infected with virus stocks containing normalized quantities of p27 antigen, and virus production was measured by reverse transcriptase assay at 2- to 3-day intervals as described previously (8, 26). The rhesus T-cell line 221-89 (1) was maintained in the presence of 100 U of interleukin-2 (IL-2)/ml (Boehringer) and 20% fetal calf serum. Infections were performed in the absence of exogenous IL-2 or in the presence of 50 U IL-2/ml and 5% fetal calf serum. Virus infectivity was deter-mined using TZM-BL and P4-CCR5 cells as described previously (12, 46).
Infectivity was measured on P4-CCR5 and TZM-BL cells using the -galactosi-dase screen from TROPIX as recommended by the manufacturer.
Statistical methods.The mean activities of HIV-2 and SIVsmnefalleles were compared by using Student’sttest. Similar results were obtained with the Mann-Whitney test. The software package StatView version 4.0 (Abacus Concepts, Berkeley, CA) was used for all calculations.
Phylogenetic analysis.Sequences were derived from the Los Alamos sequence database (http://hiv-web.lanl.gov). Phylogenetic tree construction was done with the minimum evolution criterion as implemented in MEGA3 (33). Poisson cor-rected amino acid distances were used assuming uniform rates over sites. The following search settings were used: CNI level⫽2 and max trees⫽20. Bootstrap node support was calculated from 10,000 replicates.
Nucleotide sequence accession numbers.The GenBank accession numbers for the HIV-2 and SIVsm sequences are DQ092758 to DQ92767.
RESULTS
Nef sequence and phylogenetic analysis. To investigate whether Nef function might have changed after cross-species transmission of SIVsm from sooty mangabeys to the new ma-caque or human hosts, we analyzed nefalleles derived from three sooty mangabeys (designated FWr, FYr, and FFm) and seven HIV-2 strains (Table 1). For comparison, we also deter-mined the activity of the SIVmac239 and HIV-1 NA7 nef
alleles, which have been well characterized in previous studies (21, 25, 26, 29, 47, 52). The SIVsmnefalleles were amplified directly from uncultured blood samples of naturally infected sooty mangabeys. Three of the HIV-2nefalleles were derived from the infectious molecular ROD10 (9, 41), BEN (30), and CBL23 (53) clones. The remainingnef genes were amplified from the HIV-2 CDC 310248, 310072, 310319, and 60415K strains (16, 42) obtained through the AIDS Reagent Program. These four HIV-2 strains have been obtained by coculture with PHA-stimulated normal peripheral blood mononuclear cells (PBMC) and were never passaged on transformed cell lines (16, 42). As summarized in Table 1, the HIV-2 strains origi-nated from several African countries and were obtained at different clinical stages of infection.
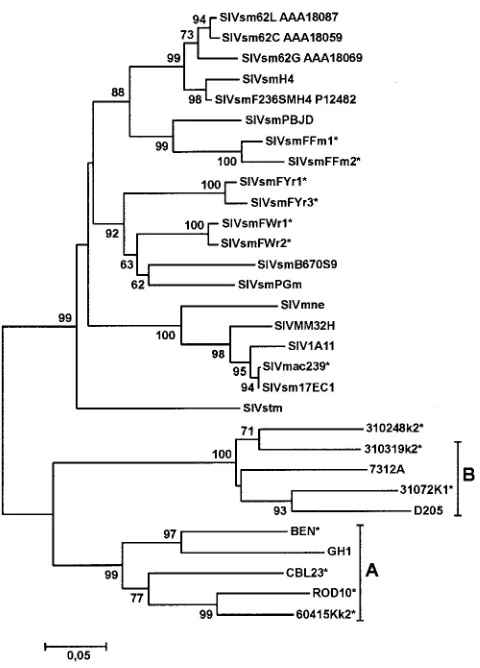
Phylogenetic analysis revealed that the HIV-2/SIV Nef se-quences formed five distinct clusters (Fig. 1). Three clusters included the various SIVsm and SIVmac Nef sequences, the fourth the HIV-2 group B strains as well as HIV-2 310248, and the fifth the HIV-2 group A ROD10, GH1, CBL23, BEN, and 60415K strains. Clustering of the 310248 Nef with that of HIV-2 group B strains was unexpected, because it has been previously demonstrated that the 310248envsequence clusters with group A ROD10 and 60415Kenvgenes (42). Thus, HIV-2 310248 might have arisen from recombination between a sub-type A and B strain. Importantly, our analysis revealed that all sequences were animal or patient specific, confirming the au-thenticity of the Nef sequences and excluding cross-sample PCR contaminations.
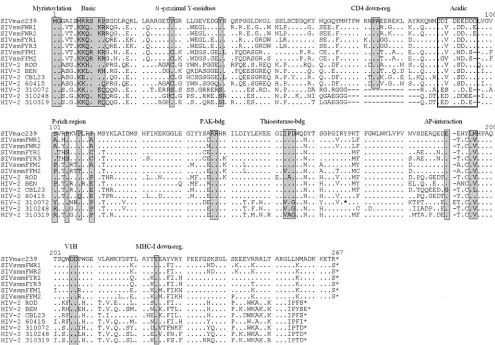
Most SIVsm and HIV-2nefgenes predicted full-length pro-teins (Fig. 2). Exceptions were the six HIV-2 310072nefORFs analyzed, which all contained a premature stop at the 149th codon (Fig. 2 and data not shown). Several domains and pro-tein interaction sites previously described to be relevant for HIV-1 function (reviewed in reference 18) were conserved in the remaining SIVsm and HIV-2 Nef sequences, including the N-terminal myristoylation signal, the acidic region, a diarginine motif, a C-proximal adaptor-protein interaction site, and a diacidic putative V1H binding site (Fig. 2). Concordant with
on November 8, 2019 by guest
http://jvi.asm.org/
their limited role for viral replication in vivo (8, 34), the N-proximal Y residues, proposed to represent endocytosis signals (6, 45), and the proline-rich region, which is involved in the interaction with cellular kinases (34), were more variable. All SIVsm FYr and HIV-2 60415K Nef sequences analyzed, for example, contain only a single proline residue at this location. In contrast, Y223, critical for MHC-I down-modulation by 239wt Nef (39, 58), was preserved at the corresponding posi-tion in all full-length SIVsm and HIV-2 Nef sequences. Resi-dues P73A74 and D204, which are important for CD4 down-modulation and efficient replication of SIVmac239 in acutely infected macaques (26), were conserved among the HIV-2 group A and SIVsm Nef sequences. Unexpectedly, however, the HIV-2 group B 310072, 310248, and 310319 Nef sequences, which are closely related to one another and were all derived from Ivory Coast blood donors (Table 1), contained a deletion of 16 amino acid residues encompassing residues P73 and A74 (Fig. 2).
Modulation of cellular receptors by HIV-2 and SIVsm Nefs.
Next, we investigated the activity of SIVsm and HIV-2 nef
alleles in modulating various human cellular receptors. nef
genes were cloned into a bicistronic vector coexpressing Nef
and GFP for fluorescence-activated cell sorter analysis (21). Of the five to six individualnefclones per SIVsm or HIV-2 sample sequenced, one or two representative alleles were selected for functional studies (Fig. 2). Western blot analysis confirmed that all bicistronic vectors containing AU1-tagged nef open reading frames expressed proteins of the expected size (Fig. 3). Altogether, however, expression levels varied, and only small amounts of HIV-2 ROD10 Nef were detected in the extracts of transfected 293T cells (Fig. 3A). Next, we used permeabilized transfected Jurkat T cells to detect both Nef and GFP expres-sion. Flow cytometric analysis showed that all HIV-2 and SIV
nef alleles analyzed were expressed, albeit with differential efficiencies (Fig. 3B). Consistent with the results of the West-ern blot analysis (Fig. 3A), expression of the HIV-2 ROD Nef in Jurkat T cells was very inefficient (Fig. 3B).
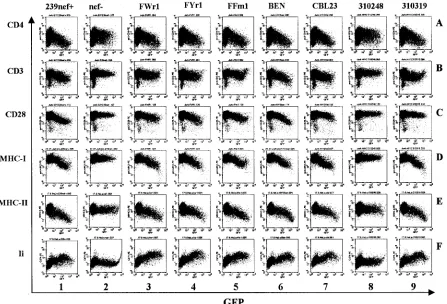
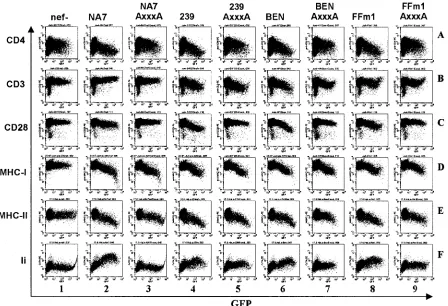
As expected (52), the 239wt Nef modulated the surface expression of all six receptors investigated, whereas a construct containing inactivating point mutations innef had no signifi-cant effects (Fig. 4, columns 1 and 2). The six SIVsmnefalleles analyzed showed comparable efficiencies in modulating cell surface expression of CD4, MHC-I, MHC-II, and Ii molecules but were less effective than 239wtnefin down-regulating CD3 and CD28 (Fig. 4, columns 3 to 5; Table 1). Similarly, the HIV-2 BEN and CBL23nefalleles affected surface expression of six cellular receptors (Fig. 4, columns 6 and 7), whereas the 310319 Nef did not efficiently down-modulate CD28 (Fig. 4, panel 9C). As expected, the severely truncated HIV-2 310072 Nef was inactive in these assays (data not shown). The 310248 Nef was also generally inactive, although the full-length pro-tein could be detected by Western blot analysis (Fig. 3). Tagged and untaggednefalleles did not differ significantly in the ability to modulate the surface expression of the six cellular factors analyzed (data not shown).
[image:4.585.45.285.66.400.2]The bicistronic vector used for flow cytometric analysis al-lows us to readily quantitate the effect of Nef on the surface expression of cellular receptors. As described previously (7, 51, 52), we calculatedn-fold up-regulation of Ii or down-modula-tion of the remaining surface receptors by dividing the mean fluorescence intensity obtained on Jurkat or HeLa CIITA cells coexpressing high levels of GFP and the differentnefalleles by the mean fluorescence obtained for cells transfected with a control construct containing a disruptednef gene. The same ranges of GFP expression (fluorescence intensities between 500 and 3,000) were used in all calculations. The activities of differentnefalleles varied considerably, particularly within the HIV-2 group, e.g., CD4 down-modulation from 4.2- to 11.8-fold between the HIV-2 ROD10 and 60415Knefalleles. Fur-thermore, the HIV-2 60415K and 310319nef alleles showed little activity in modulating CD28 and/or CD3 surface expres-sion, although they were highly active in other aspects of Nef function (Table 1). On average, however, the ability to modu-late CD4, CD28, MHC-I, MHC-II, and Ii surface expression did not differ significantly between the SIVsm and HIV-2 groups (Table 1). Notably, the five functional HIV-2nefalleles were significantly more active than the six SIVsmnef alleles analyzed in down-regulating CD3 (P ⫽0.01) (Table 1). It is also noteworthy that the 239wt Nef was more active than all remaining SIVsm and HIV-2 Nefs analyzed in down-modula-tion of CD4, CD28, and CD3 molecules. Consistent with its inefficient expression (Fig. 3), the HIV-2 ROD10 nef was
FIG. 1. Evolutionary relationships among HIV-2, SIVsm, and SIVmac Nef sequences. A phylogenetic tree was constructed using the minimum evolution criterion as implemented in MEGA3 (33). Boot-strap node support was calculated from 10,000 replicates. Nefs that were functionally characterized in this study are labeled with an aster-isk.
on November 8, 2019 by guest
http://jvi.asm.org/
poorly active in several in vitro assays, including down-modu-lation of CD4, CD28, and MHC-II (Table 1). Taken together, our results demonstrate that the ability to modulate various human receptors involved in TCR signaling and MHC antigen presentation is conserved within the SIVsm/mac/HIV-2 group. With the exception of CD3 down-modulation, SIVsm Nefs manipulated cell surface expression of human receptors with the same efficiency as HIV-2 Nefs without any adaptive changes, suggesting that functionalnefgenes likely helped the virus to evade the human immune system immediately after cross-species transmission.
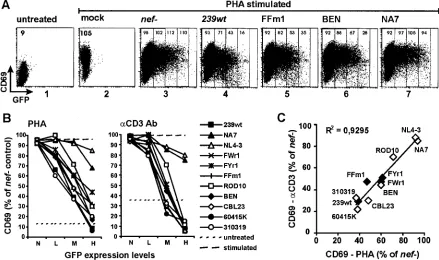
Interference of HIV-2 and SIVsm Nefs with T-cell activa-tion.Next, we utilized a transient assay (25) to study the effect of variousnefalleles on TCR-mediated T-cell activation. Stim-ulation of Jurkat T cells with PHA or CD3 MAb results in rapid induction of the early T-cell activation marker CD69 on the cell surface. Thus, induction of CD69 provides a conve-nient indicator of CD3 signaling and T-cell activation. Treat-ment of Jurkat T cells resulted in a readily detectable 5- to 10-fold increase in CD69 surface expression (Fig. 5A, compare
[image:5.585.45.540.67.412.2]panels 1 and 2). Transient transfection with bicistronic vectors allowed us to quantitate CD69 induction on cells expressing different levels of GFP and, indirectly, Nef. Cells transfected with anef-defective control construct showed similar levels of CD69 induction at different levels of GFP expression (Fig. 5A, panel 3). In contrast, the increase in CD69 expression was blocked in Jurkat T cells expressing medium to high levels of the SIVmac239, SIVsm FFm1, and HIV-2 BEN Nef proteins (Fig. 5A, panels 4 to 6). In comparison, the HIV-1 NA7 Nef had little effect on CD69 induction (Fig. 5A, panel 7). Analysis of a larger set ofnefalleles revealed that SIVmac, SIVsm, and HIV-2 Nefs efficiently inhibited both PHA- and anti-CD3 MAb-induced up-regulation of CD69 surface expression in Jurkat T cells in a dose-dependent manner, whereas HIV-1 Nefs showed little if any activity in blocking T-cell activation (Fig. 5B). The functional activities of thesenefalleles in inhib-iting PHA- and CD3-induced CD69 expression correlated sig-nificantly, suggesting that they might involve the same mech-anism (P ⬍ 0.0001) (Fig. 5C). Consistent with their higher activity in CD3 down-modulation, most HIV-2 Nefs inhibited
FIG. 2. Alignment of SIVmac239, SIVsm, and HIV-2 Nef sequences. The SIVmac239 sequence is shown in the upper row for comparison. Some conserved sequence elements in Nef, including the N-terminal myristoylation signal, N-proximal tyrosines (6, 8), PA residues known to be critical for CD4 down-modulation by 239wt Nef (26), the acidic and proline-rich regions, a diarginine motif, a C-proximal adaptor protein (AP) interaction site (10), a diacidic putative V1H binding site, and a Y residue involved in MHC-I down-regulation (39, 58), are indicated schematically. Dots indicate identity with the 239wt Nef sequence, and dashes indicate gaps introduced to optimize the alignment. The asterisk denotes the position of the premature stop codon in the HIV-2 310072 Nef. For comparison, the predicted amino acid sequence after the stop signal is shown in the alignment.
on November 8, 2019 by guest
http://jvi.asm.org/
CD69 up-regulation more efficiently than SIVsm Nefs (Table 1). However, both HIV-2 and SIVsm Nefs interfered with early T-cell activation events much more severely than HIV-1 Nefs.
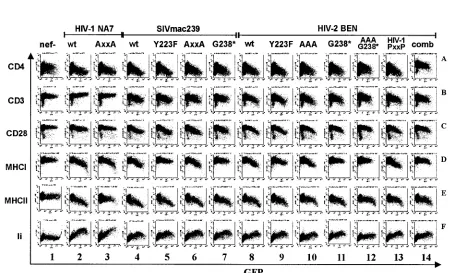
Mutational analysis of HIV-2 and SIVsm Nef proteins. It has been previously demonstrated that a dileucine motif in HIV-1 Nef is critical for down-modulation of CD4 and up-regulation of Ii but dispensable for down-up-regulation of MHC-I or -II (6, 10, 49, 57). We found that the dileucine-based sorting signal is also important for the ability of SIVsm and HIV-2 Nefs to down-modulate CD4 and to up-regulate Ii. Extending these previous studies, we investigated the functional activity of HIV-1 NA7, SIVmac239, HIV-2 BEN, and SIVsm FFm1 Nef proteins containing alanine (AXXXA) substitutions in the EXXXL motif, known to be important for adaptor protein interaction (10). These mutations also impaired down-modu-lation of CD28 (Fig. 6, row C). They had little if any disruptive effect, however, on down-regulation of CD3 and MHC-I or -II surface expression (Fig. 6, rows B, D, and E). Notably, the NA7 AXXXA Nef was entirely inactive in up-regulating Ii cell sur-face expression, whereas the mutated 239wt, BEN, and FFm1 Nefs showed some residual activity (Fig. 6, panel F, and data
not shown). Thus, compared to HIV-1 Nef, additional regions in SIVmac and HIV-2 Nef might be involved in Ii up-regula-tion. These results suggest that the “dileucine” motif is of general importance for the ability of HIV-1, HIV-2, SIVsm, and SIVmac Nefs to modulate CD4 and CD28 but dispensable for the effect on CD3, MHC-I, and MHC-II surface expression. It has been established that down-modulation of MHC-I is mediated by different domains in HIV-1 and SIVmac239 Nef (58). Three motifs in HIV-1 Nef have been implicated in MHC-I down-modulation, M20, EEEE65, and P72XXP75(5). In
contrast, the ability of the 239wt Nef to down-regulate MHC-I requires a unique C-terminal region, which is not found in HIV-1 Nef (58). Both the proline-rich and C-terminal regions are highly variable, and the C terminus of HIV-2 Nefs is trun-cated compared to SIVmac and SIVsm (Fig. 2). Several HIV-2 BEN Nef mutants were generated (Fig. 7A) to investigate whether its functional organization is reminiscent of that of HIV-1, having the same host, or of SIVmac239, showing much higher sequence similarity and belonging to the same phyloge-netic group. As expected from previous studies (34, 52), ala-nine substitutions of the central proline residues did not impair
FIG. 3. Expression of SIVsm and HIV-2 Nefs. (A) 293T cells were transfected with expression plasmids encoding the indicated AU1-tagged Nef proteins and GFP. Mock-transfected cells and a vector containing a disruptednefgene (GFP only) were used as negative controls. GFP was detected to control for transfection efficiency and-actin for total quantity of cellular protein. (B) Jurkat cells were transfected with a bicistronic vector expressing GFP and Nef or GFP alone. As described in Materials and Methods, cells were permeabilized, and Nef and GFP expression were detected simultaneously. Ranges for green fluorescence on cells defined as expressing no (N) GFP and low (L) and medium (M) levels of GFP are indicated. Compared to normal transfected Jurkat T cells, permeabilized cells contain smaller quantities of GFP. Therefore, the number of cells expressing high levels of GFP was relatively low and not used for quantitative analysis. The right panel shows the mean channel numbers of red Nef fluorescence for the three different regions of GFP expression. The results were confirmed in an independent experiment using different plasmid DNA preparations.
on November 8, 2019 by guest
http://jvi.asm.org/
239wt Nef function (Fig. 7B, column 6). In comparison, muta-tion of P72XXP75in the HIV-1 NA7 Nef reduced its activity in
down-modulating MHC-I slightly from 7.0- to 4.7-fold and MHC-II down-regulation more strongly from 7.6- to 2.3-fold at high Nef expression levels (Fig. 7B, columns 2 and 3). A Y223F mutation in HIV-2 Nef selectively impaired its ability to down-modulate MHC-I, albeit less severely than the corresponding mutation in the 239wt Nef (Fig. 7B, panels 5D and 9D) (39, 52, 58). Mutation of the three central proline (AAA) residues in HIV-2 BEN Nef had little effect (Fig. 7B, column 10), whereas C-terminal truncation (G238*) disrupted MHC-I down-mod-ulation but not other functions (Fig. 7B, column 11). We also introduced four amino acid substitutions, T103R, R105Q, R108L, and E110P, into the HIV-2 BEN Nef to evaluate whether an HIV-1 PXXP consensus region (Fig. 7A) might allow MHC-I down-modulation in the absence of the C-termi-nal region. The HIV-1 PXXP mutant Nef was fully functioC-termi-nal in all six assays (Fig. 7B, column 13). Similar to the effects on wild-type BEN and 239 Nef function, however, C-terminal truncation disrupted its effect on MHC-I (Fig. 7B, column 14). Further analysis revealed that C-terminal truncation of FFm1 Nef also impaired MHC-I down-modulation (data not shown). Our results are further evidence that different groups of pri-mate immunodeficiency viruses utilize different Nef domains to perform analogous activities and demonstrate that the
C-terminal region is required for MHC-I down-modulation by both SIV and HIV-2 Nef proteins.
Enhancement of viral infectivity and replication.Next, we investigated whether HIV-2 and SIVsm nef alleles also en-hance viral infectivity. AllnefORFs were inserted downstream of theenvgene of the proviral SIVmac239 clone as previously described for HIV-1nefgenes (31), except that the SacI site located at the 3⬘end ofenvwas used for cloning (Fig. 8A). In these constructs, the original SIVmac239nefinitiation codon and a second ATG at codon 7 of thenef ORF are mutated without changing the predicted Env sequence (31). Sequence and functional analysis confirmed that the resulting proviral SIVmac239 constructs did not contain undesired changes in
[image:7.585.71.518.71.375.2]env and expressed the appropriate HIV-2 and SIVsm Nef proteins (data not shown). As a positive control, otherwise isogenic constructs containing thenefgenes of 239wt, HIV-1 NL4-3, or the molecular SHIV-40K6 clone were generated. The 40K6 nef allele was derived from a Nef-SHIV-infected macaque with simian AIDS and is closely related to the HIV-1 group M subtype B Nef consensus sequence (31). As a negative control, we inserted anefallele containing a premature stop codon at position 93 of thenefORF (239nef*). Compared to the parental SIVmac239 clone, the genomic size of these con-structs is enlarged by about 170 nucleotides, and thenefATG initiation codon is located at a different position because of the
FIG. 4. Modulation of human cell surface receptors by SIVsm and HIV-2nefalleles. Jurkat T (rows A to D) or HeLa CIITA (rows E and F) cells were transfected with bicistronic vectors coexpressing the indicatednefalleles and GFP and assayed for surface expression of CD4, CD3, CD28, MHC-I, MHC-II, and Ii. The results were confirmed in two independent experiments and using the AU1-tagged Nef proteins.
on November 8, 2019 by guest
http://jvi.asm.org/
elimination of theenv-nefoverlap. On average, the infectivity of the modified SIVmac239 was approximately three- to four-fold reduced compared to that of the parental virus (Fig. 8B). Nevertheless, the construct containing an intact 239wtnefgene showed about 10-fold-higher activity than the otherwise iso-genic nef-defective form (Fig. 8B). Thus, this experimental system allows us to study the effect of variousnefalleles on SIVmac infectivity without the complications of overlapping
env sequences. Functional assays revealed that the 239wt, HIV-1 K6 and NL4-3, SIVsm FWr1, FYr1, FFm1, and FFm2, and HIV-2 ROD10, BEN, and 310319 nef alleles efficiently enhanced viral infectivity (Fig. 8C). In comparison, the HIV-2 CBL23 and 60415Knefgenes were only marginally active and the 310248 Nef was inactive in enhancing virion infectivity. These data demonstrate that some HIV-2 and SIVsmnefgenes increase virion infectivity with high efficiency.
Finally, we investigated the functional activity of HIV-2 and SIVsm nef alleles in enhancing viral replication. All SIV-mac239 nef recombinants replicated efficiently in CEMx174 cells (data not shown). The T-lymphoid cell line 221-89, de-rived from a rhesus monkey infected with herpesvirus saimiri (1), was utilized to evaluate the stimulatory effects of the var-iousnefalleles on SIVmac replication. In our previous studies, the abilities of variousnefalleles to stimulate SIV replication in 221-89 cells and in rhesus- or human-derived PBMC always correlated (8, 34, 40, 58), indicating that both experimental
[image:8.585.72.511.71.331.2]systems might measure analogous effects of Nef (1). However, the 221-89 assay is less variable, because SIV replication in PBMC is to some extent dependent on the blood donor. Intact 239wt and K6nefalleles enhanced viral replication moderately in the presence of exogenous IL-2 (Fig. 9, left panels) and more strongly in the absence of IL-2 (Fig. 9, right panels). The four SIVsmnefalleles consistently stimulated SIVmac replica-tion in 221-89 cells with high efficiency (representative example shown in Fig. 9A). With the exception of the 310248 Nef, the HIV-2 Nefs also enhanced viral replication (Fig. 9B). How-ever, the ROD10, BEN, CBL23, and 60415Knefalleles were clearly less active than the four SIVsmnefalleles in enhancing viral replication in both the presence and absence of exoge-nous IL-2 (Fig. 9 and Table 1). This observation is consistent with previous studies showing that the efficiency of HIV-1 replication correlates with Nef activity in CD4 down-modula-tion but not enhancement of viral infectivity. On average, the HIV-2 nef alleles also enhanced viral replication in human PBMC less efficiently than SIVsmnefgenes (data not shown). Because they were highly active in other functional aspects, it was unexpected that the HIV-2nefalleles enhanced viral rep-lication only slightly compared to the nef-defective controls. Our results demonstrate that primary SIVsmnefgenes are able to stimulate viral replication in rhesus cells. Although our data are suggestive, a larger number of samples needs to be ana-lyzed to elucidate whether HIV-2 Nefs are typically less active
FIG. 5. HIV-2 and SIVsmnefalleles inhibit CD69 induction more severely than HIV-1nefalleles. (A) Jurkat cells were either mock transfected (panels 1 and 2), transfected with a plasmid expressing GFP alone (panel 3), or transfected with plasmids expressing GFP together with the 239wt, FFm1, BEN, and NA7nefalleles (panels 4 to 7) and cultured overnight in the absence (panel 1) or presence (panels 2 to 7) of PHA and stained for CD69 expression. Ranges for green fluorescence on cells defined as expressing no GFP (N) and low (L), medium (M), or high (H) levels of GFP are indicated in panels 3 to 7. Values give mean fluorescence intensity. Similar results were obtained in two independent experiments. (B) Inhibition of CD69 induction by the indicatednefalleles. CD69 expression levels on cells coexpressing different levels of GFP and Nef are shown relative to those measured on Jurkat T cells transfected with the control construct expressing only GFP. (C) Correlation between inhibition of PHA- and CD3-induced CD69 expression. Calculations were performed for Jurkat T cells expressing medium levels of GFP and the indicated
nefalleles. Average values derived from three independent experiments are shown in panels B and C.
on November 8, 2019 by guest
http://jvi.asm.org/
in enhancing viral replication than those derived from other groups of primate lentiviruses.
DISCUSSION
In the present study, we demonstrated that, similarly to SIVmac239 Nef (52), SIVsmnefalleles derived from naturally infected sooty mangabeys are able to modulate cell surface expression of human CD4, CD3, CD28, MHC-I, MHC-II, and Ii molecules and to enhance virion infectivity and replication. Thus, SIVsm Nefs derived from primary isolates can modulate cell surface expression of human receptors without adaptive changes following animal or cell culture passage. These results resemble those of our previous study showing that primary SIVcpz nef alleles also did not require any changes to be functionally active in human cells and likely enhanced viral replication and persistence immediately after cross-species transmission to the new human host (32). We found that most HIV-2 Nefs were also functional (Table 1). Thus, none of the eight Nef functions investigated have been newly acquired or lost after cross-species transmission from sooty mangabeys to humans or macaques. It is remarkable, however, that all HIV-2 and SIVsm nef alleles investigated were less active than the SIVmac239 Nef in down-modulating CD4, CD28, and CD3 cell surface expression. Taken together, these data show that primary SIVsm Nefs are able to perform a variety of functions that most likely contribute to immune evasion and efficient
persistence in vivo (26, 39, 40, 52). Concordant with this, SIVsm infection of sooty mangabeys is characterized by high-level viremia and efficient viral replication (48, 54).
Although sooty mangabeys are not entirely resistant to SIV-induced disease (35), they are clearly less susceptible to the development of immunodeficiency than infected rhesus ma-caques, which often develop disease within 1 year after infec-tion (28). In comparison, HIV-2 causes AIDS in humans with an average incubation period of about 14 to 16 years (37, 44). Our observation that SIVsm, SIVmac, and HIV-2 Nefs per-form similar functions supports the hypothesis that host factors are important for the different clinical outcomes of infection of mangabeys, macaques, and humans. We also found, however, that all HIV-2 310072nefalleles were prematurely truncated and that 310248 Nefs showed little functional activity. Unfor-tunately, these two HIV-2 strains were derived from a blood donor with unknown clinical status (42). Thus, it remains un-clear whether the lack of HIV-2 Nef function was associated with attenuated viral replication in vivo. Notably, a high prev-alence of defective nef genes in HIV-2 infection has been previously reported (61). A larger number of samples needs to be analyzed to draw definitive conclusions, but taken together these findings suggest that disrupted or nonfunctional nef
[image:9.585.70.518.71.377.2]genes contribute to the low level of viral replication and lack of disease progression in some HIV-2-infected individuals.
FIG. 6. The dileucine-based sorting motif in HIV-2 or SIVmac Nef is not required for down-modulation of CD3, MHC-I, and MHC-II surface expression. Cells were transfected with constructs expressing either the wild-type NA7, 239, BEN, or FFm1 Nefs or mutant forms containing changes of EXXXL to AXXXA in the C-proximal adaptor protein interaction site. Assays were performed as described in Materials and Methods and confirmed in two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
Recently, we performed a comprehensive analysis of HIV-1 and SIVcpz Nef function (32). A comparison of these previous results with those of the present study shows thatnef alleles derived from the HIV-1/SIVcpz and HIV-2/SIVsm/mac groups show some remarkable functional differences. In contrast to HIV-2 and SIVsm or SIVmac Nefs, HIV-1 Nefs are generally inactive in down-regulating cell surface expression of CD3, a component of the T-cell receptor. Furthermore, HIV-2 and SIVsm Nefs are much more active than HIV-1 and SIVcpz
[image:10.585.56.509.281.554.2]Nefs in down-regulating CD28 from the cell surface and in inhibiting TCR-mediated T-cell activation. Recent studies in-dicate that chronic immune activation might play a major role in AIDS pathogenesis (11, 54). HIV-2 is less pathogenic than HIV-1, the major causative agent of AIDS (37, 44). Concor-dant with our data, it has been demonstrated that the levels of immune activation are relatively low in HIV-2 infection (38). Microarray studies in transfected Jurkat T cells have shown that HIV-1 Nef triggers a transcriptional program resembling
FIG. 7. The C terminus of HIV-2 Nef is critical for MHC-I down-modulation but not for other Nef functions. (A) Mutations introduced into HIV-2 Nef. (B) The functional activity of the indicated Nef mutants was determined as described in the legend to Fig. 4. The NA7 AXXA Nef contains the changes P72A and P75A. The mutant SIVmac239 Nefs have been described previously (34, 58). Similar results were obtained in two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
TCR-mediated T-cell activation (56). It will be of interest to determine whether or not HIV-2 Nefs trigger similar T-cell signaling pathways. Analysis of Nef function clearly indicates that moderately pathogenic HIV-2 strains interfere with TCR signaling from the cell membrane more severely than highly virulent HIV-1 strains. Efficient inhibition of TCR signaling together with lower activity in triggering T-cell activation path-ways might explain why HIV-2 causes weaker chronic immune activation and is less pathogenic than HIV-1. However, the determinants of HIV and SIV pathogenesis are complex, and other viral and host factors are clearly also important. For example, SIVmac239 expressing anefallele that performs all of these functions is highly pathogenic in rhesus macaques (28, 29). It must be considered, however, that macaques are par-ticularly susceptible to SIV-induced disease. It is noteworthy
[image:11.585.119.468.68.446.2]that the accessory vpr and vpx genes are not required for efficient viral spread and disease progression in this model (19), although Vpr might play a relevant role for AIDS patho-genesis in HIV-1-infected humans (50). SIVmac239 is non-pathogenic when reintroduced into sooty mangabeys (27). Ma-caques infected with SIVsm showed high levels of immune activation and developed CD4⫹-T-cell depletion, whereas mangabeys displayed little T-cell proliferation and remained healthy (55). Thus, the seemingly high virulence of SIVmac239 is not an inherent property of the virus itself but due to the unusually high susceptibility of macaques to SIV-induced dis-ease. Mutations in Nef that increase its ability to cause T-cell activation enhance the virulence of SIVmac239 even further and result in an acute pathogenic phenotype (14). Most im-portantly, HIV-2, which is closely related to SIVmac and also
FIG. 8. SIVsm and HIV-2nefalleles enhance viral infectivity. (A) Generation of proviral SIVmac239 constructs carrying SIVsm and HIV-2nef
alleles. As described previously (31), the original SIVmac239nefinitiation codon and a second ATG at amino acid position 7 of thenefORF were mutated to eliminate the overlap between theenvandnefopen reading frames. The variousnefgenes were cloned into the unique SacI and MluI restriction sites. (B) Infectivity of SIVmac239 without overlappingenv-nefsequences. TZM-bl cells were infected in triplicate with the parental 239wt or 239nef* viruses or with mutant forms containing an intact (SSIV239nef⫹) or a prematurely truncated (SSIV239nef*)nefgene inserted downstream ofenvand upstream of a truncated U3 region as shown in panel A. (C) Infectivity of SIVmac239 variants containing the indicated HIV-1, SIVsm, or HIV-2nefalleles. TZM-bl indicator cells were infected in triplicate with 293T cell-derived virus stocks containing 200 pg p27. Infectivity is shown relative to the recombinant virus containing the 239wtnefallele. Similar results were obtained in two independent experiments and using P4-CCR5 indicator cells.
on November 8, 2019 by guest
http://jvi.asm.org/
originates from SIVsm-infected mangabeys (16), is clearly less virulent than HIV-1 in infected humans (37, 44). Thus, when compared in the same host, the SIVsm/mac/HIV-2 group is less pathogenic than the SIVcpz/HIV-1 group. Therefore, we feel that the observation that SIVmac239 is highly virulent in macaques does not contradict the possibility that down-mod-ulation of CD3 and CD28 favors slow/nonprogressive infec-tion.
Our results indicate that HIV-2 Nefs down-modulate CD3 and inhibit T-cell activation more effectively than SIVsmnef
alleles but are less active in stimulating viral replication in 221-89 cells and in human PBMC. However, more samples need to be investigated to clarify how frequently HIV-2 nef
genes are more active in inhibiting T-cell activation and less active in stimulating viral replication than those derived from other groups of primate lentiviruses. Furthermore, HIV-2 Nefs might be more active in stimulating viral replication in their usual genomic context. Our constructs have the advantage that
neffunction can be studied using the well-characterized pro-viral SIVmac239 clone without the complications of overlap-pingenvsequences. However, the increased genomic size and the expression of Nef from a different location resulted in reduced viral infectivity and replication compared to the pa-rental SIVmac239 clone. As discussed above, we feel that
fur-ther studies on their ability to stimulate T-cell activation and viral replication might provide important new insights into the pathogenesis of HIV-1 and HIV-2 infection.
In addition to the functional differences of HIV-1/SIVcpz and HIV-2/SIVsm/mac Nefs in modulating CD3 and CD28 surface expression, it is noteworthy that some conserved Nef functions are mediated by different Nef domains. The C-ter-minal region of SIVmac239, SIVsm, and HIV-2 Nefs is highly variable and not found in HIV-1 Nef (Fig. 2). Our mutational analysis extends previous studies (39, 58) showing that this region is exclusively involved in MHC-I down-modulation by the mac239 Nef and demonstrates that it plays a similar role for HIV-2 Nef function. It has recently been shown that changes selected for in rhesus macaques infected with an SIV-mac239 mutant containing deletions near the C terminus of Nef generate an additional PXXP motif, as present in most HIV-1 Nef sequences, and restore the ability to down-modu-late MHC-I (59). In comparison, introduction of an HIV-1 PXXP consensus did not restore MHC-I down-modulation by the truncated BEN Nef. Thus, at least in the context of the BEN Nef, this proline motif is not sufficient to functionally compensate for the C-terminal region. Notably, additional variations changing amino acids in 239 Nef to those usually present in HIV-1 Nef were also observed in infected macaques (59) and might be required to restore this function. In contrast to the effect on MHC-I, down-modulation of CD4 requires similar functional domains, such as the EXXXLL motif, in the SIVcpz/HIV-1 and SIVsm/HIV-2 Nef proteins. These findings suggest that the nefgene of the unknown ancestor of these groups of primate lentiviruses might have already been able to perform this function. Some conserved domains in Nef are clearly critical for specific functions. Even within the same group of viruses, however, the requirement of specific residues seems to be often dependent on the specificnefallele investi-gated. For example, the PA motif located in the N-proximal adaptor protein interaction site in SIVmac239 Nef is required for CD4 down-modulation (26). These residues are missing in the HIV-2 group B 310319 Nef (Fig. 2), although it effectively down-regulates CD4 (Fig. 4, column 9). Thus, while the func-tional analysis of specific Nef mutants provides some impor-tant information on the functional relevance of specific se-quence motifs, general conclusions must be made with great caution.
In contrast to the effect on CD4, down-regulation of MHC-I clearly requires distinct domains in HIV-1 and HIV-2 Nefs, suggesting that this function might have been acquired inde-pendently and later during the evolution of these primate len-tiviruses. The observation that distinct domains mediate dif-ferent Nef functions might be important for optimal viral spread in vivo, because it has been shown that various HIV-1 or SIVmac Nef activities can be modulated at different stages of disease progression (2, 7, 43). Our results also indicate that HIV-1 and HIV-2 Nefs use overlapping but distinct domains to up-regulate Ii. Mutations in the EXXXLL motif entirely dis-rupted HIV-1 Nef function but had less severe effects on SIV-mac239 or HIV-2 Nefs. We are currently investigating whether additional functional domains are involved in up-regulation of Ii by SIVmac and HIV-2 Nef proteins.
Taken together, the results of the present study demonstrate that the ability of Nef to modulate cell surface expression of
FIG. 9. Enhancement of viral replication by (A) SIVsm and (B) HIV-2nefalleles. Replication kinetics of recombinant SIVmac239 variants containing the indicated SIVsm or HIV-2nefalleles in 221-89 cells in the presence (left panels) or absence (right panels) of exoge-nous IL-2 are shown. Infections were performed using virus stocks containing 10 pg p27 antigen. All infections were performed at the same time and the 239wt and nef* controls are shown in both panels for comparison. Reverse transcriptase activity was determined using a phosphorimager. PSL, photon-stimulated light emission. Symbols:■, 239wt;䊐, nef*;F, K6nefalleles;⫻, uninfected cells. Similar results were obtained in two independent experiments.
on November 8, 2019 by guest
http://jvi.asm.org/
human CD4, CD3, CD28, MHC-I, MHC-II, and Ii and to enhance virion infectivity is conserved within the SIVsm/HIV-2/SIVmac group of primate lentiviruses. HIV-1 and SIVcpz Nefs are also able to perform a striking variety of functions. Nevertheless, compared to SIVsm and HIV-2 Nefs, they are generally unable to down-regulate CD3 and also have little effect on CD28 cell surface expression (32). It will be of inter-est to further evaluate whether these differences in Nef func-tion contribute to the low pathogenicity of HIV-2 compared to HIV-1 in the human host.
ACKNOWLEDGMENTS
We thank Thomas Mertens for constant support and encourage-ment, Ingrid Bennett for critical reading of the manuscript, and Ainee McKnight for providing the HIV-2 CBL23nefallele.
This work was supported by the Wilhelm-Sander Foundation, the Deutsche Forschungsgemeinschaft, and a grant from the Elitefo ¨rder-programm BW to J.M.
REFERENCES
1.Alexander, L., Z. Du, M. Rosenzweig, J. U. Jung, and R. C. Desrosiers.1997. A role for natural simian immunodeficiency virus and human immunodefi-ciency virus type 1 Nef alleles in lymphocyte activation. J. Virol.71:6094– 6099.
2.Arganaraz, E. R., M. Schindler, F. Kirchhoff, M. J. Cortes, and J. Lama. 2003. Enhanced CD4 down-modulation by late stage HIV-1 nef alleles is associated with increased Env incorporation and viral replication. J. Biol. Chem.278:33912–33919.
3.Bell, I., C. Ashman, J. Maughan, E. Hooker, F. Cook, and T. A. Reinhart. 1998. Association of simian immunodeficiency virus Nef with the T-cell receptor (TCR) zeta chain leads to TCR down-modulation. J. Gen. Virol. 79:2717–2727.
4.Benson, R. E., A. Sanfridson, J. S. Ottinger, C. Doyle, and B. R. Cullen.1993. Downregulation of cell-surface CD4 expression by simian immunodeficiency virus nef prevents viral super infection. J. Exp. Med.177:1561–1566. 5.Blagoveshchenskaya, A. D., L. Thomas, S. F. Feliciangeli, C. H. Hung, and
G. Thomas.2002. HIV-1 Nef downregulates MHC-I by a PACS-1- and PI3K-regulated ARF6 endocytic pathway. Cell111:853–866.
6.Bresnahan, P. A., W. Yonemoto, and W. C. Greene.1999. Cutting edge: SIV Nef protein utilizes both leucine- and tyrosine-based protein sorting path-ways for down-regulation of CD4. J. Immunol.163:2977–2981.
7.Carl, S., T. C. Greenough, M. Krumbiegel, M. Greenberg, J. Skowronski, J. L. Sullivan, and F. Kirchhoff.2001. Modulation of different human im-munodeficiency virus type 1 Nef functions during progression to AIDS. J. Virol.75:3657–3665.
8.Carl, S., A. J. Iafrate, S. M. Lang, N. Stolte, K. Matz-Rensing, D. Fuchs, C. Stahl-Hennig, J. Skowronski, and F. Kirchhoff.2000. Simian immunodefi-ciency virus containing mutations in N-terminal tyrosine residues and in the PxxP motif in Nef replicates efficiently in rhesus macaques. J. Virol.74:4155– 4164.
9.Clavel, F., M. Guyader, D. Gue´tard, M. Salle, L. Montagnier, and M. Alizon. 1986. Molecular cloning and polymorphism of the human immune deficiency virus type 2. Nature324:691–695.
10.Craig, H. M., M. W. Pandori, and J. C. Guatelli.1998. Interaction of HIV-1 Nef with the cellular dileucine-based sorting pathway is required for CD4 down-regulation and optimal viral infectivity. Proc. Natl. Acad. Sci. USA 95:11229–11234.
11.Dalgleish, A. G., and K. J. O’Byrne.2002. Chronic immune activation and inflammation in the pathogenesis of AIDS and cancer. Adv. Cancer Res. 84:231–276.
12.Derdeyn, C. A., J. M. Decker, J. N. Sfakianos, X. Wu, W. A. O’Brien, L. Ratner, J. C. Kappes, G. M. Shaw, and E. Hunter.2000. Sensitivity of human immunodeficiency virus type 1 to the fusion inhibitor T-20 is modulated by coreceptor specificity defined by the V3 loop of gp120. J. Virol.74:8358– 8367.
13.Desrosiers, R. C., and D. J. Ringler.1989. Use of SIV for AIDS research. Intervirology30:301–312.
14.Du, Z., S. M. Lang, V. G. Sasseville, A. A. Lackner, P. O. Ilyinskii, M. D. Daniel, J. U. Jung, and R. C. Desrosiers.1995. Identification of anefallele that causes lymphocyte activation and acute disease in macaque monkeys. Cell82:665–674.
15.Fultz, P. N.1993. Nonhuman primate models for AIDS. Clin. Infect. Dis. 17(Suppl. 1):S230–S235.
16.Gao, F., L. Yue, A. T. White, et al.1992. Human infection by genetically diverse SIVSM-related HIV-2 in west Africa. Nature358:495–499. 17.Gao, F., L. Yue, D. L. Robertson, S. C. Hill, H. Hui, R. J. Biggar, A. E.
Neequaye, T. M. Whelan, D. D. Ho, and G. M. Shaw.1994. Genetic diversity of human immunodeficiency virus type 2: evidence for distinct sequence subtypes with differences in virus biology. J. Virol.68:7433–7447. 18.Geyer, M., O. T. Fackler, and B. M. Peterlin.2001. Structure-function
rela-tionships in HIV-1 Nef. EMBO Rep.2:580–585.
19.Gibbs, J. S., A. A. Lackner, S. M. Lang, M. A. Simon, P. K. Sehgal, M. D. Daniel, and R. C. Desrosiers.1995. Progression to AIDS in the absence of a gene forvprorvpx. J. Virol.69:2378–2383.
20.Glushakova, S., J. Munch, S. Carl, T. C. Greenough, J. L. Sullivan, L. Margolis, and F. Kirchhoff.2001. CD4 down-modulation by human immu-nodeficiency virus type 1 Nef correlates with the efficiency of viral replication and with CD4⫹T-cell depletion in human lymphoid tissue ex vivo. J. Virol. 75:10113–10117.
21.Greenberg, M. E., A. J. Iafrate, and J. Skowronski.1998. The SH3 domain-binding surface and an acidic motif in HIV-1 Nef regulate trafficking of class I MHC complexes. EMBO J.17:2777–2789.
22.Hahn, B. H., G. M. Shaw, K. M. De Cock, and P. M. Sharp.2000. AIDS as a zoonosis: scientific and public health implications. Science287:607–614. 23.Howe, A. Y. M., J. U. Jung, and R. C. Desrosiers.1998. Zeta chain of the
T-cell receptor interacts with Nef of simian immunodeficiency virus and human immunodeficiency virus type 2. J. Virol.72:9827–9834.
24.Hua, J., and B. R. Cullen.1997. Human immunodeficiency virus types 1 and 2 and simian immunodeficiency virus Nef use distinct but overlapping target sites for downregulation of cell surface CD4. J. Virol.71:6742–6748. 25.Iafrate, A. J., S. Bronson, and J. Skowronski.1997. Separable functions of
Nef disrupt two aspects of T cell receptor machinery: CD4 expression and CD3 signaling. EMBO J.16:673–684.
26.Iafrate, A. J., S. Carl, S. Bronson, C. Stahl-Hennig, T. Swigut, J. Skowronski, and F. Kirchhoff.2000. Disrupting surfaces of Nef required for down-regu-lation of CD4 and for enhancement of virion infectivity attenuates simian immunodeficiency virus replication in vivo. J. Virol.74:9836–9844. 27.Kaur, A., R. M. Grant, R. E. Means, H. McClue, M. Feinberg, and R. P.
Johnson.1998. Diverse host responses and outcomes following SIVmac239 infection in sooty mangabeys and rhesus macaques. J. Virol.72:9597–9611. 28.Kestler, H., T. Kodama, D. Ringler, M. Marthas, N. Pedersen, A. Lackner, D. Regier, P. Sehgal, M. Daniel, N. King, and R. Desrosiers.1990. Induction of AIDS in rhesus monkeys by molecularly cloned simian immunodeficiency virus. Science248:1109–1112.
29.Kestler, H. W., D. J. Ringler, K. Mori, D. L. Panicali, P. K. Sehgal, M. D. Daniel, and R. C. Desrosiers.1991. Importance of thenefgene for mainte-nance of high virus loads and for development of AIDS. Cell65:651–662. 30.Kirchhoff, F., K. D. Jentsch, B. Bachmann, A. Stuke, C. Laloux, W. Luke, C.
Stahl-Hennig, J. Schneider, K. Nieselt, M. Eigen, and G. Hunsmann.1990. A novel proviral clone of HIV-2: biological and phylogenetic relationship to other primate immunodeficiency viruses. Virology177:305–311.
31.Kirchhoff, F., J. Munch, S. Carl, N. Stolte, K. Ma¨tz-Rensing, D. Fuchs, P. T. Haaft, J. L. Heeney, T. Swigut, J. Skowronski, and C. Stahl-Hennig.1999. The human immunodeficiency virus type 1nefgene can to a large extent replace simian immunodeficiency virusnefin vivo. J. Virol.73:8371–8383. 32.Kirchhoff, F., M. Schindler, N. Bailer, H. Renkema, K. Saksela, V. Knoop,
M. C. Mu¨ller-Trutwin, M. L. Santiago, F. Bibollet-Ruche, M. T. Dittmar, J. L. Heeney, B. H. Hahn, and J. Mu¨nch.2004. Nef proteins from simian immunodeficiency virus-infected chimpanzees interact with p21-activated kinase 2 and modulate cell surface expression of various human receptors. J. Virol.78:6864–6874.
33.Kumar, S., K. Tamura, and M. Nei.2004. MEGA3: integrated software for molecular evolutionary genetics analysis and sequence alignment. Brief. Bioinform.5:150–163.
34.Lang, S. M., A. J. Iafrate, C. Stahl-Hennig, E. M. Kuhn, T. Nilein, M. Haupt, G. Hunsmann, J. Skowronski, and F. Kirchhoff.1997. Association of simian immunodeficiency virus Nef with cellular serine/threonine kinases is dispensable for the development of AIDS in rhesus macaques. Nat. Med. 3:860–865.
35.Ling, B., C. Apetrei, I. Pandrea, R. S. Veazey, A. A. Lackner, B. Gormus, and P. A. Marx.2004. Classic AIDS in a sooty mangabey after an 18-year natural infection. J. Virol.78:8902–8908.
36.Lundquist, C. A., M. Tobiume, J. Zhou, D. Unutmaz, and C. Aiken.2002. Nef-mediated downregulation of CD4 enhances human immunodeficiency virus type 1 replication in primary T lymphocytes. J. Virol.76:4625–4633. 37.Marlink, R. G., D. Ricard, S. M. Boup, P. J. Kanki, J. L. Romet-Lemonne,
I. N. Doye, K. Diop, M. A. Simpson, F. Greco, M. J. Chou, V. Degruttola, C. C. Hsieh, C. Boye, F. Barin, F. Denis, M. F. McLane, and M. Essex.1988. Clinical, hematological, and immunologic cross-sectional evaluation of indi-viduals exposed to human immunodeficiency virus type-2 (HIV-2). AIDS Res. Hum. Retrovir.4:137–148.
38.Michel, P., A. T. Balde, C. Roussilhon, G. Aribot, J. L. Sarthou, and M. L. Gougeon.2000. Reduced immune activation and T cell apoptosis in human immunodeficiency virus type 2 compared with type 1: correlation of T cell apoptosis with beta2 microglobulin concentration and disease evolution. J. Infect. Dis.181:64–75.
39.Mu¨nch, J., N. Stolte, D. Fuchs, C. Stahl-Hennig, and F. Kirchhoff.2001. Efficient class I MHC down-regulation by simian immunodeficiency virus
on November 8, 2019 by guest
http://jvi.asm.org/
Nef is associated with a strong selective advantage in infected rhesus ma-caques. J. Virol.75:10532–10536.
40.Mu¨nch, J., A. Janardhan, N. Stolte, C. Stahl-Hennig, P. Ten Haaft, J. L. Heeney, T. Swigut, F. Kirchhoff, and J. Skowronski.2002. T-cell receptor: CD3 down-regulation is a selected in vivo function of simian immunodefi-ciency virus Nef but is not sufficient for effective viral replication in rhesus macaques. J. Virol.76:12360–12364.
41.Naidu, Y. M., H. W. Kestler III, Y. Li, C. V. Butler, D. P. Silva, D. K. Schmidt, C. D. Troup, P. K. Sehgal, P. Sonigo, M. D. Daniel, and R. C. Desrosiers.1988. Characterization of infectious molecular clones of simian immunodeficiency virus (SIVmac) and human immunodeficiency virus type 2: persistent infection of rhesus monkeys with molecularly cloned SIVmac. J. Virol.62:4691–4696.
42.Owen, S. M., D. Ellenberger, M. Rayfield, S. Wiktor, P. Michel, M. H. Grieco, F. Gao, B. H. Hahn, and R. B. Lal.1998. Genetically divergent strains of human immunodeficiency virus type 2 use multiple coreceptors for viral entry. J. Virol.72:5425–5432.
43.Patel, P. G., M. T. Yu Kimata, J. E. Biggins, J. M. Wilson, and J. T. Kimata. 2002. Highly pathogenic simian immunodeficiency virus mne variants that emerge during the course of infection evolve enhanced infectivity and the ability to downregulate CD4 but not class I major histocompatibility complex antigens. J. Virol.76:6425–6434.
44.Pepin, J., G. Morgan, D. Dunn, S. Gevao, M. Mendy, I. Gaye, N. Scollen, R. Tedder, and H. Whittle.1991. HIV-2-induced immunosuppression among asymptomatic West African prostitutes: evidence that HIV-2 is pathogenic, but less so than HIV-1. AIDS5:1165–1172.
45.Piguet, V., Y. L. Chen, A. Mangasarian, M. Foti, J. L. Carpentier, and D. Trono.1998. Mechanism of Nef-induced CD4 endocytosis: Nef connects CD4 with the mu chain of adaptor complexes. EMBO J.17:2472–2481. 46.Platt, E. J., K. Wehrly, S. E. Kuhmann, B. Chesebro, and D. Kabat.1998.
Effects of CCR5 and CD4 cell surface concentrations on infection by mac-rophagetropic isolates of human immunodeficiency virus type 1. J. Virol. 72:2855–2864.
47.Regier, D. A., and R. C. Desrosiers.1990. The complete nucleotide sequence of a pathogenic molecular clone of SIV. AIDS Res. Hum. Retrovir.6:1221– 1231.
48.Rey-Cuille, M. A., J. L. Berthier, M. C. Bomsel-Demontoy, Y. Chaduc, L. Montagnier, A. G. Hovanessian, and L. A. Chakrabarti.1998. Simian im-munodeficiency virus replicates to high levels in sooty mangabeys without inducing disease. J. Virol.72:3872–3886.
49.Riggs, N. L., H. M. Craig, M. W. Pandori, and J. C. Guatelli.1999. The dileucine-based sorting motif in HIV-1 Nef is not required for down-regu-lation of class I MHC. Virology258:203–207.
50.Ru¨cker, E., J.-C. Grivel, J. Mu¨nch, F. Kirchhoff, and L. Margolis.2004. Vpr and Vpu are important for efficient human immunodeficiency virus type 1
replication and CD4⫹T-cell depletion in human lymphoid tissue ex vivo. J. Virol.78:12689–12693.
51.Schindler, M., S. Wu¨rfl, P. Benaroch, T. C. Greenough, R. Daniels, P. Easterbrook, M. Brenner, J. Mu¨nch, and F. Kirchhoff.2003. Down-modu-lation of mature major histocompatibility complex class II and up-reguDown-modu-lation of invariant chain cell surface expression are well-conserved functions of human and simian immunodeficiency virusnefalleles. J. Virol.77:10548– 10556.
52.Schindler, M., J. Mu¨nch, C. Stahl-Hennig, J. Skowronski, and F. Kirchhoff. 2004. Comprehensive analysis of Nef functions selected in simian immuno-deficiency virus-infected macaques. J. Virol.78:10588–10597.
53.Schulz, T. F., D. Whitby, J. G. Hoad, T. Corrah, H. Whittle, and R. A. Weiss. 1990. Biological and molecular variability of human immunodeficiency virus type 2 isolates from The Gambia. J. Virol.64:5177–5182.
54.Silvestri, G., D. L. Sodora, R. A. Koup, M. Paiardini, S. P. O’Neil, H. M. McClure, S. I. Staprans, and M. B. Feinberg.2003. Nonpathogenic SIV infection of sooty mangabeys is characterized by limited bystander immuno-pathology despite chronic high-level viremia. Immunity18:441–452. 55.Silvestri, G., A. Fedanov, S. Germon, N. Kozyr, W. J. Kaiser, D. A. Garber,
H. McClure, M. B. Feinberg, and S. I. Staprans. 2005. Divergent host responses during primary simian immunodeficiency virus SIVsm infection of natural sooty mangabey and nonnatural rhesus macaque hosts. J. Virol. 79:4043–4054.
56.Simmons, A., V. Aluvihare, and A. McMichael.2001. Nef triggers a tran-scriptional program in T cells imitating single-signal T cell activation and inducing HIV virulence mediators. Immunity14:763–777.
57.Stumptner-Cuvelette, P., S. Morchoisne, M. Dugast, S. Le Gall, G. Raposo, O. Schwartz, and P. Benaroch.2001. HIV-1 Nef impairs MHC class II antigen presentation and surface expression. Proc. Natl. Acad. Sci. USA 98:12144–12149.
58.Swigut, T., A. J. Iafrate, J. Mu¨nch, F. Kirchhoff, and J. Skowronski.2000. Simian and human immunodeficiency virus Nef proteins use different sur-faces to down-regulate class I major histocompatibility antigen expression. J. Virol.74:5691–5701.
59.Swigut, T., L. Alexander, J. Morgan, J. Lifson, K. G. Mansfield, S. Lang, R. P. Johnson, J. Skowronski, and R. C. Desrosiers.2004. Impact of Nef-mediated downregulation of major histocompatibility complex class I on immune response to simian immunodeficiency virus. J. Virol.78:13335– 13344.
60.Swigut, T., N. Shody, and J. Skowronski.2001. Mechanism for down-regu-lation of CD28 by Nef. EMBO J.20:1593–1604.
61.Switzer, W. M., S. Wiktor, V. Soriano, A. Silva-Graca, K. Mansinho, I. M. Coulibaly, E. Ekpini, A. E. Greenberg, T. M. Folks, and W. Heneine.1998. Evidence of Nef truncation in human immunodeficiency virus type 2 infec-tion. J. Infect. Dis.177:65–71.