Specific Recognition of 5
=
GG Sequence for RTPase Activity
Liya Hu,aDar-Chone Chow,bJohn T. Patton,cTimothy Palzkill,b,dMary K. Estes,dand B. V. Venkataram Prasada,d
Verna and Marrs McLean Department of Biochemistry and Molecular Biology,aDepartment of Pharmacology,band Department of Molecular Virology and Microbiology,d
Baylor College of Medicine, Houston, Texas, USA, and National Institute of Allergy and Infectious Diseases, National Institutes of Health, Bethesda, Maryland, USAc
Rotavirus nonstructural protein NSP2, a functional octamer, is critical for the formation of viroplasms, which are exclusive sites for replication and packaging of the segmented double-stranded RNA (dsRNA) rotavirus genome. As a component of replication intermediates, NSP2 is also implicated in various replication-related activities. In addition to sequence-independent single-stranded RNA-binding and helix-destabilizing activities, NSP2 exhibits monomer-associated nucleoside and 5=RNA triphospha-tase (NTPase/RTPase) activities that are mediated by a conserved H225 residue within a narrow enzymatic cleft. Lack of a 5=
␥-phosphate is a common feature of the negative-strand RNA [(ⴚ)RNA] of the packaged dsRNA segments in rotavirus. Strik-ingly, all (ⴚ)RNAs (of group A rotaviruses) have a 5=GG dinucleotide sequence. As the only rotavirus protein with 5=RTPase activity, NSP2 is implicated in the removal of the␥-phosphate from the rotavirus (ⴚ)RNA. To understand how NSP2, despite its sequence-independent RNA-binding property, recognizes (ⴚ)RNA to hydrolyze the␥-phosphate within the catalytic cleft, we determined a crystal structure of NSP2 in complex with the 5=consensus sequence of minus-strand rotavirus RNA. Our studies show that the 5=GG of the bound oligoribonucleotide interacts extensively with highly conserved residues in the NSP2 enzy-matic cleft. Although these residues provide GG-specific interactions, surface plasmon resonance studies suggest that the C-ter-minal helix and other basic residues outside the enzymatic cleft account for sequence-independent RNA binding of NSP2. A novel observation from our studies, which may have implications in viroplasm formation, is that the C-terminal helix of NSP2 exhibits two distinct conformations and engages in domain-swapping interactions, which result in the formation of NSP2 octa-mer chains.
R
otavirus, a member of theReoviridae family, causes severe life-threatening infantile gastroenteritis worldwide (7). It is a large nonenveloped icosahedral virus with three concentric capsid layers that enclose a genome consisting of 11 segments of double-stranded RNA (dsRNA) (16,31). The rotavirus genome encodes six structural (VPs) and six nonstructural (NSPs) proteins (7). Transcription of the genome segments takes place within the in-tact double-layered particles (DLPs), which are produced by the removal of the outer capsid layer during cell entry (7). This pro-cess, which yields positive-strand RNAs [(⫹)RNA] that are capped at the 5=end, is carried out by the viral polymerase (VP1) and the capping enzyme (VP3) located inside the DLPs. Subse-quent processes, including synthesis of dsRNA segments from (⫹)RNAs by VP1 and their packaging into newly assembled DLPs, take place in specialized compartments called viroplasms (7, 23). During genome replication, a consensus sequence [3=CS(⫹)], 5=-UGUGACC-3=at the 3=end of the (⫹)RNA, con-served among group A rotaviruses, is specifically recognized by VP1 to produce minus-strand RNA [(⫺)RNA] with a comple-mentary CS [5=CS(⫺)] that terminates with a highly conserved GG motif at the 5=end (18,35,39,40) (Fig. 1). A strikingly con-sistent feature of the rotavirus (⫺)RNAs is the absence of␥ -phos-phate at their 5=ends (9,20,37). This highly conserved feature suggests that removal of the␥-phosphate has an obligatory func-tion during virus replicafunc-tion. Among the rotavirus proteins, only NSP2 exhibits RNA 5=triphosphatase (RTPase) activity (37).NSP2 is a multifunctional enzyme that is implicated in various replication-related activities. Along with NSP5, it is essential for the formation of viroplasms, sites of rotavirus genome replication and DLP assembly. As one of the components of early replication intermediates, NSP2 is suggested to directly interact with VP1 and
partially replicated viral RNA (2,3,12,13). Biochemical studies with recombinant NSP2 show that it possesses several properties that are consistent with its intricate role in genome replication/ packaging. NSP2 binds to single-stranded-RNA (ssRNA) without any observed sequence specificity and has the ability to destabilize duplex RNA (32,34). In addition to the 5=RTPase activity men-tioned above, NSP2 exhibits nucleotidyl triphosphatase (NTPase) (32) and nucleoside diphosphate kinase (NDP kinase) activities (14), all of which are mediated by a highly conserved H225 as the catalytic residue. The RNA-binding and helix-destabilizing ities of NSP2, neither of which depends upon its enzymatic activ-ities, are thought to be used to relax secondary structures of mRNA templates for dsRNA synthesis by VP1 (34). Precisely how the enzymatic activities of NSP2 are utilized during genome rep-lication/packaging needs to be elucidated, and yet their necessity is strongly supported by complementation assays with enzymati-cally defective NSP2, which show the abrogation of dsRNA syn-thesis without compromising the functional oligomeric state of NSP2 or viroplasm formation (33).
The structure of NSP2, which readily self-assembles into an octamer, has been characterized by both X-ray crystallography and electron cryomicroscopy (cryo-EM) techniques (10, 11). NSP2 subunits, related by 4-2-2 symmetry, form a donut-shaped
Received15 May 2012Accepted10 July 2012
Published ahead of print18 July 2012
Address correspondence to B. V. Venkataram Prasad, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/JVI.01201-12
on November 7, 2019 by guest
http://jvi.asm.org/
octamer with a 35 Å wide central hole and four exterior grooves that are lined predominantly by basic residues. The NSP2 mono-mer has distinct N- and C-terminal domains separated by a deep cleft. Crystallographic studies of NSP2 with ATP analogs show that nucleoside substrates bind within the cleft, and that␥ -phos-phate hydrolysis, involving a phosphohistidine intermediate, is mediated by H225 (14). However, how NSP2 recognizes 5=RNA, which is shown to be a better substrate than NTPs for␥-phosphate hydrolysis (37), how the narrow catalytic cleft accommodates the 5=nucleotides of RNA, and how, in the context of rotavirus repli-cation, NSP2 makes sequence-specific interactions with con-served 5=GG dinucleotide remain unclear.
In this study, to understand the structural basis for how NSP2 recognizes rotavirus (⫺)RNA as a substrate for RTPase activity, we carried out crystallographic studies of NSP2 with RNA oligo-nucleotides, including one that corresponds to 5=CS(⫺). Our re-sults show that the 5=CS(⫺) oligoribonucleotide fully engages the NSP2 catalytic cleft through extensive contacts between 5=GG and highly conserved amino acid residues in the cleft. A novel obser-vation from our crystallographic studies, which may be relevant in the context of viroplasm formation, is that the C-terminal helix of NSP2 adopts two different conformations and engages in a do-main-swapping interaction to link neighboring NSP2 octamers. Surface plasmon resonance (SPR) experiments show that deletion of this basic C-terminal helix significantly reduces NSP2 affinity for RNA, which suggests that while residues in the cleft confer specificity for the GG dinucleotide, the C-terminal helix and other basic residues outside the cleft in the NSP2 octamer are involved in sequence-independent RNA binding by NSP2 and play a role in its helix-destabilizing activity.
MATERIALS AND METHODS
Protein expression, purification, and crystallization.The NSP2 gene from simian rotavirus strain SA11 was cloned into the bacterial expression vector pQE60 (Qiagen) with a C-terminal His tag following a thrombin cleavage site and expressed inEscherichia colistrain M15(pREP4). This His-tagged protein was purified using Ni-nitrilotriacetic acid (Ni-NTA) affinity chromatography (Qiagen). After this step, the His tag was
re-moved using thrombin (Hematologic Tech). Untagged NSP2 was further purified over Ni-NTA affinity and HiTrap SP HP cation-exchange chro-matography columns (GE Healthcare), dialyzed into 10 mM Tris-HCl (pH 7.2)–50 mM NaCl–1 mM dithiothreitol, and concentrated to 20 to 30 mg/ml.
Mutagenesis of NSP2. The NSP2 C-terminal deletion mutant (NSP2⌬CT) was produced by PCR amplification. An expression vector of pQEg8/KR⌬T encoding the C-terminal deletion and the K223A and R227A mutations was produced by PCR amplification using pQE60g8/⌬T as the template and the primers 5=-TAACAAGTTTGCTGTAATCACACATGGT
GCGGGTCATTATGCAATTGTAAAGTATTCATCAGTTGCTAAT-3=
and 5=-ATTAGCAACTGATGAATACTTTACAATTGCATAATGACCC GCACCATGTGTGATTACAGCAAACTTGTTA-3=. The bases in these primers used to change the codons in NSP2 gene are denoted in boldface. Crystallization.SA11 NSP2/RNA (5=-P-GGU CAC AU, octa-riboad-enyl nucleotide, purchased from Dharmacon RNAi Technologies was co-crystallized by hanging-drop vapor diffusion. Drops were composed 2l of protein-RNA mixed with 2l of well solution (50 mM sodium caco-dylate [pH 6.5], 70 mM NaCl, 1 mM EDTA, 10 to 12% PEG 4000). Crys-tals were transferred in a single step to the mother liquid supplemented with 20% glycerol for 1 min and then flash-frozen in liquid nitrogen for diffraction data collection.
Structure determination and refinement. Data were collected at 100K at beamline 19-ID or 19-BM at the Advanced Photon Source and processed using either using HKL2000 (22), d*trek (25), or IMOSFLM (41). The structures of NSP2-RNA complexes were determined by molec-ular replacement (MR) method using NSP2 structure (Protein Data Bank [PDB] no. 1L9V) as a search model. The MR solution was obtained using the program PHASER (19). Model building was performed using COOT (6). Iterative rounds of refinement, which included simulated annealing, translation/liberation/screw (TLS), and noncrystallographic symmetry constrains, and model building were carried using PHENIX (1). In the NSP2-5=CS(⫺) complex, the ligand density was validated using compos-ite and simulated annealing omit maps using either the PHENIX (1) or the CNS (4) packages. The final model of this complex has anRwork⫽20.63% and anRfree⫽17.21% with 97.7% of all residues within the most favored or allowed regions in the Ramachandran plot. The data collection and refinement statistics are summarized inTable 1. Ligand interactions were analyzed using COOT (6) and LIGPLOT (38) with donor to acceptor distances between 2.6 and 3.2 Å for hydrogen bonding interactions and C-C distances between 3.4 and 4.5 Å for hydrophobic interactions. The
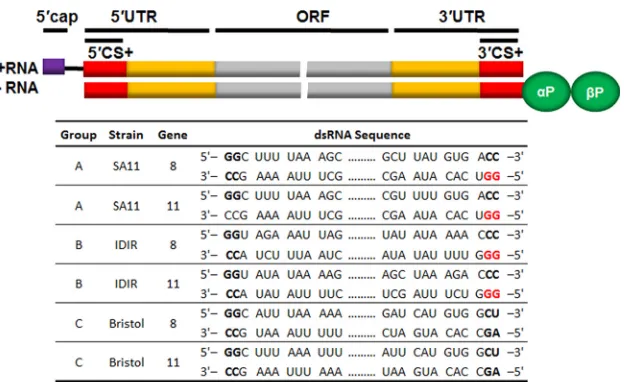
FIG 1Diagram schematic of a rotavirus dsRNA. The top schematic represents a dsRNA with the open reading frame (ORF) shown in gray, the 5=and 3=
untranslated regions (UTRs) shown in orange, and the consensus sequences (CS) shown in red. The lower table shows the representative sequences of the 5=and 3=CS from different groups, strains, and gene segments of rotavirus. The conserved “GG” dinucleotide is shown in red.
Hu et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:2.585.138.450.63.254.2]electrostatic potential surface of the NSP2 octamer was calculated by using PyMol (29). Figures were generated using Chimera (24).
NSP2⌬CTalso crystallized under the same conditions as the NSP2-5=CS(⫺) complex. Diffraction data for these crystals were collected at the Baylor College of Medicine crystallography core facility using Rigaku FR-E⫹SuperBright rotating anode. NSP2⌬CTcrystals diffracted to⬃3.4 Å, and data processing, using d*TREK (25), indicated a P4 space group. Further data analysis using SCALA in CCP4 (41) and X-triage in PHENIX (1) indicated anisotropic diffraction and twinning. MR using PHASER (19), with previously determined NSP2 monomeric subunit without the residues 294 to 317 as a search model, provided a unique solution with a high z-score. The crystallographic asymmetric unit consists of five tail-to-tail interacting dimers. Four of these constitute one octamer, and the remaining dimer in association with three other dimers related by crys-tallographic 4-fold symmetry forms another octamer. The structure re-finement was carried out with appropriate twinning corrections using REFMAC5 (41). The data collection and refinement statistics are summa-rized inTable 1.
SPR analysis.Biacore 3000 was used to analyze the binding kinetics of NSP2 to RNA oligonucleotides. In these studies, 5=phosphorylated and 3=
biotinylated oligonucleotides—5=-P-GGU CAC AUA A-biotin-3=(GG nucleotide) and 5=-P-CCU CAC AUA A-biotin-3= (CCnucleotide)— purchased from IDT DNA Technologies were immobilized on an SA sen-sor chip with an analyteRmaxin the 100- to 200-resonance unit (RU) range. Binding experiments were performed with HBS-P buffer (10 mM HEPES [pH 7.4], 150 mM NaCl, 5 mM MgCl2, 0.05% surfactant P20). Ten concentrations (5 to 500 nM) of NSP2 were injected at 20°C with a flow rate of 15l/min. The kinetic parameters and the dissociation con-stants (Kd) were calculated using BIAevaluation 4.1.1(Biacore Life Sci-ences). Initial fitting of the sensograms of wild-type NSP2 using a 1:1
Langmuir binding model produced high2values and subsequently were fit using a heterogeneous ligand parallel reactions (HLPR; two-site bind-ing) model. Sensograms of the NSP2⌬CTand NSP2⌬CTK223A/R227A mutant were fit to a 1:1 Langmuir binding model.
Protein structure accession numbers.Coordinates and structure fac-tors of the structures discussed here have been deposited in the PDB under accession numbers 4G0A (NSP2-RNA complex) and 4G0J (NSP2⌬CT).
RESULTS
Untagged NSP2 construct and cocrystallization with oligoribo-nucleotides.All previously reported structural studies were car-ried out with a C-terminal His-tagged NSP2 (10,11,14). To avoid any possible interference of the basic His tag with negatively charged RNA, a new construct was designed such that the His tag could be removed (see Materials and Methods). With this untagged NSP2, crystallization trials in the presence of oligo-nucleotides, 5=-GGUCACAU-3=, which corresponds to the 5=CS of rotavirus (⫺)RNAs [5=CS(⫺)] and other oligoriboadenyl-nucleotides were carried out using Natrix, a commercially avail-able screen for nucleic acids and protein-nucleic acid complexes. Structures were determined by molecular replacement (MR) us-ing the previously determined NSP2 monomer structure (PDB no. 1L9V) as the phasing model. Although we were able to obtain crystals of NSP2 in the presence of each of the RNA nucleotides mentioned above, and these crystals diffracted to resolutions ranging from 2.2 to 3.2 Å, only cocrystals of NSP2-5=CS(⫺), dif-fracting to 2.1-Å resolution, showed strong interpretable density for the ligand. Despite several attempts, including various buffer conditions, pH, ionic strength, and ligand concentrations, no in-terpretable density for the ligand was observed with other oligori-bonucleotides. The data collection and refinement statistics for NSP2 in complex with the 5=CS(⫺) is presented inTable 1.
Structure of NSP2-RNA complex.In contrast to His-tagged NSP2, which crystallized in the I422 space group with one mole-cule in the asymmetric unit (10,14), analysis of the diffraction data from the cocrystal of untagged NSP2-5=CS(⫺) indicated a different space group, P3121, with four NSP2 molecules in the
crystallographic asymmetric unit. In all the four subunits, both 2Fo-Fc and Fo-Fc maps clearly indicated extra density that was unaccounted for by NSP2 but could be interpreted as the terminal GGU of the 5=CS RNA. The modeled nucleotide was further val-idated by computing composite and simulated annealing omit maps (Fig. 2A). All four subunits in the crystallographic asymmet-ric unit show the same structural features with two distinct do-mains separated by a cleft. A striking difference, however, is that in one of the subunits, the C-terminal helix is moved away from the C-terminal domain (henceforth referred to as open conforma-tion), and in the other three subunits it remains closer to this domain (closed conformation) as in the His-tagged apo-NSP2 structure (Fig. 2BandC). The GG of the 5=CS (⫺) RNA binds deep within the cleft. Although partial density for the next nucle-otide (U) could be observed at lower contour levels, there is no interpretable density for the remaining five bases as they emerge from the cleft, indicating that they are not constrained by any specific interactions with NSP2.
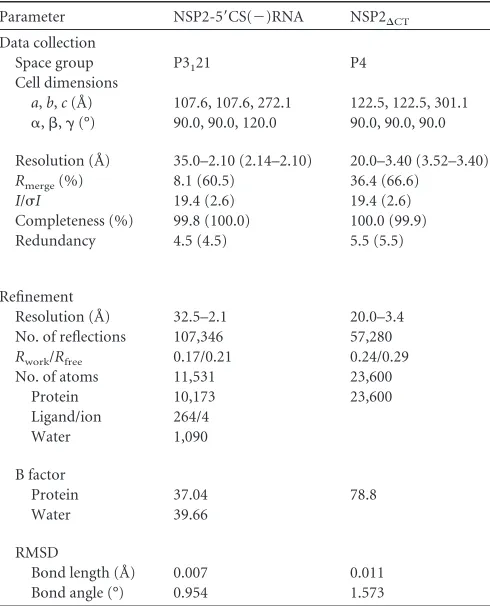
Octamer formation. NSP2 subunits in the NSP2-5=CS(⫺) complex structure show octamer formation that is consistent with the previous structural and biochemical studies of NSP2, which indicate that NSP2 readily forms an octamer and this quaternary organization is the functional form (30,32). In the previous crys-TABLE 1Diffraction data collection and refinement statistics of NSP2/
RNA complex and NSP2⌬CTa
Parameter NSP2-5=CS(⫺)RNA NSP2⌬CT
Data collection
Space group P3121 P4
Cell dimensions
a,b,c(Å) 107.6, 107.6, 272.1 122.5, 122.5, 301.1
␣,,␥(°) 90.0, 90.0, 120.0 90.0, 90.0, 90.0
Resolution (Å) 35.0–2.10 (2.14–2.10) 20.0–3.40 (3.52–3.40)
Rmerge(%) 8.1 (60.5) 36.4 (66.6)
I/I 19.4 (2.6) 19.4 (2.6)
Completeness (%) 99.8 (100.0) 100.0 (99.9)
Redundancy 4.5 (4.5) 5.5 (5.5)
Refinement
Resolution (Å) 32.5–2.1 20.0–3.4
No. of reflections 107,346 57,280
Rwork/Rfree 0.17/0.21 0.24/0.29
No. of atoms 11,531 23,600
Protein 10,173 23,600
Ligand/ion 264/4
Water 1,090
B factor
Protein 37.04 78.8
Water 39.66
RMSD
Bond length (Å) 0.007 0.011
Bond angle (°) 0.954 1.573
aValues in parentheses are for the highest-resolution shell.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:3.585.41.286.86.389.2]tallographic studies of the His-tagged NSP2 in the I422 space group, application of the crystallographic 4-2-2 symmetry leads to octamer formation consisting of two tail-to-tail interacting te-tramers. In the present P3121 space group, the four NSP2
mole-cules in the asymmetric unit, which are related by a noncrystallo-graphic 4-fold symmetry, form a tetramer with extensive interactions between the subunits, and two of these tetramers, related by the crystallographic 2-fold axis, form the typical NSP2 octamer (Fig. 3AandB). Unlike the I422 octamer, in which the
subunits of each tetramer are equivalent because of the crystallo-graphic 4-fold axis, in the P3121 tetramer, conformational
changes in the C-terminal helix break this equivalence. Except for this difference, all of the other structural features in the P3121
octamer, including the 35-Å central hole (Fig. 3B) along the 4-fold axis and the basic grooves at the 2-fold axes (Fig. 3CandD), are the same as in the I422 octamer.
NSP2-RNA interactions.The binding of 5=CS(⫺) by NSP2 involves a network of hydrogen bond interactions, several hydro-FIG 2Structure of NSP2-5=CS(⫺)RNA complex. (A) Simulated annealing composite omit map showing the 5=terminal nucleotides in a (2ⱍFobservedⱍ ⫺ ⱍFcalculatedⱍ) map calculated by using the protein only as the phasing model and contoured at⬃1.1. Modeled RNA is shown in a ball-and-stick representation
(yellow) inside the map, with the nitrogen and the oxygen atoms colored in blue and red, respectively. (B) Ribbon diagram representation of an NSP2 monomer in the “closed” conformation. (C) Ribbon diagram representation of an NSP2 monomer in the “open” conformation.
FIG 3NSP2 octamer formation. (A) NSP2 tetramer in the crystallographic asymmetric unit as viewed along the pseudo 4-fold axis of the tetramer with bound RNA shown in a sphere representation. (B) NSP2 octamer formed by the tetramers related by the crystallographic 2-fold axis, as viewed along the pseudo 4-fold axis of the octamer. (C) NSP2 octamer viewed along the 2-fold axes showing the RNA-binding cleft. (D) Electrostatic surface potential surface of the NSP2 octamer (calculated without bound nucleotides) showing the groove (dashed black line) and two distinct electropositive patches: one near the cleft (white star) and the other at either end of the groove corresponding to the locations of the C-terminal helices in open (dashed black arrow) and closed (black arrow) conformations. Positively and negatively charged residues are shown in blue and red, respectively.
Hu et al.
on November 7, 2019 by guest
http://jvi.asm.org/
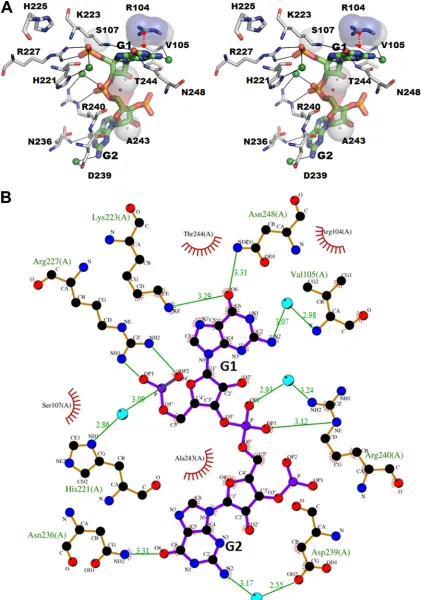
[image:4.585.113.474.65.211.2] [image:4.585.99.488.394.665.2]phobic contacts, and a distinct cation-interaction between the 5=GG dinucleotide and the amino acid residues within the NSP2 catalytic cleft (Fig. 4). The side chains of K223 and N248 form hydrogen bonds with the base of G1, and R227 makes bidentate hydrogen bonds with two of the phosphate oxygens of G1. In addition, the G1 residue of RNA is further stabilized by a cation- interaction between the side chain of Arg104 and the pyrimidine ring of G1. Consistent with the suggestion that NSP2 uses the same catalytic mechanism for both RTPase and NTPase activities (37), the terminal G1 nucleotide of the 5=CS(⫺) binds within the cata-lytic cleft at the same location as nonhydrolyzable ATP analogs in previous cocrystal structures (14).
The G2 of 5=CS(⫺) also makes a significant number of contacts with the NSP2 residues in the cleft. The phosphate oxygen of G2 participates in direct hydrogen bonding interaction with the side chain of R240, and its base with the side chain of N236. In addition to these electrostatic interactions, the sugars and bases of GG con-tribute to hydrophobic interactions that involves NSP2 residues V105, S107, D239, A243, and T244 (Fig. 4AandB). The confor-mation of the bound GG and its interactions with NSP2 are almost identical between the NSP2 subunits in the tetramer irrespective of the conformational state of the C-terminal helix. Compared to the previously published apo-NSP2 structure, the binding of 5=CS(⫺) does not induce any significant changes in NSP2, except for the slight movement toward the bound RNA of an␣-helix (amino acids [aa] 235 to 243) and a loop (aa 251 to 256) on one side of the basic cleft.
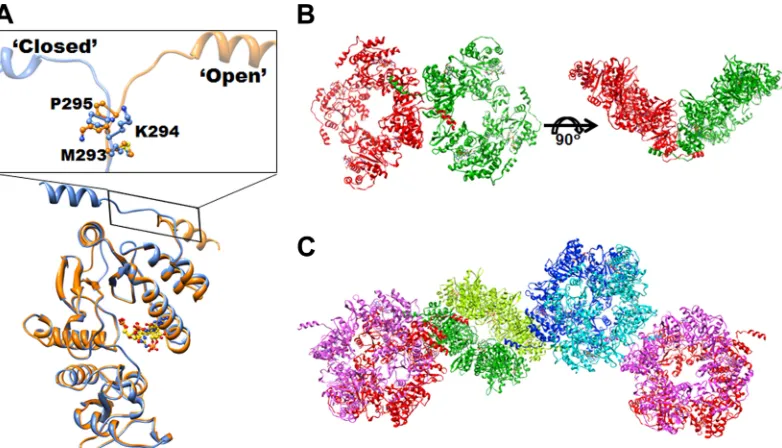
The C-terminal helix participates in domaswapping in-teractions.As mentioned above, the C-terminal helix of NSP2 adopts distinct “closed” and “open” conformations. The C-termi-nal helix in the open conformation engages in interoctamer inter-actions, akin to domain swapping. The pocket vacated by the C-terminal helix in one octamer is occupied by an open-confor-mation C-terminal helix from the adjacent symmetry-related oc-tamer such that the exchanged C-terminal helix mimics a C-ter-minal helix in the closed conformation. With each octamer consisting of two C-terminal helices on either side in the open conformation, the domain-swapping interactions leads to the for-mation of a chain of NSP2 octamers in the crystal lattice (Fig. 5C). Changes in the backbone dihedral angles of M293, K294, and P295 lead to the C-terminal helix switching its conformation (Fig. 5A andB). The domain-swapping interactions bring the neighboring octamers in the chain to closely interact with one another through several strong hydrophobic contacts involving residues L191, P196, and I146 with a buried surface area of⬃1,300 Å2between
them. Such a domaswapping interaction, leading to strong in-teroctamer association, is likely an important factor in the context of viroplasm formation.
SPR analysis of NSP2-RNA binding.To further understand the RNA-binding characteristics of NSP2, particularly in regard to RNA nucleotides that lack the 5=GG, and to examine whether the C-terminal helix, which contains several basic residues, plays any role in RNA binding, we carried out binding experiments using SPR. Previous studies showed that mutating some of the basic residues that line the groove, particularly residues in the N-termi-nal domain, diminishes RNA binding (37). Electrostatic potential surface of the NSP2 octamer shows that in addition to a strong electropositive patch close to the cleft, the C-terminal helices of the 2-fold related NSP2 subunits form another strong positively charged patch at either end of the groove (Fig. 2D). We
hypothe-sized that the basic C-terminal helix of NSP2, in addition to the basic residues in the N-terminal domain, is important for se-quence-independent RNA binding of NSP2.
For our SPR experiments, we cloned, expressed, and purified a deletion construct, NSP2⌬CT, which lacks residues 295 to 317.
Size-exclusion chromatography of NSP2⌬CTshowed a very similar
elution profile to that of wild-type NSP21-317aa, indicating that the
C-terminal truncation did not affect the formation of NSP2 oc-tamers (Fig. 6AandB). Although we were able to obtain diffract-ing crystals of this deletion construct, crystals diffracted to a lower resolution (⬃3.4 Å) and showed anisotropic diffraction and twin-ning that resulted in relatively highRmergevalues (Table 1).
De-spite these challenges, phasing by molecular replacement was suc-cessful. The structure refined withRworkandRfreevalues of 26 and
30%, respectively, and showed that NSP2⌬CT forms native-like
octamers (Fig. 6C) consistent with the results from size exclusion chromatography.
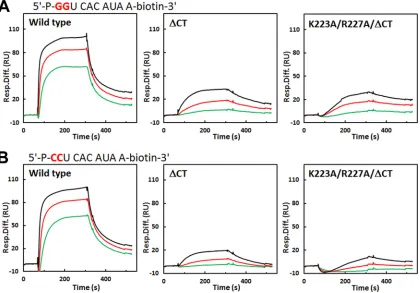
For RNA-binding analysis by SPR, we used a biotinylated de-caribonucleotide with a sequence corresponding to the 5=CS(⫺) and another decaribonucleotide in which the 5=GG was replaced with CC. The sensogram of RNA binding with wild-type NSP2 (wtNSP2) was best fitted using an HLPR (two-site) binding model (Fig. 7). The2value of fitting using HLPR was consistently and
significantly lower than using 1:1 Langmuir model (one-site) for all of the concentrations of NSP2. This data analysis shows that although the binding affinities of both GG and CC nucleotides are similar, the GG nucleotide has a slightly higher binding affinity compared to the CC nucleotide and it exhibits significantly (⬃ 2-fold) higher association for one of the sites (“site 1”) (Fig. 7and Table 2). In light of our crystallographic analysis with the GG nucleotide and the previous cryo-EM studies of NSP2 with decari-boadenylnucleotide showing an accumulation of extra density across the groove regions (including the C-terminal helix) (11), one qualitative interpretation is that while both GG and CC can bind to the electropositive grooves (including the C-terminal he-lix) in a sequence-independent manner (groove binding), the GG nucleotide additionally binds cleft region through sequence-spe-cific interactions. Such a preferential binding of the GG nucleotide within the cleft may contribute to its higher binding affinity. Al-though the CC nucleotide can potentially bind NSP2 interacting with some basic residues inside the cleft, this interaction would lack the hydrogen bonding with N236 and N248, cation- inter-action, and some of the stabilizing hydrophobic contacts found in the NSP2-GG interaction.
With NSP2⌬CT, the sensogram fits well to a 1:1 Langmuir
model, with low2values. The binding parameters clearly show that deletion of the C-terminal helix has a profound effect on RNA binding (Fig. 7,Table 2). In the absence of the C-terminal helix, the binding affinity for both GG and CC nucleotides is signifi-cantly reduced. Of particular note, the rate of association (kon) is
considerably reduced with NSP2⌬CTversus wtNSP2, suggesting
that the basic C-terminal helix also has an electrostatic steering effect on RNA binding to NSP2. After deletion of the C-terminal helix, the difference between GG and CC binding to NSP2 be-comes more pronounced with the apparent binding affinity of the GG nucleotide significantly higher than of the CC nucleotide. Since the binding is isolated to the cleft and other nearby basic residues in the groove, preferential binding of the GG nucleotide within the cleft is clearly accentuated. This is further substantiated by a proportional reduction in the binding affinities of the GG and
on November 7, 2019 by guest
http://jvi.asm.org/
FIG 4(A) Stereo view of interactions between NSP2 and 5=CS of (⫺)RNA. The hydrogen bond interactions between the NSP2 residues (gray) and RNA are displayed as dashed lines. Residues involved in hydrophobic contacts are shown as spheres. A red dashed line indicates the cation-interaction between R104 and G1. (B) Detailed interactions between SA11 NSP2 and RNA, as determined using LIGPLOT (36). The amino acid residues of NSP2 and the RNA involved in the interactions are labeled. Water molecules mediating RNA recognition are represented by small spheres in cyan. The carbon, nitrogen, and oxygen atoms are shown in black, blue, and red colored circles, respectively. Hydrogen bonds are shown as green lines between the respective donor and acceptor atoms, along with the acceptor-donor distance. The NSP2 residues involved in the hydrophobic interactions are indicated by an arc with spokes radiating toward the RNA atoms that they make contact with. The contacted atoms are shown with spokes radiating back.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:6.585.81.508.35.635.2]CC residues upon mutating basic residues K223 and R227 within the cleft to alanine (Fig. 7,Table 2). Despite these mutations, since the other residues involved in GG specific interactions such as R104 are not altered, GG is likely to exhibit residual cleft binding, contributing to the differential affinities between GG and CC nu-cleotides.
DISCUSSION
NSP2 exhibits both sequence-specific and sequence-indepen-dent RNA binding.Our studies provide a structural rationale for the seemingly contradictory results of biochemical studies show-ing that RNA bindshow-ing by NSP2 is sequence independent (13,32) and yet it can target rotavirus (⫺)RNA with a highly conserved 5= GG motif for RTPase activity (37). Our crystallographic studies, together with SPR analysis, indicate that NSP2 exhibits both types of RNA binding. When presented with an RNA that has a GG motif at the 5=end, NSP2 can target it for RTPase activity through extensive sequence-specific interactions that involve highly con-served residues within the enzymatic cleft. NSP2 can also bind to RNA in a sequence-independent manner through the C-terminal helix and other basic residues that line the grooves of the octamer. A new finding from our SPR analysis is that the C-terminal helix has a profound effect on the RNA binding of NSP2. This helix with its ability to exist in two distinct conformations is likely involved in enhancing both types of RNA binding through electrostatic steering effects. It can be surmised that the quaternary organiza-tion of NSP2 subunits into an octamer allows the integraorganiza-tion of both GG-specific RNA “cleft binding” with sequence-indepen-dent “groove binding.” The 4-2-2 octamer organization of the NSP2 subunits, which also leads to a 2-fold related disposition of the basic residues along the grooves, may be the basis for the ob-served helix-destabilizing activity of NSP2.
It is clear from both our SPR analysis and previous studies (32)
that NSP2 binds to any single-stranded RNA. However, we have not been successful in visualizing crystallographically the binding of oligoribonucleotides other than the 5=GG oligonucleotide. A reasonable explanation is that the binding of the non-GG nucle-otides, which involve interactions between the phosphate groups of the RNA and basic residues that line the groove, is nonspecific and varies within and between the NSP2 octamers and thus be-comes transparent in crystallographic analysis. It is for the same reason that the remaining five nucleotides of the 5=CS(⫺) used in our crystallographic studies were not visualized. These nucleo-tides that emerge from the cleft can bind the octamer groove in multiple orientations.
NTPase and RTPase activities share the same catalytic mech-anism.The binding of 5=CS(⫺) within the cleft and proximal to the catalytic residue H225, similar to that of ATP analogs in pre-vious crystallographic studies (14), is consistent with biochemical studies (37) showing that both RTPase and NTPase activities are mediated by H225 and that they share a catalytic mechanism in-volving transient phosphorylation of H225. However, compared to the binding of the ATP analogs in which only the terminal phosphates and the sugar moiety of the nucleoside could be ob-served in the crystal structures (14), our crystallographic studies show that the entire terminal GG is firmly anchored to the cleft and makes substantial interactions with cleft residues. This obser-vation is in accordance with previous biochemical studies that indicate 5=RNA binds to NSP2 with tighter affinity than the NTPs and that the former is a better substrate for␥-phosphate hydroly-sis (37). The observation that most residues involved in coordi-nating the binding of the 5=CS GG motif are highly conserved among the group A rotaviruses strongly support the notion that a GG-specific interaction is relevant in the context of rotavirus rep-lication. The hydrolysis of the␥-phosphate of rotavirus (⫺)RNAs may indeed be a consequence of this specific recognition. Al-FIG 5Domain swapping of the C-terminal helix of NSP2. (A) Superposition of the NSP2 subunits with C-terminal helix in open (orange), and closed (blue) conformations. The dihedral angle changes in residues M293, K294, and P295 lead to the C-terminal helix switching its conformation. (B) Two NSP2 tetramers (green and red) interact with each other through swapped C-terminal helices. (C) Domain-swapping interactions of the C-terminal helices between the octamers lead to the formation of a chain of NSP2 octamers in the crystal lattice.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:7.585.97.488.66.290.2]though it is currently unclear as to which of the enzymatic activi-ties of NSP2 predominates during rotavirus replication, a strong possibility is that NSP2 uses different combinations of these cata-lytic activities or switching from one to another during genome replication and packaging.
In the viroplasms, which are the exclusive sites for rotavirus genome/encapsidation, only RNA species with a 5=GG sequence that can be substrates for 5= RTPase activity are the nascent (⫺)RNAs, since the 5=ends of (⫹)RNAs are capped. NSP2 does not exhibit capping or the cap-binding activity. A relevant ques-tion is whether NSP2 is the only protein that is responsible for the removal of␥-phosphate from these (⫺)RNAs. Currently, NSP2 is the only rotavirus protein that has been experimentally shown to have 5=RTPase activity (37). Another candidate is rotavirus VP3, which is responsible for 5=capping of the (⫹)RNA during
endog-enous transcription (5,8,17). The formation of 5=caps on rota-virus (⫹)RNA requires removal of the ␥-phosphate. However, conservation of the GG motif in the rotavirus (⫺)RNAs together with the highly conserved nature of the NSP2 residues that inter-act with this motif argues for NSP2 as the most likely candidate responsible for the␥-phosphate hydrolysis of (⫺)RNAs of rota-virus.
Although the absence of a␥-phosphate is a characteristic fea-ture of rotavirus (⫺)RNAs, its significance during virus replica-tion is unclear. Whether it is a countermeasure against host anti-viral response triggered by triphosphate RNA such as activation of the RIG-I pathways (28,42) or whether it is also necessary for a particular function during rotavirus genome replication/encapsi-dation requires further study. It is conceivable that the specific binding of the NSP2 to the 5=CS of the (–)RNA by NSP2 is re-FIG 6NSP2⌬CTforms octamers. (A) Size-exclusion chromatography of wide-type and mutant NSP2s using Superose 6 (GE Healthcare), with elution volumes
for each peak shown in the figure. (B) SDS-PAGE analysis of purified NSP2, NSP2⌬CT, and NSP2K223A/R227A/⌬CT. (C) Crystal structure NSP2⌬CToctamer
determined to 3.4-Å resolution (above) compared to the previously published NSP2 I422 octamer (11) (below). Two NSP2 subunits related by the 2-fold axis and involved in the tail-to-tail interaction of two NSP2 tetramers are shown in green and gold, respectively. The rest subunits of the octamers are shown in tan.
Hu et al.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:8.585.94.493.64.486.2]quired to confer structural organization of the newly synthesized duplex RNA around VP1 for subsequent encapsidation. Each VP1-VP3 heterodimeric complex inside the DLPs may associate with one segment to allow simultaneous transcription of dsRNA segments (15, 26, 36). Based on previous biochemical studies, which strongly suggest the possibility of direct interaction be-tween NSP2 and VP1 (3,12), one model that can be envisioned is that the specific recognition of 5=CS of (⫺)RNA and sequence-independent binding of the 3=end of the (⫹)RNA of the duplex as it emerges from VP1 allows for proper structural organization of the genome segment around VP1. This might then allow for sub-sequent packaging into assembling core particles such that each segment is associated with one VP1 molecule inside the assembled particles, which then would be conducive for simultaneous
tran-scription of the genome segments during endogenous transcrip-tion.
The C-terminal helix exhibits two distinct conformational states.A novel observation from our studies with untagged NSP2 is that the C-terminal helix of NSP2 exhibits two distinct confor-mational states, referred to here as the closed and open conforma-tions. In the previous X-ray crystallographic studies of His-tagged NSP2, in the presence or in the absence of NTP analogs, the C-terminal helix exists only in the closed conformation. It is possible that the addition of His6residues to the C terminus
[image:9.585.84.504.68.361.2]prevents this conformational change. What triggers this confor-mational change in only one subunit of the tetramer is puzzling. From the observation that all subunits irrespective of the confor-mational state of their C-terminal helices show density for the FIG 7RNA-binding assays using SPR. (A) Representative sensograms for wild-type and mutant NSP2 at 50 nM (green), 100 nM (red), and 200 nM (black) binding to GG RNA. (B) Representative sensograms for wild-type and mutant NSP2 binding to CC RNA with the same color scheme shown in panel A. The association lasts 240 s and is followed by disassociation for 180 s. The deducedkon,koff, and apparentKdvalues are shown inTable 2.
TABLE 2Binding of “GG” RNA and “CC” RNA to NSP2 and NSP2 mutantsa
Protein RNA kon1 (M⫺
1s⫺1) k off1 (s⫺
1) k
on2 (M⫺
1s⫺1) k off2 (s⫺
1) K
d1 (nM) Kd2 (nM)
Wild type GG 1.50⫻106 3.80⫻10⫺2 1.98⫻105 1.99⫻10⫺3 25.3 10.1
CC 7.30⫻105 2.60⫻10⫺2 1.76⫻105 2.33⫻10⫺3 35.6 13.2
⌬CT GG 8.26⫻104 5.18⫻10⫺3 62.7
CC 5.42⫻104 8.76⫻10⫺3 161.6
K223A/R227A/⌬CT GG 3.56⫻104 3.33⫻10⫺3 93.5
CC 1.81⫻104 6.41⫻10⫺3 354.1
a
Association rate constants (kon), dissociation rate constants (koff), and apparent dissociation constants (Kd) were obtained from SPR assays. Sensograms of wild-type NSP2 were fit to a heterogeneous ligand (parallel reactions), whereas sensograms of the⌬CT and K223A/R227A/⌬CT mutant NSP2 were fit to a 1:1 (Langmuir) binding model.
on November 7, 2019 by guest
http://jvi.asm.org/
[image:9.585.40.546.610.706.2]bound RNA, it is unlikely that the RNA-binding affects this switching. However, without further studies we cannot rule out subtle cooperativity between the subunits, i.e., binding of RNA initially to one subunit triggers the open conformation, along with small changes in the intersubunit interactions, thus allowing the remaining subunits to bind RNA but without causing a confor-mational change in the C-terminal helix. A simple explanation for this, however, is that the conformational change is the result of crystallization conditions and crystal packing forces. Nonetheless, the ability of the C-terminal helix to exist in distinct conforma-tional states is clearly evident.
C-terminal helix is involved in domain-swapping interac-tions.Another interesting and novel observation from our struc-tural analysis is the involvement of the C-terminal helix in an open conformation in the domain-swapping interactions resulting in the formation of chain of NSP2 octamers. Two residues, K294 and P295, in the loop region that connects the C-terminal helix to the rest of the C-terminal domain provide a switch for the open and closed conformations. Proline residues are frequently associated with domain-swapping interactions in proteins (21, 27). Both K294 and P295 are highly conserved in the NSP2 of both group A and C rotavirus, which implies that these residues are functionally significant. Considering that NSP2 is critical for viroplasm forma-tion and is one of the most abundant proteins in the viroplasm with concentrations similar to those found in crystals, it stands to reason that the domain-swapping interaction provides an under-lying mechanism for viroplasm formation.
In conclusion, our studies have provided novel insights into RNA-binding characteristics of NSP2, a multifunctional protein that is critical for rotavirus replication. We have provided a struc-tural basis for specific recognition of the 5=CS of rotavirus (⫺)RNA by NSP2 and how the symmetric organization of the NSP2 octamer facilitates both specific and sequence-independent RNA binding. A novel observation from our studies is that the C-terminal helix of NSP2 exhibits two distinct confor-mations and engages in domain-swapping interactions that may have functional implications during viroplasm formation. Our SPR analysis highlights the importance of the C-terminal helix in RNA binding of NSP2 and suggests that it plays a role in electro-static steering of the RNA. Although these SPR studies have pro-vided a simple qualitative picture of the RNA-binding character-istics of NSP2, more sophisticated binding analyses are required to fully comprehend the kinetics of this interaction in the context of the symmetric organization of the NSP2 octamer. Further struc-tural and functional studies are required to understand how NSP2 with multiple binding partners uses its RNA-binding and enzy-matic activities to coordinate the complex and dynamic processes of viroplasm formation, genome replication, and encapsidation in rotavirus.
ACKNOWLEDGMENTS
We acknowledge the support from National Institutes of Health (NIH) grants AI36040 (B.V.V.P.) and AI080656 and P30 DK56338 (M.K.E.) and of the Robert Welch Foundation (Q1279) to B.V.V. P.J.T.P. was sup-ported by the Intramural Research Program of the National Institute of Allergy and Infectious Disease, NIH. SBC-CAT 19ID at Advanced Photon Source is supported by the U.S. Department of Energy, Basic Energy Sci-ences, Office of Science, under contract W-31-109-Eng-38.
We thank Zenobia Taraporewala, Kristen Ogden, and Marco Morelli for helpful comments on the manuscript. We acknowledge the use of the synchrotron beamlines at Argonne National Laboratories (SBC-CAT
19ID), Chicago, IL, for diffraction data collection. We thank their staff for excellent help.
REFERENCES
1.Adams PD, et al.2010. PHENIX: a comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. D Biol. Crystal-logr.66:213–221.
2.Aponte C, Poncet D, Cohen J.1996. Recovery and characterization of a replicase complex in rotavirus-infected cells by using a monoclonal anti-body against NSP2. J. Virol.70:985–991.
3.Arnoldi F, Campagna M, Eichwald C, Desselberger U, Burrone OR.
2007. Interaction of rotavirus polymerase VP1 with nonstructural protein NSP5 is stronger than that with NSP2. J. Virol.81:2128 –2137.
4.Brunger AT, et al.1998. Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr. D Biol. Crystallogr.54:905–921.
5.Chen D, Luongo CL, Nibert ML, Patton JT.1999. Rotavirus open cores catalyze 5=-capping and methylation of exogenous RNA: evidence that VP3 is a methyltransferase. Virology265:120 –130.
6.Emsley P, Lohkamp B, Scott WG, Cowtan K.2010. Features and devel-opment of Coot. Acta Crystallogr. D Biol. Crystallogr.66:486 –501. 7.Estes MK, Greenberg HB.Rotaviruses.InKnipe DM, et al. (ed), Fields
virology, 6th ed, in press. Lippincott/The Williams & Wilkins Co, Phila-delphia, PA.
8.Fukuhara N, Nishikawa K, Gorziglia M, Kapikian AZ.1989. Nucleotide sequence of gene segment 1 of a porcine rotavirus strain. Virology173: 743–749.
9.Imai M, Akatani K, Ikegami N, Furuichi Y.1983. Capped and conserved terminal structures in human rotavirus genome double-stranded RNA segments. J. Virol.47:125–136.
10. Jayaram H, Taraporewala Z, Patton JT, Prasad BV.2002. Rotavirus protein involved in genome replication and packaging exhibits a HIT-like fold. Nature417:311–315.
11. Jiang X, et al.2006. Cryoelectron microscopy structures of rotavirus NSP2-NSP5 and NSP2-RNA complexes: implications for genome replica-tion. J. Virol.80:10829 –10835.
12. Kattoura MD, Chen X, Patton JT.1994. The rotavirus RNA-binding protein NS35 (NSP2) forms 10S multimers and interacts with the viral RNA polymerase. Virology202:803– 813.
13. Kattoura MD, Clapp LL, Patton JT.1992. The rotavirus nonstructural protein, NS35, possesses RNA-binding activity in vitro and in vivo. Virol-ogy191:698 –708.
14. Kumar M, et al.2007. Crystallographic and biochemical analysis of ro-tavirus NSP2 with nucleotides reveals a nucleoside diphosphate kinase-like activity. J. Virol.81:12272–12284.
15. Lawton JA, Estes MK, Prasad BV.1997. Three-dimensional visualization of mRNA release from actively transcribing rotavirus particles. Nat. Struct. Biol.4:118 –121.
16. Li Z, Baker ML, Jiang W, Estes MK, Prasad BV. 2009. Rotavirus architecture at subnanometer resolution. J. Virol.83:1754 –1766. 17. Liu M, Mattion NM, Estes MK.1992. Rotavirus VP3 expressed in insect
cells possesses guanylyltransferase activity. Virology188:77– 84. 18. Lu X, et al.2008. Mechanism for coordinated RNA packaging and
ge-nome replication by rotavirus polymerase VP1. Structure16:1678 –1688. 19. McCoy AJ, et al.2007. PHASER crystallographic software. J. Appl.
Crys-tallogr.40:658 – 674.
20. McCrae MA, McCorquodale JG.1983. Molecular biology of rotaviruses. V. Terminal structure of viral RNA species. Virology126:204 –212. 21. Miller KH, Karr JR, Marqusee S.2010. A hinge region cis-proline in
ribonuclease A acts as a conformational gatekeeper for C-terminal domain swapping. J. Mol. Biol.400:567–578.
22. Otwinowski Z, Minor W.1997. Processing of X-ray diffraction data collected in oscillation mode,InCarter CW, Jr, Sweet RM (ed), Methods in enzymology, vol 276, p 307–326. Academic Press, New York, NY. 23. Patton JT, Silvestri LS, Tortorici MA, Vasquez-Del Carpio R,
Tarap-orewala ZF.2006. Rotavirus genome replication and morphogenesis: role of the viroplasm. Curr. Top. Microbiol. Immunol.309:169 –187. 24. Pettersen EF, et al. 2004. UCSF Chimera: a visualization system for
exploratory research and analysis. J. Comput. Chem.25:1605–1612. 25. Pflugrath JW.1999. The finer things in X-ray diffraction data collection.
Acta Crystallogr. D Biol. Crystallogr.55:1718 –1725.
Hu et al.
on November 7, 2019 by guest
http://jvi.asm.org/
26. Prasad BV, et al.1996. Visualization of ordered genomic RNA and local-ization of transcriptional complexes in rotavirus. Nature382:471– 473. 27. Rousseau F, Schymkowitz JW, Wilkinson HR, Itzhaki LS.2001.
Three-dimensional domain swapping in p13suc1 occurs in the unfolded state and is controlled by conserved proline residues. Proc. Natl. Acad. Sci. U. S. A.98:5596 –5601.
28. Schmidt A, et al.2009. 5=-triphosphate RNA requires base-paired struc-tures to activate antiviral signaling via RIG-I. Proc. Natl. Acad. Sci. U. S. A.
106:12067–12072.
29. Schrödinger.2010. The PyMOL molecular graphics system, v1.3. Schrödinger, LLC, Munich, Germany.
30. Schuck P, Taraporewala Z, McPhie P, Patton JT.2001. Rotavirus non-structural protein NSP2 self-assembles into octamers that undergo ligand-induced conformational changes. J. Biol. Chem.276:9679 –9687. 31. Settembre EC, Chen JZ, Dormitzer PR, Grigorieff N, Harrison SC.
2011. Atomic model of an infectious rotavirus particle. EMBO J.30:408 – 416.
32. Taraporewala Z, Chen D, Patton JT.1999. Multimers formed by the rotavirus nonstructural protein NSP2 bind to RNA and have nucleoside triphosphatase activity. J. Virol.73:9934 –9943.
33. Taraporewala ZF, et al.2006. Structure-function analysis of rotavirus NSP2 octamer by using a novel complementation system. J. Virol.80: 7984 –7994.
34. Taraporewala ZF, Patton JT.2001. Identification and characterization of
the helix-destabilizing activity of rotavirus nonstructural protein NSP2. J. Virol.75:4519 – 4527.
35. Tortorici MA, Broering TJ, Nibert ML, Patton JT. 2003. Template recognition and formation of initiation complexes by the replicase of a segmented double-stranded RNA virus. J. Biol. Chem.278:32673–32682. 36. Trask SD, Ogden KM, Patton JT.Interactions among capsid proteins
orchestrate rotavirus particle functions. Curr. Opin. Virol. in press. 37. Vasquez-Del Carpio R, Gonzalez-Nilo FD, Riadi G, Taraporewala
ZF, Patton JT.2006. Histidine triad-like motif of the rotavirus NSP2 octamer mediates both RTPase and NTPase activities. J. Mol. Biol.
362:539 –554.
38. Wallace AC, Laskowski RA, Thornton JM.1995. LIGPLOT: a program to generate schematic diagrams of protein-ligand interactions. Protein Eng.
8:127–134.
39. Wentz MJ, Patton JT, Ramig RF. 1996. The 3=-terminal consensus sequence of rotavirus mRNA is the minimal promoter of negative-strand RNA synthesis. J. Virol.70:7833–7841.
40. Wentz MJ, Zeng CQ, Patton JT, Estes MK, Ramig RF.1996. Identifi-cation of the minimal replicase and the minimal promoter of negative-strand synthesis, functional in rotavirus RNA replication in vitro. Arch. Virol. Suppl.12:59 – 67.
41. Winn MD, et al.2011. Overview of the CCP4 suite and current develop-ments. Acta Crystallogr. D Biol. Crystallogr.67:235–242.
42. Yoneyama M, Fujita T.2009. RNA recognition and signal transduction by RIG-I-like receptors. Immunol. Rev.227:54 – 65.