0022-538X/80/12-0816/13$02.00/0
Analysis
of
Herpes
Simplex
Virus
Nucleoprotein
Complexes
Extracted
from Infected Cells
PIER F. PIGNATTII* AND ENZO CASSAI2
Istituto diGenetica, Universitd diPavia,27100Pavia,'andIstituto diMicrobiologia, Universitadi
Ferrara,
44100
Ferrara,2
Italy
HEp-2 cells were infected with herpes simplex virus type 1 and labeled with
[3H]thymidine and '4C-amino acids. Infected cells or nucleipreparedfromthem
wereextractedwithTriton X-100 andNaCl,utilizingamethod
recently
described,and thelow-speed supernatant(extract) waspartially purified bysedimentation on sucrosegradients. Anucleoproteincomplexwhichsedimentedas awidepeak
around 200S was identified. The
nucleoprotein complex
contained viral DNA,which bandedattheexpecteddensityinCsClisopycnic gradientsandwasintact
after measurements taken on electron microscopic
photographic
enlargements.Theautoradiographic patternof
"C-labeled
proteinsafterelectrophoresis
showedthatonlya few of thevirus-specificpolypeptideswerepresentin thenucleoprotein
complexes,inparticular, VP5, VP12,VP15.2, VP19, andVP24.Cellularhistones
were absent. The extractsand thenucleoprotein complexes werecentrifugedto
equilibriuminmetrizamidedensity gradientswithoutprefixation. Electron
micro-scopic direct visualization of thenucleoprotein complexesaftersucrose or
metri-zamidepurificationrevealed thattheproteinswere
preferentially
associatedwithoneendofthe DNAmolecule andformed
large irregular
terminalthickenings
orcapsid-like transparent shellsenclosingpolyglobularcores.Nonucleosomeswere
observed on herpes simplex virus nucleoprotein complexes. The same type of
complexwas detected after phosphonoacetic acid addition, and grossly altered
nucleocapsidswereformed.
Nucleoprotein complexes (NPC) of small
DNA tumor viruses showing chromatin-like
structureshave been isolated fromcellsinfected
with polyoma virus (4) and simian virus 40 (4,
10,22)orfrompapilloma (7)and BK(18)virions.
The isolation of soluble minichromosomes has
greatly accelerated structural and functional
studies of these viruses. As to medium-sized
DNA tumor viruses, the replicative
intermedi-ates of adenoviruses show a different type of
structure which is devoid ofnucleosomes (15).
For the class oflarge DNA tumor viruses, the
herpesviruses, no such soluble complexeshave
asyetbeendescribed.
Herpesvirions areschematicallycomposedof
a centrally located core surrounded by three
concentricstructures: aproteincapsid about 100
nmindiameter,thetegument, and theenvelope
(23). Electronmicroscopicobservation ofherpes
simplex virus (HSV)-purified nucleocapsids (8)
andofthin sections ofHSV-infected cells (8, 12,
19) has allowed the identification ofviral core
particles containing DNA which might be
spooled aroundacentral mass ofproteins.
Tolook for the presence of HSV NPCduring
productiveinfection,wehaveused apreparation
methodwedescribedrecently (21) which allows
the rapid extraction of HSV DNA from the
infected cells without contaminationwith
cellu-larchromatin. This paper reports the isolation
ofHSV NPC and theanalysis of theirstructure
and composition. A tentative interpretation of
the NPC role in the growth cycle of
herpesvi-ruses based on present data and on previous
results is
provided.
MATERIALS AND METHODS Cell culture and virus infection. HEp-2 cells weregrown in Eagle minimal essential medium sup-plemented with 10%fetal calfserum.When thecells approached confluency, theywereinfected withHSV type 1 (HSV-1), strain F (6),at amultiplicityof1to 5 PFU/cellfor 2 hinphosphate-bufferedsaline supple-mented with 1%inactivatedcalf serum. After aspira-tion of the viralinoculumandwashing,thecells were maintained inEagle minimal essential medium sup-plemented with 1% dialyzed fetal calfserum and la-beled for the times stated in each experiment with
['3H]thymidine (5 1Ci/ml). Labelingmedium with
"C-amino acids (0.7,ICi/ml) contained one-tenth ofthe normalaminoacid concentration. The cellmonolayers were washedtwice with ice-cold phosphate-buffered saline, scraped witharubberpoliceman,collectedby low-speed centrifugation, andfrozenat-80°C.
Preparation of NPC. Extractswere prepared as described previously(21)bythawinginfected cell sam-plesand mixing them with9volumes oflysing solution containing 10mMTris-hydrochloride (pH7.9)-0.25%
816
on November 10, 2019 by guest
http://jvi.asm.org/
HSV DNA-PROTEIN COMPLEXES 817
Triton X-100-0.2 M NaCI-l0mM EDTA. Infected-cell nuclei were prepared according to the technique of Burgoyne et al. (3), and then extracts were prepared as described above. The suspension was clarified by low-speed centrifugation, and the supernatant (ex-tract) waslayered on linear 20 to 40% (wt/vol) sucrose gradients with 0.6ml of a 77% (wt/vol) sucrose cushion which contained0.01MTris-hydrochloride (pH 7.9)-0.2 MNaCI-1 mM EDTA. The gradients were centri-fuged in anSW40rotor at35,000 rpm and 4°C for 30 min.Fractions werecollected from the bottom of the tube,and acid-insolubleradioactivity was determined bytheglass fiber filter procedure (2).
Cytoplasmic nucleocapsids were prepared with the detergent method ofGibson and Roizman (9) and then weresubjected to velocity sedimentation analysis as describedpreviously.
CsClequilibrium density gradients. The DNA samplesweredeproteinized by incubation in the pres-ence of 100 ug of proteinase K per ml-0.2% sodium dodecyl sulfate-l0 mM EDTA-10 mM Tris-hydro-chloride(pH 7.5) for2hat37°C. CsClwasthen added and thesolutionwasadjustedto adensity of 1.720 g/
cm3. The DNA solution was centrifuged in a 65 Ti rotor at 40,000 rpm and20°C for 62 h, and fractions of 0.16mlwere collected from the bottom of the tube. Acid-insoluble radioactivity was counted as above. Ethidium bromide fluorescence was scored under a UVlamp after spottingasample of each fraction on WhatmanGF/C filters and soaking them inasolution containing0.5,ugof ethidiumbromide per ml in Tris-hydrochloride (pH 7.5)-lmMEDTA.
Metrizamide density gradients. Metrizamide
(Nyegaard & Co., Oslo,Norway) solutions were pre-pared inabuffercontaining 10 mM Tris-hydrochloride (pH 7.5)-0.2 M NaCl-I mMEDTA, filtered onglass fiber paper(GF/A Whatman) filters,andadjustedto pH 7.5 with NaOH. Discontinuous gradients were preparedasdescribedpreviously(22) andcentrifuged in the65 Ti rotor at 40,000 rpm and4°C for 39 h. Fractions werecollected,anddensity wasdetermined accordingtotheformula p=3.350c-3.462(1),where cis the refractive index.
Acrylamide gel electrophoresis and
autora-diography. Virion polypeptides (VP) and
infected-cell polypeptides were prepared as described
previ-ously (11, 13). Nucleoprotein sampleswereprepared
by dilutingappropriate sucrosegradient fractions in 10mMTris-hydrochloride (pH 7.5) and bypelleting
the NPC in the65Tirotor at40,000 rpm and4°C for 3 h. The pellets were thencollected and treated as described above. Calfthymushistonespurchasedfrom
Sigma Chemical Co. (St. Louis, Mo.) were used as
electrophoreticstandards.Electrophoresis onsodium
dodecyl sulfate-containing polyacrylamide gel slabs
cross-linked withdiallyltartardiamideandgel analysis
wereperformedasdescribedpreviously(11).
Electron microscopy. NPC were prepared for electronmicroscopic observation withoutprior fixa-tion,using theproteinlessmethod of Dubochetetal. (5)asdescribed previously (4, 22).Theaqueous mi-crodroptechniqueofKleinschmidt (17)wasalso uti-lized for DNA visualization. The samples were ob-served withaPhilipselectronmicroscopeEM300at
60kV,andphotographsweretakenatmagnifications
rangingfromx14,700 to x36,100. Measurements were done on photographic enlargements: diameters were determined with a lenswith0.1-mm graduations, and DNA contour lengths were determined with a map ruler. Calibration was based on examination of cross-grating replicas (Polaron Equipment Ltd., Watford, England).
RESULTS
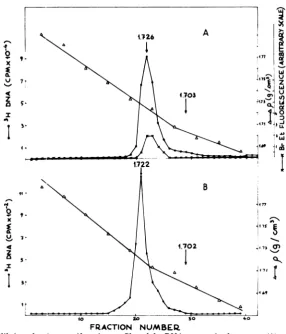
Analysis of the extracts. HEp-2 cells were
infected with HSV-1, grown in thepresence of
[3H]thymidine, collected, and extracted with
Triton X-100 and NaCl-containingbuffer at
var-ious times after infection. In the extracts only
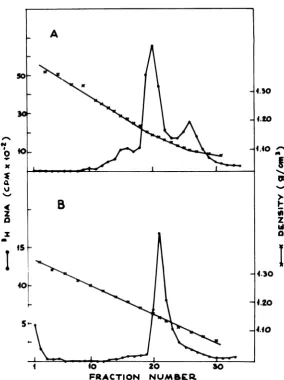
onepeak of DNAwas present, and it banded at
1.726g/cm3inCsCl gradients (Fig. 1A), a density
correspondingtotheexpected HSV-1 (strain F) DNAvalue (16). The absence ofcellularDNA
in the extracts was confirmed by the lack of
ethidium bromidefluorescence in the
appropri-ate gradient fractions (Fig. 1A), and it is in
accord with previous results(21)obtained with
cells which had been labeled with
['4C]thymi-dine before viral infection. The extracts
con-tainedincreasingamountsof viralDNA atlater timesafter infection.
Preparation andpreliminary characteri-zation of NPC.
Samples
of the extracts werelayeredon 20 to40%sucrosegradientsand
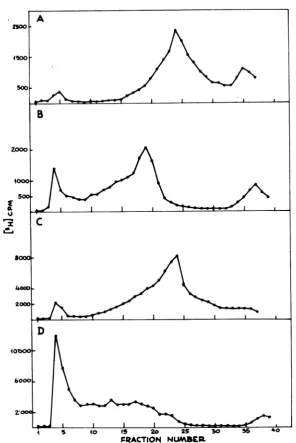
cen-trifugedasdescribed above.Abroadpeak
sedi-menting
atabout 200Sappeared (Fig. 2A),
which could be shiftedtothe HSV DNA sedimentation value afterproteinase K deproteinization (21). NPC were absent in cells collected 3 and 6 hpostinfection, appeared at 9 h, and then were
present atall latertimes
tested, i.e.,
12, 18, 32,and48hafter
infection,
when celllysis usually
occurred. Thesamesedimentationvelocity
wasobserved for NPC obtained at different times after infection.
Figure
2B shows theresults ofanextraction fromanuclear
preparation.
AnNPC with thesamesedimentation velocityasin Fig.2A was
identified;
thepeak
wasbroader,
andrelatively moreviral DNA (in fractions 33 and 34) and mature
nucleocapsids (on
the sucrosecushion in fractions4 and
5)
werepresent. The DNA from thesucrose-purified
NPC bandedinCsCl
isopycnic
gradients
as asingle species
dis-playing viralDNA density (seeFig.
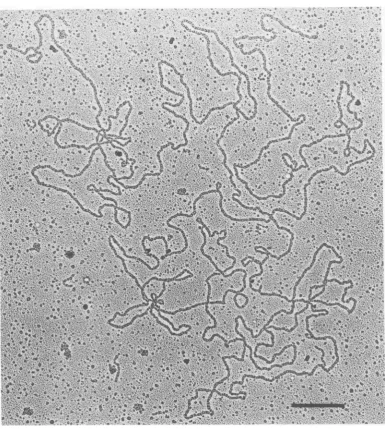
1B). DNAmolecules about45
pm
long
werepredominantly
observed in the NPC peak after
deproteiniza-tion. Figure 3 shows one such
filament,
whichwasfoundtobe 46.2
,m long.
Proteins present in NPC. To
analyze
theprotein component of the
complex,
infectedHEp-2 cellswere labeled with
14C-amino
acidsand then
collected;
theextracts weresubjected
to
velocity sedimentation
as described above.Figure 4 shows that, whereas most
14C-labeled
VOL. 36,1980
on November 10, 2019 by guest
http://jvi.asm.org/
818 PIGNATTI AND CASSAI
A
to
I
4..
11
-x
%-O
4c
a
5-I
3.
4.703
I
B
1.702
]
2_
R.471
N ,,
10°
*4.71 JL
.
1*
IA'
117
q75
FRACTION
NUMBER.
FIG. 1. Equilibriumdens.tycentrifugationprofilesofthe DNApresentintheextract(A)and in the NPC (B). The cellswerelabeled from5to 18hafter infectionwith[3H]thymidine, andthenextraction, sucrose gradientpurification,sampledeproteinization, andCsClcentrifugationwereperformedasdescribed in the text.(A) Triton-NaClextrac&; (B)pooled fractions15to22ofthegradientshowninFig. 2A,dialyzed against
10mMTris-hydrochloride (pH 7.5)-10mM EDTAfor16hat4'C.Arrows indicate observedpeak density
(1.726 and 1.722g/cm3) or expected cell DNA density (1.703and 1.702g/cm3), as determined inparallel
gradients.
polypeptidespresentintheextractweresoluble
and did not penetrateint-o the
gradient,
about1%of the labeledprotein
pool
waspresentintheNPC
peak,
which coincidedwith the [3H]DNApeakshown inFig. 4D.All experimentinwhich
14C-labeled
amino acids werealreadypresent inthe medium24hbefore viral infection
(Fig.
4B)
gaveresults similar tothose shown in
Fig.
4A,and in
particular,
noincreaseintheNPCradio-activity peak was observed. The
analysis
ofacontrol uninfected cell extract prepared at the
sametime after"4C-aminoacid
labeling
for48h(Fig.4C) showed thatno cellularproteins
sedi-mented at around200S in the absence of viral
infection, but all remained at the top of the gradient.
To determine the protein species associated
with viral DNA in the NPC, the appropriate
sucrosegradientfractionswerediluted, pelleted,
electrophoresed,andautoradiographed.The
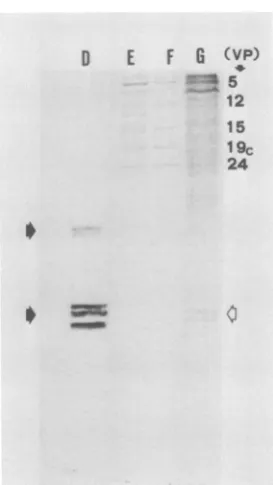
au-toradiographic pattern of
"C-labeled
proteinspresentin theNPCis shown inFig. 5.Itcanbe
noted that all of theproteinspresentinthe NPC
(Fig. 5, lane B) isoelectrophoresed with some
corresponding infected-cell and virion
polypep-tides. From the analysis of the electrophoretic
migration rates in this and other gels, the follow-ing conclusions were reached: VP5, -12, -15.2,
-19,and-24werealways present as clear bands
in theNPC. In addition, some
small-molecular-weight polypeptides, perhaps degradation
prod-ucts, migrated with the electrophoretic front.
Sometimes traces ofVP7, VP8.5, VP13, VP14,
and VP16werealso detected. It should be noted
thatmanyvirion polypeptides were always
ab-sent from the complex, in particular, some of
those represented most often in the viral
parti-cles, suchasVP8, VP17, VP18, and VP22. The
on November 10, 2019 by guest
http://jvi.asm.org/
[image:3.496.120.407.71.406.2]HSV DNA-PROTEIN COMPLEXES 819
A
I II
S I I
4 5 40 45 to Zs 30 35
B
4 5 40 15 20 25 s0 35
FRArTION N UM
FIG. 2. Sedimentationvelocity profiles of HSV-infectedcellular(A)ornuclear(B)extracts. Infectedcells
werelabeledfrom5 to 18 hafter infectionwith[3H]thymidine, collected,and dividedinto twosamples.One
samplewasextracteddirectly (A),whereasfromthe othersamplenucleiwerefirstisolated andthen thesame
extractionprocedurewasapplied (B).The twosamplesweredepositedontosucrosegradientsandcentrifuged asdescribed in thetext.Directionofsedimentation isfrom righttoleft.Thearrowindicates the sedimentation position ofanHSV-1(strain F)DNA marker.
NPCwas thusparticularly rich inthree capsid
proteins (VP5, VP19,andVP24),whereas itwas
devoid ofothers, e.g.,VP22.
Slightlymodifiedgelsin whichtheacrylamide
concentration was raised from 8.5 to 15% were
utilized for a better separation of cellular his-tones. One such electrophoretic analysis is shown in Fig. 6. Histones (Fig. 6, D) were not detected in HSV-1 NPC (Fig. 6, F)orinvirions
(Fig. 6, E),buttheywerepresentinthe
infected-cellpolypeptides (Fig. 6, G).
A searchwasmade forenzymesand cofactors retained in the NPC by investigating
endoge-nous DNA and RNApolymerizingactivities in
the sucrose gradient fractions. Cellulara, fi, y,
and HSV DNApolymerases were allpresent in
the extracts (20),whereas noendogenous HSV
DNA synthetic activity was detected in the
NPC. Endogenous RNA polymerase activity
wasdeterminedaccordingtoWilhelmetal.(25):
no [3H]uridine monophosphate was
incorpo-rated bythe viral NPC fractions. Extraction of the infected cells with 0.4 MNaClordialysisof
pooledfractionsdid notchangethe results.
Structure of the NPC. Samples obtained from thesucrosegradientfractions
correspond- I-v
s-VOL. 36,1980
on November 10, 2019 by guest
http://jvi.asm.org/
[image:4.496.96.391.61.460.2]6'
0
b Q
42, J6 is,~ .4, .&,
411~~~~~~~~~~~~~~~~~~~
:,¶,..
*
4
.~~ *%.ILA *0~
*2% J:..7.
FIG. 3. Electronmicrograph ofan intactHSVDNAmoleculepresentintheNPCpreparation. Kleinschmidt (17)procedure. Bar,0.5 Wn.
ingtotheNPCpeakwereutilized withoutprior wereinterpreted to be slightly expanded,
capsid-fixation for electron
microscopic observation,
us- like shellsenclosingviral cores. The coreglob-ingthetechnique ofDubochetetal. (5). Afew ules weresometimeslinearly arranged (see Fig.
examplesofthestructureswhichweredetected 7A and B), but more often were in groups of
areshown inFig.7AtoG. Electron-denseirreg- three at an angle to each other and with a
ularglobulararrayswerepresent insideroughly common central contact (see Fig. 7C to G).
hexagonal electron-translucent capsules, with About 90%of the particles had DNA emerging
diametersrangingfrom 110 to 120 nm (Fig. 7B from them with noproteins associated (Fig. 7A
toF).Exceptionally, amuchlarger particle(ca. to E).Often two distinct DNA filaments clearly
150nm) wasobserved (Fig. 7G). As acompari- appeared to enter and exit from the described
son, viral nucleocapsids observed under the structure (Fig. 7A, D, and E). Exact
measure-sameconditions showed diameters of about100 ments of DNAcontour length were quite
diffi-nm(Fig. 7H andI).Thenucleoprotein particles cult with thesepreparations: the filament shown
820
PIGNATTI AND CASSAIon November 10, 2019 by guest
http://jvi.asm.org/
[image:5.496.71.457.70.500.2]VOL. 36, 1980
in Fig. 7A was about 28 jim long; other values
obtainedwerein the rangeof22to 29jim,except in one case in which a 43-,im-long filament was observed. In about 10% of the cases, no DNA
filament appeared to originate from the
parti-cles,asfromthoseshown inFig. 7F and G.
Extracts and sucrose gradient-purified
com-plexesweresubjected to isopycnic centrifugation inmetrizamide densitygradients, a method that
allows nucleoprotein density determinations
IX 0J6 WI 04 S OLa IL U0 W
2I
2-i
III a A3a
I I o u .5 3 .4 FRACTION NUMAEAFIG. 4. Sedimentationvelocityprofiles of
HSV-in-fectedcell extracts.(A) Cells labeledfrom5to 18 h after infectionwith
"4C-amino
acids. (B) Cellsincu-batedfor24 hbefore infectionandfrom5to 18 hafter infectionwith14C-aminoacids.(C)Controluninfected cells labeledinparallel for48 hwith
"4C-amino
acids. (D) Cells grown in the presence of[3H]thymidine from5to18 hafter infection.Allcellswerecollectedatthesametime, extracted, andcentrifuged as
de-scribedin thetext.Directionofsedimentation isfrom righttoleft.
HSV DNA-PROTEIN COMPLEXES 821
A R0
VP
1-2
5 m '
8 8.57 -12 13
is4
A 15.2 16 AM 17 18 19 22 23 24 A IJ t. ICP ft~ 2 faw.. 4a-c 5 7 8 10 12 17 20 24 25 29 32 33 36 39 40 41 _ 43 44FIG. 5. Autoradiogram oftheproteinspresentin
the NPCafter electrophoresisonpolyacrylamide gels.
(A) HSV-1 (strain F) virion polypeptides (VP) ob-tained from 4C-amino acid-labeled virions, num-beredaccording toHeineetal. (11). (B)NPC
poly-peptides obtainedfrom fractions 16and 17 ofthe
sucrosegradientshown inFig.4A. (C)Infected-cell "4C-labeledpolypeptides (ICP) obtained 18 h after
infection,numberedaccordingtoHoness and
Roiz-man(13).Molecularweightsofselectedpolypeptides
aregivenas areference (13): VP5= 155,000; VP12
= 87,000; VP19 = 53,000; VP24 = 25,000. Sample
preparation, electrophoresis, and autoradiography
wereperformedasdescribed in thetext.
without prior fixation of the complexes. The
extract (Fig. 8A) showedthe presence of a main
nucleoprotein peak atabout 1.15
g/cm3,
with aheavier shoulderat 1.20
g/cm3
and alighter peakat 1.12
g/cm3,
which probably corresponds tofree DNA (1). The sucrose-purified NPC (Fig.
8B) showed a single peak at about 1.15
g/cm3.
Asthebuoyant density in metrizamide depends
upon the ratio ofproteins to nucleic acids in the
complexes,itwascalculated (according to Birnie
etal.[1]) thatthe HSV NPC contained slightly
fewerproteins than DNA by weight. For
com-parison, it can berecalled that purified virions have a protein-to-DNA weight ratio of about
10.7 (11). Samples from the metrizamide
gra-wp
on November 10, 2019 by guest
http://jvi.asm.org/
[image:6.496.270.415.72.381.2] [image:6.496.43.235.189.572.2]822
PIGNATTI AND CASSAID
E
F
G
(VP)
12 15 19c 24
FIG. 6. Electrophoretic analysis of the polypep-tides present in the NPC after electrophoresis on modified gels andstaining. (D)Histones. Black ar-rows indicate histone Hl and histones H2A, H2B, H3, H4, respectively. (E, F,andG)SameasFig. 5A, B, and C, respectively. Clear arrow indicates the position ofhistone bandsin(G). Sample preparation andelectrophoresis wereperformed asdescribed in thetext.
dient fractions were directly prepared for
elec-tronmicroscopyasdescribed.Someexamplesof
the observed structures are shown in Fig. 9.
Proteinswere locatedmostly atone endofthe
DNAfilament, asshownintheoverallviews of
Fig. 9A andB.Figures9C to Fareenlargements
ofprotein-associatedDNAterminifrom
metri-zamidefractionsshowing large (upto about 500
nm in length) masses with globular
substruc-tures.Thebackgrounddebriswasinterpretedto
be smallribonucleoprotein particleswhich
sed-iment at about the same density as the HSV NPC in metrizamide gradients (14).The DNA
filamentemergingfrom such nucleoprotein
ag-gregatesor"heads" almostinvariablyshoweda smaller(Fig. 9A) orlarger (Fig. 9B) loop,
some-timesfoldingbackonitself almostentirely,asin
Fig. 9B. Thefilament shown in Fig.9A was27
pmlong; thatin Fig.9Bwas25pmlong. Other
lengthmeasuresfell between 25 and 30,m.
Phosphonoaceticacid inhibition ofvirion
biosynthesis. To searchfor different kinds of
NPCduringvirusformation,infectedcells were
growninthepresenceofphosphonoacetic acid,
an inhibitor of HSV DNA
synthesis
and virus assembly. In an experiment in which 100,g of phosphonoacetic acid perml was given at 6 h and the cells were then collected 24 h after infection, the percentage of[3H]thymidine
in-corporation into acid-precipitable DNA wasabout7.5,whereas itwas28% in controlcellsnot
treated with the drug. When the extracts were
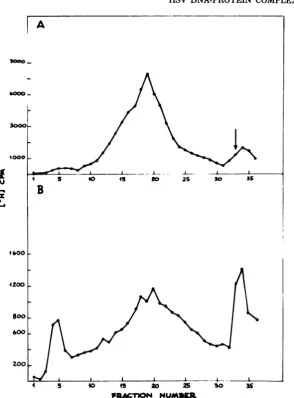
analyzedonslightly modified velocity sedimen-tation gradients (Fig. 10), very similar nucleo-protein peakswereevident both in thepresence
(Fig. 10A) and in the absence (Fig. 10C) of phosphonoacetic acid. From thesamecells
viri-ons wereprepared accordingtoGibson and
Roiz-man(9)andsubjectedtovelocity sedimentation analysis (Fig. 10B and D). A newheavier sedi-mentingpeak of NPC appeared in the phospho-noacetic acid-treated cells (Fig. 10B), whereas nucleocapsids accumulatedonthesucrose cush-ion inthe untreated control (Fig. 10D).
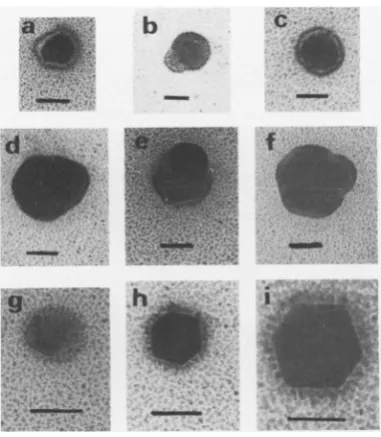
Electronmicroscopic observation of the peak fraction of Fig. 10B revealed the presence of
aberrant nucleocapsids,asshownin Fig. 11. Two
differentgroupscouldbeidentified: the first less well represented group consisted of particles
with diametersof about110 to 120nmexhibiting
structural alterations, the most common one
being the bendingoutofoneside of their
shell,
as shown inFig. lld and f. The second,larger
groupincluded particles whichwere about half
aslarge (widths ranging from50 to 75nm) and
altered: sometimes the above-described
protru-sionswereobserved, asinFig. llband e;
some-times more uniform variants were seen, as in
Fig. lla, c, and g; and sometimes miniature
capsids about 50 nm insize and with irregular
sidelengthswereseen, as inFig.llhand i.
DISCUSSION
HSV NPC have been isolated from infected cells andcharacterizedby using thesame
meth-ods utilizedpreviously (4, 22) forthe extraction
andanalysis ofpolyoma virus and simian virus 40minichromosomes from infected cellsand
nu-clei. Viral complexes devoid ofchromatin
con-tamination wereextracted similarly, but
differ-ences in NPC composition and structure were
observed. Onlysomevirus-codedproteins were
associatedwithHSV DNA, whereascellular
his-toneswere bound to polyoma virus and
simian
virus 40 genomes. Moreover, electron
micro-scopic
visualization
showed that the HSVpoly-peptides were located primarily at one end of
the DNA molecule, whereas on the contrary,
nucleosomes were distributed uniformly in the
small viralminichromosomes mentioned above.
HSVDNAthusseems to be an exception to the
eucaryotic "nucleosome rule,"possibly together
on November 10, 2019 by guest
http://jvi.asm.org/
[image:7.496.89.226.72.315.2]HSV DNA-PROTEIN COMPLEXES 823
w' '*
t.X *;;ss) w+X 44 * fw.
'I
*''*!'''
" ', ,'* "' .,*'1.' .'l,.'.'Lz'L'jS+.-,' ';- h @ '.'t p;'.', ..L'
., -,, ..,^..,, ., ... ,e;
'.
tN
''.' '";¢-ee;
'.'' .;;'':"v-'-s' ', ': ' *. ",' ;; .'':'' sr'. .: '. "'' .' ,, !, \.-' @,
X, ''">s'0,":,/
' .' '",i',:',,§ ", /':
:-,'..,',,,;,6...;;.'',
@,[email protected]
*g;'
'-t".':.,.' ;D *E ,:. ',',.s
''"-z@. ,>-
'sl'DX
',, ,. '.. X ..
{' ,v'; % 'i' " 4'
, +P+,§+ ;'S'<, .R,-. .''S
... '! '; ,, .. ..
,-, ..- -. ... ...
" .' " " '' ". "; .' .. *'
',"'.'-''',@' " ',' -s'-.''',
i¢X-}*w,',
^sS+*ff*,S,','),;;
''''.''
,' ''',','.'' ''_'r' .'t .s,';eP .. ,..,
*'-. lt '.'. " '"' ', *'.' "'8 ''.
:'.' 1.'.,,:'.';, '. .'' -;;' ". ,'.' ,' ;,e, ,' ,'. "' ':' ,'.' ,".., :. ' _ '.,.
*'."-','''',0Xf::XaS1,:
.., ,- . .
''',''.,'.,z, ',. -. ,- " ' '-.. j; '" ','v''
*..is.' ''-.'*-'', ,',*',"' .'
0
;
fft
AnA
-K;t3 H <5 t
<,: .' *,,'l. -, .,
*
-'1%1
,,: * *: ,s
i-.. . . .
...
.....
.._;
^ '_ _'
* e *-
j--r 8 *
_ S *_ , ,,', , t . *',
v
w
.. .'w.
^ o_s,,' ' .
.,, _, .
.; ,, ' _. ; .., '. _ ,
. +
.'' '-.' /.' / o
.;f m-'_s
'', *, ., ,.*_Je{';
t. .*- ,, ' . '.
_*r;
}*\@I4,4a85
'iX
;'.'.,''*.s',,X*b,-;*j,
a,X''.'
i,e'.t>t--';N
s *@v*',,*%,X@,h.
*4
*-@.@',
'.-' . ': ., d t
"' ^' -' '' ^ ' . ,- '
F ., ++ {
';,'' i'' "s+ '.;'.
* _ t.,
*w S *''* -'@ '-#-*'^"'' 1>*r ;'-;'''t'.';. w
, ,
vZ..
%X
3'- ,,s' y#;#-,*ov
. .. :, '''.
;:< -, z-, ;;,-. r..
* *< @* r ;
'-X-'#"'''s ",''--,',-,0''.'
*, ', . ._, .'
VOL. 36,1980
Cc
V.)
cc 04
I..
V.)
L.
cc;
o ,
V.
ccc cc_
s
CV.)
o
~cq
o.
on November 10, 2019 by guest
http://jvi.asm.org/
824
4 x
v
z
a
I
I
p
z
ad
I
FRACTION NUMBER.
FIG. 8. Metrizamidedensity gradientanalysis ofacellextract(A)andNPCfrom pooledsucrosegradient
fractions (B). Infected cells werelabeled with[3H]thymidine from 6to 19hafter infection, collected, and extractedasdescribed in thetext.Discontinuousmetrizamidegradientswerepreparedandrun asdetailed in thetext.
with another class of DNA tumor
viruses,
theadenoviruses (15).
Concerningthe virusgrowth cycle,wepropose
thefollowinginterpretationofourdata.(i) Upon
HSV-1 infection of HEp-2 cells in culture, a
DNAreplicationcomplexisformedfirst,similar
tothatwehave
prepared
with thesamemethodsfrom HSV-1-infected HEp-2 cells (20) which
contains viral DNApolymerase and other
fac-torssufficient for in vitro continuation of DNA
synthesis. (ii) Structural components are then
synthesized, and the DNA ispacked intoNPC
similar in appearance andprotein composition
to those described here. As the DNA "tails"
emerging from the nucleoprotein heads were
usually shorter than expected full length (24),
whereas afterdeproteinization45-,um-long
mol-eculeswere
observed,
wesuggestthatsomeviral DNAmightbecompactedinthe heads. Nucleo-capsid proteins VP5, VP15.2, VP19, and VP24could begood candidates for the role of starting
viral DNA encapsidation in vivo. As an
alter-nativeinterpretation,ifcapsidswerepreformed,
there mightbe smallamountsofother proteins
which "compact" the DNA into the capsid. We
would not detect such proteins if they were
present in only a few copies per capsid. The
NPC described here, which aredevoid of DNA
replicatingandtranscribingactivity, might
rep-resentbotha naturalintermediate in virion
as-sembly and a somehow more stable form in
particle disassembly, obtained by a controlled
on November 10, 2019 by guest
http://jvi.asm.org/
[image:9.496.118.403.68.449.2]HSV DNA-PROTEIN COMPLEXES 825
C .
t;eRl w'-4q.qsl
tt,sw >: :
st> .
St
.3 9..
I.
4 S
*0. .
''F
4t.
I
t
A
'.1 .
S
VOL. 36,1980
L.
Cob
Co
Co
*0
S.
C)
Co
S. .
o C.
S. C.e
on November 10, 2019 by guest
http://jvi.asm.org/
826 PIGNATTI AND CASSAI
rRACTION NUMBER
FIG. 10. Sedimentationanalysisoftheeffectsofphosphonoaceticacid additiontoHSV-infectedcells.Cells weregrown in the presence (A and B) and in the absence (C and D) of phosphonoacetic acid.
[3H]-thymidinewas added 6 hafter infection. Cellswerecollected24hafter infection.NPC(A andC) and virion (B and D)preparationprocedures were then applied. The samples were layered onto 20 to 50%sucrose gradientson77% cushions andcentrifugedat30,000 rpm and4°Cfor25min.Otherproceduresaredescribed
inthetext.Directionof sedimentation isfrom righttoleft.
disruption process. It does not seemlikely that
theNPC are artifacts of thepreparation
proce-dures,astheyareabsentfrom uninfected cells,
theirquantityincreases withtime after infection,
they are present also in isolated nuclei, they
appear inreduced amounts when virion
biosyn-thesis is reducedbyphosphonoacetate,andthey
can beobserved afterappropriate treatment of
isolated cytoplasmic virions. Moreover, as can
be inferred from other examples, very similar preparation procedures have yielded
informa-tiononpolyomavirus andsimian virus40NPC
inside the cell which have been confirmedbya
variety of different methods.(iii)TheNPC could
finallyassemble intonucleocapsids,astepwhich
canbe inhibitedbyphosphonoacetic acid,which
on November 10, 2019 by guest
http://jvi.asm.org/
[image:11.496.116.413.67.511.2]HSV DNA-PROTEIN COMPLEXES 827
University for theuseof their electronmicroscope,andCesare
Mussi fortechnical assistance.
[image:12.496.34.225.67.286.2]f
FIG. 11. Electron micrographs of
paredfromcellsinfected withHSVwc
presence ofphosphonoacetate.
Samp
from fraction19of thesucrosegradiei
JOB and prepared by the techniqueof
(5).Bar,50nm.
allowsthebiosynthetic processto
alittlebeyondstepii while variant
areformed, probably for the lack
essarystructural polypeptides. TI tionconstants of the complexespr
1,ii,andiiishowaprogressivemcr
value of naked viral DNA to th nucleocapsids. This could be ex
greatercompactnessobtained by t addition of the appropriate vira DNA. We should finally mention lication complex (step i) activit3 already detected incellscollected
tion, reaching itsmaximumvalue.
decliningthereafter to lowerlevel tionofNPC, onthe contrary
(ste]
a different time course: they firsicells collected 9 h postinfection creased in quantity upto 18h. Th of HSV NPC appearance in the
thus closely parallels that of viric
(23).
ACKNOWLEDGMENTS This work was supported by Consiglio
Ricerche (CNR) contribution CT 78.01424.' gettoFinalizzato Virusgrant79.00379.84,an
1225.
We thank Bernard Roizman for helpful constructive criticism, the CNR Centro l'Istochimicaand theIstitutodi AnatomiaCc
LITERATURE CITED
1. Birnie, G. D., D.Rickwood, and A. Hell. 1973. Buoyant densities and hydration of nucleic acids, proteins and nucleoproteincomplexes inmetrizamide. Biochimn. Bio-phys. Acta 331:283-294.
2 Bollum, F. J.1966. Filterpaperdisktechniques for
as-sayingradioactive macromolecules,p.296-300. In G. L.
Cantoni and D. R. Davies (ed.), Procedures in nucleic acidresearch.Harper and Row Publishers,New York. 3. Burgoyne,L.A., M. A. Waqar, and M. R. Atkinson.
1970.Calcium-dependent priming of DNA synthesis in isolatedratliver nuclei.Biochem.Biophys. Res.
Com-mun.39:254-259.
4. Cremisi, C., P. F. Pignatti, 0. Croissant, and M.
Yaniv. 1976.Chromatin-likestructuresinpolyoma
vi-rusandsimian virus 40lyticcycle.J. Virol.17:204-211. 5.Dubochet, J., M. Ducommun, M. Zollinger, and E. Kellenberger. 1971. A new preparation method for
dark-field electronmicroscopy ofbiomacromolecules.J.
Ultrastruct. Res.35:147-167.
6. Ejercito, P. M., E. D. Kieff, and B. Roizman. 1968.
Characterizationofherpes simplex virus strains differ-.---'-.t ing in their effectsonsocialbehaviour of infectedceUs.
J. Gen. Virol.2:357-364.
particles pre- 7. Favre, M.,F.Breitburd,0.Croissant, andG. Orth.
ad growninthe 1977. Chromatin-like structures obtainedafter alkaline ales were taken disruption of bovine and humanpapillomaviruses. J.
rtt
showere
taknFVirol.21:1205-1209.xtshown in Fig. 8. Furlong, D.,H.Swift, and B.Roizman.1972.
Arrange-Dubochetetal. mentofherpesvirusdeoxyribonucleic acid in thecore.
J.Virol. 10:1071-1074.
9. Gibson, W., and B. Roizman. 1972. Proteins specified by herpes simplex virus. VIII. Characterization and proceed only composition of multiplecapsid forms of subtypes1and
2.J.Virol. 10:1044-1052.
nucleocapsids 10. Griffith,J. D. 1975. Chromatinstructure: deduced from
ofsome nec- aminichromosome.Science187:1202-1203.
ie sedimenta- 11. Heine,J.W.,R. W.Honess, E.Cassai,and B.
Roiz-,esentinsteps man. 1974. Proteinsspecified byherpes simplexvirus.
reasefirom the virionpolypeptides type1 14:640-651.
iat of mature 12. Henry,C.J. 1974.The coreparticles ofherpessimplex
,plained by a virus.Can. J. Microbiol. 20:731-734.
the successive 13. Honess, R. W., and B. Roizman.1973. Proteins specified
proteins to by herpes simplex virus. XI.Identification and relative molarratesof synthesis of structural andnonstructural that the rep- herpesviruspolypeptidesin the infected cell. J. Virol.
was in fact 12:1347-1365.
3h postinfec- 14. Houssais,J. F. 1975. Separationdesparticules
ribonu-at6 to 9 h and cleoprot6iques(Hn-RNP) des noyaux decellulesL, en
deuxclassesparcentrifugationisopycniqueengradients
Ls.Theforma- demetrizamide.FEBSLett. 56:341-347.
p ii), followed 15. Kedinger,C., 0.Brison, F.Perrin, and J. Wilhelm.
t appeared in 1978. Structural analysis of viralreplicative
intermedi-and then in- atesisolated fromadenovirustype 2-infectedHeLa cell
timen
cou-e
nuclei. J. Virol. 26:364-379.e time course 16. Kieff, E. D., S. L. Bachenheimer, and B. Roizman.
infected cells 1971. Size,composition, and structure ofthe
deoxyri-mn production bonucleic acid of herpes simplexvirus subtypes1and 2.
J.Virol. 8:125-132.
17. Kleinschmidt, A. K. 1968. Monolayer techniquesin elec-tron microscopy of nucleic acid molecules. Methods Enzymol. 12B:361-377.
oNazionale delle 18. Meneguzzi,G., P. F. Pignatti,G.Barbanti-Brodano,
04, byCNR Pro- and G. Milanesi. 1978. Minichromosome from BK dby NATOgrant virusas atemplatefortranscriptionin vitro.Proc.Natl.
Acad. Sci. U.S.A. 75:1126-1130.
l suggestions and 19. Nii, S., and I. Yasuda. 1975. Detection of viral cores
di Studio per having toroidstructuresineight herpesviruses. Biken DmparataofPavia J. 18:41-46.
b
c-
As-I. .
.h
--.-VOL. 36, 1980
on November 10, 2019 by guest
http://jvi.asm.org/
PIGNATTI AND
20.Pignatti, P. F.,E.Cassai, and U.Bertazzoni. 1979.
Herpes simplex virus DNA synthesis in a partially purified solubleextractfrom infected cells. J. Virol.32: 1033-1036.
21. Pignatti,P.F.,E.Cassai,G.Meneguzzi, N.
Chenci-ner,and G. Milanesi. 1979.Herpessimplex virusDNA
isolation from infected cells with anovel procedure. Virology93:260-264.
22. Pignatti, P. F., C. Cremisi, 0. Croissant, and M.
Yaniv.1975.SV40nucleoprotein complexes:structural
modificationsafter isopycniccentrifugation in metriza-midegradients.FEBS Lett.60:369-373.
23. Roizman, B., and D. Furlong.1974.Thereplication of herpesviruses,p.229-403. In H.Fraenkel-Conrat and R. R. Wagner (ed.), Comprehensive virology, vol. 3.
Plenum Press, New York.
24. Wadsworth, S., R. J.Jacob,and B.Roizman. 1975. Anatomyof herpes simplex virus DNA. II. Size,
com-position,andarrangementof inverted terminal repeti-tions.J.Virol. 15:1487-1497.
25. Wilhelm, J.,0.Brison, C. Kedinger, and P.
Cham-bon.1976. Characterization of adenovirus type 2
tran-scriptional complexes isolatedfrominfected HeLacell
nuclei. J.Virol.19:61-81.
J. VIROL.